Introduction
Prostate Cancer (PCa) is the second most frequently
diagnosed cancer among men and the fifth leading cause of cancer
mortalities worldwide (1). Mortality
rates for PCa have been decreasing in the majority of more
developed countries, but continue to rise in certain Asian
countries, particularly in China (1).
The primary diagnostic tools used to obtain evidence of PCa include
digital rectal examination, serum concentration of PSA and
transrectal ultrasound guided biopsies (2). Diagnosis depends on the presence of
adenocarcinoma in operative specimens and prostate biopsy cores
(2). The factors driving the increase
of incidence in prostate cancer are not entirely understood;
however, they may include gradual implementation of
prostate-specific antigen screening and improved biopsy techniques,
or the effect of an increasingly westernized lifestyle, in
particular dietary habits (1).
Despite recent hormonal therapies exhibiting a survival advantage
during treatment with inhibitors, including abiraterone acetate
(17α-hydroxylase inhibitor) and enzalutamide (androgen receptor
inhibitor), the median duration of response is <1 year (2). Furthermore, the treatment options for
recurrent and metastatic PCa are limited and predominantly
palliative (2).
Epithelial-mesenchymal transition (EMT) is an early
embryonic development program, in which cells convert from the
epithelial to the mesenchymal state (3). During this process, the epithelial cells
acquire mesenchymal cell morphology through the suppressing the
epithelial markers and inducing mesenchymal markers (4). It has been shown to have a critical role
during prostate cancer progression and metastasis (5). Another important signaling pathway is
Wingless (WNT)/β-catenin (6) and
activation of Wnt/β-catenin signaling induces motility,
invasiveness, cell-fate decisions and the maintenance of
self-renewal potential (7). Recent
studies indicate that, remarkably, Wnt/β-catenin signaling is
essential for the biological process of bone metastasis in various
types of cancers, particularly in PCa (8).
Oldhamianoside II, atriterpenoidsaponin, may be
isolated from Gypsophila oldhamiana, a plant that is rich in
saponins (9). A previous study
demonstrated that oldhamianoside II was able to effectively inhibit
the proliferation and migration of tumor cells, as well as tumor
angiogenesis (10). These results
indicated the feasibility of oldhamianoside II for further clinical
application. A previous study demonstrated that oldhamianoside II
exerted antitumor activity via inhibition of C-X-C motif chemokine
receptor 4, matrix metalloproteinase 2, phosphoinositide
3-kinase/protein kinase B and extracellular signal-related
kinase/mitogen activated protein kinase (10); however, the specific mechanisms
underlying the antitumor effects of oldhamianoside II in PCa remain
unclear. The present study aimed to further elucidate these
mechanisms, and revealed that oldhamianoside II inhibited the
proliferation and invasion of PCa cells by regulating the process
of epithelial-mesenchymal transition (EMT) and the activity of the
Wnt/β-catenin signaling pathway.
Materials and methods
Cell culture and treatment
VCaP and PC3 human PCa cell lines were purchased
from American Type Culture Collection (Manassas, VA, USA) and
maintained in Dulbecco's modified Eagle's medium (HyClone; GE
Healthcare, Chicago, IL, USA) or RPMI-1640 (HyClone; GE Healthcare)
supplemented with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). For the experiments, cells
were cultured at 37°C and stimulated with oldhamianoside II, which
was extracted and isolated as previously described (9). Cycloheximide (50 µg/ml, CHX;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) or MG132 (30 µM,
Sigma-Aldrich; Merck KGaA) were applied to stimulate PCa cells for
6h before oldhamianoside II treatment.
MTT assay
An MTT assay was performed to detect to the cell
viability as previously described (11). Briefly, 1×104 cells per
well in 200 µl medium were seeded into a 96-well plate and then
subjected to the indicated treatments for 24–72 h at 37°C. For
detection, 20 µl MTT was added to each well and incubated for 4 h.
Subsequently, the cultured medium was discarded and 150 µl dimethyl
sulfoxide was added. The absorbance was then determined at 490 nm,
and all experiments were performed in triplicate.
Migration assay
For the migration assay, 1×104 cells per
well in serum-free medium were plated into 24-well Transwell
chambers with polycarbonate filter inserts (pore size, 8.0–10.0 µm;
Corning Incorporated, Corning, NY, USA). The lower chambers
contained normal growth medium with 10% fetal bovine serum.
