Introduction
Solid tumors are often malignant and are a serious
threat to human health. At present, lung carcinoma is the most
common type of malignancy and hepatocellular carcinoma (HCC) is the
second most common cause of cancer-associated mortality (1,2). Surgical
resection followed by radiotherapy and/or chemotherapy remains a
common treatment for malignant tumors (3,4). However,
these treatments are rarely curative. Therefore, the development of
specific and highly efficient therapies is important. Human
umbilical cord mesenchymal stem cells (hUCMSCs), derived from the
umbilical cord matrix, have been proposed as a promising tool for
the attenuation of tumor growth and metastasis (5).
hUCMSCs are a type of adult stem cell exhibiting
primitive stem cell characteristics, including self-renewal and
multi-potency. Following specific induction, hUCMSCs may
differentiate into cardiomyocytes, neuron-like cells, skeletal
muscle, endothelial cells and pancreatic islet-like cell clusters
(6). Therefore, the initial research
on hUCMSCs primarily focused on their multi-directional
differentiation ability, which is applicable for repairing damaged
tissues. At present, human MSCs used in experiments are primarily
acquired from adult bone marrow. Compared with bone marrow-derived
MSCs, hUCMSCs have several advantages for use in cell-based
therapy, including improved expansion capability, painless
collection procedures and lower risk of viral contamination. In
addition, these MSCs also share important characteristics,
including low immunogenicity, cytokine secretion and
transdifferentiation, which favor their potent application in the
development of novel tumor therapies (7–9).
Along with the frequent research into the use of
hUCMSCs as a tool for cancer treatment, the interactions between
hUCMSCs and malignant cells have been increasingly reported.
However, there are numerous inconsistent results regarding the
effects of hUCMSCs on tumors. In breast carcinoma models, the
interferon-β-transfected hUCMSCs were capable of migrating to tumor
sites and attenuating the proliferation of human triple-negative
breast carcinoma MDA-MB-231 and Hs578T cell lines (10). For glioma, hUCMSCs exerted antitumor
effects by inhibiting proliferation, modulating the cell cycle in
G0/G1 phase, and downregulating the expression of β-catenin and
c-Myc in C6 glioma cells (11);
hUCMSCs may also induce apoptosis in glioma U251 cells, resulting
in the significant upregulation of apoptotic genes, including
caspase-3 and caspase-9, and the significant downregulation of
anti-apoptotic genes, including survivin and X-linked inhibitor of
apoptosis (12). By contrast, other
studies (13,14) have indicated that hUCMSCs may promote
tumor proliferation in the tumor microenvironment. For example,
hUCMSCs activated by macrophages promote the proliferation and
migration of gastric epithelial cells and gastric cancer cells
(13); in addition, esophageal
carcinoma (EC) cells recruit hUCMSCs, and hUCMSCs promote EC cell
migration and invasion illustrated by the upregulation of
metastasis-related proteins, including matrix metalloproteinase
(MMP)-2 and MMP-9 in EC cells co-cultured with hUCMSCs (14).
In order to better understand the interactions
between hUCMSCs and specific tumor types, the present study further
examined this controversial issue by analyzing the underlying
mechanisms of the influence of hUCMSCs on the lung cancer A549 cell
line and the hepatocellular cancer BEL7402 cell line via indirect
and direct co-culture. The results indicated that hUCMSCs inhibit
lung cancer and HCC cell progression by inducing apoptosis and
targeting Wnt signaling, and possibly via direct cell-to-cell
contact.
Materials and methods
Cell culture
hUCMSCs were isolated from the umbilical cords of
healthy neonates delivered in local hospitals with the written
informed consent of their mothers. The research protocol was
approved by the Institutional Review Board of the School of Life
Science and Biopharmacology of Guangdong Pharmaceutical University
(Guangzhou, China). Separation, expansion and identification of
hUCMSCs were performed as previously described (15). Cells at passage 3 to 8, displaying a
homogeneous MSC immunophenotype and multipotent differentiation
potential into adipocytic, osteoblastic and chondrocytic lineages
(data not shown), were selected for experiments.
The human lung cancer A549 cell line and the human
HCC BEL7402 cell line were obtained from the American Type Culture
Collection (Manassas, VA, USA) and were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Zhejiang Tianhang
Biotechnology Co., Ltd., Hangzhou, China) at 37°C in a humidified
atmosphere containing 5% CO2, according to the American
Type Culture Collection protocols.
Treatment of tumor cells with
conditioned medium from hUCMSCs
hUCMSCs were cultured in Dulbecco's modified Eagle's
medium (DMEM)/F12 (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS to 100% confluence. The culture medium
was then harvested, filtered through 0.22-µm pore sterile filters
(Pall Life Sciences, Port Washington, NY, USA), and stored at −80°C
in aliquots until use. A mixture of the aforementioned supernatant
derived from hUCMSCs and RPMI-1640 medium containing 10% FBS at a
volume ratio of 3:2, designated as hUCMSC-CM, was subsequently used
to treat tumor cell lines at 37°C for 72 h. During the incubation
time, the hUCMSC-CM was replaced every 24 h (16) to prevent the rapid acidification of
culture medium due to the pre-mixing of hUCMSC supernatant with
fresh medium and to replenish the degradation of bioactive
substances in hUCMSC-CM. In the control group, the same tumor cells
were cultured with DMEM/F12 medium without FBS for 72 h.
Cell cycle analysis
A total of 1×106 cells were harvested,
washed twice with cold phosphate-buffered saline (PBS), fixed in
70% cold ethanol and stained with 5 µg/ml propidium iodide (PI;
Invitrogen; Thermo Fisher Scientific, Inc.) in the dark for 15 min
at 37°C. The DNA content of the stained cells was detected using a
flow cytometer (Gallios; Beckman Coulter, Inc., Brea, CA, USA) and
analyzed using ModFit software (Verity Software House, Inc.,
Topsham, ME, USA).
Transwell migration assay
After culturing at 37°C for 72 h with or without
hUCMSC-CM, the A549 and BEL7402 cells (1×105 cells/well)
were plated into the top chambers of Transwell plates (Corning
Incorporated, Corning, NY, USA). RPMI-1640 medium containing 10%
FBS was placed into the bottom chambers, followed by incubation at
37°C in an atmosphere containing 5% CO2 for 24 h. The
cells that did not penetrate the polycarbonate membrane were
removed with cotton stickers. The membrane was then fixed in 100%
methanol at room temperature for 15 min. Subsequently, the
morphological differences between the two tumor cell types became
less clear. To distinguish them properly, two different staining
agents, eosin and crystal violet, which are common in Transwell
assays of tumor cells (17,18), were used to stain A549 cells and
BEL7402 cells at room temperature for 5 min, respectively. The
migratory ability of tumor cells was determined by counting the
number of cells that had penetrated the membrane using a light
microscope (Olympus Corporation, Tokyo, Japan), in at least 5
fields for each assay (magnification, ×400).
