Introduction
Colorectal cancer (CRC) is one of the most common
causes of cancer-related deaths worldwide, with high mortality and
morbidity in both genders (1–3). Current therapeutic strategies against
CRC consist of curative surgery and postoperative adjuvant
chemotherapy. Despite the slow and steady improvement in the
overall 5-year survival rate during the past decades (3,4), prognosis
of patients with CRC remains poor due to post-surgical recurrence
and fatal distant metastasis. According to a recent statistic, the
5-year relative survival ranges from >90% for patients with
stage I disease to slightly >10% in patients with stage IV
disease (5). Therefore, it is a great
urgency to search for novel biomarkers, suitable intervention
strategies in order to improve patient outcomes.
With the rapid development of genome and
transcriptome sequencing technologies and implementation of
genomics consortiums, such as ENCODE and FANTOM, the classic view
of the transcriptome landscape and its mRNA-centric paradigm for
transcript annotation has undergone a fundamental change (6). Current knowledge has well recognized
that the vast majority of genome serves as the template for the
transcription of noncoding RNAs. For example, small noncoding RNAs,
particularly microRNAs, have been extensively investigated for
several decades, and their biological functions in various cancers
have been uncovered (7). However, as
the fundamental elements of noncoding RNAs, long noncoding RNAs
(lncRNAs) has not received a wide recognition and the function of
lncRNAs in numerous cancers still remains to be elucidated.
LncRNAs are defined as a class of RNAs with length
>200 nucleotides. Ever since its discovery, lncRNAs have been
identified as key regulators of various biological processes,
including cell proliferation, differentiation, apoptosis,
migration, and invasion (8). The
lncRNA UCC, for instance, has recently been identified to promote
CRC cell growth and invasion by sponging miR-143 (9).
LncRNA BX357664 is a novel lncRNA that has been
identified from the analysis of normalized microarray data and
further quantitative polymerase chain reaction (qPCR) analysis.
BX357664 might be a potential biomarker involved in renal cell
carcinoma (10). Furthermore, it was
demonstrated that BX357664 regulates cell proliferation and
epithelial-to-mesenchymal transition (EMT) via inhibition of
transforming growth factor (TGF)-β1/p38/heat shock protein (HSP) 27
signaling in renal cell carcinoma (11). These data implicated that BX357664
might have critical roles in human tumorigenesis. However, the role
of BX357664 in other types of solid tumors remains largely
unknown.
The present study aimed to investigate the role of
BX357664 in human CRC. To this end, the expression levels of
BX357664 in human CRC tissues and in CRC cell lines were examined.
Expression of BX357664 was then modulated in order to assess the
functional changes of CRC cells in response to BX357664 alteration.
Cell survival was also examined. The present data imply that
BX357664 may be a critical tumor suppressor that might serve as a
drug target in the treatment of CRC in the clinic.
Materials and methods
Human samples
Colorectal cancer tissues from 80 patients (age
range, 40–80 years; mean age, 58 years; male:female ratio, 47:33),
admitted to the Department of General Surgery, Qilu Hospital of
Shandong University (Jinan, China) between March 2016 and March
2017, were collected via surgical resection, frozen into liquid
nitrogen immediately once dissected from patients, and stored at
−80°C. Matched adjacent non-cancerous tissues were also obtained
from each CRC patient. All patients expressed their full intentions
to participate in the present study and written consent forms were
obtained from each patient. The present study was approved by the
Ethics Committee of the Anqiu People's Hospital (Weifang,
China).
Cell culture and plasmid
transfection
Control cells HIEC-6 were purchased from the
American Type Tissue Collection (Manassas, VA, USA). CRC cell lines
COLO 205, HCT116 and HT-29 were purchased from the Cell Bank of
Chinese Academy of Sciences (Shanghai, China). All of the cell
lines were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) as well as 1% antibiotics
(penicillin/streptomycin). The cell lines were cultured in a 37°C
incubator with 5% CO2 and the culture media were
replaced every other day, otherwise as stated. The full sequence of
BX357664 was cloned from human genome extracted from normal human
liver, and then cloned into pcDNA 3.1 vector (catalog no., 52535;
Addgene, Inc., Cambridge, MA, USA) at the sites of the restriction
enzymes EcoRI and XhoI. The correct sequence was
confirmed by sequencing using the T7 promoter forward primer
(Shenggong Biology Engineering Technology Service, Ltd., Shanghai,
China). The transfections were performed with Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufactures' instructions, using a dose of 2.5 µl for 1.5 µg
plasmid DNA. Six h following transfection, the culture medium was
replaced with fresh DMEM.
RNA isolation and reverse
transcription (RT)-qPCR
Total RNA from CRC patient tissues and cultured CRC
cells were extracted with TRIzol reagent (Takara Biotechnology Co.,
Ltd., Dalian, China) in a dilution of 1 ml for each well in a
six-well plate. The RNA quality and quantity were determined using
a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.)
at the absorbance of 260 and 280 nm. Reverse transcription of
first-strand cDNA was performed with PrimeScript RT Master Mix
(Perfect Real Time; Takara Biotechnology Co., Ltd.), following the
manufacturer's protocol. All PCR reactions were performed in an ABI
PRISM 7900 Real-Time system (Thermo Fisher Scientific, Inc.) with
the SYBR Premix Ex Taq kit (Takara Biotechnology Co., Ltd.). The
thermocycling protocol was as follows: Initial denaturation at 95°C
for 2 min, followed by 35 repeats of the three-step cycling program
consisting of 30 sec at 95°C (denaturation), 1 min at 53°C (primer
annealing) and 30 sec at 72°C (elongation), followed by a final
extension step for 10 min at 72°C. The housekeeping gene GAPDH was
included as an internal control. Primers were purchased from
Shenggong Biology Engineering Technology Service, Ltd. (Shanghai,
China) with the following sequences: BX357664, forward,
5′-GGCGTGGTTTTGATGGAGTG-3′, and reverse,
5′-AGGCTGCAGAGTTGAGATCG-3′; GAP DH, forward,
5′-GTGGACATCCGCAAAGAC-3′ and reverse, 5′-AAAGGGTGTAACGCAACTA-3′.