Following incubation at 37°C for 24 h, the cells in the upper
compartment were removed and the migrated cells were fixed and
stained using the crystal violet method. Cells were quantified in
five randomly selected microscopic fields using light microscope
(magnification, ×200) and all experiments were performed in
triplicate.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted using an RNeasy Mini kit
(Qiagen), and RNA (1 µg) was reverse-transcribed to cDNA using a
SuperScript II cDNA synthesis kit (Invitrogen). Subsequently, 1 µg
of cDNA product was used for amplification with the PCR
amplification kit (Takara) as previously described (12). The details of the primers for each
human gene were as follows: E-cadherin forward,
5′-TCATGAGTGTCCCCCGGTAT-3′, and reverse,
5′-TCTTGAAGCGATTGCCCCAT-3′; N-cadherin forward,
5′-CGCCATCCGCTCCACTT-3′, and reverse, 5′-GCTGATGACAAATAGCGGGC-3′;
vimentin forward, 5′-GGACCAGCTAACCAACGACA-3′, and reverse,
5′-AAGGTCAAGACGTGCCAGAG-3′; cyclin D1 (CCND1) forward,
5′-CACACGGACTACAGGGGAGT-3′, and reverse,
5′-GATGGTTTCCACTTCGCAGC-3′; c-JUN forward,
5′-GTGCCGAAAAAGGAAGCTGG-3′, and reverse,
5′-CTGCGTTAGCATGAGTTGGC-3′; axin 2 (AXIN2) forward,
5′-AAACGCAATGGGAAAGGCAC-3′, and reverse,
5′-TGTGCTTTGGGCACTATGGG-3′; Dickkopf-1 (DKK1) forward,
5′-AAGTGTGGTGGCTTCCAAGG-3′, and reverse,
5′-CCGGCCACATGAGTAAGAGG-3′; protein phosphatase 2 catalytic subunit
β (PPP2CB) forward, 5′-ACCATGCCAATGGTCTCACA-3′, and reverse,
5′-AACATGAGGCTCACCACGAC-3′; secreted frizzled-related protein 5
(SFRP5) forward, 5′-CACTCGGATACGCAGGTCTT-3′, and reverse,
5′-CACTCGGATACGCAGGTCTT-3′; and GAPDH (internal loading control)
forward, 5′-AATGGGCAGCCGTTAGGAAA-3′, and reverse,
5′-GCGCCCAATACGACCAAATC-3′. The reaction was performed at 94°C for
30 sec, 56°C for 30 sec and 72°C for 30 sec, for 36 cycles. GAPDH
served as the loading control. Analysis of relative gene expression
data using real-time quantitative PCR and the 2−ΔΔCq
method (13). All reactions were
performed in triplicate.
Western blot analysis
The nuclear proteins were extracted using Nuclear
and Cytoplasmic Extraction Reagents kits (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Western blotting was performed as
previously described (14). The
primary antibodies against E-cadherin (1:1,000; cat no. 3195),
N-cadherin (1:1,000; cat no. 13116), β-catenin (1:1,000; cat no.
8480) and Vimentin (1:1,000; cat no. 5741) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA) and the antibodies
against cyclin D1 (1:1,000; cat no. ab134175), c-Jun (1:1,000; cat
no. ab32137) and Lamin A (1:1,000; cat no. ab108922) were purchased
from Abcam (Cambridge, UK). For the loading control,
HRP-conjugated-GAPDH antibodies (1:1,000; cat no. sc-293335; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) were used. The primary
antibodies were incubated at 4°C overnight, and the second antibody
labeled with horseradish-peroxidase was subsequently incubated at
37°C for 1 h. SuperSignal West Pico kit (Thermo Fisher Scientific,
Inc.) was used to visualize the protein bands.
Statistical analysis
Statistical analyses of the data were performed
using SPSS version 16 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard deviation and were analyzed by
one-way analysis of variance followed by Tukey's post hoc test,
considering oldhamianoside II treatment and cell viability at each
time point, or vehicle and oldhamianoside II treatment at different
concentrations. P<0.05 was considered to indicate a
statistically significant difference.