Cell apoptosis assay
An Annexin V-fluorescein isothiocyanate (FITC)/PI
apoptosis detection kit (MBL International Co., Woburn, MA, USA)
was used to examine the percentage of apoptotic cells. In brief,
1×105 cells were washed with cold PBS and re-suspended
in binding buffer provided in the aforementioned kit. Cells were
incubated at room temperature with 10 µl Annexin V-FITC and 5 µl PI
for 15 min, and were then analyzed using a Gallios flow cytometer
(Beckman Coulter, Inc.) and FlowJo software 7.6.1 (FlowJo LLC,
Ashland, OR, USA).
Western blot analysis
Protein expression was measured via western blot
analysis. Total protein was extracted from A549 and BEL7402 cells
with ice-cold RIPA lysis buffer (Beyotime Institute of
Biotechonology, Shanghai, China), and the protein concentration was
determined using Bradford reagent (Beyotime Institute of
Biotechnology). A quantity of 30 µg total protein per lane was
separated by 10% SDS-PAGE and transferred onto nitrocellulose
membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membranes were then blocked by 5% non-fat milk for 1 h at 4°C under
agitation, followed by incubation with primary antibodies at 4°C
overnight against EphA5 (cat. no. sc-1014; 1:1,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), Bcl-2 (cat. no. 32124;
1:1,000; Abcam, Cambridge, MA, USA), pro caspase-7 (cat. no. 32067;
1:1,000; Abcam), β-catenin (cat. no. 8480; 1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) and c-Myc (cat. no. 5605;
1:1,000; Cell Signaling Technology, Inc.). An antibody against
GAPDH (cat. no. AB-P-R001; 1:1,000; Hangzhou Goodhere Biotechnology
Co., Ltd., Hangzhou, China; http://www.goodhere.com/) was used as the loading
control. Membranes were incubated with a horseradish
peroxidase-conjugated goat anti-rabbit IgG secondary antibody (cat.
no. ZB-2301; OriGene Technologies, Inc., Beijing, China) at a
dilution of 1:5,000 for 1 h at room temperature. Protein bands were
visualized by incubating the membranes with enhanced
chemiluminescence reagent (Bio-Rad Laboratories, Inc.) and recorded
on X-film (Kodak, Rochester, NY, USA). Protein expression was
quantified by densitometry using ImageJ software 1.51 (National
Institutes of Health, Bethesda, MD, USA).
Cell co-culture and detection
Cell fusion experiments based on direct contact
between tumor cell lines and hUCMSCs were performed as previously
described (19,20) with slight modification. Tumor cells
(A549 and BEL7402) and hUCMSCs were labeled with DIO and DID
fluorescent dyes (Invitrogen; Thermo Fisher Scientific, Inc.),
respectively, according to the manufacturer's protocols. Next, a
total of 1×104 DIO-labeled tumor cells were mixed with
the DID-labeled hUCMSCs, at a ratio of 1:1, in 100-mm glass bottom
dishes or 6-well cell culture plates containing DMEM/F12
supplemented with 10% FBS and 1% penicillin/streptomycin; these
were incubated at 37°C in a humidified atmosphere containing 5%
CO2. Monochrome fluorescence-labeled hUCMSCs or tumor
cells cultured alone were set as controls. The co-cultured cells
were observed under a confocal laser scanning microscope (FluoView
FV1000, Olympus Corporation) at a magnification of ×400 every 2–3
