Introduction
CD24 is a glycosyl-phosphatidylinositol-anchored
protein with a mucin-type structure that resides exclusively in
membrane microdomains. CD24 is often highly expressed in human
solid tumors (including lung, brain, gastric, pancreatic, colon,
prostate, breast and ovarian cancers, cholangiocarcinoma,
medulloblastoma and cutaneous malignant melanoma) and is associated
with poor prognosis (1–6). Experimental studies performed over the
last decade suggest a role of CD24 in tumor growth and processes
associated with metastatic spread, including adhesion, migration
and invasion (7–9). Downregulation of the expression of CD24
in vitro (using short-hairpin interfering RNA) and in
vivo (using monoclonal antibodies) was demonstrated to decrease
the viability of ovarian cancer cells and induce cell apoptosis
(10,11). In a study by Sagiv et al
(9), knockdown of CD24 expression
with the use of monoclonal antibodies or small interfering (si)RNA
was demonstrated to inhibit growth and invasion of colorectal and
pancreatic cancer cells. Suyama et al (12) also reported that CD24 gene transfer or
inhibition of signal transducer and activator of transcription may
represent a novel therapeutic strategy against refractory breast
cancer. CD24+ cells were also demonstrated to exhibit enhanced
chemoresistance as compared to CD24− subpopulations
(2,13). Furthermore, addition of SWA11
monoclonal antibody to inhibit CD24 expression strongly sensitized
A549 lung cancer cells to gemcitabine (14).
However, the underlying mechanisms via which the
expression of CD24 is regulated and the role of CD24 in tumor
progression and chemoresistance have remained to be fully
elucidated. None of the known CD24-associated proteins fully define
the role of CD24 in chemotherapy. In previous work by our group, a
series of potential CD24-interacting proteins was identified by
using high-throughput mass spectrometry (data unpublished). These
included glucose-regulated protein 78 (GRP78), which belongs to the
family of heat shock proteins and is also referred to as
immunoglobulin (Ig) heavy-chain binding protein (BiP). Elucidation
of the association between CD24 and GRP78 may help define the role
of CD24 in tumor progression and possibly the development of a
novel approach to treat colorectal cancer (CRC).
GRP78 is a major endoplasmic reticulum (ER)
chaperone with Ca2+ binding ability; it also serves as
an ER stress signaling regulator (15). Upregulation of GRP78 expression in
cancer cells has been reported in several cancer types, including
those of the urinary, digestive, mammary, cerebral and respiratory
systems, and osteosarcoma (16–19). The
presence of GRP78 autoantibodies in the sera of cancer patients is
generally associated with poor prognosis, as GRP78 promotes tumor
cell proliferation, survival and metastasis by suppressing ER
stress-induced apoptosis (20–25).
Overexpression of GRP78 was also reported to correlate with
chemoresistance. In recent studies, GRP78 was demonstrated to
confer resistance against doxorubicin, bortezomib (26), VP-16 (27), temozolomide (28), paclitaxel and cisplatin (29). Furthermore, treatment with GRP78
inhibitor or knockdown of GRP78 was reported to potentiate
chemotherapy-induced apoptosis in most of the cancer types
mentioned above. However, the mechanisms by which GRP78 promotes
malignant attributes of cancer cells have remained largely elusive.
Of note, GRP78 and CD24 have a similar role in tumor progression
and chemoresistance (30,31). A previous study by our group
identified GRP78 as a CD24-interacting protein, which suggests a
possible link between the mechanism of action of GRP78 and CD24.
Understanding the interaction between GRP78 and CD24 may lead to
the further elucidation of the mechanisms of tumor progression.
Materials and methods
Cell lines
The HT29, HT8, SW480 and colo205 human CRC cell
lines were obtained from the American Type Culture Collection
(Manassas, VA, USA) and cultured in RPMI-1640 medium containing 10%
fetal calf serum (Bio Whittaker, Verviers, Belgium), 2 mM
glutamine, 100 IU/ml penicillin and 100 µg/ml streptomycin
(Cellgro; Mediatech Inc., Manassas, VA, USA) at 37°C in a
humidified atmosphere containing 5% CO2.
Chemicals and antibodies
Oxaliplatin (L-OHP) and the GRP78 inhibitor
vomitoxin (VT) were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). L-OHP was stored as a 20 mM solution in
dimethyl sulfoxide (DMSO) at −20°C, and was diluted with RPMI 1640
medium prior to use with the highest concentration of DMSO in
culture being 0.05% (v/v). The rabbit polyclonal anti-GRP78/Bip
antibody (cat no. ab21685) was purchased from Abcam Ltd (Cambridge,
MA, USA). Anti-CD24-SWA11 monoclonal antibody used for
immunoprecipitation (IP) was obtained from Professor Hans-Peter
Altevogt (Department of Translational Immunology, German Cancer
Research Center, Heidelberg, Germany), in whose lab the antibody
was generated (32). Anti-CD24
monoclonal antibody (cat. no. SC-7034) used for western blotting
and IP, β-actin antibody (cat. no. SC130301) and horseradish
peroxidase-conjugated goat anti-rabbit IgG (cat. no. SC2004) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA).