Results
Oldhamianoside II inhibits the
proliferation and invasion of PCa cell lines
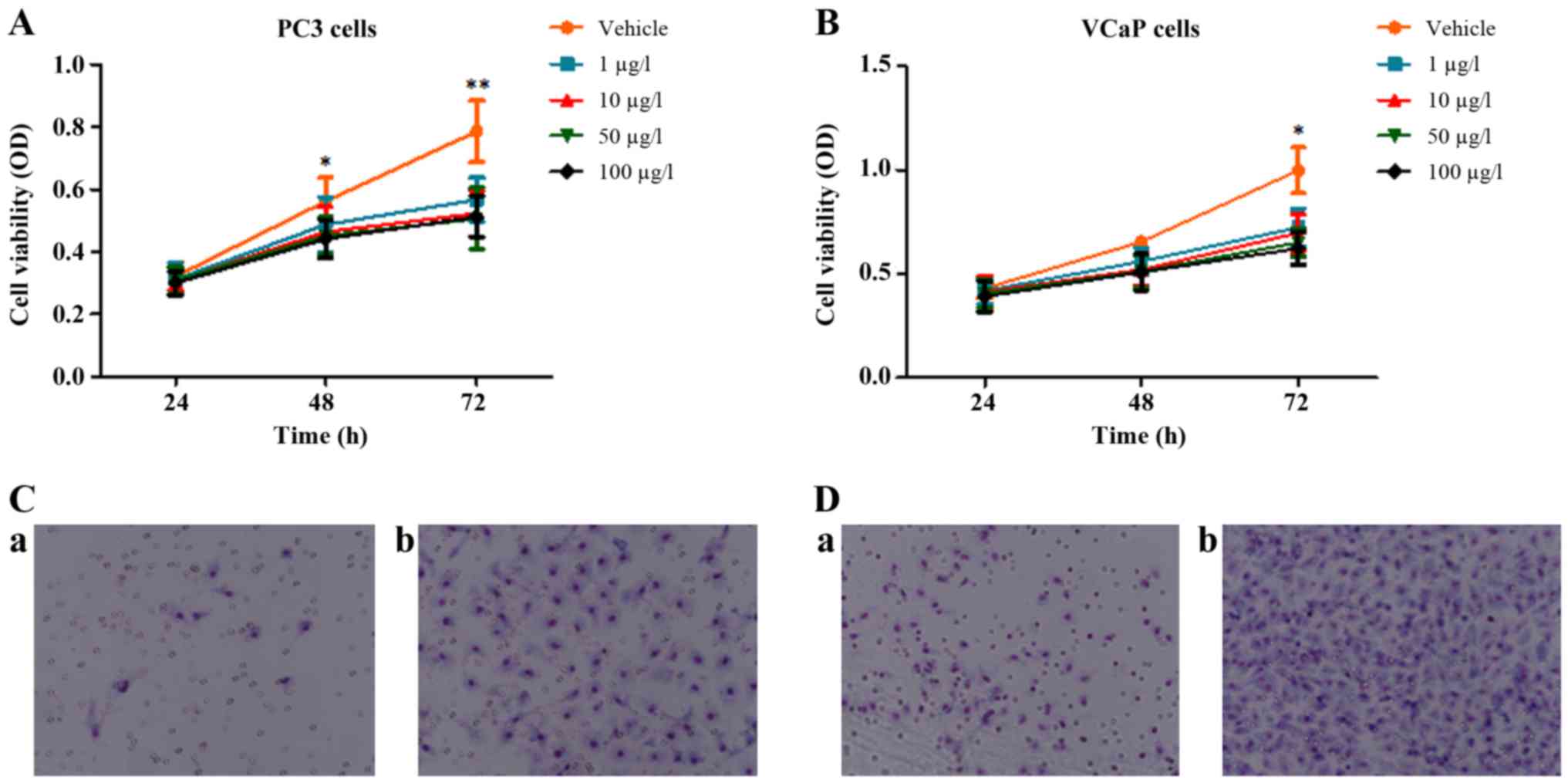
VCaP and PC3 human PCa cell lines were treated to
detect the effects of oldhamianoside II on the biological activity
of tumor cells. Consistently with a previous study (10), the MTT analysis demonstrated that
oldhamianoside II treatment at various concentrations decreased the
viability of PC3 and VCaP cells (Fig. 1A
and B). In addition, a Transwell migration assay demonstrated
that 50 µg/l oldhamianoside II significantly inhibited the
migration ability of PC3 (Fig. 1C)
and VCaP cells (Fig. 1D).