All quantitative data were normalized to GAPDH using the
2−ΔΔCq method (12).
Colony formation assay
HCT116 and HT-29 cells were transfected with pcDNA
3.1 vector with or without BX357664 sequence in six-well plates
with a density of 200 cells/well. After 2 weeks in a 37°C
incubator, cell colonies that contained >50 cells were counted
by staining with crystal violet (0.5%, 10 min) and observation
under a light microscope with a magnification of ×200 (Nikon
Corporation, Tokyo, Japan).
Cell cycle analysis
Prior to cell cycle analysis, HCT116 and HT-29 cells
were transfected with empty vector plasmid or BX357664 plasmid for
48 h. Next, cells were collected with low speed centrifugation (840
× g, 5 min, 4°C) and fixed with pre-iced ethanol (70%) for 10 min
at 4°C. The cells were washed and re-suspended in cold PBS and
incubated at 37°C for 30 min with 10 mg/ml RNase and 1 mg/ml
propidium iodide (PI; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The % of cells in each phase of the cell cycle was
determined using the Cell Quest Pro acquisition software (BD
Biosciences, Franklin Lakes, NJ, USA).
Cell viability determination
Both HCT116 and HT-29 cells were seeded in a 96-well
plate at a concentration of 1,000 cells/well. Following incubation
for 24 h, cells were transfected with BX357664-expressing plasmid
or control vector. Cell proliferation was examined in consecutive 5
days with a Cell Titer 96 AQueous Non-Radioactive Cell
Proliferation kit (Promega Corporation, Madison, WI, USA), as per
the manufacturers' protocol. The cell proliferative rate in each
group was determined by measuring the absorbance at 490 nm using a
microplate reader (Tecan, Männedorf, Switzerland). For data
presentation, absorbance for control cells at the first day was set
as 1 and used for normalization. Absorbance values for other groups
were then normalized to that of control at the first day.
Therefore, cell viability ratio was calculated as the absorbance in
others groups relative to that in control group at the first
day.
Transwell assay
For cell migration assays, HCT116 and HT-29 cells
were transfected with corresponding plasmid for 48 h and then
trypsinized, collected by low-speed centrifugation (840 × g, 4°C, 5
min) in serum-free medium. A total of 1×104 cells (200
µl) were seeded into the upper chamber of a Corning Transwell
chamber (8 µm pore). The lower chamber was filled with 600 µl
medium containing 10% FBS. Afterwards, the plate was incubated at
37°C and the cells are allowed to migrate for 24 h. Afterwards, the
membrane was fixed with pre-cooled methanol and stained with
crystal violet (1%) for 5 min at room temperature. Cell migration
was assessed by counting the cells that had migrated through the
membrane. Five random fields were selected and images were captured
under a Nikon light microscope (Nikon Corporation, Tokyo, Japan) at
a magnification of ×100. For cell invasion assays, the membrane was
pre-coated with Matrigel (Corning Incorporated, Corning, NY, USA)
for 6 h in 37°C incubator.
Wound-healing assay
HCT116 and HT-29 cells were transfected with
BX357664-expressing or control plasmid, cultured in DMEM in a
six-well culture plate at a density of 5×105 cells/well
and allowed to grow to a confluence of 90% overnight. Next day, the
culture medium was replaced with serum-free DMEM, and then two
lines were scratched in the single cell layer with a 10 µl pipette
tip and the cells were washed with PBS three times. Following
incubation for 12 h, images of the migrating cells were observed
and captured using a Nikon light microscope at the cross of two
straight lines.
Western blot analysis
Molecules included in the process of EMT and cell
cycle progression were evaluated by western blot analysis. Briefly,
HCT116 and HT-29 cells were transfected with BX357664-expressing
plasmid for 48 h, and then total proteins were prepared with lysis
buffer (RIPA; Beyotime Institute of Biotechnology, Nantong, China)
on ice and quantified using a Bio-Rad protein assay reagent (Thermo
Fisher Scientific, Inc.). Equal amounts of protein (50 µg) were
separated with 10% SDS-PAGE and transferred to a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA). The membrane was
blocked for 1 h with 5% skimmed milk at room temperature and then
incubated with primary antibodies overnight at 4°C. The primary
antibodies against Cyclin B1 (cat. no. sc-70898; 1:1,000),
cell-division cycle (CDC) 25C (cat. no. sc-327; 1:1,000), Cyclin D1
(cat. no. sc-4074; 1:1,000), E-Cadherin (cat. no. sc-71009;
1:1,000), N-Cadherin (cat. no. sc-53488; 1:1,000) and GAPDH (cat.
no. sc-32233; 1:1,000) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). After washing with TBST for
4 times (8 min each), the membrane was incubated with secondary
goat-anti-rabbit (cat. no. sc-2004) or goat-anti-mouse (cat. no.
sc-2005) antibody (Santa Cruz Biotechnology, Inc.) for 1 h at 37°C
with a dilution of 1:1,000. Finally, the proteins were quantified
using ECL Prime Western Blotting Detection reagent (GE Healthcare,
Chicago, IL, USA) and an ImageQuant LAS 4000 Mini Biomolecular
Imager (GE Healthcare).
Flow cytometric analysis of cell
apoptosis
The annexin V/PI assay was performed as per the
manufacturer's instructions (Invitrogen; Thermo Fisher Scientific,
Inc.). Briefly, HCT116 and HT-29 cells were plated into 6-well
plates and transfected with control or BX357664-expressing plasmid
for 48 h. Afterwards, cells were washed with cold PBS, trypsinized,
and re-suspended in 100 µl of binding buffer with 2.5 µl
fluorescein isothiocyanate (FITC)-conjugated annexin V and 1 µl PI
(100 µg/ml). Afterwards, cells were incubated at room temperature
for 15 min in the dark. A total of at least 10,000 cells were
collected and analyzed by flow cytometry for both cell lines (Cell
Quest Pro acquisition software, BD Biosciences, Franklin Lakes, NJ,
USA).