Plasmids and small interfering
(si)RNA
Full-length human CD24 was amplified by polymerase
chain reaction (PCR). The primers for CD24 were as follows:
Forward, 5′-GTTGTTGGATCCATGGGCAGAGCAATGGT-3′ and reverse,
5′-GTTGTTCTCGAGcgAGAGTAGAGATGCAGAAGAGAG-3′. The pcDNA4-CD24-myc
plasmid (33) was constructed by
inserting this PCR product into the pcDNA4 Myc-His B vector
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
verified by sequencing. GRP78 siRNA (siGENOME SMART pool GRP78;
cat. no. M-008198-01) and control siRNA (siGENOME Non-Targeting
siRNA Pool #1; cat. no. D-001206-13-20) were obtained from
Dharmacon (Lafayette, CO, USA). The sequences of the CD24 siRNA and
control siRNA were 5′-UCUCUCUUCUGCAUCUUUAdTdT-3′ and
5′-CUACCUAUGCAGAUUUAUUdTdT-3′, respectively. Transfection of siRNAs
was performed using the Lipofectamine® 2000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocols.
Cell viability assay
Cell viability was determined using a CellTiter-Blue
assay kit (Promega Corp., Madison, WI, USA), according to the
manufacturer's instructions. In brief, cells were seeded in 96-well
plates at a density of 4×103 cells/well and allowed to
attach for 24 h followed by exposure to L-OHP at different
concentrations (1, 2.5, 5 and 10 µM) for 12, 24 or 48 h.
Subsequently, 20 µl CellTiter-Blue reagent was added into each well
and the cells were incubated for another 2 h at 37°C. The
fluorescence was then recorded [540Ex/600Em]
using a fluorescence plate reader (BioTek Synergy 2; BioTek,
Winooski, VT, USA). Assays were performed in triplicate for each
experiment and the mean cell viability was normalized to that in
the vehicle (DMSO)-treated controls. Each experimental data-point
was generated from at least three independent experiments.
Colony forming assay
500 HT29 cells were seeded in each hole of 6-hole
plate. HT29 cells were treated with or without GRP78 inhibitor VT
(1 µg/ml) for 2 h, followed by incubation with different
concentrations of L-OHP or vehicle (DMSO) for another 24 h. The
medium was refreshed and the cells were incubated at 37°C in an
atmosphere with 95% humidity containing 5% CO2 for 14
days. The cells were stained with crystal violet for 30 mins at
room temperature and the number of colonies was counted. At least
three independent experiments were performed for each treatment.
The results were expressed as a percentage of colony growth
relative to that in the control group.
Co-IP and western blot analysis
Total protein lysates of HT29 cells were obtained by
incubation on ice for 30 min in IP/lysis buffer [50 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, pH 7.5, 150 mM
NaCl, 5 mM EDTA, 1% Triton X-100, 1 mM phenylmethyl
sufonylfluoride, 2 µg/ml pepstatin A and 1X protease inhibitor
cocktail (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)], followed
by centrifugation at 15,000 × g for 20 min at 4°C. Subsequently, 25
µl protein G beads (Zymed; Thermo Fisher Scientific, Inc.) were
added to preclear the supernatant at 4°C for 1 h, followed by
centrifugation at 1000 × g at 4°C for 10 min. The supernatant was
stored and protein concentration was assessed. The samples were
then incubated with 5–10 µg of the indicated antibody or control
mouse IgG (Pierce; Thermo Fisher Scientific, Inc.) for 1 h at 4°C
and then with 30 µl protein G beads for 2 h at 4°C. After 4 washes
in IP/lysis buffer, the beads were boiled in 40 µl of 2X Laemmli
buffer for 5 min and the eluted proteins (30 µg/lane) were
subjected to 12.5% SDS-PAGE. After the separation, the proteins
were transferred onto polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The membrane was blocked in
5% non-fat milk in 1% Tris-buffered saline containing 0.1% Tween-20
(TBST)_for 1 h at room temperature, and then probed separately with
primary antibodies (1:500) overnight at 4°C. Subsequently, the
membranes were washed thrice for 10 min with TBST and incubated
with the appropriate secondary antibody (cat. no. SC2004) for 1 h
at room temperature. After 3 washes, the immunostained proteins
were detected by chemiluminescence (Pierce™ ECL Western
Blotting Substrate, cat. no. 32106; Thermo Fisher Scientific,
Inc.).
