Introduction
The growth rate of prostate cancer is slow and early
symptoms are often not obvious and are easily overlooked, which may
delay diagnosis and initiation of treatment (1,2). The
molecular mechanisms underlying prostate cancer metastasis remain
unclear, and there is a requirement for the identification of
molecular markers for metastasis to enable the treatment and to
improve the prognosis of patients with prostate cancer.
The membrane-associated Annexin family is a group of
highly evolutionarily conserved calcium-dependent
phospholipid-binding proteins (3).
The Annexin family has numerous critical functions in cell
signaling, inflammation, cell proliferation and differentiation,
and maintenance of the extracellular matrix integrity (4,5).
Downregulation or dysfunction of Annexins may serve important roles
in the development and progression of malignant tumors. Annexin A2
(ANXA2) is one of nearly 20 members of the Annexin superfamily and
contains a ~70 amino acid conserved repeat region (6). ANXA2 is involved in diverse cellular
functions, including cell apoptosis, membrane transport, signal
transduction and cell-cell interaction (7). ANXA2 is a multifunctional protein that
serves an important regulatory role in biofilm formation,
endocytosis and exocytosis, osteoblast formation, bone resorption,
DNA synthesis, and cell proliferation and differentiation (8). ANXA2 is also associated with a number of
cellular activities by connecting membrane protein complexes with
the actin cytoskeleton, and is involved in ion channel formation,
plasminogen activation and cellular matrix interactions (9,10).
Epithelial-mesenchymal transition (EMT) is a highly
regulated process by which epithelial cells transform into a
mesenchymal cell phenotype (11). EMT
is associated with primary tumor invasion, cell migration and
secondary metastasis formation in a variety of tumor types,
particularly in those of epithelial origin (12,13).
Bioinformatics analysis is a major tool for the
identification of putative microRNA (miRNA/miR)-targeted genes. The
present study used bioinformatics software analysis to identify
ANXA2 mRNA as a target of miR-206 and an aim of the present
study was to examine the molecular mechanism by which miR-206
targeting of ANXA2 regulates the EMT, and the invasive and
metastatic activity of prostate cancer cells.
Materials and methods
Patients and tissue specimens
A total of 110 male patients were enrolled in the
present study; 60 with prostate cancer (median age, 72.8 years; age
range, 56–85 years), 30 with metastatic prostate cancer (median
age, 73.5 years; age range, 57–85 years), and 20 with benign
prostatic hyperplasia (BPH; median age, 68.6 years; age range,
52–83 years) as control. Patients with prostate cancer included in
the present study received no preoperative medication and had no
history of surgical castration or radiotherapy. Patients with BPH
had no long-term medication history prior to surgery. Tissue
samples were obtained by surgical resection at the Department of
Urology at the Second Affiliated Hospital, University of South
China (Hengyang, China) and were stored at −80°C prior to use. All
specimens were reviewed independently by two senior pathologists
and the diagnoses were confirmed by histopathological examination.
The present study was certified by the Ethics Committee of the
Second Affiliated Hospital of University of South China, and all
participants provided written informed consent.
Cell lines
The prostate cancer PC-3 cell line was purchased
from The Cell Centre of Central South University (Changsha, China).
Cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), supplemented with 10% bovine
serum albumin, and were incubated at 37°C in a 5% CO2
atmosphere.
Reagents
The immunohistochemical streptavidin peroxidase
(S-P) kit and 3,3′-diaminobenzidine developer were obtained from
Fuzhou Maixin Biotech Co., Ltd. (Fuzhou, Fujian province, China).
Mouse anti-human monoclonal antibodies against ANXA2, GAPDH,
E-cadherin, N-cadherin and β-actin were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Lipofectamine 2000 was
purchased from Invitrogen; Thermo Fisher Scientific, Inc., the
Transwell assay kit was purchased from Corning Incorporated
(Corning, NY, USA), and Matrigel was obtained from BD Biosciences
(Franklin Lakes, NJ, USA).
Bioinformatics analysis
miRNAs predicted to bind to ANXA2 mRNA were
identified using the miRWalk online program, which contains 10
software programs (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk/predictedmirnagene.html).
The miRNAs with the highest predicted binding scores were
identified using miRanda software (version: August 2010 release;
http://www.microrna.org/microrna/home.do), which
computes thermodynamic stability scores and sequence conservation
scores.
Immunohistochemistry (IHC)
The prostate tissue specimens were fixed using by
10% formalin for 24–48 h at room temperature, and then embedded in
paraffin. The sample was sliced into sections 4 µm thick.
Immunohistochemical staining of prostate tissue specimens was
performed using the S-P immunohistochemical method (14). The cytoplasmic staining intensity was
scored by two pathologists as follows: No color, negative (−); pale
yellow, weakly positive (+); brown, positive (++); and tan,
strongly positive (+++). The percentage of tissue samples with
positive expression was calculated as [(total number of samples
with weakly positive + positive + strongly positive staining)/total
number of samples evaluated] ×100.
RNA extraction
Total RNA was extracted from fresh prostate cancer
and BPH tissues by homogenization using TRIzol reagent (Thermo
Fisher Scientific, Inc. Waltham, MA, USA). Following incubation for
5 min at room temperature, the samples were mixed with 200 ml of
chloroform, incubated for 5 min at room temperature, and then
centrifuged at 12,000 × g for 15 min at 4°C. The supernatant was
removed, combined with 200 ml isopropanol, mixed by inversion,
incubated for 10 min at room temperature, and centrifuged at 12,000
× g for 15 min at 4°C. The supernatant was removed and the pellet
was washed by addition of 1 ml ethanol followed by centrifugation
at 12,000 × g for 15 min at 4°C. The supernatant was removed and
the pellet was vacuum dried. Finally, the RNA was resuspended in
deionized water and subsequently used for reverse
transcription-quantitative polymerase chain reactions
(RT-qPCR).