Determination of caspase
activities
The activities of caspase-3, caspase-8 and caspase-9
were determined by specific caspase activity kits (Beyotime
Institute of Biotechnology), following the manufacturer's
instructions. Briefly, cells were transfected with plasmids for 48
h. Afterwards, cell lysates were collected by low speed
centrifugation (840 × g, 5 min, 4°C). An equal amount of 10 µl
proteins from each sample were added into 96-well plates and mixed
with an aliquot of 80 µl reaction buffer supplied with caspase
substrates (2 mM). Following incubation at 37°C for 4 h, caspase
activities were determined using a Tecan plate reader at an
absorbance of 450 nm.
Statistical analysis
All experiments were repeated at least three times
in triplicate, showing reproducible results. All data were
presented as the mean ± standard deviation, except if otherwise
stated. Student's t-test analysis was used for the comparison of
numerical variables between two groups. One-way analysis of
variance was used for comparisons of numerical variables among
multiple groups, followed by the LSD post hoc test. Data were
analyzed with GraphPad Prism 6 (GraphPad Software Inc., San Diego,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
BX357664 is downregulated in CRC
patient tissues and cell lines
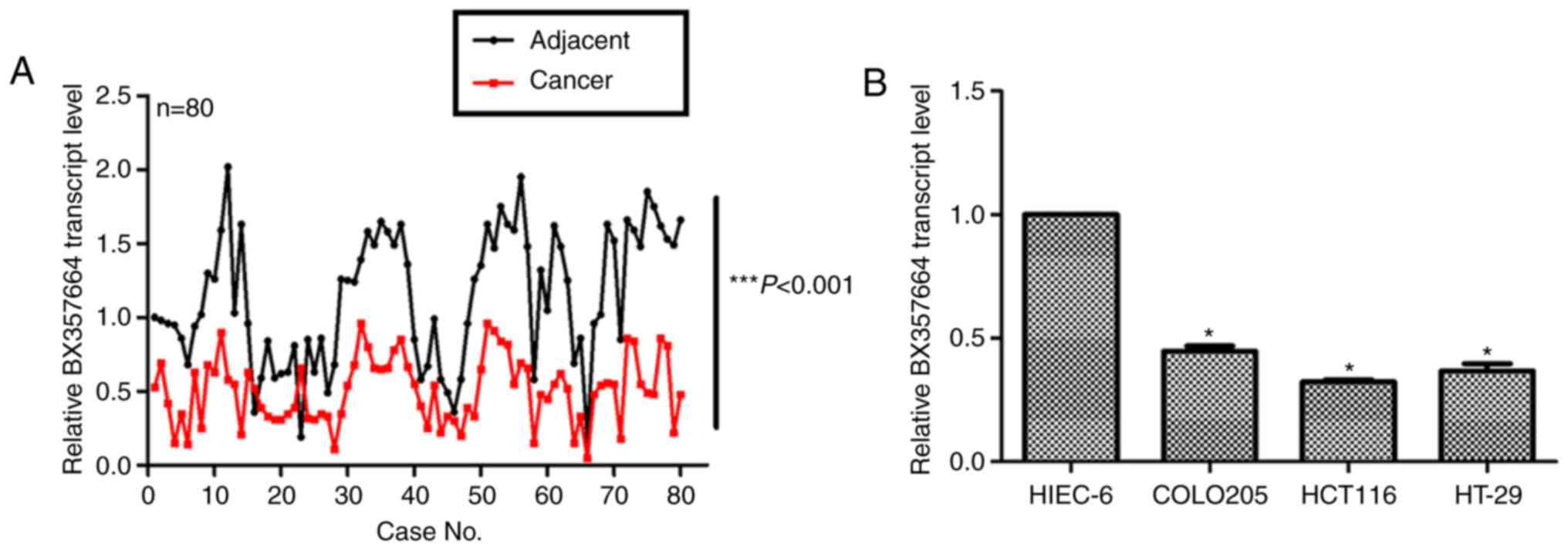
Initially, the expression profile of BX357664 was
examined in clinical CRC tissues. To this end, a total of 80 cases
with clinically diagnosed CRC were collected. Adjacent
non-cancerous tissues were included in the analysis as a control.
As illustrated in Fig. 1A, the
transcription levels of BX357664 in cancerous tissues were
significantly lower compared with adjacent normal tissues. The mean
transcription level of BX357664 in cancerous tissue was only 25–50%
of that in the adjacent ones (Fig.
1A; P<0.001). Furthermore, compared with the normal
intestinal epithelial control HIEC-6 cells, the expression of
BX357664 in CRC cell lines (COLO205, HCT116 and HT-29) was
significantly reduced (Fig. 1B).
These data suggested that BX357664 was downregulated in CRC.