Statistical analysis
GraphPad Prism 5 software (GraphPad Software, Inc.,
La Jolla, CA, USA) was used for data processing. For the colony
formation assay, data were analyzed with Microsoft®
Excel (version 14.0.7180.5002, Microsoft Corp., Redmond, WA, USA)
and plotted in Prism 5. Values are expressed as the mean ± standard
deviation or standard error as indicated. Differences between
groups were evaluated using one-way analysis of variance. Multiple
comparisons between the groups were performed using the
Student-Neuman-Keuls test. P<0.05 was considered to indicate a
statistically significant difference.
Results
L-OHP decreases CD24 expression in CRC
cells
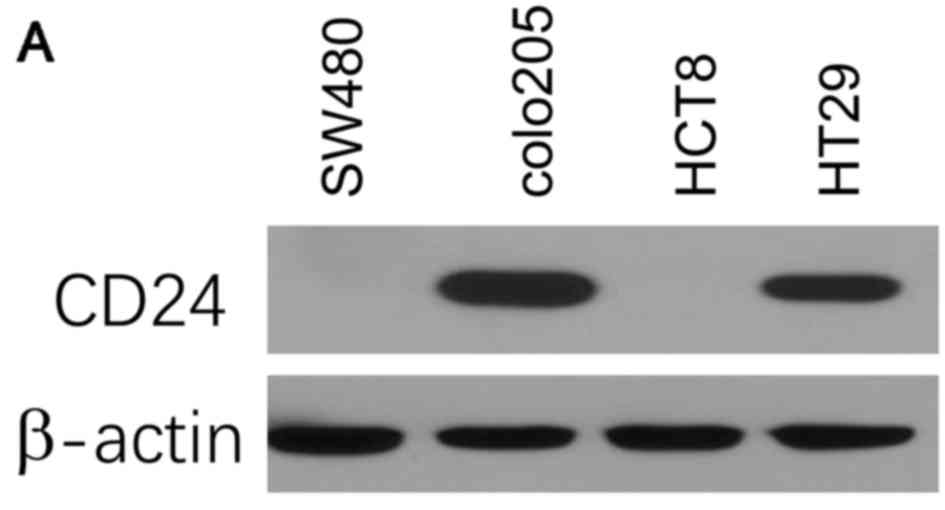
First, the expression of CD24 was assessed in a
panel of CRC cell lines, including SW480, colo205, HCT8 and HT29,
using immunoblotting analysis. The results indicated that the CRC
cell lines expressed varying levels of CD24. The HT29 and colo205
cell lines had high expression levels of CD24, while the expression
was undetectably low in SW480 and HCT8 cells (Fig. 1A).
To assess the effect of L-OHP on CD24 expression,
HT29 cells were treated with vehicle or L-OHP at various
concentrations for 24 h. The expression of CD24 was determined
using western blot analysis. As presented in Fig. 1B, the protein levels of CD24 in HT29
cells treated with L-OHP at 1, 2.5 and 5 µM were decreased to 63,
51 and 21%, respectively, compared with those in the DMSO vehicle
control. These results suggested that L-OHP decreased the
expression of CD24 in a dose-dependent manner. An L-OHP dose of 5
µM was chosen for further experiments.