RT-qPCR
PCR amplification of miR-206 was performed using a
two-step method using an RT-qPCR kit (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). RT were performed in a water bath at 50°C for
60 min and 15 min with inactivated reverse transcriptase
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 70°C in a water
bath. Primers were designed based on the precursor sequence of
miR-206 by Primer Premier 5.0 (Primer Biosoft, Paolo Alto, CA,
USA). Primer sequences were as follows: miR-206 forward,
5′-TGCTTCCCGAGGCCACATGC-3′ and reverse, 5′-GTGTGTGGTTTCGGCAAGTG-3′;
and U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. All primers were synthesized by
Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The reactions were
performed on a fluorescence RT-qPCR instrument, according to the
manufacturer's protocol. The PCR reaction system is as follows: 2.5
µl dNTP (2.5 mM each); 2.5 µl 10× PCR buffer; 1.5 µl
MgCl2 solution; 1 U Taq polymerase; 0.25× SYBRGreen I
(Sigma-Aldrich; Merck KGaA) (final concentration); 1 µl 10 µM PCR
forward and reverse primer; 1 µl cDNA; water (to a total volume of
25 µl). The U6 reaction was as follows: 95°C for 5 min and 35
cycles of 95°C for 10 sec, 59°C for 15 sec, 72°C for 20 sec and
82°C for 5 sec. The miRNA reaction was as follows: 95°C for 15 min
and 40 cycles of 94°C for 15 sec, 55°C for 30 sec and 70°C for 30
sec). The 2−ΔΔCq was used to quantify the results
(15).
Western blot analysis
To extract total cellular protein, tissues or cells
were mixed with precooled lysis buffer (Sigma-Aldrich; Merck KGaA),
centrifuged for 10 min at 4°C at 10,000 × g, and placed on ice for
30 min. Following centrifugation, the supernatants were removed and
protein concentrations were determined using a Bradford assay. A
total of 40 µg protein was separated using SDS-PAGE (10% gel). The
proteins were denatured by incubation for 5 min at 100°C, separated
by gel electrophoresis and transferred onto polyvinylidene
difluoride membranes. The membranes were blocked at room
temperature with in 5% skimmed milk powder for 1–2 h and were then
washed and incubated with the following primary antibodies:
Anti-ANXA2 (1:1,000; cat. no. 28385), anti-β-actin
(1:10,000; cat. no. 58673), anti-vimentin (1:1,500; cat. no. 6260),
anti N-cadherin (1:1,500; cat. no. 8424) and anti-E-cadherin
(1:1,500; cat. no. 8426), overnight at 4°C, and added the secondary
antibody (1:1,000; HRP-conjugated mouse anti-rabbit IgG; cat. no.
2357) for 2 h at temperature. All the primary antibodies and the
secondary antibody were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). The proteins were treated using enhanced
chemiluminescence chromogenic solution (Boster Biological
Engineering Co., Ltd. Wuhan, China). Visualization of the protein
on the membrane occurred upon exposure to X-ray film. The Gray
value was detected by Image J software (version K1.45; National
Institutes of Health, Bethesda, MD, USA).
In vitro cell invasion assay
All the cell lines were cultured in RPMI-1640 and
10% fetal bovine serum (Hangzhou Sijiqing Biological Engineering
Co., Ltd., Hangzhou, China). PC-3 cells were seeded onto 6-well
plates (~2×105 cells/plate) and cultured to 80%
confluence prior to transfection at 37°C. Cells were transfected
with miR-206 mimic or inhibitor sequences or a negative control
(NC) sequence using a Lipofectamine 2000 transfection kit (Santa
Cruz Biotechnology, Inc.). After 48 h of transfection, the cell
extractive was analyzed to detect the activity of the reported
gene. For the Transwell invasion assay, Matrigel was diluted at
1:100 in serum-free medium and 100 µl was added to coat the upper
chamber at room temperature for 24 h. To detect invasion, the upper
chamber was removed, and the cells in the membrane were fixed in
ethanol at room temperature for 30 min and stained with 0.5%
crystal violet at room temperature for 20 min. The number of
invaded tumor cells in the membrane was counted under an optical
microscope (Nikon Corporation, Tokyo, Japan) at ×400
magnification.
Statistical analysis
Data were analyzed using SPSS version 17 software
(SPSS, Inc., Chicago, IL, USA). Data are expressed as the mean ±
standard deviation. Comparisons were performed using an unpaired
Student's t-test or Mann-Whitney U test for two groups or with a
one-way analysis of variance, followed by the least significant
difference post hoc test, for three groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miRNA-ANXA2 interactions predicted by
miRWalk
Using miRWalk, which combines the power of 10
programs, the following 12 miRNAs predicted to interact with
ANXA2 mRNA were identified: hsa-miR-206, hsa-miR-1,
hsa-miR-9, hsa-miR-613, hsa-miR-185, hsa-miR-425, hsa-miR-890,
hsa-miR-155, hsa-miR-20b, hsa-miR-520h, hsa-miR-579 and
hsa-miR-767-5p (Table I).
 | Table I.Results of intersection analysis by
miRWalk. |
Table I.