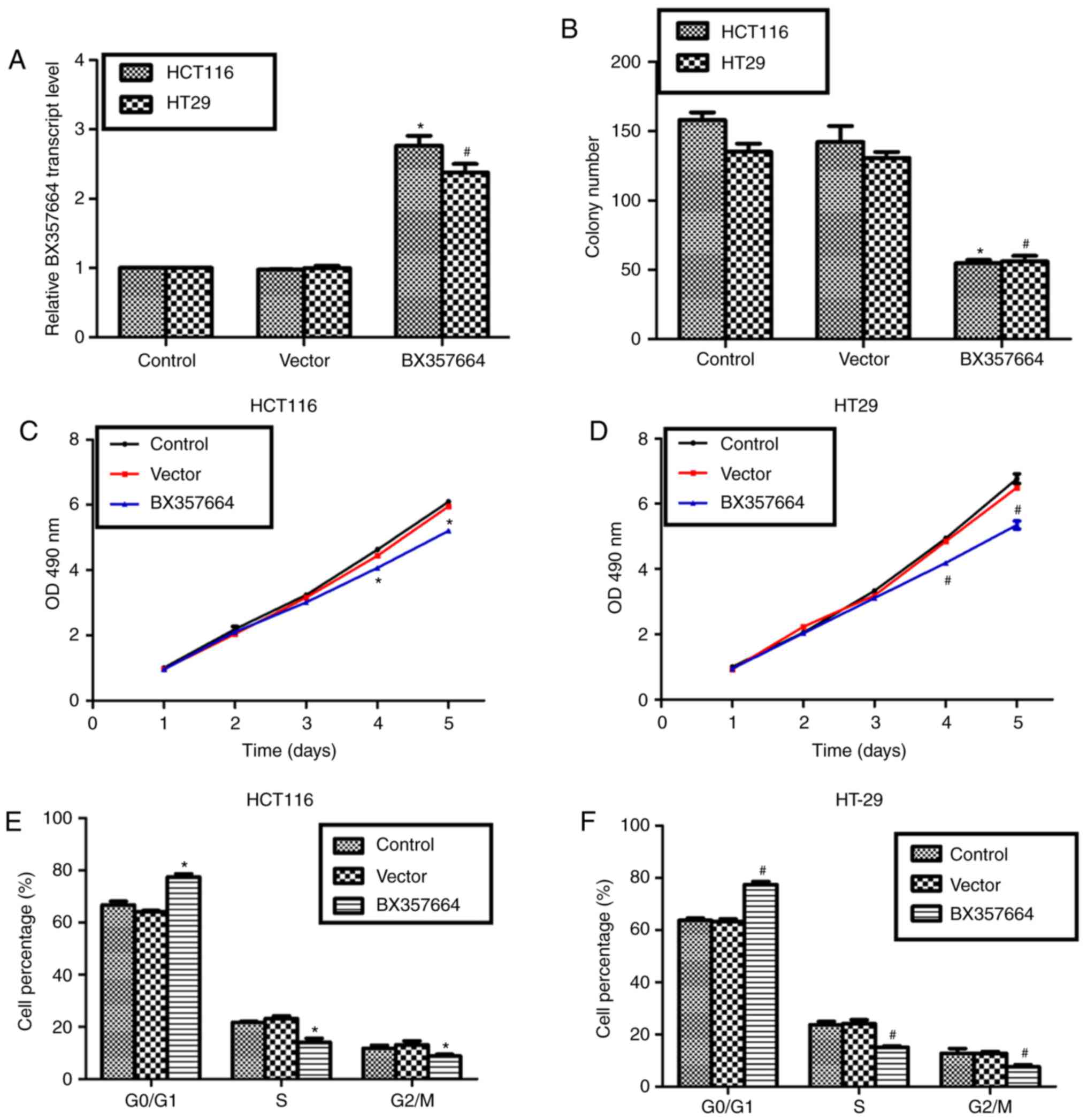
Overexpression of BX357664 inhibits
cell growth and arrests cell cycle in HCT116 and HT-29 cells
Next, BX357664 was overexpressed in HCT116 and HT-29
cells by transfection of an overexpression construct. Analysis of
transcription levels of BX357664 revealed that the overexpression
construct successfully elevated the expression of BX357664 in both
cell lines by up to 2–3 fold, compared with the cells transfected
with empty vector (Fig. 2A). With the
aid of this overexpression system, colony formation and cell
viability assays were performed. As presented in Fig. 2B, it was observed that transfection of
a vector plasmid resulted in insignificant changes on cell
capacities to form colonies. However, compared with an average of
150 colonies in control HCT116 cells and 135 in control HT-29
cells, BX357664-overexpressing cells exhibited a significantly
reduced average of 50 colonies (Fig.
2B). In the cell viability assay, it was observed that the
number of BX357664-overexpressing cells was ~70% of control HCT116
cells on the 5th day (Fig. 2C), while
the number of BX357664-overexpressing cells was ~65% of control
HT-29 cells on the 5th day (Fig. 2D).
In addition, cell cycle progression was assessed in control and
BX357664-overexpressing cells. Following transfection of BX357664
into HCT116 cells, the % of cells in the G0/G1 phase was
significantly increased, whereas the % of cells in the S and G2/M
phases was decreased accordingly (Fig.
2E). Likewise, in the HT-29 cells, cells were more accumulated
in the G0/G1 phase following overexpression of BX357664, whereas
control HT-29 cells were significantly more accumulated in the S
and G2/M phases (Fig. 2F). These data
suggested that overexpression of BX357664 led to cell growth
inhibition and cell cycle arrest at the G0/G1 phase.