days and the images obtained were analyzed using FV10-ASW 1.6
software (Olympus Corporation) according to the manufacturer's
protocol. On day 8 of co-culture, the hybrids (double-stained
cells) were further confirmed using a Gallios flow cytometer
(Beckman Coulter, Inc.) and the fusion efficiency was calculated
using FlowJo software 7.6.1 (FlowJo LLC).
Statistical analysis
Statistical analysis was performed using SPSS
version 20.0 (IBM Corp., Armonk, NY, USA) software and GraphPad
Prism 6 software (GraphPad Software, Inc., La Jolla, CA, USA).
Quantitative data are expressed as the mean ± standard deviation.
The Student's t-test was used to test the probability of
significant differences between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
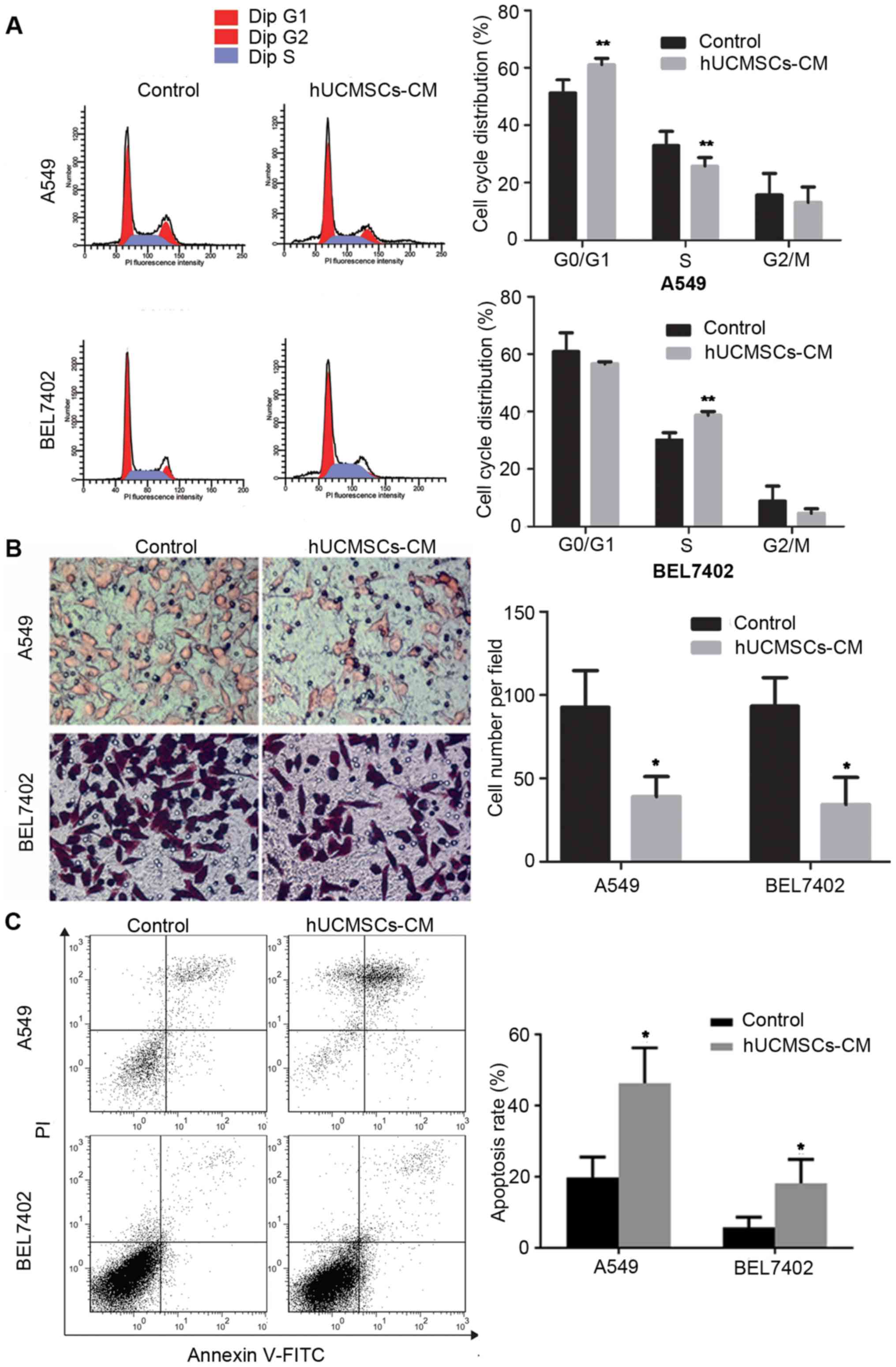
hUCMSC-CM influences the cell cycle of
A549 and BEL7402 cells
We hypothesized that the influence of the
supernatant from hUCMSCs on the growth of A549 and BEL7402 tumor
cells may be associated with altered cell cycle progression. As
expected, the analysis of cell cycle distribution indicated that
A549 tumor cells were arrested in the G1 phase, as illustrated by
an increased number of cells in the G1 phase (Fig. 1A). This was accompanied by a
corresponding reduction in the number of cells in the S phase
following incubation with hUCMSC-CM. The results for BEL7402 cells
treated with hUCMSC-CM were different, with an increased cell
number in the S phase and a corresponding reduction in the G1 phase
population (Fig. 1A). Overall,
analysis of the cell cycle distribution revealed that hUCMSC-CM
inhibited the proliferation of tumor cells by arresting the cell
cycle in specific phases.
hUCMSC-CM inhibits the migration of
A549 and BEL7402 cells
The effect of hUCMSC-CM on the migration of A549 and
BEL7402 tumor cells was investigated using 24-well polycarbonate
Transwell inserts. The results revealed that fewer A549 and BEL7402
cells were able to cross the membrane following treatment with
hUCMSC-CM, compared with the control group (P<0.05; Fig. 1B). Therefore, hUCMSC-CM markedly
reduced the migratory abilities of A549 and BEL7402 tumor
cells.
hUCMSC-CM induces the apoptosis of A549 and BEL7402
cells. In order to investigate whether apoptosis was involved in
the hUCMSC-mediated inhibition of tumor cell growth, flow
cytometric analysis was performed using Annexin V/PI staining. As
demonstrated in Fig. 1C, incubation
with hUCMSC-CM induced a significant increase in the percentage of
apoptotic cells, with 46.27±9.96% of A549 tumor cells and
18.23±6.66% of BEL7402 tumor cells in late-stage apoptosis
(P<0.05; Fig. 1C). Taken together,
these results demonstrated that hUCMSC-CM induced the apoptosis of
cancer cells.
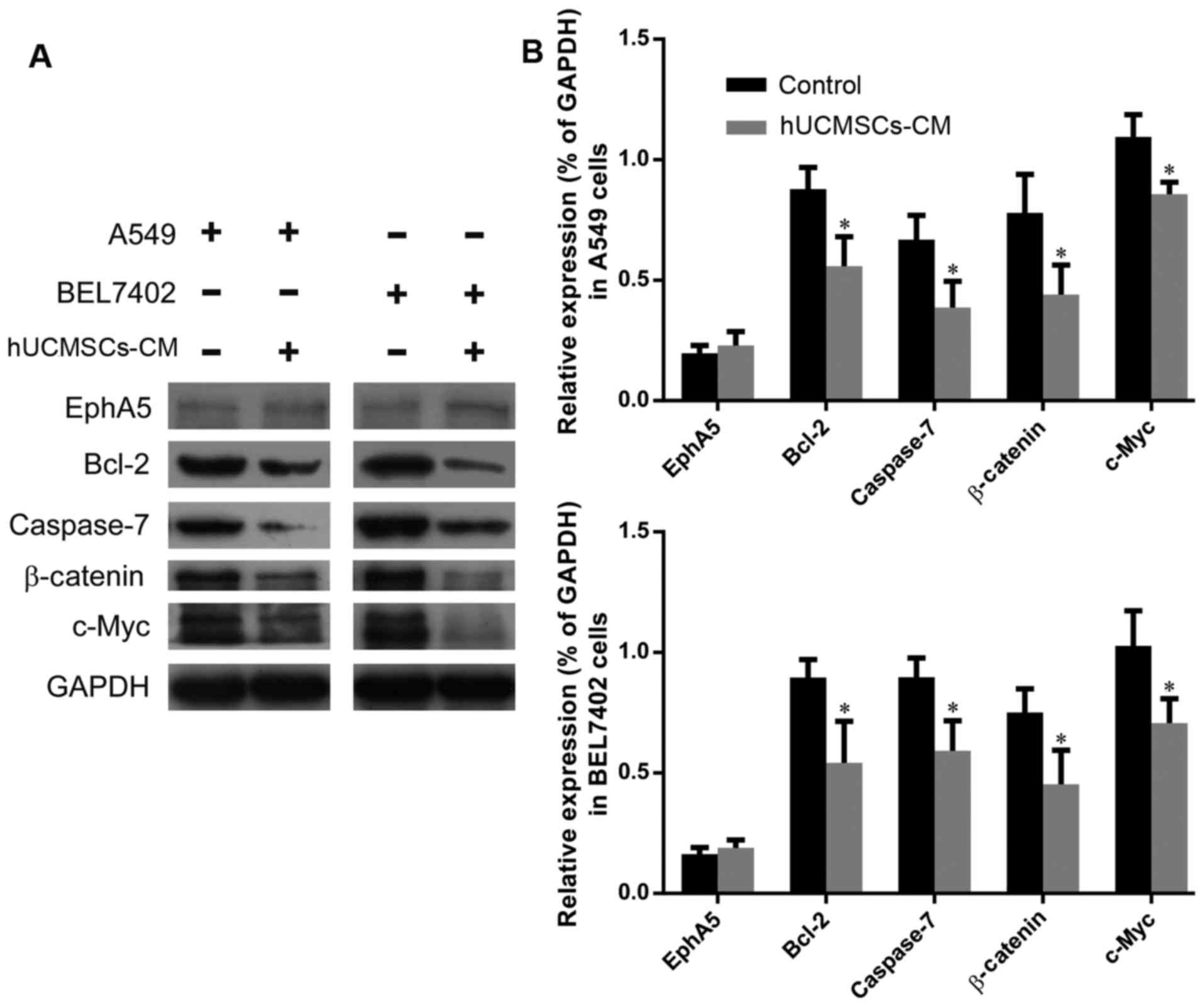
hUCMSC-CM modulates molecular
expression changes in A549 and BEL7402 cells
The results of western blot analysis revealed that
hUCMSC-CM slightly increased the expression of EphA5, which has
been identified as a biomarker of cancer cell dormancy. hUCMSC-CM
also resulted in the downregulation of Bcl-2 and caspase-7 in these
two tumor cell types (Fig. 2A and B).