Results of intersection analysis by
miRWalk.
| Gene name | miRNA | DIANAmT | miRanda | miRDB | miRWalk | RNAhybrid | PICTAR4 | PICTAR5 | PITA | RNA22 | Target scan | SUM |
|---|
| ANXA2 | hsa-miR-206 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 6 |
| ANXA2 | hsa-miR-1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 6 |
| ANXA2 | hsa-miR-890 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 5 |
| ANXA2 | hsa-miR-9 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 5 |
| ANXA2 | hsa-miR-613 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 5 |
| ANXA2 | hsa-miR-185 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 5 |
| ANXA2 | hsa-miR-425 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 5 |
| ANXA2 | hsa-miR-155 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 4 |
| ANXA2 | hsa-miR-20b | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 4 |
| ANXA2 | hsa-miR-520h | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 4 |
| ANXA2 | hsa-miR-579 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 4 |
| ANXA2 | hsa-miR-767-5b | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 4 |
Optimal miRNA-ANXA2 interactions
predicted by miRanda
miRNAs with the highest binding scores for
ANXA2 mRNA were identified using miRanda software, which
incorporates thermodynamic and sequence conservation scores. This
analysis identified the following 4 miRNAs that met the imposed
criteria (mirSVR, ≤-0.1; phastCons, 0.5–0.7): miR-206, miR-1,
miR-613 and miR-425. miR-206 was predicted to bind to ANXA2
mRNA with the highest stability and specificity, suggesting that it
may exhibit preferential binding to ANXA2 mRNA (Table II).
| Table II.Scores of mirSVR and PhastCons among
12 predicted miRNAs. |
Table II.
Scores of mirSVR and PhastCons among
12 predicted miRNAs.
|
| mirSVR score | Ranking | PhastCons
score |
|---|
| hsa-miR-206 | −1.1965 | 1 | 0.6944 |
| hsa-miR-1 | −1.1964 | 2 | 0.6944 |
| hsa-miR-613 | −1.1865 | 3 | 0.6944 |
| hsa-miR-425 | −1.1787 | 4 | 0.6451 |
| hsa-miR-520h | −0.8678 | 5 | 0.6697 |
| hsa-miR-185 | −0.7357 | 6 | 0.6459 |
| hsa-miR-579 | −0.4511 | 7 | 0.5177 |
| hsa-miR-9 | −0.2252 | 8 | 0.6459 |
| hsa-miR-155 | −0.2098 | 9 | 0.5084 |
| hsa-miR-767 | −0.1488 | 10 | 0.5022 |
| hsa-miR-890 | −0.1193 | 11 | 0.6944 |
| hsa-miR-20b | −0.0235 | 12 | 0.6697 |
Expression of ANXA2 protein in
prostate cancer and BPH tissues
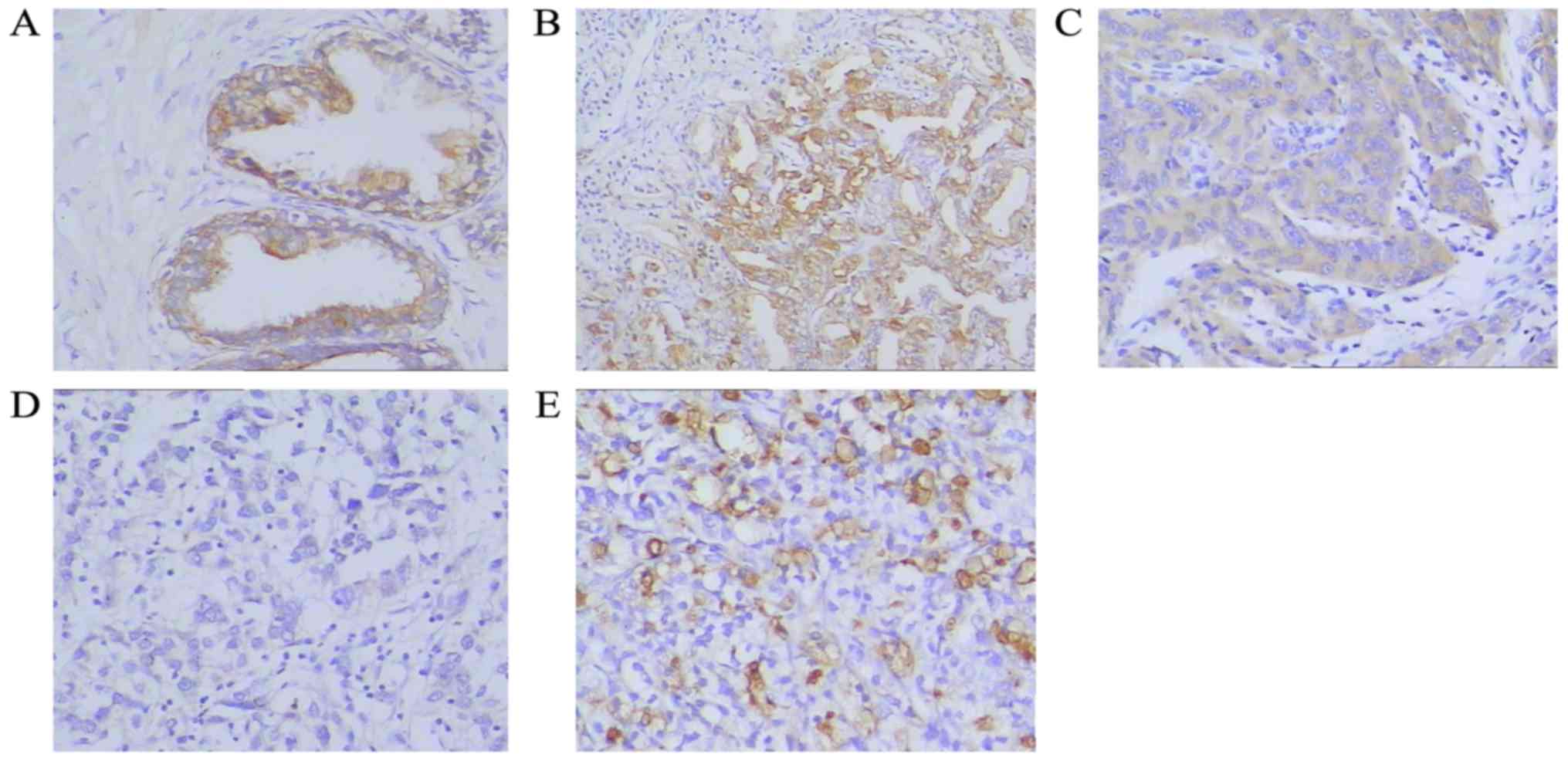
Expression of ANXA2 protein in tissue specimens was
assessed by IHC. The proportion of samples positively stained for
ANXA2 was significantly higher in the BPH samples (95%, 19/20)
compared with the prostate cancer samples (51.7%, 31/60;
P<0.001). Additionally, 76.7% (23/30) of the metastatic prostate
cancer tissues exhibited positive ANXA2 staining, which was a
significantly higher proportion than that of the prostate cancer
samples (P<0.001 Fig. 1; Table III).