CD24 antagonizes L-OHP-induced
cytotoxicity
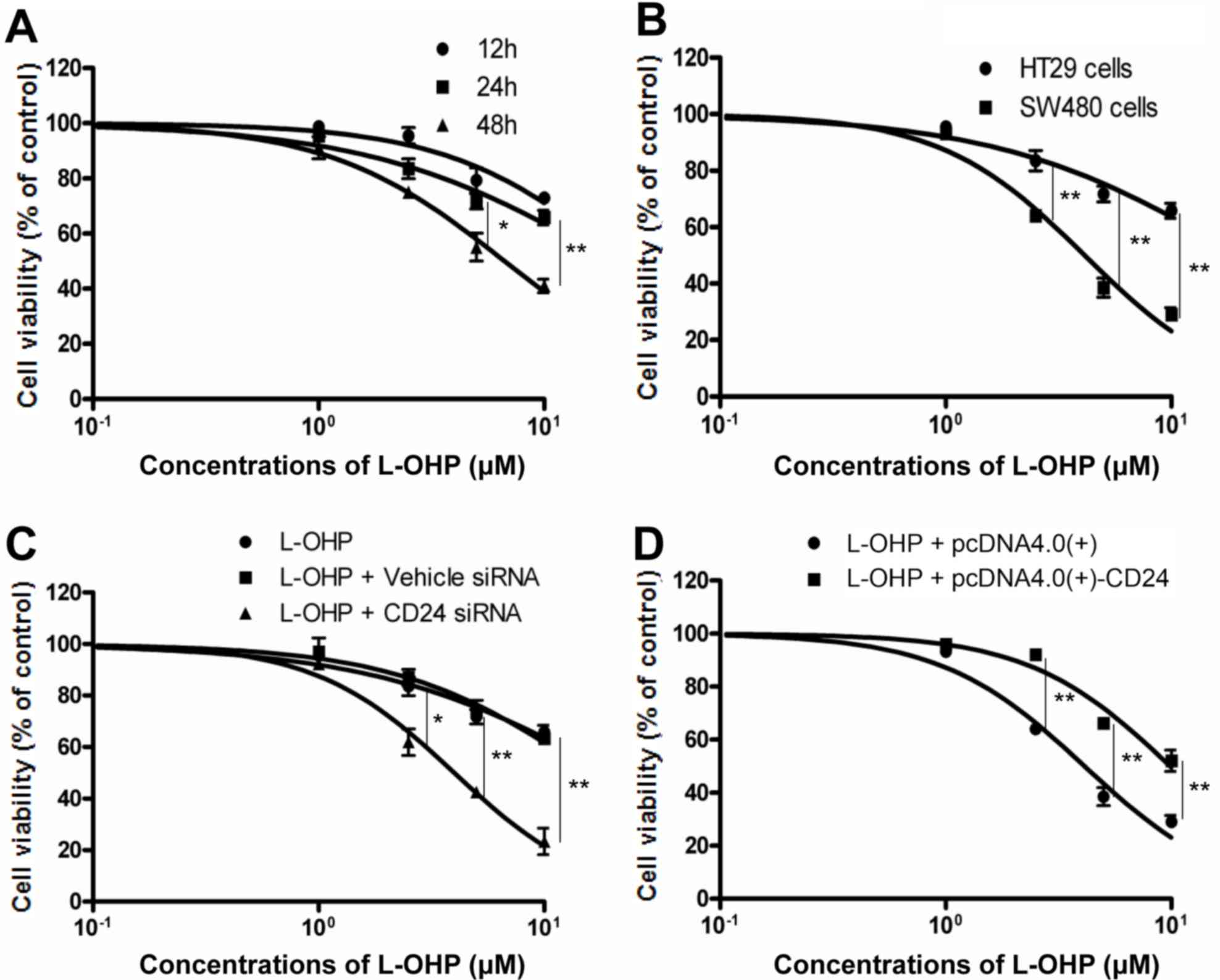
HT29 CRC cells were treated with different
concentrations of L-OHP (1, 2.5, 5 or 10 µM) for 12, 24 or 48 h. As
presented in Fig. 2A, the
concentration leading to a 50% reduction in the cell number
(IC50) of L-OHP at 12, 24 and 48 h was determined as
22.35±1.295, 19.89±0.815 and 6.683±0.0363 µM, respectively. The
cytotoxic effect of L-OHP on HT29 cells with high expression of
CD24 and SW480 cells with low expression of CD24 was then compared.
The IC50 was 19.89±0.815 and 4.13±0.245 µM, respectively
(Fig. 2A and B). To determine the
role of CD24 in L-OHP-induced cytotoxicity, CD24 expression in HT29
cells was knocked down using specific targeted siRNA. HT29 cells
were transfected with vehicle siRNA or CD24 siRNA, and then
incubated with different concentrations of L-OHP for 24 h. As
presented in Fig. 2C, CD24 siRNA
decreased the IC50 of L-OHP from 19.89±0.815 to
3.998±0.042 µM. Vehicle siRNA had almost no effect on the
cytotoxicity of L-OHP. The effect of ectopic CD24 expression on the
cytotoxic effect of L-OHP was then examined in SW480 cells, which
were transiently transfected with a CD24 expression plasmid,
pcDNA4-CD24-myc plasmid, or a vehicle control plasmid. SW480 cells
were observed to develop resistance to L-OHP after transfection
with CD24 expression plasmid; the IC50 increased from
4.13±0.25 to 9.90±0.43 µM (Fig.
2D).