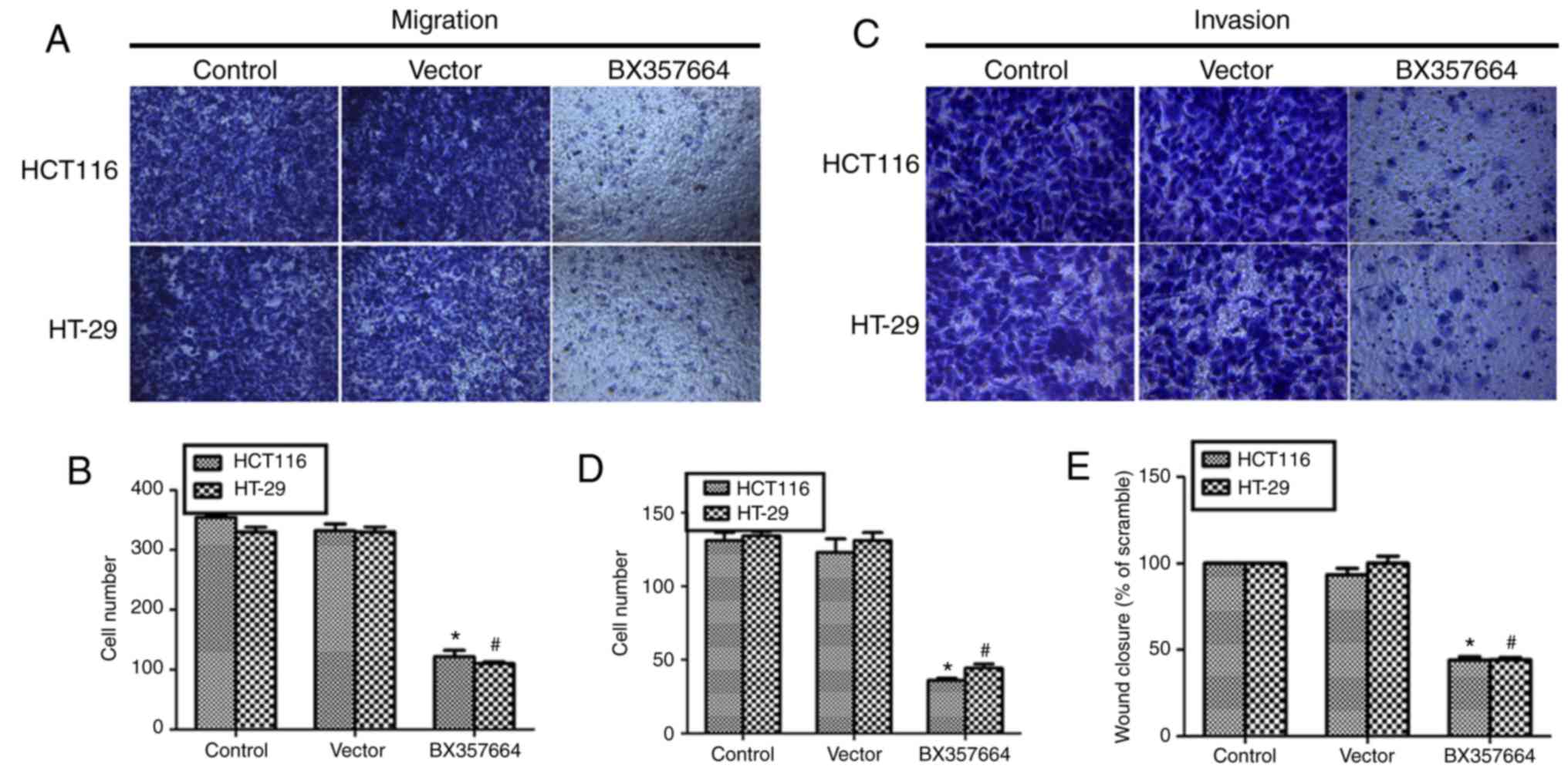
Overexpression of BX357664 inhibits
cell migration and invasion in HCT116 and HT-29 cells
The effects of BX357664 overexpression on cell
migration and invasion were next examined. In the transwell
migration assay, there were visibly less cells migrated to the
lower chamber observed in the BX357664-transfected group (Fig. 3A). Quantification of transmigrated
cells demonstrated that almost 340 cells migrated to the lower
chamber in the control groups, while only ~100 cells with BX357664
overexpression were observed in the lower chamber, indicating a 70%
decrease of migration capacity (Fig.
3B). Similarly, in the transwell invasion assay, cells that
invaded into the lower chamber were visibly fewer in the
BX357664-transfected HCT116 and HT-29 cells (Fig. 3C). In fact, an average of 126 control
HCT116 cells were counted in the lower chamber while 34 cells were
counted in the lower chamber of the BX357664-overexpressing HCT116
cells (Fig. 3D). For the HT-29 cell
line, only 36 cells invaded through the Matrigel compared with ~128
cells in the control groups (Fig.
3D). Finally, in the wound healing assay,
BX357664-overexpressing cells displayed significantly reduced
capacities to recover the scratched wound, as evidenced by the
lower rate of wound closure compared with the control groups