| Table III.Expression of ANXA2 in prostate
cancer and BPH tissues. |
Table III.
Expression of ANXA2 in prostate
cancer and BPH tissues.
|
|
| Immunohistochemical
staining scores |
|---|
|
|
|
|
|---|
|
| n | Weak positive or
negative (0–2) | Positive (3–4) | Strong positive
(5–6) | P-value |
|---|
| BPH | 20 | 1 | 3 | 16 |
<0.001a |
| Prostate
cancer |
|
|
|
|
<0.001b |
|
Highly-differentiated | 20 | 6 | 12 | 2 |
|
|
Moderately-differentiated | 34 | 18 | 16 | 0 |
|
|
Poorly-differentiated | 6 | 5 | 1 | 0 |
|
| Metastatic prostate
cancer | 30 | 7 | 8 | 15 |
<0.001c |
Association between the expression of
miR-206 and ANXA2 in prostate cancer and BPH tissues
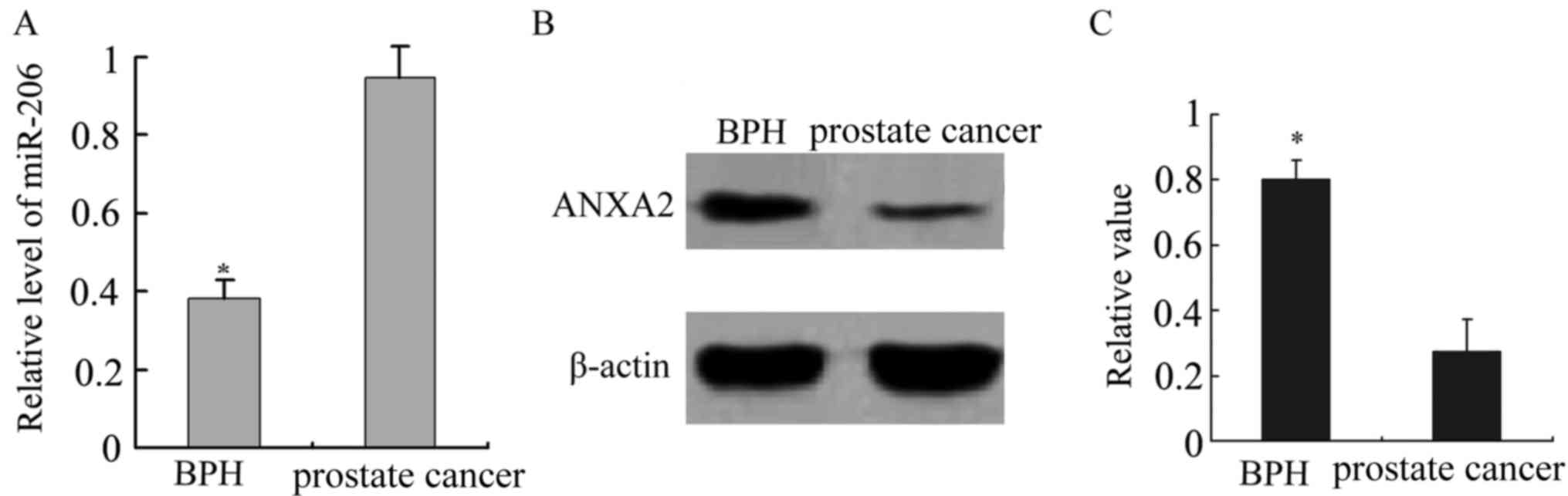
The expression of miR-206 was significantly higher
in prostate cancer tissues than in BPH tissues, while an inverse
expression pattern was observed for ANXA2 (P<0.001; Fig. 2). miR-206 expression was significantly
increased in the prostate cancer samples compared with the BPH
samples (P<0.001). ANXA2 expression levels were significantly
decreased in the prostate cancer samples, but increased in the BPH
samples (P<0.001). By contrast, miR-206 was lowly expressed and
ANXA2 was highly expressed in the BPH samples. Therefore, there was
a negative association between the expression of miR-206 and
ANXA2.