Effect of L-OHP on GRP78 and CD24
To elucidate the underlying regulatory mechanisms
associated with CD24, it was attempted to identify proteins
associated with IT, as to date, none of the known CD24-associated
proteins has provided any insight into its role in the response or
resistance to chemotherapy, to the best of our knowledge. In
previous work by our group, a series of CD24-associated proteins
was identified (data unpublished), and further research focused on
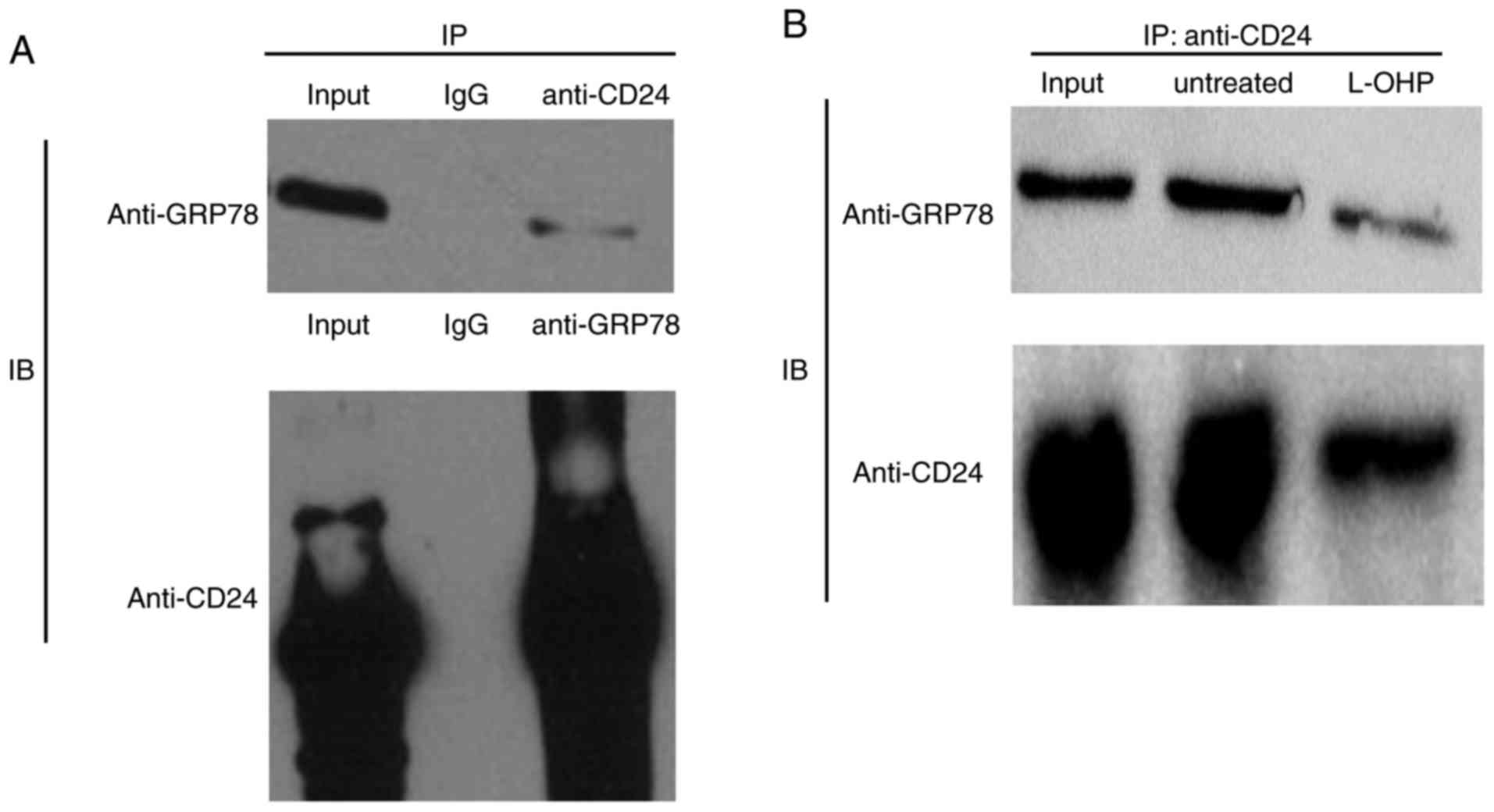
the proteins involved in chemotherapy. Co-IP was performed to
further verify the physical association between GRP78 and CD24
indicated in the previous study. As presented in Fig. 3A, the co-IP assay demonstrated that
endogenous CD24 associated with endogenous GRP78 in HT29 cells. To
investigate the effect of L-OHP on the interaction between GRP78
and CD24, HT29 cells were treated with 5 µM L-OHP and the
interaction was assessed using a co-IP assay. The results
demonstrated that GRP78 physically associated with CD24 in HT29
cells and that L-OHP suppressed the expression of GRP78 and CD24
(Fig. 3B).