(Fig. 3E). These findings suggested
that overexpression of BX357664 inhibited cell migration and
invasion in CRC cells.
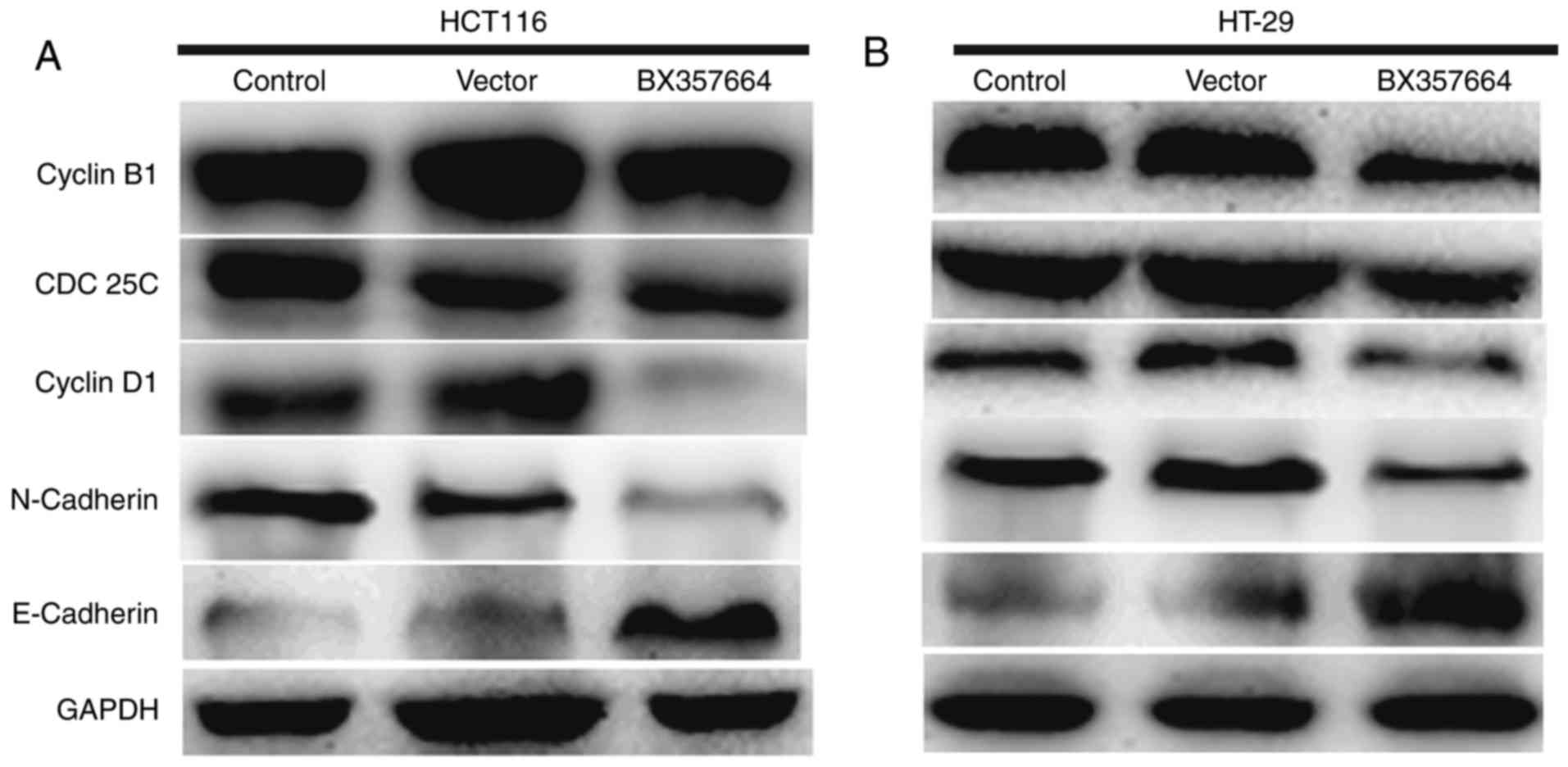
Overexpression of BX357664 alters cell
cycle regulator and EMT marker expression
In view of the above observations, the expression of
key cell cycle regulators and EMT markers was examined in HCT116
and HT-29 cells with or without BX357664 overexpression. In HCT116
cells, overexpression of BX357664 decreased the protein levels of
Cyclin B1, CDC25C and Cyclin D1 (Fig.
4A). In addition, expression of the epithelial marker
E-cadherin was upregulated, while expression of the mesenchymal
marker N-cadherin was inhibited following overexpression of
BX357664 (Fig. 4A). Similar results
were also observed in HT-29 cells (Fig.
4B). These data suggested that overexpression of BX357664
altered the cell cycle progression and EMT processes.
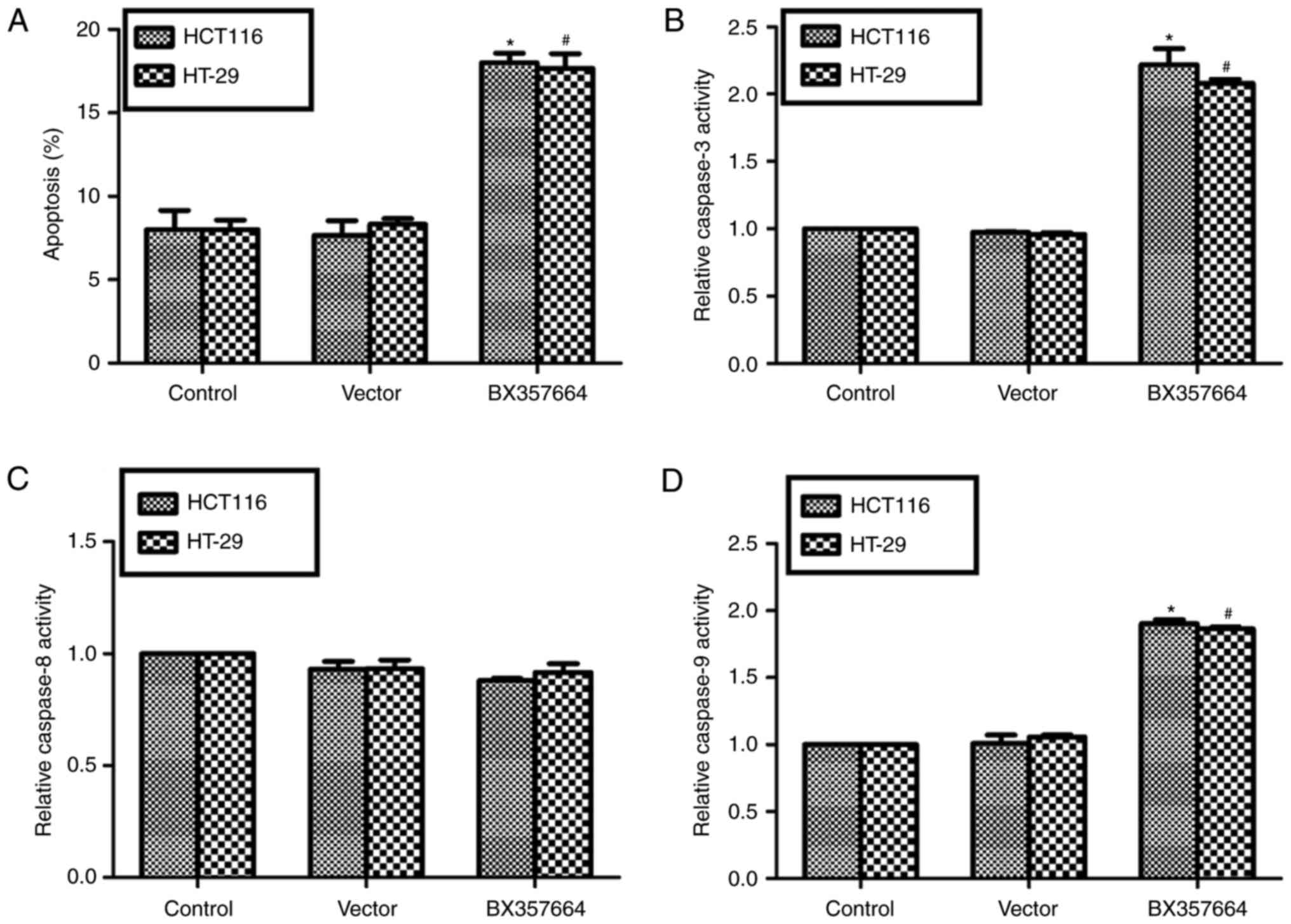
Overexpression of BX357664 promotes
cell apoptosis in HCT116 and HT-29 cells
Since BX357664 overexpression arrested cell cycle at
G0/G1 phase, we further assessed whether cell apoptosis was
regulated by BX357664 overexpression. The results demonstrated that
the cell apoptosis rate was ~2-fold increased following
transfection of BX357664 into HCT116 cells or HT-29 cells, compared
with the control groups (Fig. 5A).