The primary antibody against caspase-7 (cat. no. 32067; Abcam) that
was used in the present study only recognizes the pro-form but does
not react with cleaved forms of caspase-7. Accordingly, the
caspase-7 bands in Fig. 2A presented
the pro-form, but not the cleaved form of caspase-7. Wnt signaling
is known to serve critical roles in regulating the proliferation
and progression of tumor cells. Therefore, the involvement of Wnt
signaling was investigated in the present study. The expression
levels of β-catenin and c-Myc, two key signaling molecules in the
Wnt pathway, were downregulated in A549 and BEL7402 tumor cells
treated with hUCMSC-CM. Taken together, these results indicated the
involvement of cell apoptosis molecules, as well as that of the Wnt
signaling pathway, in the inhibitory effects of hUCMSCs on A549 and
BEL7402 tumor cells.
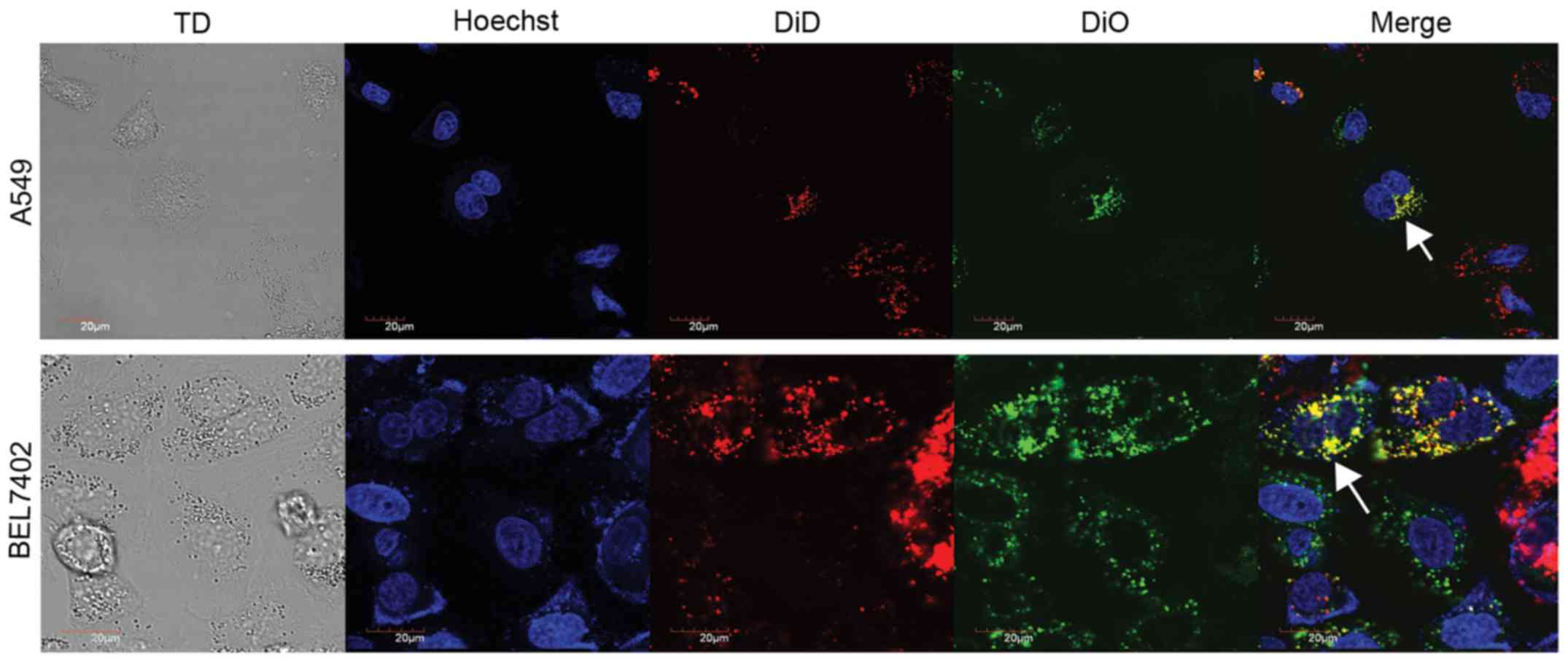
Co-culture of hUCMSCs with A549 and
BEL7402 cells induces cell fusion
hUCMSCs were mixed with the two solid tumor cell
types by direct cell-to-cell co-culture. The cell fusion event and
cell fusion efficiency were demonstrated using confocal
laser-scanning microscopy and flow cytometry, respectively. After
co-culture for 72 h in vitro, the hUCMSCs were revealed to
have merged into A549 or BEL7402 tumor cells, as illustrated by the
existence of bi-nucleated hybrid cells of dual color (DiO and DiD
double-stained cells), which were detected by confocal
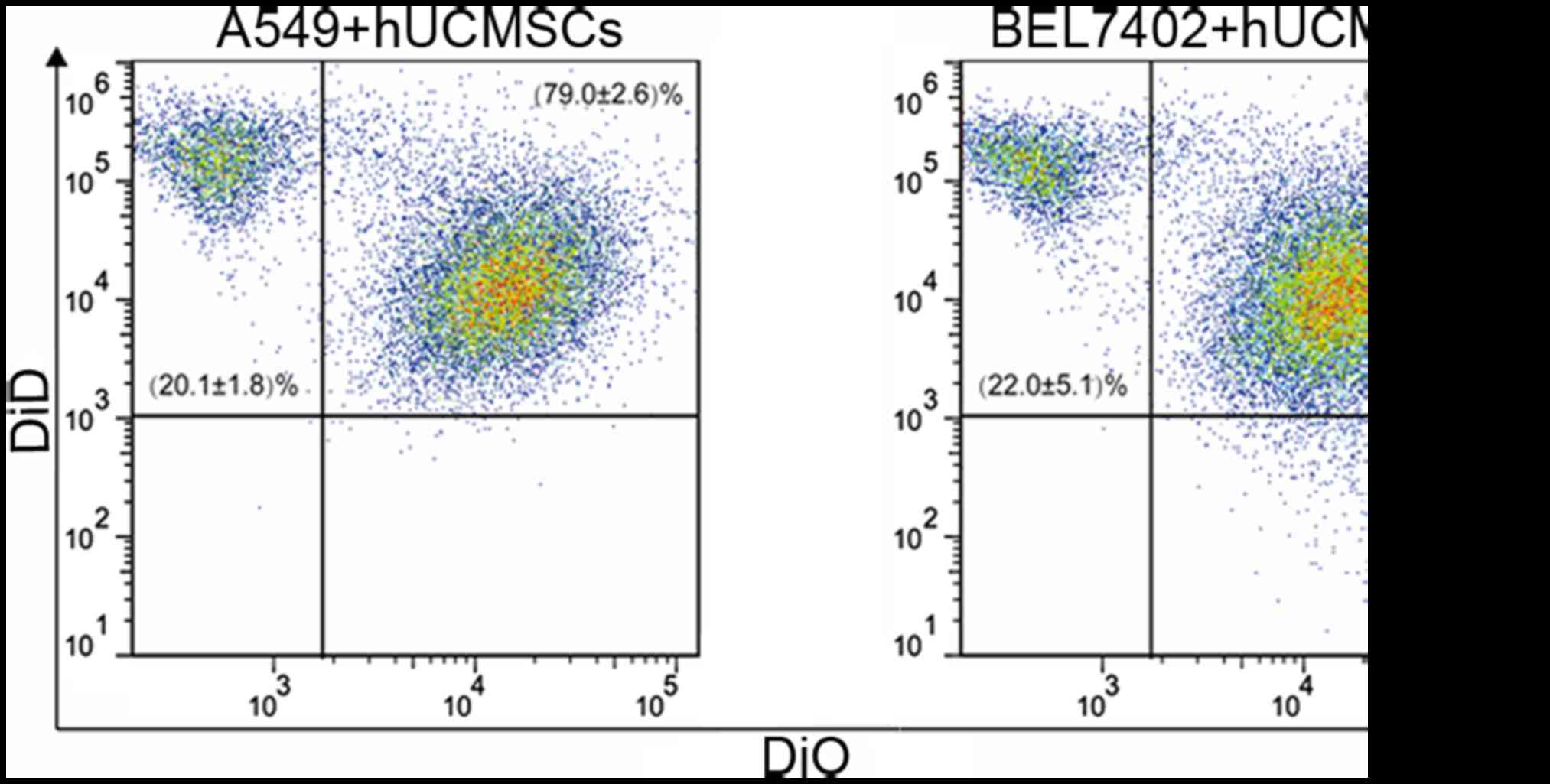
laser-scanning microscopy (Fig. 3).
Following co-culture for 8 days, flow cytometry revealed that the
fusion efficiencies were approximately 79.0±2.6% in A549 cells and
76.4±4.3% in BEL7402 cells (Fig.
4).
Discussion
Increasing evidence has indicated that adult stem
cells may be effective therapeutic tools for cancer therapy
(21,22). hUCMSCs are a group of adult stem cells
that are easy to collect, may be expanded extensively in
vitro and have a low immunogenicity. It is important to study
the effects of hUCMSCs on tumor growth in order to develop novel
therapies for the treatment of cancer. Recently, certain studies
have reported that hUCMSCs have an intrinsic ability to attenuate
the growth of several types of cancer cells. Ma et al
(23) directly injected hUCMSCs into
an immunodeficient xenograft mouse model transplanted with
MDA-MB-231 breast cancer stem cells. The hUCMSCs reduced tumor
volume and tumor weight in these mice. Zhang et al (24) reported that modified hUCMSCs that were