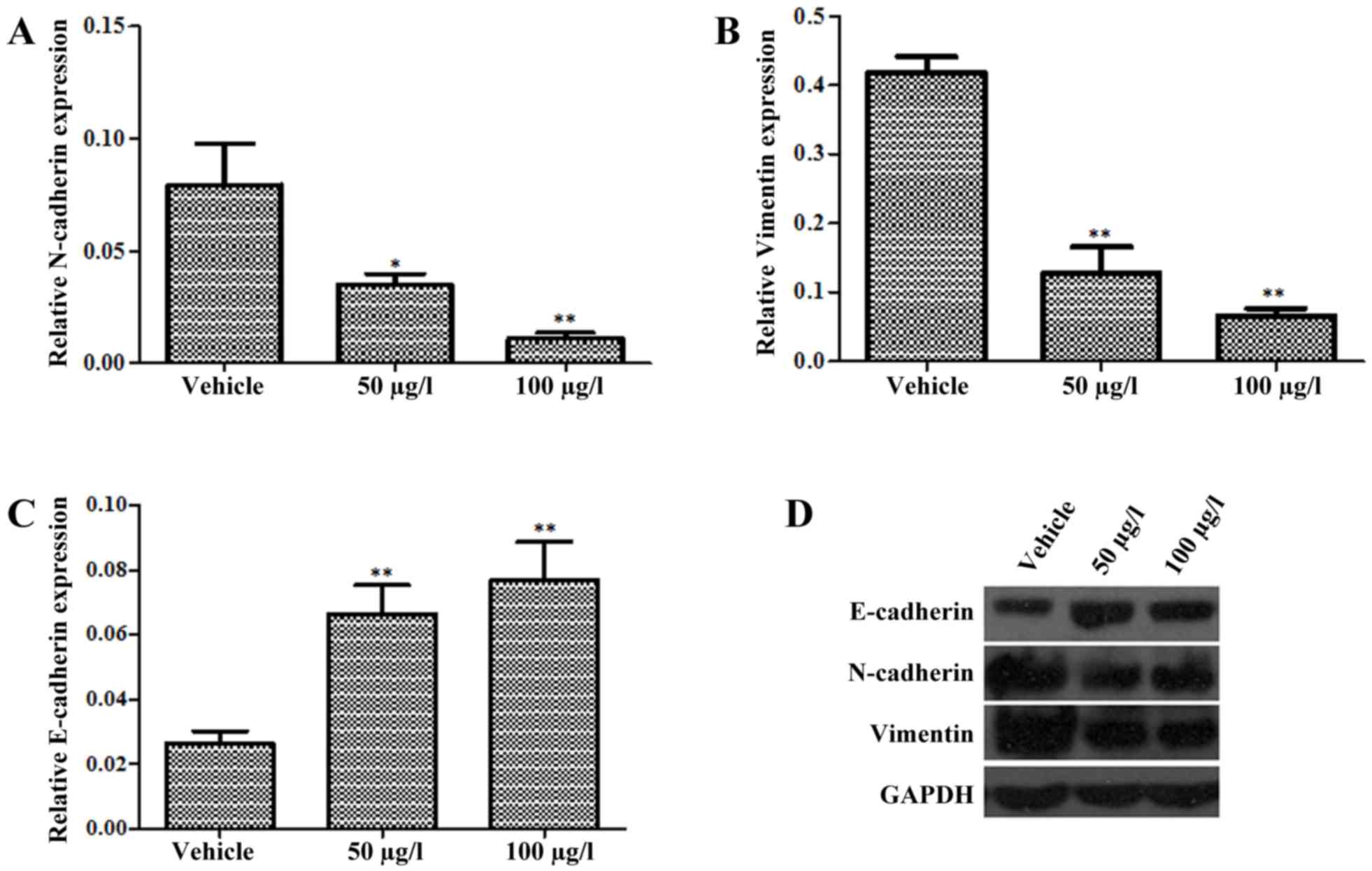
Oldhamianoside II inhibits EMT in VCaP
cells
EMT is associated with the metastasis and
progression of PCa (15). As
presented in Fig. 2A, treatment with
50 and 100 µg/l oldhamianoside II in VCaP cells for 72 h suppressed
EMT, as indicated by the suppression of N-cadherin (Fig. 2A) and Vimentin (Fig. 2B) and the induction of E-cadherin
(Fig. 2C) at the mRNA level.
Furthermore, following the treatment of VCaP cells with
oldhamianoside II at 50 and 100 µg/l for 48 h, western blot
analysis demonstrated that the level of E-cadherin protein was
significantly elevated, whereas the levels of Vimentin and
N-cadherin were decreased significantly (Fig. 2D).