In vitro invasion assay of a prostate
cancer cell line
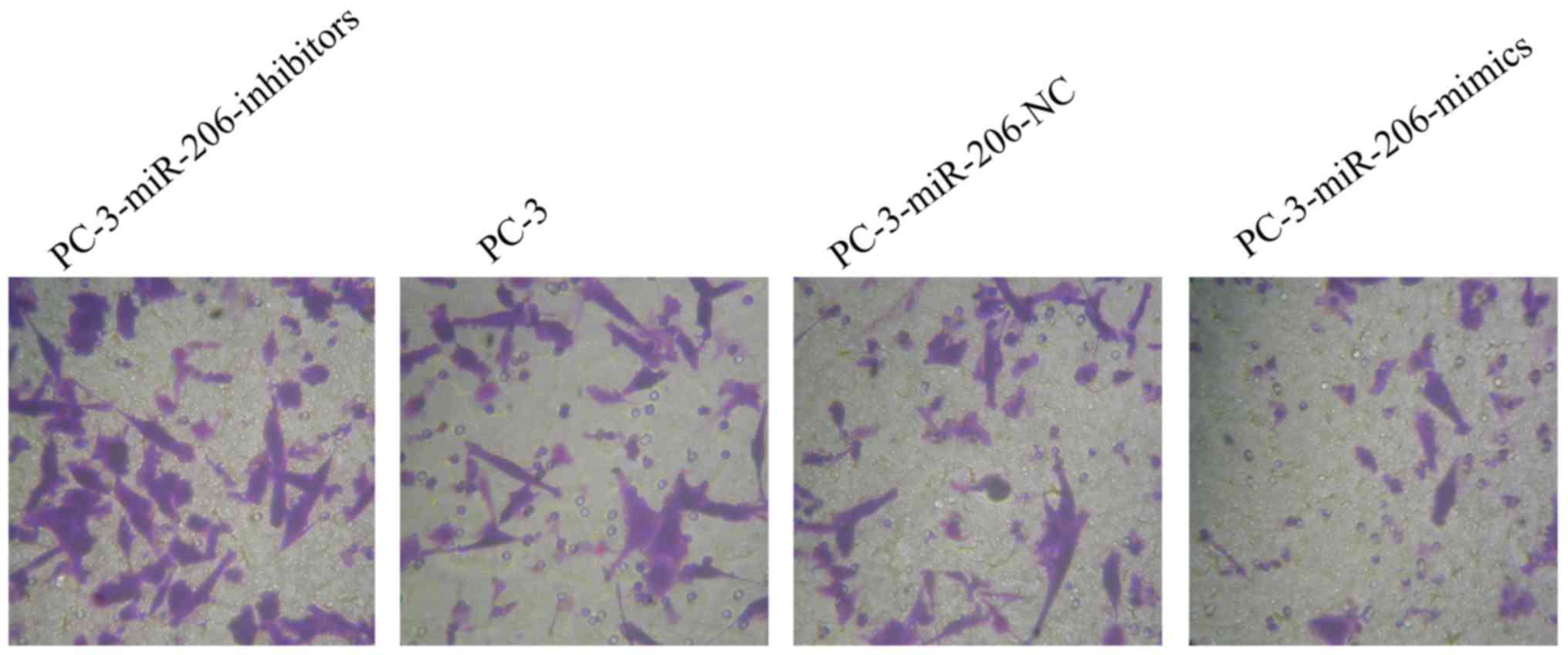
The invasion assay demonstrated that a significantly
lower number of PC-3-miR-206-inhibitor cells exhibited invasive
behavior than untransfected PC-3 or PC-3-miR-206-NC cells (Fig. 3; P<0.001, data not shown). By
contrast, the number of invasive PC-3-miR-206-mimic cells was
significantly higher than the number of invasive PC-3 or
PC-3-miR-206-NC cells. These results indicated that downregulation
of miR-206 may promote the invasive capacity of PC-3 prostate
cancer cells in vitro (Table
IV; Fig. 3; P<0.001).
| Table IV.Changes in the effect of miR-206
expression on the invasion of prostate cancer cells in
vitro. |
Table IV.
Changes in the effect of miR-206
expression on the invasion of prostate cancer cells in
vitro.
| Group | no. invaded cells
(/HPF) | P-value |
|---|
|
PC-3-miR-206-inhibitors | 107±16a | <0.001 |
| PC-3 | 52±9 |
|
|
PC-3-miR-206-NC | 48±8 |
|
|
PC-3-miR-206-mimics | 21±4b | <0.001 |
Association between the expression of
miR-206 and that of ANXA2 and WNT pathway proteins
Downregulation of miR-206 in PC-3-miR-206-inhibitor
cells was associated with increased expression of ANXA2 and
E-cadherin and decreased expression of N-cadherin and vimentin
(Fig. 4). By contrast, upregulation
of miR-206 in PC-3-miR-206-mimic cells resulted in downregulation
of ANXA2 and E-cadherin and upregulation of N-cadherin and vimentin
(P<0.05; Fig. 4). PC-3 and
PC-3-miR-206-NC cells exhibited no significant differences in the
expression of ANXA2, E-cadherin, N-cadherin or vimentin (Fig. 4).