transfected with the inerleukin-21 gene inhibited the proliferation
of ovarian cancer cells in vitro and in vivo.
Subramanian et al (25)
confirmed that hUCMSCs did not transform into tumor-associated
fibroblasts, making them safer than bone marrow MSCs.
In our previous study, hUCMSCs were successfully
separated from the umbilical cords of healthy donors (15). hUCMSCs have the general
characteristics of MSCs. The aim of the present study was to
investigate the effects of hUCMSCs on the malignant behaviors,
including proliferation, migration and survival capabilities, of
the two types of solid tumor cells in vitro. Cell
proliferation is specifically controlled in the G1 phase and the
G1/S phase transition in the cell cycle. When cell cycle analysis
was conducted, significantly more tumor cells were observed in the
G1/S phase following treatment with hUCMSC-CM in vitro.
However, the cell cycle arrest in these two types of tumor cells
was different. Lung cancer A549 cells were arrested in the G0/G1
phase, while HCC BEL7402 cells were arrested in the S phase of the
cell cycle. Therefore, we hypothesized that hUCMSCs may interfere
with the growth of tumor cells through regulating cell cycle arrest
in different phases. These outcomes also suggest that the
anticancer effect of hUCMSCs may depend on the specific types of
tumor cells. Metastasis is one of the most important biological
behaviors of malignant tumors. The present study demonstrated that
hUCMSCs also significantly inhibited the migratory behavior of the
two types of tumor cells in Transwell chambers. Furthermore, it was
revealed that hUCMSC-CM significantly increased the proportion of
Annexin V/PI-positive cells. Consistently, the western blot
analysis revealed that hUCMSC-CM inhibited the expression of
apoptosis-related proteins, including Bcl-2 and pro caspase-7, in
A549 and BEL7402 cells. Caspase-7 has been identified as a major
contributor to the execution of apoptosis. During apoptosis,
caspase-7 is activated through proteolytic processing to produce
the mature and active subunits, resulting in the decrease of
pro-caspase-7 and emergence of cleaved caspase-7 (26). Therefore, the downregulation of
pro-caspase-7, as well as that of Bcl-2, indicated the activation
of intrinsic and extrinsic apoptosis pathways, and constituted the
molecular mechanisms underlying the hUCMSC-induced apoptosis of
tumor cells.