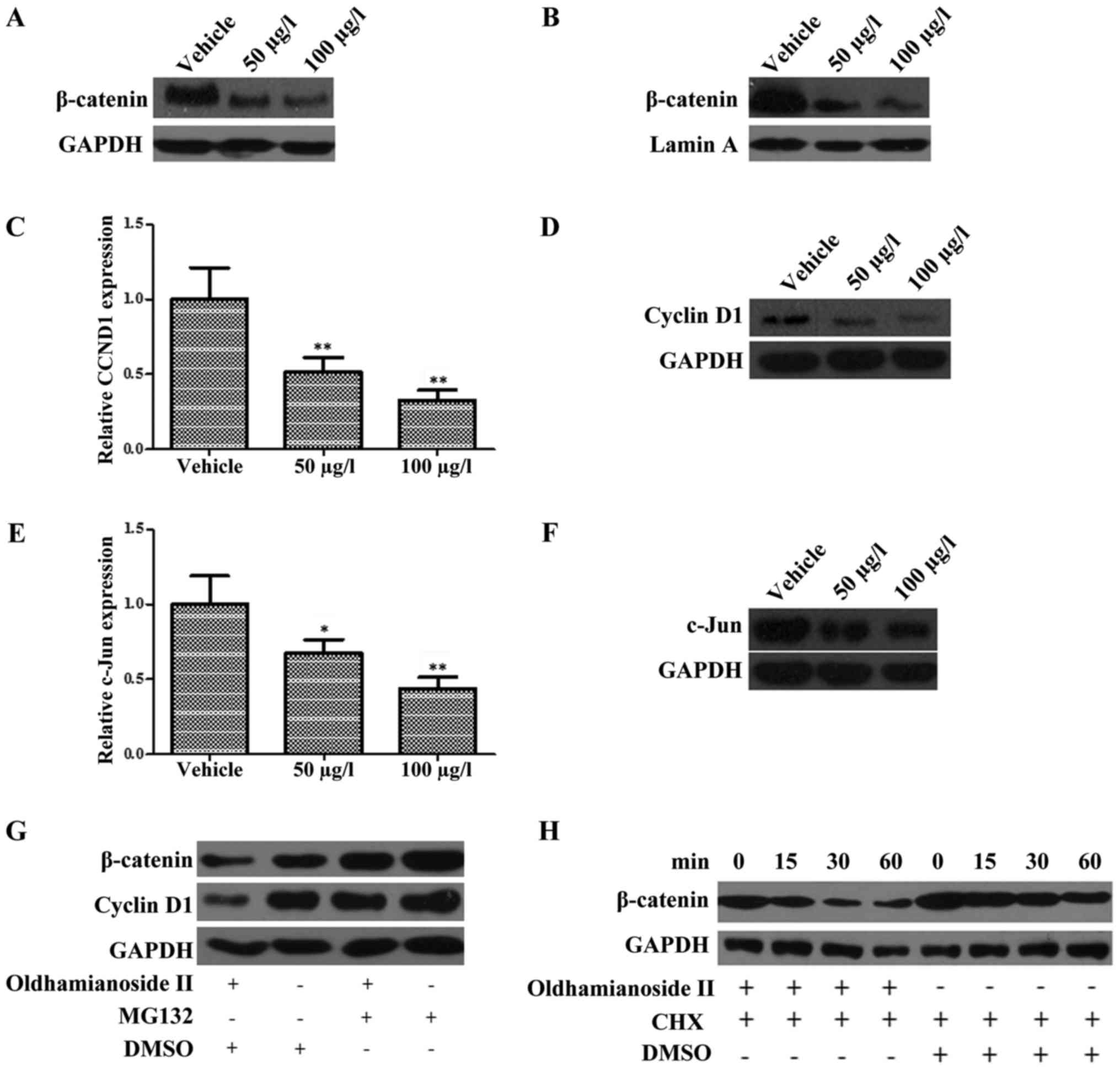
Oldhamianoside II inhibits the
expression and activity of β-catenin in PCa cells
Based on the aforementioned results, the present
study investigated whether oldhamianoside II was able to regulate
the activity of the Wnt/β-catenin signaling pathway in PCa cells.
Therefore, VCaP cells were treated with oldhamianoside II and the
level of β-catenin was determined by western blotting. The total
β-catenin protein level (Fig. 3A) and
its nuclear content (Fig. 3B) were
significantly suppressed in VCaP cells following oldhamianoside II
treatment at 50 and 100 µg/l. Subsequently, the expression levels
of the target genes of β-catenin, CCND1 and c-JUN were also
evaluated in response to oldhamianoside II treatment (16,17). As
shown in Fig. 3C-F, oldhamianoside II
treatment induced a significant decrease in the expression levels
of these target genes at the mRNA (Fig.
3C and E) and protein (Fig. 3D and
F) levels.
As β-catenin is well-validated to be targeted for
ubiquitin-mediated proteolysis (18),
we subsequently investigated whether oldhamianoside II was able to
regulate the proteolysis of β-catenin. As presented in Fig. 3G, the effect of oldhamianoside II on
β-catenin and its target cyclin D1 was blocked in the presence of
the proteasome inhibitor MG132, which indicated that oldhamianoside
II promoted proteasome-mediated degradation of β-catenin.
Consistently, oldhamianoside II treatment resulted in a significant
decrease in the half-life of β-catenin (Fig. 3H).