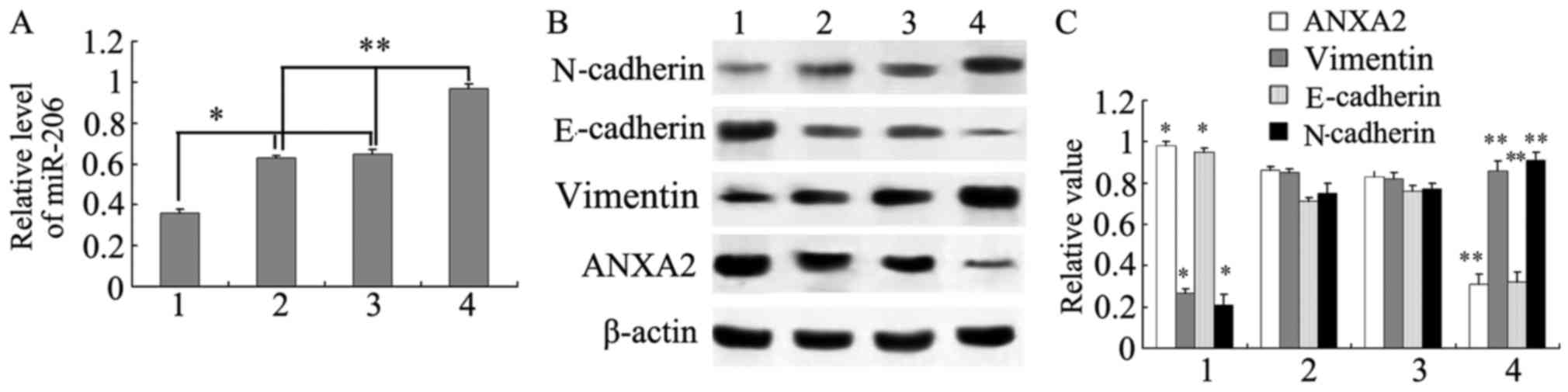 | Figure 4.Association between the expression of
miR-206, ANXA2, E-cadherin, N-cadherin and vimentin in prostate
cancer cell lines. (A) Relative expression level of miR-206. (B)
Western blot analysis of protein expression in 1)
PC-3-miR-206-inhibitor, 2) PC-3, 3) PC-3-miR-206-NC and 4)
PC-3-miR-206-mimic cells. (C) Statistical analysis of western
blotting results. *P<0.05, PC-3-miR-206 inhibitor vs. PC-3 or
PC-3-miR-206-NC; **P<0.05, PC-3-miR-206 mimic vs. PC-3 or
PC-3-miR-206-NC. ANXA2, Annexin A2; NC, negative control; miR,
microRNA; HPF, high power field. |
The results indicated that transfection of miR-206
mimics into PC-3 cells inhibited the expression of ANXA2, resulting
in the downregulation of E-cadherin and vimentin and the
upregulation of N-cadherin. However, transfection of miR-206
inhibitors into PC-3 cells upregulated ANXA2 expression and
concomitantly downregulated E-cadherin. Therefore, the miR-206
target ANXA2 regulates the expression of E-cadherin,
N-cadherin and vimentin and may be involved in the EMT.
Discussion
Changes in the expression and distribution of ANXA2
have been detected in many types of cancer, such as laryngeal
cancer, pancreatic cancer, breast cancer, and so on (15–17). ANXA2
expression has also been associated with the differentiation of
squamous cell lung cancer (18).
ANXA2 content was demonstrated to be high in human hepatocellular
cancer cells but not in embryonic tissues, normal liver cells or
injured liver cells, suggesting that ANXA2 may serve a key role in
hepatocellular carcinoma (19). A
study undertaken by Kirshner et al (20) revealed that respiratory cilia, pleural
mesothelial and alveolar epithelial cells exhibit a high expression
of ANXA2. Expression of ANXA2 in prostate cancer differs from that
in other tumor types. Immunohistochemical studies have demonstrated
that ANXA2 highly expressed in >50% of glands in the majority
(>85%) of patients with BPH (21).
A number of studies have demonstrated that changes in the
expression and distribution of ANXA2 are associated with the
invasion and metastasis of various tumor types (22,23).
However, the exact molecular mechanism by which this occurs remains
unclear.
In the present study, a significantly higher
proportion of BPH samples than prostate cancer samples were
positively stained for ANXA2 protein. Furthermore, a significantly
higher proportion of metastatic cancer samples than prostate cancer
samples expressed ANXA2. Therefore, the expression of ANXA2 may be
a potential marker for the progression and development of prostate
cancer.
The interaction between miRNAs and mRNAs occurs
through complementary binding of base pairs. However, the
interaction is not usually fully complementary, and several
programs have been developed to predict miRNA-mRNA binding and to
identify the specific target genes of miRNAs (24). Due to the fact that the bioinformatics
programs use algorithms that focus on different aspects of the
interactions between miRNAs and mRNAs, the prediction results are
not entirely consistent between programs. Therefore, a previous
study employed a combination of predictive programs for analyses
(25). In the present study,
ANXA2 mRNA was predicted by miRWalk online program, which
contains 10 software programs), to be a target for 12 miRNAs, of
which the interaction with miR-206 was predicted to have the
highest stability and specificity. This suggested that miR-206 may
preferentially target ANXA2 mRNA.
The present study also revealed that the expression
of miR-206 was higher than that of ANXA2 in prostate cancer
samples, while an inverse association was observed in BPH tissues,
suggesting a negative association between the expression of miR-206
and ANXA2. These findings further supported the possibility that
ANXA2 may be a target of miR-206.
Previous studies have demonstrated that miR-206 acts
as a tumor suppressor, and that its expression is downregulated in
many tumor types compared with normal cells or tissues (26). The expression of miR-206 is
downregulated in renal cell carcinoma, and overexpression of
miR-206 targets cell division kinase CDK9 and CDK4, inducing cell
cycle arrest and inhibiting cell proliferation (26). miR-206 expression is also
downregulated in colon cancer cells, and overexpression of miR-206
targets d-MET and inhibits cell proliferation and metastasis
(27). miR-206 directly targets the
oncogenes Kirsten rat sarcoma proto-oncogene and ANXA2, thereby
acting as tumor suppressor in human pancreatic ductal
adenocarcinoma cells by blocking cell cycle progression, and cell
proliferation, migration and invasion (28). The miRNA functional analysis and a
luciferase reporter assay demonstrated that miR-206 effectively
downregulated the expression of ANXA2 by binding to the
3′-untranslated region of the ANXA2 mRNA in pulmonary artery smooth
muscle cells (29). However, there
have been no previous studies on miR-206 expression in prostate
cancer.