To further investigate other molecular mechanisms by
which hUCMSCs inhibited tumor cells, the present study focused on
the influence of hUCMSC-CM on Wnt signaling and tumor dormancy
biomarkers in A549 and BEL7402 tumor cells.
Wnt/β-catenin signaling has been demonstrated to
serve an important role in regulating tumor initiation and
progression in various malignant tumor types, including lung cancer
and HCC (27,28). Furthermore, Wnt signaling may regulate
genes that are involved in cell-cycle regulation and cell
apoptosis, including Cyclin and Bcl-2 (29). Therefore, we hypothesized that this
signaling pathway may be involved in governing the inhibitory
effects of hUCMSCs on lung cancer A549 cells and HCC BEL7402 cells.
β-catenin is a key mediator in Wnt signaling, regulating multiple
cellular functions. Stabilized β-catenin translocates into the
nucleus, binds with T-cell factor/lymphoid-enhancing factor
transcriptional factors and regulates the expression of downstream
β-catenin-dependent genes, including cyclin D1 and c-Myc (30). The results of western blot analysis in
the present study demonstrated that the treatment of A549 and
BEL7402 cells with hUCMSC-CM resulted in the downregulation of
β-catenin. To further assess whether known β-catenin targets were
also reduced, the expression of c-Myc was analyzed. Consistent with
the observed reduction in β-catenin, the expression levels of c-Myc
in these two tumor cell types receiving hUCMSC-CM were also
downregulated. These molecular data revealed that the
hUCMSC-mediated tumor inhibition was associated with the depression
of Wnt signaling in the two tumor cell types. It has been
demonstrated that MSCs may secret bioactive molecules, including
signaling proteins, cytokines and chemokines, into culture medium
(31). Some of these soluble factors
may have helped the triggering of the downregulation of Wnt
signaling in tumor cells.
Tumor dormancy describes a process through which
malignant cells exit the cell cycle and survive in a quiescent
state (32). Namely, the dormant
cancer cells usually experience the static stage at which they do
not proliferate or no longer enter the cell cycle. The maintenance
or reversal of tumor dormancy involves the changes of a series of
malignant biological behaviors, including the metastasis,
proliferation and apoptosis of tumor cells in a specific
environment. Accordingly, one possible explanation for tumor
dormancy is proliferative arrest, and the other is the equilibrium
between proliferation and apoptosis (32). At the beginning of the present study,
it was observed that hUCMSCs induced cell cycle arrest, an increase
in apoptosis and a decrease in the migration of A549 and BEL7402
tumor cells, which indicated the possible involvement of dormancy
mechanisms. However, it has been demonstrated that the interactions
between cancer cells and their normal neighbors act as an important
microenvironmental control of the onset and maintenance of dormancy
during cancer development (33). MSCs
are one of the critical components in the tumor microenvironment.
We hypothesized that MSCs may also serve a role in tumor dormancy.
On the basis of the aforementioned speculations, the present study
aimed to investigate the expression of dormancy-specific biomarkers
(34). EphA5, one of the tumor
dormancy markers that was detected, was upregulated at the mRNA and
protein levels in the two tumor cell types following incubation
with hUCMSC-CM. This may be a possible explanation for how hUCMSCs
support tumor dormancy. Similarly, Glick and Yuspa (35) observed that the tumor microenvironment
may inhibit or reverse the malignant characteristics of tumor cells
that are in a dormant state, and tumors are able to develop only
when the stability of the microenvironment is disrupted.
Alt-Holland et al (36) also
proposed that the signaling network interaction between tumor cells
and adjacent normal cells may control tumor growth and maintain the
dormancy of tumor cells.
The majority of solid tumor cells and MSCs are
adherent cells. Therefore, in order to avoid the interference of
MSCs with the detection of tumor cells, the majority of experiments
prefer to culture tumor cells with conditioned medium from MSCs.
However, MSCs will inevitably come into contact with tumor cells
after entering the body when they are used for tumor therapy. To
better reflect this situation, in the present study, hUCMSCs were
co-cultured with the two solid tumor cell types by direct
cell-to-cell contact. With confocal scanning, bi-nucleated hybrid
cells were observed due to the fusion of hUCMSCs with the
co-cultured tumor cells, and it was re-affirmed by flow cytometry.
Specifically, hybrid cells with two clear nuclei were observed
until the end of 6 days of confocal tracking in the present study
(data not shown), which may aid in distinguishing cell fusion from
other mechanisms, including phagocytosis among cells as well as
endocytosis of MSC-secreted exosomes to a certain extent.
Phagocytosis refers to the process of specifically engulfing and
destroying particulate targets via diverse mechanisms (37). Targets of phagocytosis include
microorganisms, dead or dying cells, and environmental debris. By
contrast, cell fusion is a nuclear reprogramming process that
involves fusing two or more cell types to form a single identity
and generally does not cause deadly damage to the two sides of the
fusion (19). However, membranous
vesicle transport, particularly the exosome-mediated endocytosis,
is one of the important mechanisms by which mesenchymal stem cells
exert their biological functions, possibly including the
communication between MSCs and tumor cells (38). Exosomes and other extracellular
vesicles belong to subcellular components without nuclear
structures, although they usually contain cell-specific proteins,
lipids and nucleic acids. However, in the present study,
bi-nucleated cells were observed under confocal microscope, which
indicated the direct fusion of hUCMSCs into tumor cells.
Considering the limitations of the present study, including the
absence of electron microscopy data, the aforementioned observation
does not exclude the involvement of exosomes or other mechanisms,
but emphasized the potential roles of cell fusion in the crosstalk
between MSCs and tumor cells.