Suppression of GRP78 decreases the
expression of CD24 and enhances L-OHP-induced cytotoxicity
To assess whether altered levels of GRP78 affect
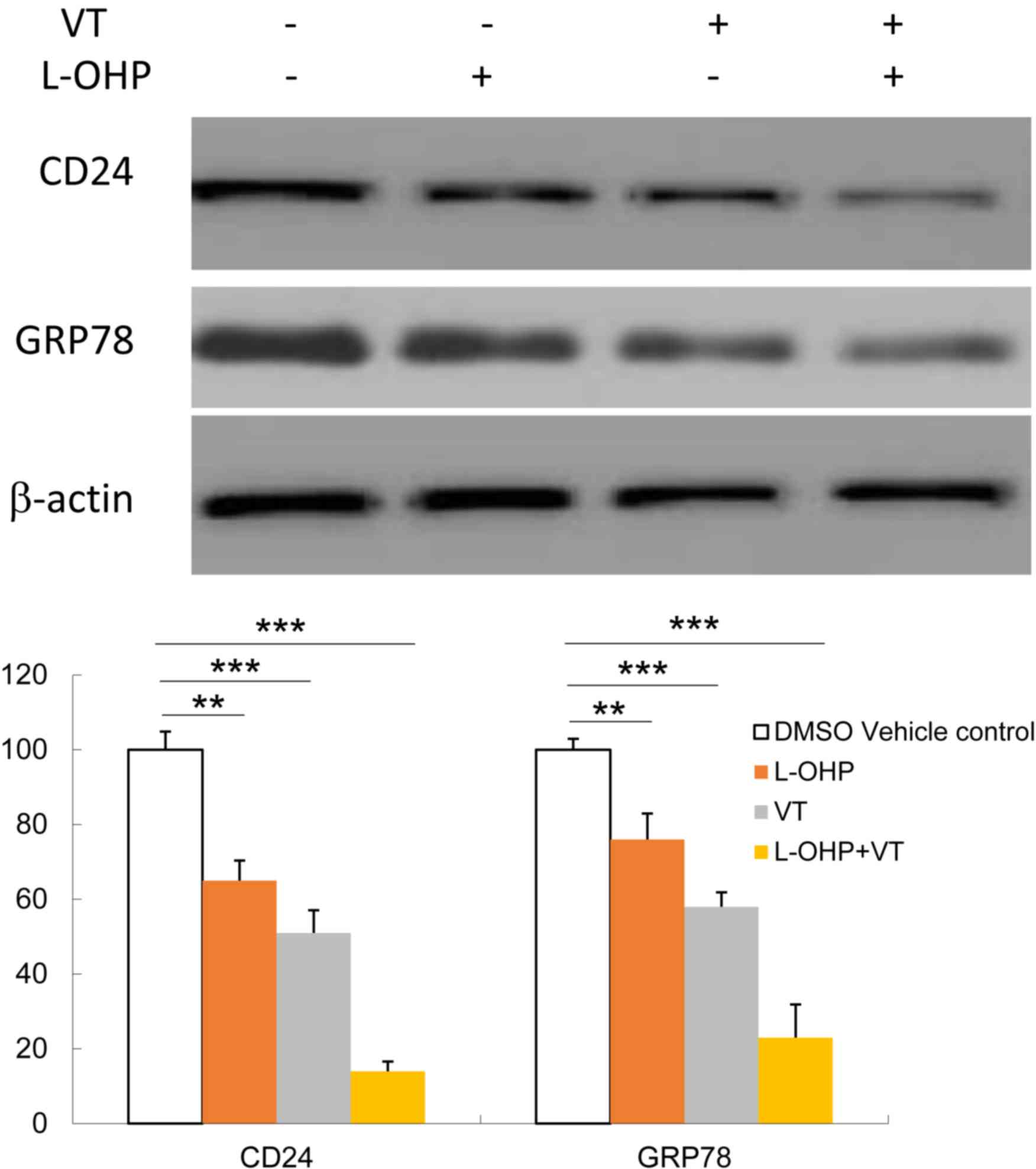
CD24 expression, First, HT29 cells were treated with GRP78
inhibitor VT at 1 µg/ml for 2 h, then treated with L-OHP at 5 µM
for 24 h. Then CD24 and GRP78 expression were determined by western
blot analysis. As presented in Fig.
4, VT caused a significant downregulation of CD24 expression in
HT29 cells with or without L-OHP treatment. L-OHP significantly
decreased the expression of CD24, and that of GRP78 to a slightly
lesser extent. Furthermore, inhibition of GRP78 by VT potentiated
the inhibition of CD24 by L-OHP.