EMT is a complex and dynamic process. One of the
early events is remodeling of the cytoskeleton, which is
accompanied by increased expression of mesenchymal markers,
including N-cadherin and vimentin, and decreased expression of
epithelial markers, such as E-cadherin (30). A previous study demonstrated that low
expression of E-cadherin may induce EMT (31). N-cadherin is a cell surface adhesion
molecule and a major structural element in intercellular adhesion.
Its principal function is to mediate cell adhesion and migration
(32). The expression of N-cadherin
is increased in epithelial malignant tumors and is associated with
the EMT (33). Recent studies have
demonstrated that E-cadherin and N-cadherin are aberrantly
expressed in numerous tumor types and serve an important role in
the occurrence, development, invasion and metastasis of tumors
(34,35).
In the present study, miR-206 was demonstrated to
promote the invasive ability of prostate cancer cells in
vitro. Increased expression of miR-206 inhibited the expression
of ANXA2, resulting in downregulation of E-cadherin, and
upregulation of N-cadherin and vimentin. By contrast, decreased
expression of miR-206 increased the expression of ANXA2, resulting
in upregulation of E-cadherin and downregulation of N-cadherin and
vimentin. These results indicated that miR-206 may bind to ANXA2
and inhibit its expression, which, in turn, regulates the EMT and
inhibits the invasion and metastasis of prostate cancer cells.
Acknowledgements
Not applicable.
Funding
The Hunan Province Science Foundation (grant no.
2015JC3088).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
NY contributed to the conception and design of the
work. LW, JL, LL, JH and XC acquired and analyzed the data for the
work. ZL designed the work and corrected the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was certified by the Ethics
Committee of the Second Affiliated Hospital of University of South
China, and all participants provided written informed consent.
Consent for publication
All participants provided written consent agreeing
for publication of the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nandana S and Chung LW: Prostate cancer
progression and metastasis: Potential regulatory pathways for
therapeutic targeting. Am J Clin Exp Urol. 2:92–101.
2014.PubMed/NCBI
|
|
2
|
Simons JW: Prostate cancer immunotherapy:
Beyond immunity to curability. Cancer Immunol Res. 2:1034–1043.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Monastyrskaya K: Functional association
between regulatory RNAs and the annexins. Int J Mol Sci.
19:E5912018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mussunoor S and Murray GI: The role of
annexins in tumour development and progression. J Pathol.
216:131–140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mortimer JC, Laohavisit A, Macpherson N,
Webb A, Brownlee C, Battey NH and Davies JM: Annexins:
Multifunctional components of growth and adaptation. J Exp Bot.
59:533–544. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang M, Chen D, Zhen Z, Ao J, Yuan X and
Gao X: Annexin A2 positively regulates milk synthesis and
proliferation of bovine mammary epithelial cells through the mTOR
signaling pathway. J Cell Physiol. 233:2464–2475. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun MY, Xing RH, Gao XJ, Yu X, He HM, Gao
N, Shi HY, Hu YY, Wang QX, Xu JH and Hou YC: ANXA2 regulates the
behavior of SGC-7901 cells. Asian Pac J Cancer Prev. 14:6007–6012.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jeon YR, Kim SY, Lee EJ, Kim YN, Noh DY,
Park SY and Moon A: Identification of annexin II as a novel
secretory biomarker for breast cancer. Proteomics. 13:3145–3156.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lokman NA, Elder AS, Ween MP, Pyragius CE,
Hoffmann P, Oehler MK and Ricciardelli C: Annexin A2 is regulated
by ovarian cancer-peritoneal cell interactions and promotes
metastasis. Oncotarget. 4:1199–1211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang HJ, Yao DF, Yao M, Huang H, Wang L,
Yan MJ, Yan XD, Gu X, Wu W and Lu SL: Annexin A2 silencing inhibits
invasion, migration, and tumorigenic potential of hepatoma cells.
World J Gastroenterol. 19:3792–3801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zare M, Bastami M, Solali S and Alivand
MR: Aberrant miRNA promoter methylation and EMT-involving miRNAs in
breast cancer metastasis: Diagnosis and therapeutic implications. J
Cell Physiol. 233:3729–3744. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng J, Fu Z, Guo J, Lu W, Wen K, Chen W,
Wang H, Wei J and Zhang S: Overexpression of peroxiredoxin 2
inhibits TGF-β1-induced epithelial-mesenchymal transition and cell
migration in colorectal cancer. Mol Med Rep. 10:867–873. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bezdenezhnykh N, Semesiuk N, Lykhova O,
Zhylchuk V and Kudryavets Y: Impact of stromal cell components of
tumor microenvironment on epithelial-mesenchymal transition in
breast cancer cells. Exp Oncol. 36:72–78. 2014.PubMed/NCBI
|
|
14
|
Mou H, Yu L, Zheng X, Liao Q, Hou X and Wu
Y: p16 gene expression in pancreatic cancer tissue and its
importance in diagnosis. J Biol Regul Homeost Agents. 31:1043–1047.
2017.PubMed/NCBI
|
|
15
|
Luo S, Xie C, Wu P, He J, Tang Y, Xu J and
Zhao S: Annexin A2 is an independent prognostic biomarker for
evaluating the malignant progression of laryngeal cancer. Exp Ther
Med. 14:6113–6118. 2017.PubMed/NCBI
|
|
16
|
Murphy AG, Foley K, Rucki AA, Xia T,
Jaffee EM, Huang CY and Zheng L: Stromal Annexin A2 expression is
predictive of decreased survival in pancreatic cancer. Oncotarget.