It has been widely demonstrated that numerous cell
types in the tumor microenvironment are able to merge with
malignant cells by cell fusion (39,40). As
one of the critical components in the tumor microenvironment, MSCs
are also a putative fusogenic candidate. Similarly, the study of
Wei et al (19) co-cultured
RFP-expressing MSCs with eGFP-expressing lung cancer H441 cells
without any fusogenic agent and demonstrated that MSCs fuse
spontaneously with lung cancer cells. Transcriptome profiles
revealed that the lung cancer cells are reprogrammed to slow growth
and a stem-like state upon MSC fusion, accomplished by the
restoration of p21 function and the upregulation of forkhead box
F1, a putative tumor suppressor (19). Wang et al (20) also generated fusion progeny by fusing
DiD-labeled MSCs and DiO-labeled esophageal carcinoma cells with
PEG1500, and confirmed that the fusion aids in controlling the
malignant phenotype of esophageal cancer cells.
In summary, the results of the present study
suggested that hUCMSCs may inhibit the malignant biological
behaviors of human lung cancer and hepatocellular cancer cells
in vitro by activating cell apoptosis and inhibiting Wnt
signaling. hUCMSCs also have the potential ability to induce tumor
dormancy, at least through the mechanism of cell cycle arrest. In
addition, the present study provided evidence to support
spontaneous cell fusion between hUCMSCs and tumor cells, which may
contribute to the antitumor effects of hUCMSCs. Unlike individual
molecules, including protein and DNA/RNA, each complete cell is
functionally independent and the cell-cell interaction involves
complex mechanisms. As a preliminary attempt to investigate such a
complicated issue, the present study may only provide limited
conclusions for the time being. Notably, the results of the present
study provide valuable clues for in-depth research on the
interactions between MSCs and tumors, including the identification
of bioactive molecules in hUCMSC-CM as upstream modulators of Wnt
signaling. Therefore, the mechanisms emphasized in the present
study warrant further efforts by using additional approaches and
more suitable models in the future.
Acknowledgements
The authors wish to thank Dr Danliang Chen of the
First Affiliated Hospital of Jinan University (Guangdong, China)
and Dr Liqin Zhao in the Third Affiliated Hospital of Southern
Medical University (Guangdong, China) for their help in umbilical
cord collection.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81502973), the
Guangdong Natural Science Foundation of China (grant nos.
2014A030310259 and 2015A030310310) and the Innovative School
Project of Guangdong Pharmaceutical University (grant no.
2014KQNCX142).
Availability of data and materials
The datasets analyzed during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
XL and YY designed the experiments. YY performed the
hUCMSC isolation. CZ performed tumor cell culture. YY, CZ, and XC
performed Transwell assay and western blot analysis. YY and XC
performed flow cytometric analyses. CT and XL performed confocal
laser scanning and image analysis. HC performed cell co-culture.
YY, CZ, and XL wrote the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Human umbilical cord collection for research was
approved by the institutional review board of the School of Life
Science and Biopharmacology of Guangdong Pharmaceutical University
(Guangzhou, China). MSCs were isolated from umbilical cords with
donor's informed consent.
Consent for publication
The patients consented to the publication of this
data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang W, Zhang L, Liu L, Zheng Y, Zhang Y,
Yang S, Shi R and Wang S: Chemosensitizing effect of shRNA-mediated
ERCC1 silencing on a Xuanwei lung adenocarcinoma cell line and its
clinical significance. Oncol Rep. 37:1989–1997. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rojas A, Zhang P, Wang Y, Foo WC, Muñoz
NM, Xiao L, Wang J, Gores GJ, Hung MC and Blechacz B: A positive
TGF-β/c-KIT feedback loop drives tumor progression in advanced
primary liver cancer. Neoplasia. 18:371–386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baskar R, Yap SP, Chua KL and Itahana K:
The diverse and complex roles of radiation on cancer treatment:
Therapeutic target and genome maintenance. Am J Cancer Res.
2:372–382. 2012.PubMed/NCBI
|
|
4
|
Lee DH, Ahn Y, Kim SU, Wang KC, Cho BK,
Phi JH, Park IH, Black PM, Carroll RS, Lee J and Kim SK: Targeting
rat brainstem glioma using human neural stem cells and human
mesenchymal stem cells. Clin Cancer Res. 15:4925–4934. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ciavarella S, Dominici M, Dammacco F and
Silvestris F: Mesenchymal stem cells: A new promise in anticancer
therapy. Stem Cells Dev. 20:1–10. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang J, Miao Y, Chang Y, Zhang F, Wang Y
and Zheng S: Condition medium of HepG-2 cells induces the
transdifferentiation of human umbilical cord mesenchymal stem cells
into cancerous mesenchymal stem cells. Am J Transl Res.
8:3429–3438. 2016.PubMed/NCBI
|
|
7
|
Liao W, Zhong J, Yu J, Xie J, Liu Y, Du L,
Yang S, Liu P, Xu J, Wang J, et al: Therapeutic benefit of human
umbilical cord derived mesenchymal stromal cells in intracerebral
hemorrhage rat: Implications of anti-inflammation and angiogenesis.
Cell Physiol Biochem. 24:307–316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong YS, Lin N, Deng MH, Zhang FC, Tang
ZF and Xu RY: Deficient proliferation of bone marrow-derived
mesenchymal stem cells in patients with chronic hepatitis B viral
infections and cirrhosis of the liver. Dig Dis Sci. 55:438–445.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bieback K, Kem S, Kluter H and Eichler H:
Critical parameters for the isolation of mesenchymal stem cells
from umbilical cord blood. Stem Cells. 22:625–634. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen CJ, Chan TF, Chen CC, Hsu YC, Long CY
and Lai CS: Human umbilical cord matrix-derived stem cells
expressing interferon-β gene inhibit breast cancer cells via
apoptosis. Oncotarget. 7:34172–34179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma S, Liang S, Jiao H, Chi L, Shi X, Tian
Y, Yang B and Guan F: Human umbilical cord mesenchymal stem cells
inhibit C6 glioma growth via secretion of dickkopf-1 (DKK1). Mol
Cell Biochem. 385:277–286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang C, Lei D, Ouyang W, Ren J, Li H, Hu J
and Huang S: Conditioned media from human adipose tissue-derived
mesenchymal stem cells and umbilical cord-derived mesenchymal stem
cells efficiently induced the apoptosis and differentiation in
human glioma cell lines in vitro. Biomed Res Int. 2014:1093892014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang M, Cai J, Huang F, Zhu M, Zhang Q,
Yang T, Zhang X, Qian H and Xu W: Pre-treatment of human umbilical
cord-derived mesenchymal stem cells with interleukin-6 abolishes
their growth-promoting effect on gastric cancer cells. Int J Mol
Med. 35:367–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang X, Li Z, Ma Y, Gao J, Liu S, Gao Y
and Wang G: Human umbilical cord mesenchymal stem cells promote
carcinoma growth and lymph node metastasis when co-injected with
esophageal carcinoma cells in nude mice. Cancer Cell Int.