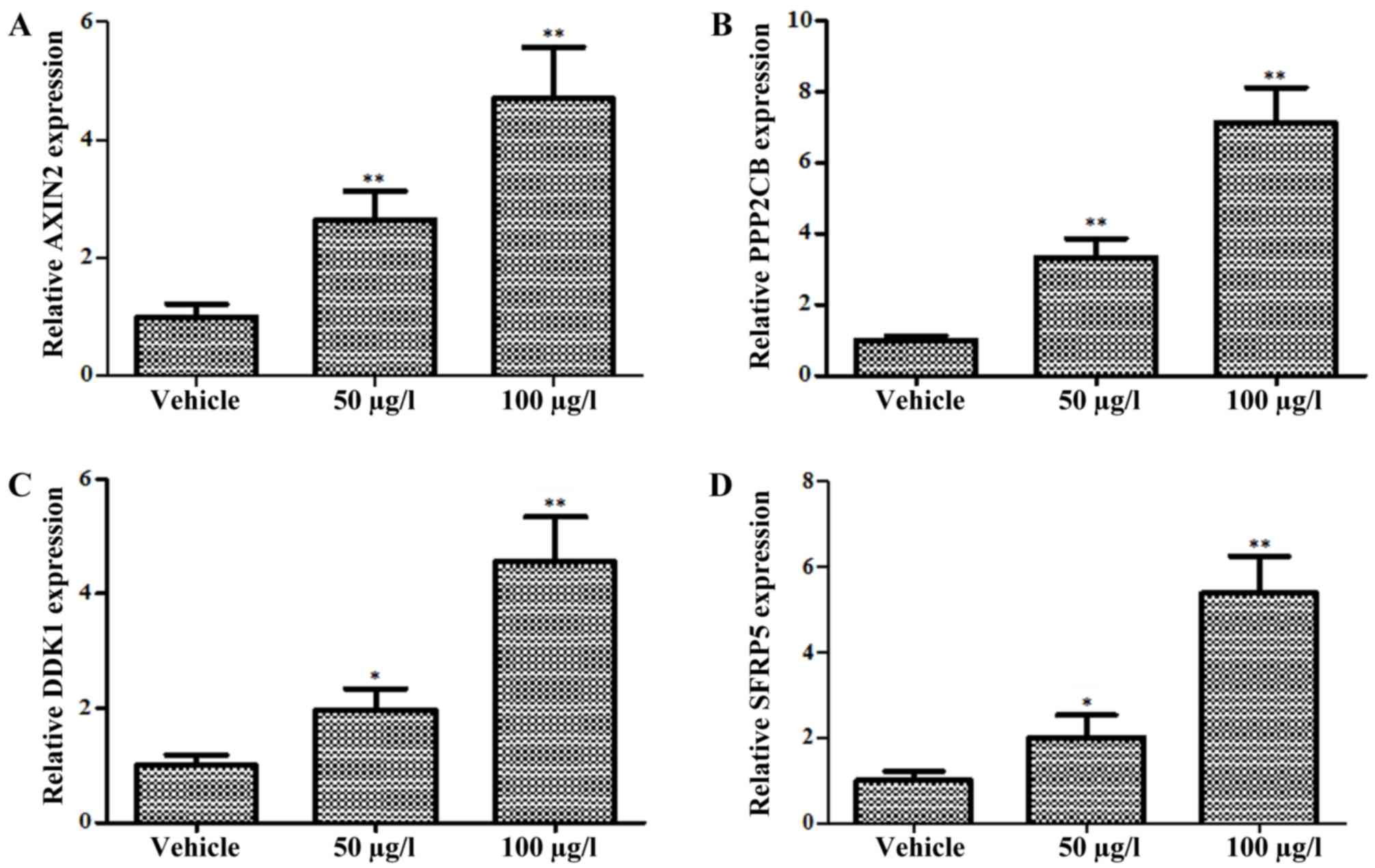
Oldhamianoside II promotes β-catenin
degradation by inducing expression of Wnt antagonists
A previous study demonstrated that antagonists of
the Wnt signaling pathway promoted β-catenin degradation by
interfering with the binding of Wnt proteins to their receptors or
organizing the destruction complex (19). To further confirm the regulation of
oldhamianoside II on the Wnt/β-catenin signaling pathway, the
expression level of natural inhibitors towards Wnt signaling were
detected following oldhamianoside II treatment. Four
well-characterized antagonists (AXIN2, PPP2CB, DKK1 and SFRP5)
targeting Wnt/β-catenin signaling (20) were selected for investigation, and the
results revealed that AXIN2 (Fig.
4A), PPP2CB (Fig. 4B), DKK1
(Fig. 4C) and SFRP5 (Fig. 4D) were upregulated significantly in
oldhamianoside II-treated cells.