Detection of key apoptosis executors revealed that the activity of
caspase-3 was significantly enhanced following BX357664
overexpression in both CRC cell lines (Fig. 5B). The activity of caspase-8 remained
largely unchanged among the treatment groups (Fig. 5C). The activity of caspase-9 was
significantly enhanced by overexpression of BX357664 in both HCT116
cells and HT-29 cells (Fig. 5D).
These findings suggested that cell apoptosis was significantly
induced by BX357664 overexpression in CRC cells.
Discussion
Colorectal cancer (CRC) represents a great burden
for patients worldwide. It is reported that >1.2 million
patients are diagnosed with CRC annually, of which ~600,000 die
from the disease (5). The cornerstone
of therapy for CRC is surgery; however surgery is effective
predominantly for localized CRC. In patients suffering from
metastatic CRC, only 10–25% is applicable for surgical resection
(13). Despite neoadjuvant
radiotherapy (for patients with rectal cancer) and adjuvant
chemotherapy (for patients with stage III/IV and high-risk stage II
colon cancer), only 30–40% of patients survive 5 years and over
(14). Therefore, new diagnostic and
prognostic tools and a detailed understanding of colorectal
carcinoma development need to be explored as soon as possible
(15).
Accumulating evidence has demonstrated that a body
of lncRNAs exert its tissue specificity in various types of cancer
and serve an important role in human tumorigenesis and cancer
progression (16,17). Emerging lncRNAs have been identified
to associate with tumor progression or suppression of CRC,
including lncRNA UCC (9), lncRNA CHRF
(18) and lncRNA TUSC7 (19). BX357664 was initially identified by
microarray analysis in renal cell carcinoma, and its full-length is
650 nt. BX357664 was predicted to have the least potential of
protein coding by the online Coding Potential Assessment Tool
(20). BX357664 was reported to be
downregulated in renal cell carcinoma and to negatively regulate
the cell proliferation and EMT processes (11). However, the role of BX357664 in other
solid tumors remains largely unknown. In view that the same
molecule may play different roles in different types of cancer, the
role of BX357664 in other tumors merits further exploration. This
detailed information would largely enhance our understanding of
human tumorigenesis and of the functional roles of BX357664 in
human cancers.
The present study examined the role of BX357664 in
human CRC, and it represents the first report about this lncRNA in
human CRC. Similarly to the report demonstrating that BX357664
negatively regulated cell proliferation and EMT in human renal
carcinoma (11), the present study
also demonstrated a significant role of BX357664 on human CRC.
First, the present study examined the downregulation of this lncRNA
in human CRC tissues and cultured cells, which was consistent with
that in renal carcinoma (11).
Second, based on its lower transcript level in CRC, BX357664 was
overexpressed in two CRC cell lines, HCT116 and HT-29, and the
effects on cell proliferation, migration and invasion were
examined. Consistent with the observations in renal carcinoma
(11), overexpression of BX357664 led
to significantly decreased cell proliferation, migration and
invasion capacities in CRC cells.
To further verify the observations in cell
proliferation, migration and invasion, additional cell cycle and
western blot analysis of cell cycle regulators and EMT markers were
performed. Cell cycle arrest is a hallmark of proliferation
inhibition and an indicator of tumor suppression (21). Cyclin B1, CDC25C and Cyclin D1 are key
regulators of the cell cycle. Deregulation of these proteins is
associated with aberrant tumor growth (21). The present western blot analysis
revealed that the protein levels of Cyclin B1, CDC25C and Cyclin D1
were consistently decreased following overexpression of BX357664 in
CRC cells, consistent with the proliferation inhibition mediated by
BX357664. Additionally, EMT, the process where tumor cells obtain
highly mesenchymal properties and lose the epithelial phenotype, is
significantly associated with metastasis (21). N-cadherin is a mesenchymal marker and
E-cadherin is an epithelial marker (22). The present western blot analysis
detected that expression of the mesenchymal marker N-cadherin was
decreased while the epithelial marker E-cadherin was increased
following BX357664 overexpression in CRC cells, consistent with the
migration and invasion inhibition effects mediated by BX357664.
Therefore, the present results strongly suggested that BX357664
exerted inhibition effects on CRC proliferation, migration and
invasion.
Third, cell cycle arrest results in cell apoptosis
in human cancers (23,24). Therefore, the role of BX357664 in cell
apoptosis was further examined. The present results demonstrated
that BX357664 overexpression significantly increased cell apoptosis
rates, which further supported the cell cycle arrest and cell
proliferation inhibition mediated by BX357664 in CRC cells. Taken
together, the present findings were consistent with the previous
study in renal cell carcinoma (11),
and suggest that BX357664 may serve as a tumor suppressor in a wide
range of solid tumors.