14:932014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan Y, Lu X, Chen X, Shao H and Huang S:
Jagged1 contributes to the drug resistance of Jurkat cells in
contact with human umbilical cord-derived mesenchymal stem cells.
Oncol Lett. 6:1000–1006. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qiao L, Xu ZL, Zhao TJ, Ye LH and Zhang
XD: Dkk-1 secreted by mesenchymal stem cells inhibits growth of
breast cancer cells via depression of Wnt signalling. Cancer Lett.
269:67–77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ba Q, Li J, Huang C, Qiu H, Li J, Chu R,
Zhang W, Xie D, Wu Y and Wang H: Effects of benzo [a]pyrene
exposure on human hepatocellular carcinoma cell angiogenesis,
metastasis and NF-κB signaling. Environ Health Perspect.
123:246–254. 2015.PubMed/NCBI
|
|
18
|
Tian LL, Yue W, Zhu F, Li S and Li W:
Human mesenchymal stem cells play a dual role on tumor cell growth
in vitro and in vivo. J Cell Physiol. 226:1860–1867. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei HJ, Nickoloff JA, Chen WH, Liu HY, Lo
WC, Chang YT, Yang PC, Wu CW, Williams DF, Gelovani JG and Deng WP:
FOXF1 mediates mesenchymal stem cell fusion-induced reprogramming
of lung cancer cells. Oncotarget. 5:9514–9529. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Fan H, Zhou B, Ju Z, Yu L, Guo L,
Han J and Lu S: Fusion of human umbilical cord mesenchymal stem
cells with esophageal carcinoma cells inhibits the tumorigenicity
of esophageal carcinoma cells. Int J Oncol. 40:370–377.
2012.PubMed/NCBI
|
|
21
|
Di GH, Jiang S, Li FQ, Sun JZ, Wu CT, Hu X
and Duan HF: Human umbilical cord mesenchymal stromal cells
mitigate chemotherapy-associated tissue injury in a pre-clinical
mouse model. Cytotherapy. 14:412–422. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang C, Yang SJ, Wen Q, Zhong JF, Chen
XL, Stucky A, Press MF and Zhang X: Human-derived normal
mesenchymal stem/stromal cells in anticancer therapies. J Cancer.
8:85–96. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma Y, Hao X, Zhang S and Zhang J: The in
vitro and in vivo effects of human umbilical cord mesenchymal stem
cells on the growth of breast cancer cells. Breast Cancer Res
Treat. 133:473–485. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Wang J, Ren M, Li M, Chen D, Chen
J, Shi F, Wang X and Dou J: Gene therapy of ovarian cancer using
IL-21-secreting human umbilical cord mesenchymal stem cells in nude
mice. J Ovarian Res. 7:82014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Subramanian A, Shu-Uin G, Kae-Siang N,
Gauthaman K, Biswas A, Choolani M, Bongso A and Chui-Yee F: Human
umbilical cord wharton's jelly mesenchymal stem cells do not
transform to tumor-associated fibroblasts in the presence of breast
and ovarian cancer cells unlike bone marrow mesenchymal stem cells.
J Cell Biochem. 113:1886–1895. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ghoshal A and Ghosh SS: Antagonizing
canonical Wnt signaling pathway by recombinant human sFRP4 purified
from E. coli and its implications in cancer therapy. Mol Cell
Biochem. 418:119–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei Y, Shen N, Wang Z, Yang G, Yi B, Yang
N, Qiu Y and Lu J: Sorafenib sensitizes hepatocellular carcinoma
cell to cisplatin via suppression of Wnt/β-catenin signaling. Mol
Cell Biochem. 381:139–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Baek SH, Kioussi C, Briata P, Wang D,
Nguyen HD, Ohgi KA, Glass CK, Wynshaw-Boris A, Rose DW and
Rosenfeld MG: Regulated subset of G1 growth-control genes in
response to derepression by the Wnt pathway. Proc Natl Acad Sci
USA. 100:3245–3250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu J, Han G, Liu H and Qin C: Suppression
of cholangiocarcinoma cell growth by human umbilical cord
mesenchymal stem cells: A possible role of Wnt and Akt signaling.
PLoS One. 8:e628442013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Caplan AI and Correa D: The MSC: An injury
drugstore. Cell Stem Cell. 9:11–15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Aguirre-Ghiso JA: Models, mechanisms and
clinical evidence for cancer dormancy. Nat Rev Cancer. 7:834–846.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barkan D, El Touny LH, Michalowski AM,
Smith JA, Chu I, Davis AS, Webster JD, Hoover S, Simpson RM,
Gauldie J and Green JE: Metastatic growth from dormant cells
induced by a col-I-enriched fibrotic environment. Cancer Res.
70:5706–5716. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Almog N, Ma L, Raychowdhury R, Schwager C,
Erber R, Short S, Hlatky L, Vajkoczy P, Huber PE, Folkman J and
Abdollahi A: Transcriptional switch of dormant tumors to
fast-growing angiogenic phenotype. Cancer Res. 69:836–844. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Glick AB and Yuspa SH: Tissue homeostasis
and the control of the neoplastic phenotype in epithelial cancers.
Semin Cancer Biol. 15:75–83. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alt-Holland A, Zhang W, Margulis A and
Garlick JA: Microenvironmental control of premalignant disease: The
role of intercellular adhesion in the progression of squamous cell
carcinoma. Semin Cancer Biol. 15:84–96. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
David DM and Goodridge HS: Information
processing during phagocytosis. Nat Rev Immuno. 12:492–502. 2012.
View Article : Google Scholar
|
|
38
|
Lavoie JR and Rosu-Myles M: Uncovering the
secretes of mesenchymal stem cells. Biochimie. 95:2212–2221. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jacobsen BM, Harrell JC, Jedlicka P,
Borges VF, Varella-Garcia M and Horwitz KB: Spontaneous fusion with
and transformation of mouse stroma by, malignant human breast
cancer epithelium. Cancer Res. 66:8274–8279. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mortensen K, Lichtenberg J, Thomsen PD and
Larsson LI: Spontaneous fusion between cancer cells and endothelial
cells. Cell Mol Life Sci. 61:2125–2131. 2004. View Article : Google Scholar : PubMed/NCBI
|