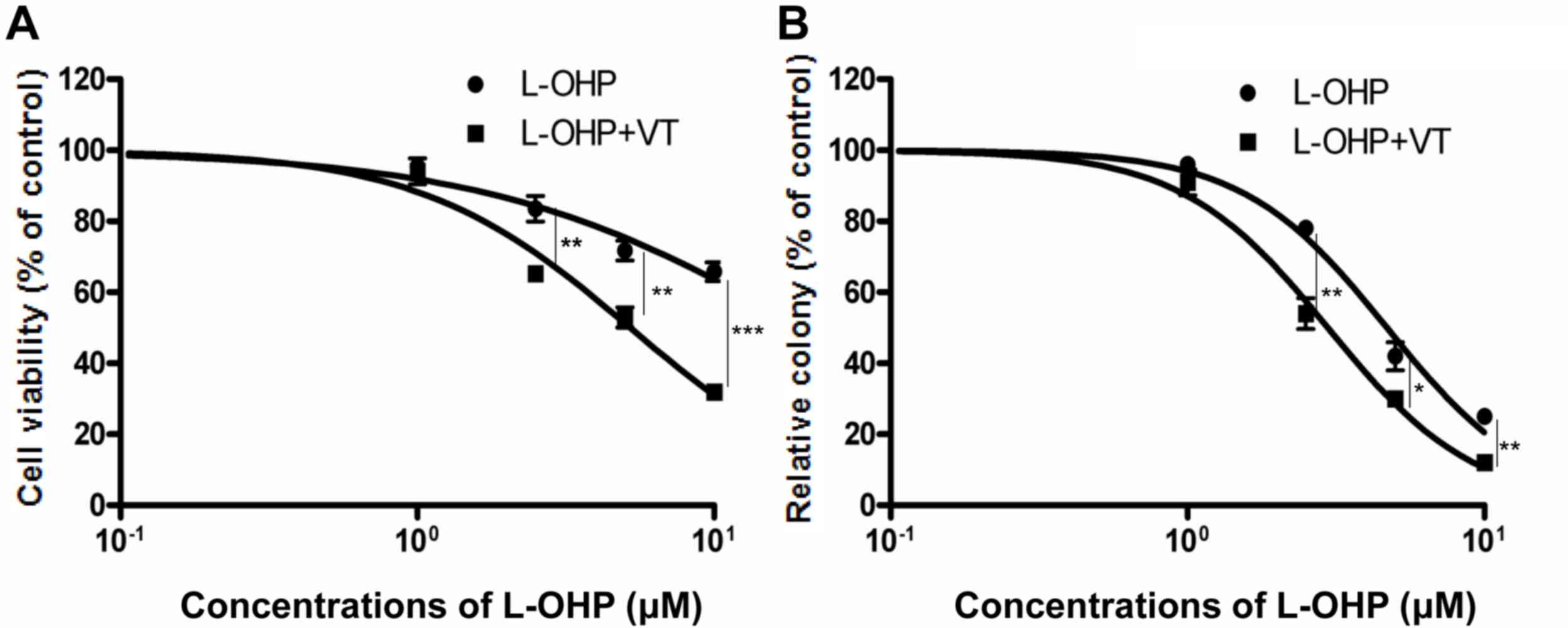
The cell viability assay and colony forming assay
confirmed the indication that suppression of GRP78 sensitized HT29
cells to L-OHP-induced cytotoxicity. As presented in Fig. 5A, suppression of GRP78 using VT
resulted in a significant decrease in the amount of viable cells;
the IC50 of L-OHP alone and in combination with VT was
19.89±0.82 and 5.216±0.63 µM, respectively. In the colony forming
assay, after exposure to 1 µg/ml VT for 2 h, HT29 cells were
treated with various concentrations of L-OHP for 24 h and then
incubated for another 10–14 days, followed by determination of the
number of colonies. The IC50 of L-OHP alone and in
combination with VT was 4.695±1.02 and 2.97±0.96 µM, respectively
(Fig. 5B). The IC50 values
obtained in the colony forming assay were lower than those of the
cytotoxicity assay, which indicated that certain cells were
inhibited, losing their ability to proliferate, and form colonies.
The colony assay determines the cologenicity of cells, which act as
seed cells to form metastasis or recurrent tumors, while the cell
viability assay may reflect the acute effect of the drug on the
primary tumor. The results of the colony forming assay are likely
to be more representative of the scenario of an in vivo
experiment than the cell viability assay.
In summary, the results of the present study
indicated that CD24 antagonized L-OHP-induced cytotoxicity and that
GRP78 was involved in this process. In addition, the potential role
of GRP78 in the regulation of the sensitivity of CRC cells to
L-OHP-induced cytotoxicity was examined. It was indicated that
GRP78 physically interacts with CD24 and that downregulation of
GPR78 using GPR78 synthesis inhibitor sensitized CRC cells to
L-OHP-induced cytotoxicity, at least in part by inhibition of
CD24.
Discussion
CRC is the fourth most common cause of
cancer-associated mortality after lung, gastric and liver cancer.