Discussion
Oldhamianoside II, a novel triterpenoidsaponin, was
first chemically isolated from Gypsophila oldhamiana,
commonly called ‘Xia Cao’ in China (9); it contains a triterpenoidaglycone and
eight sugar chains. A previous study demonstrated that
oldhamianoside II may inhibit tumor growth and angiogenesis, and
that the migration of tumor cells was also suppressed following
oldhamianoside II treatment (10).
Consistent with previous evidence, the present study demonstrated
that oldhamianoside II inhibited the proliferation and invasion of
PCa cells.
EMT progression is characterized by cytoskeletal
reorganization, followed by the suppression of cell adhesion,
finally leading to enhanced cell motility. Numerous studies have
validated that EMT is closely associated with tumor progression and
metastasis (21). In the present
study, by characterizing the expression level of epithelial and
mesenchymal markers, it was demonstrated that oldhamianoside II
inhibited EMT in PCa cells. Together, the aforementioned results
corroborate the inhibitory role of oldhamianoside II during PCa
progression.
Aberrantly activated Wnt/β-catenin signaling may
induce tumor formation and progression (22). β-catenin is a transcription factor
that is involved in the Wnt signaling pathway and serves an
important role in oncogenesis in combination with T-cell factor
(Tcf) and protein kinase D1 (23,24). It
has previously been reported that β-catenin was dysregulated in and
associated with the development of numerous types of cancer,
including PCa (25), and that the
shuttle between the cytoplasm and nucleus was pivotal for its pro-
or antitumor functions (26).
Therefore, a study focusing on the regulation of β-catenin would
highlight the pathological mechanism underlying PCa and also
provide novel treatment targets. The results of the present study
revealed that oldhamianoside II decreased the expression level of
β-catenin and its downstream target genes, CCND1 and c-Jun,
resulting in the suppression of PCa growth. Notably, oldhamianoside
II was able to promote the proteasome-mediated degradation of
β-catenin. These results further supported the potential
application of oldhamianoside II in the treatment of PCa.
Wnt/β-catenin signaling is frequently activated due
to the suppression of its antagonists (27). To further investigate the mechanism
underlying the effects of oldhamianoside II on Wnt/β-catenin
signaling, the present study also detected the expression levels of
Wnt antagonists, including AXIN2, PPP22B, DKK1 and SFRP5 (28). Oldhamianoside II treatment was found
to induce the expression of these genes. A previous study
demonstrated that antagonists of the Wnt signaling pathway
inhibited its activity in various ways; among these, DKK1 inhibited
the interaction between the receptor and its ligand (29), while AXIN2 and PPP2CB could form a
destruction complex to degrade β-catenin (30). Together with the present study, these
findings suggest that oldhamianoside II may inhibit PCa progression
via targeting β-catenin through various mechanisms.
In conclusion, the present study demonstrated a
novel molecular mechanism, involving the reversal of EMT and
inhibition of β-catenin signaling, through which the anticancer
effects of oldhamianoside II are mediated. Oldhamianoside II may
act as a chemopreventive agent to inhibit or delay the progression
of PCa by promoting β-catenin degradation and inhibiting its
transcriptional activity. The potential clinical application of
oldhamianoside II in PCa treatment requires further study.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81572544 and 81772760), The
Key Research and Development Project of Shandong (grant no.
2016GSF201166), Scientific research fund of Jinan University
(XKY1522), Supporting Fund For Teacher's research of Jining Medical
University (JY2017FS001), The Shandong Taishan Scholarship (grant
no. tsqn20161076) and The Innovation Project of Shandong Academy of
Medical Sciences.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SL and LW contributed to study conception and
design. KL, XZ, JZ, TW, HD, FJ, DL, YC and YL contributed to the
acquisition of data. KL, XZ, SL and LW contributed to the analysis
and interpretation of data.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
All authors approved the final version to be
published.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heidenreich A, Aus G, Bolla M, Joniau S,
Matveev VB, Schmid HP and Zattoni F: European Association of
Urology: EAU guidelines on prostate cancer. Eur Urol. 53:68–80.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:ra82014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kong D, Banerjee S, Ahmad A, Li Y, Wang Z,
Sethi S and Sarkar FH: Epithelial to mesenchymal transition is
mechanistically linked with stem cell signatures in prostate cancer
cells. PLoS One. 5:e124452010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nauseef JT and Henry MD:
Epithelial-to-mesenchymal transition in prostate cancer: Paradigm
or puzzle? Nat Rev Urol. 8:428–439. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Giles RH, van Es JH and Clevers H: Caught
up in a Wnt storm: Wnt signaling in cancer. Biochim Biophys Acta.