8:106405–106414. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shetty P, Patil VS, Mohan R, D'souza LC,
Bargale A, Patil BR, Dinesh US, Haridas V and Kulkarni SP: Annexin
A2 and its downstream IL-6 and HB-EGF as secretory biomarkers in
the differential diagnosis of Her-2 negative breast cancer. Ann
Clin Biochem. 54:463–471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui JW and Wang YL: Expression and
function of Annexin II in lung cancer tissue. Asian Pac J Trop Med.
6:150–152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong Z, Yao M, Zhang H, Wang L, Huang H,
Yan M, Wu W and Yao D: Inhibition of Annexin A2 gene transcription
is a promising molecular target for hepatoma cell proliferation and
metastasis. Oncol Lett. 7:28–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kirshner J, Schumann D and Shively JE:
CEACAM1, a cell-cell adhesion molecule, directly associates with
annexin II in a three-dimensional model of mammary morphogenesis. J
Bio Chem. 278:50338–50345. 2003. View Article : Google Scholar
|
|
21
|
Yee DS, Narula N, Ramzy I, Boker J,
Ahlering TE, Skarecky DW and Ornstein DK: Reduced annexin II
protein expression in high-grade prostatic intraepithelial
neoplasia and prostate cancer. Arch Pathol Lab Med. 131:902–908.
2007.PubMed/NCBI
|
|
22
|
Ohno Y, Izumi M, Kawamura T, Nishimura T,
Mukai K and Tachibana M: Annexin II represents metastatic potential
in clear-cell renal cell carcinoma. Br J Cancer. 101:287–294. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang W, Zhao P, Xu XL, Cai L, Song ZS,
Cao DY, Tao KS, Zhou WP, Chen ZN and Dou KF: Annexin A2 promotes
the migration and invasion of human hepatocellular carcinoma cells
in vitro by regulating the shedding of CD147-harboring
microvesicles from tumor cells. PLoS One. 8:e672682013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oh M, Rhee S, Moon JH, Chae H, Lee S, Kang
J and Kim S: Literature-based condition-specific miRNA-mRNA target
prediction. PLoS One. 12:e01749992017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk-database: Prediction of possible miRNA binding sites by
‘walking’ the genes of three genomes. J Biomed Inform. 44:839–847.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiao H, Xiao W, Cao J, Li H, Guan W, Guo
X, Chen K, Zheng T, Ye Z, Wang J and Xu H: miR-206 functions as a
novel cell cycle regulator and tumor suppressor in clear-cell renal
cell carcinoma. Cancer Lett. 374:107–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ren XL, He GY, Li XM, Men H, Yi LZ, Lu GF,
Xin SN, Wu PX, Li YL, Liao WT, et al: MicroRNA-206 functions as a
tumor suppressor in colorectal cancer by targeting FMNL2. J Cancer
Res Clin Oncol. 142:581–592. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Keklikoglou I, Hosaka K, Bender C, Bott A,
Koerner C, Mitra D, Will R, Woerner A, Muenstermann E, Wilhelm H,
et al: MicroRNA-206 functions as a pleiotropic modulator of cell
proliferation, invasion and lymphangiogenesis in pancreatic
adenocarcinoma by targeting ANXA2 and KRAS genes. Oncogene.
34:4867–4878. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen L, Li YS, Cui J, Ning JN, Wang GS,
Qian GS, Lu KZ and Yi B: MiR-206 controls the phenotypic modulation
of pulmonary arterial smooth muscle cells induced by serum from
rats with hepatopulmonary syndrome by regulating the target gene,
annexin A2. Cell Physiol Biochem. 34:1768–1779. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scanlon CS, Van Tubergen EA, Inglehart RC
and D'Silva NJ: Biomarkers of epithelial-mesenchymal transition in
squamous cell carcinoma. J Dent Res. 92:114–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martínez-Ramírez AS, Garay E,
García-Carrancá A and Vázquez-Cuevas FG: The P2RY2 receptor induces
carcinoma cell migration and EMT through cross-talk with epidermal
growth factor receptor. J Cell Biochem. 117:1016–1026. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carvalho-Cruz P, Alisson-Silva F,
Todeschini AR and Dias WB: Cellular glycosylation senses metabolic
changes and modulates cell plasticity during epithelial to
mesenchymal transition. Dev Dyn. 247:481–491. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kaushik NK, Kaushik N, Yoo KC, Uddin N,
Kim JS, Lee SJ and Choi EH: Low doses of PEG-coated gold
nanoparticles sensitize solid tumors to cold plasma by blocking the
PI3K/AKT-driven signaling axis to suppress cellular transformation
by inhibiting growth and EMT. Biomaterials. 87:118–130. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu AQ, Ding Y, Li CL, Yang Y, Yan SR and
Li DS: TALEN-induced disruption of Nanog expression results in
reduced proliferation, invasiveness and migration, increased
chemosensitivity and reversal of EMT in HepG2 cells. Oncol Rep.
35:1657–1663. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen HN, Yuan K, Xie N, Wang K, Huang Z,
Chen Y, Dou Q, Wu M, Nice EC, Zhou ZG and Huang C: PDLIM1
Stabilizes the E-Cadherin/β-Catenin complex to prevent
epithelial-mesenchymal transition and metastatic potential of
colorectal cancer cells. Cancer Res. 76:1122–1134. 2016. View Article : Google Scholar : PubMed/NCBI
|