The mechanisms by which BX357664 exerts its
functions in CRC remain to be elucidated. The present data
demonstrated that cell apoptosis was induced following
overexpression of BX357664. The activities of caspase-3 and
caspase-9, but not caspase-8, were significantly enhanced by
BX357664 overexpression. These data indicated that BX357664 might
inhibit CRC progression via a caspase-3 and caspase-9-dependent
apoptosis pathway. Since BX357664 is a novel lncRNA whose role in
solid tumors remains largely unknown, and there exists only one
previous study that reported that TGF-β1/p38/HSP27 signaling was
regulated by BX357664 in renal cell carcinoma (11), the molecular mechanisms by which
BX357664 exerts its properties in CRC would merit further
investigation.
In summary, the present study demonstrated that
BX357664 was downregulated in clinical CRC tissues compared with
adjacent normal tissues. Overexpression of BX357664 decreased cell
proliferation, migration and invasion capacities in CRC cells. The
present data suggested that BX357664 might have critical
suppressive effects on human tumorigenesis and serve as a tumor
suppressor. These data provided evidence that overexpression of
BX357664 might be a valuable therapeutic strategy for a wide range
of solid tumors, such as CRC and renal cell carcinoma.
References
|
1
|
Ferlay J, Parkin DM and Steliarova-Foucher
E: Estimates of cancer incidence and mortality in Europe in 2008.
Eur J Cancer. 46:765–781. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ren JS, Masuyer E and Ferlay J:
Global estimates of cancer prevalence for 27 sites in the adult
population in 2008. Int J Cancer. 132:1133–1145. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo P, Huang ZL, Yu P and Li K: Trends in
cancer mortality in China: An update. Ann Oncol. 23:2755–2762.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
de Hoon M, Shin JW and Carninci P:
Paradigm shifts in genomics through the FANTOM projects. Mamm
Genome. 26:391–402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Martens-Uzunova ES, Bottcher R, Croce CM,
Jenster G, Visakorpi T and Calin GA: Long noncoding RNA in
prostate, bladder and kidney cancer. Eur Urol. 65:1140–1151. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen L, Wang W, Cao L, Li Z and Wang X:
Long Non-Coding RNA CCAT1 acts as a competing endogenous RNA to
regulate cell growth and differentiation in acute myeloid leukemia.
Mol Cells. 39:330–336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang FT, Chen WY, Gu ZQ, Zhuang YY, Li
CQ, Wang LY, Peng JF, Zhu Z, Luo X, Li YH, et al: The novel long
intergenic noncoding RNA UCC promotes colorectal cancer progression
by sponging miR-143. Cell Death Dis. 8:e27782017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qin C, Han Z, Qian J, Bao M, Li P, Ju X,
Zhang S, Zhang L, Li S, Cao Q, et al: Expression pattern of long
non-coding RNAs in renal cell carcinoma revealed by microarray.
PLoS One. 9:e993722014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Y, Qian J, Li X, Chen W, Xu A, Zhao K,
Hua Y, Huang Z, Zhang J, Liang C, et al: Long noncoding RNA
BX357664 regulates cell proliferation and epithelial-to-mesenchymal
transition via inhibition of TGF-β1/p38/HSP27 signaling in renal
cell carcinoma. Oncotarget. 7:81410–81422. 2016.PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tomlinson JS, Jarnagin WR, DeMatteo RP,
Fong Y, Kornprat P, Gonen M, Kemeny N, Brennan MF, Blumgart LH and
D'Angelica M: Actual 10-year survival after resection of colorectal
liver metastases defines cure. J Clin Oncol. 25:4575–4580. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Abdalla EK, Vauthey JN, Ellis LM, Ellis V,
Pollock R, Broglio KR, Hess K and Curley SA: Recurrence and
outcomes following hepatic resection, radiofrequency ablation and
combined resection/ablation for colorectal liver metastases. Ann
Surg. 239:818–827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li L and Luo HS: G-Protein signaling
protein-17 (RGS17) is upregulated and promotes tumor growth and
migration in human colorectal carcinoma. Oncol Res. 2017.(Epub
ahead of print).
|
|
16
|
Prensner JR, Iyer MK, Balbin OA,
Dhanasekaran SM, Cao Q, Brenner JC, Laxman B, Asangani IA, Grasso
CS, Kominsky HD, et al: Transcriptome sequencing across a prostate
cancer cohort identifies PCAT-1, an unannotated lincRNA implicated
in disease progression. Nat Biotechnol. 29:742–749. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li H, Ma SQ, Huang J, Chen XP and Zhou HH:
Roles of long noncoding RNAs in colorectal cancer metastasis.
Oncotarget. 8:39859–39876. 2017.PubMed/NCBI
|
|
18
|
Tao Y, Han T, Zhang T, Ma C and Sun C:
LncRNA CHRF-induced miR-489 loss promotes metastasis of colorectal
cancer via TWIST1/EMT signaling pathway. Oncotarget.
30:36410–36422. 2017.
|
|
19
|
Xu J, Zhang R and Zhao J: The novel long
noncoding RNA TUSC7 inhibits proliferation by sponging MiR-211 in
colorectal cancer. Cell Physiol Biochem. 41:635–644. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang L, Park HJ, Dasari S, Wang S, Kocher
JP and Li W: CPAT: Coding-Potential assessment tool using an
alignment-free logistic regression model. Nucleic Acids Res.
41:e742013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sherr CJ: Cancer cell cycles. Science.
274:1672–1677. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: Emt: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu XY, Xia P, Yu M, Nie XC, Yang X, Xing
YN, Liu YP, Takano Y and Zheng HC: The roles of REIC gene and its
encoding product in gastric carcinoma. Cell Cycle. 11:1414–1431.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du Y, Gong J, Tian X, Yan X, Guo T, Huang
M, Zhang B, Hu X, Liu H, Wang Y, et al: Japonicone a inhibits the
growth of non-small cell lung cancer cells via
mitochondria-mediated pathways. Tumour Biol. 36:7473–7482. 2015.
View Article : Google Scholar : PubMed/NCBI
|