1653:1–24. 2003.PubMed/NCBI
|
|
7
|
Rabbani SA, Arakelian A and Farookhi R:
LRP5 knockdown: Effect on prostate cancer invasion growth and
skeletal metastasis in vitro and in vivo. Cancer Med. 2:625–635.
2013.PubMed/NCBI
|
|
8
|
Dai J, Hall CL, Escara-Wilke J, Mizokami
A, Keller JM and Keller ET: Prostate cancer induces bone metastasis
through Wnt-induced bone morphogenetic protein-dependent and
independent mechanisms. Cancer Res. 68:5785–5794. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun JY, Zhong Y, Yin JT, Liu L, Wang B and
Zuo CX: Isolation and structure identification of triterpenoids
from Gypsophila oldhamiana. Chin Trad Herbal Drugs.
36:644–646. 2005.
|
|
10
|
Wang FL, Sun JY, Wang Y, Mu YL, Liang YJ,
Chong ZZ, Qin SH and Yao QQ: Oldhamianoside II, a new
triterpenoidsaponin, prevents tumor growth via inducing cell
apoptosis and inhibiting angiogenesis. Oncol Res. 20:369–376. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang L, Zhang J, Yang X, Chang YW, Qi M,
Zhou Z, Zhang J and Han B: SOX4 is associated with poor prognosis
in prostate cancer and promotes epithelial-mesenchymal transition
in vitro. Prostate Cancer Prostatic Dis. 16:301–307. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Zheng Y, Xu H, Yan X and Chang X:
Investigate pathogenic mechanism of TXNDC5 in rheumatoid arthritis.
PLoS One. 8:e533012013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Li Y, Yang X, Yuan H, Li X, Qi M,
Chang YW, Wang C, Fu W, Yang M, et al: ERG-SOX4 interaction
promotes epithelial-mesenchymal transition in prostate cancer
cells. Prostate. 74:647–658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smith BN and Odero-Marah VA: The role of
Snail in prostate cancer. Cell Adh Migr. 6:433–441. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:29–26. 2009. View Article : Google Scholar
|
|
17
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fischer JA: Deubiquitinating enzymes:
Their roles in development, differentiation, and disease. Int Rev
Cytol. 229:43–72. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arend RC, Londoño-Joshi AI, Straughn JM Jr
and Buchsbaum DJ: The Wnt/β-catenin pathway in ovarian cancer: A
review. Gynecol Oncol. 131:772–779. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Koch A, Waha A, Hartmann W, Hrychyk A,
Schüller U, Waha A, Wharton KA Jr, Fuchs SY, von Schweinitz D and
Pietsch T: Elevated expression of Wnt antagonists is a common event
in hepatoblastomas. Clin Cancer Res. 11:4295–4304. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bullions LC and Levine AJ: The role of
beta-cateninin cell adhesion, signal transduction, and cancer. Curr
Opin Oncol. 10:81–87. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mulholland DJ, Dedhar S, Coetzee GA and
Nelson CC: Interaction of nuclear receptors with the
Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know? Endocr
Rev. 26:898–915. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sundram V, Chauhan SC, Ebeling M and Jaggi
M: Curcumin attenuates β-catenin signaling in prostate cancer cells
through activation of protein kinase D1. PLoS One. 7:e353682012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yokoyama NN, Shao S, Hoang BH, Mercola D
and Zi X: Wnt signaling in castration-resistant prostate cancer:
Implications for therapy. Am J Clin Exp Urol. 15:27–44. 2014.
|
|
26
|
Park JH, Kwon HY, Sohn EJ, Kim KA, Kim B,
Jeong SJ, Song JH, Koo JS and Kim SH: Inhibition of Wnt/β-catenin
signaling mediates ursolic acid-induced apoptosis in PC-3 prostate
cancer cells. Pharmacol Rep. 65:1366–1374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Robinson DR, Zylstra CR and Williams BO:
Wnt signaling and prostate cancer. Curr Drug Targets. 9:571–580.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kypta RM and Waxman J: Wnt/β-catenin
signaling in prostate cancer. Nat Rev Urol. 9:418–428. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kagey MH and He X: Rationale for targeting
the Wnt signalling modulator Dickkopf-1 for oncology. Br J
Pharmacol. 174:4637–4650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng AS, Lau SS, Chen Y, Kondo Y, Li MS,
Feng H, Ching AK, Cheung KF, Wong HK, Tong JH, et al: EZH2-mediated
concordant repression of Wnt antagonists promotes
β-catenin-dependent hepatocarcinogenesis. Cancer Res. 71:4028–4039.
2011. View Article : Google Scholar : PubMed/NCBI
|