L-OHP is widely used as the first-line chemotherapeutic agent in
patients with CRC. However, resistance of human CRC cells to the
available chemotherapeutic agents is considered a major obstacle to
successful treatment (34).
Therefore, a better understanding of the molecular mechanisms
underlying L-OHP resistance may be helpful in developing more
effective treatments for CRC.
The present study demonstrated that the expression
of CD24 contributes to the resistance of CRC cells against
L-OHP-induced cytotoxicity (2,13). The
HT29 cell line with high expression of CD24 and the SW480 cell line
with low expression of CD24 were selected to explore the role of
CD24 in modulating the cytotoxic effect of L-OHP on CRC cells. The
SW480 cells were more sensitive to L-OHP than HT29 cells.
Furthermore, knockdown of CD24 using siRNA sensitized the HT29
cells to L-OHP and ectopic expression of CD24 made SW480 cells
resistant to L-OHP. These results indicate that CD24 expression
antagonizes the cytotoxic effect of L-OHP. Decades of research have
demonstrated that CD24 promotes cancer cell proliferation and
migration; furthermore, overexpression of CD24 is associated with
enhanced chemoresistance and poor prognosis (1–6).
Therefore, CD24 has been described as a novel predictor of
responsiveness to chemotherapy and an adverse prognostic marker in
the context of several malignancies (3). The present study confirmed that CD24
overexpression antagonizes L-OHP-induced cytotoxicity; this may be
a possible mechanism for the acquisition of L-OHP resistance by CRC
cells. Identifying the underlying regulatory mechanisms of CD24
expression will help to improve the effectiveness of L-OHP.
A previous study by our group identified a series of
potential CD24-interacting proteins, which included GRP78. The
present study revealed that GRP78 was coordinately expressed with
CD24 expression. In HT29 cells, GRP78 and CD24 were highly
expressed, and suppression of GRP78 using a pharmacological
inhibitor resulted in a significant decrease in CD24 expression.
The co-IP assay indicated that GRP78 physically associates with
CD24. Therefore, it was deduced that GRP78 may directly promote
CD24 expression. Co-IP assay revealed that L-OHP treatment
decreased the expression of GRP78 and CD24, which may be due to
suppression of the interaction between GRP78 and CD24 by L-OHP.
Follow-up experiments are required to confirm this hypothesis.
Treatment of HT29 cells with L-OHP induced a significant
downregulation of CD24 expression, while GRP78 expression was also
downregulated. Therefore, L-OHP maybe reduce the interaction
between GRP78 and CD24. In addition, combined treatment with L-OHP
and GRP78 inhibitor decreased the expression of CD24 to a great
extent compared with separate drug treatments. However, these
results are preliminary, and follow-up experiments are required for
verification. It was hypothesized that inhibition of GRP78 by VT
sensitizes HT29 cells to L-OHP, which may result in a higher
cytotoxic effect. The cytotoxicity assay indicated that GRP78
inhibitor sensitized HT29 cells to L-OHP, which confirmed the
present hypothesis. However, we have only completed some
preliminary experiments, follow-up experiments still need to do to
verify the results in the future.
In conclusion, the present results indicate that
CD24 antagonizes L-OHP-induced cytotoxicity and that upregulation
of CD24 may be a possible resistance mechanism in CRC cells. GRP78
was indicated to promote CD24 expression and may be a promising
target of anti-tumor therapy. Sensitization of CRC cells to
L-OHP-induced cytotoxicity by inhibition of GRP78 was closely
associated with down-regulation of CD24. Thus, combination of drugs
capable of suppressing GRP78 may enhance the effectiveness of L-OHP
in the treatment of CRC and may help improve the prognosis of
patients with CRC.
Acknowledgements
The authors would like to thank Professor Hans-Peter
Altevogt (Department of Translational Immunology, German Cancer
Research Centre, Heidelberg, Germany) for providing anti-CD24-SWA11
monoclonal antibody. The antibody was generated in this lab
(32).
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81572938).
Availability of data and materials
The analyzed data sets generated during the study ar
the corresponding author on reasonable request.
Authors' contributions
XW conceived and designed the experiments. JX, YC,
SH and FC performed the experiments. SH analyzed the data.
Ethical approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bretz N, Noske A, Keller S, Erbe-Hofmann
N, Schlange T, Salnikov AV, Moldenhauer G, Kristiansen G and
Altevogt P: CD24 promotes tumor cell invasion by suppressing tissue
factor pathway inhibitor-2 (TFPI-2) in a c-Src-dependent fashion.
Clin Exp Metastasis. 29:27–38. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Keeratichamroen S, Leelawat K, Thongtawee
T, Narong S, Aegem U, Tujinda S, Praditphol N and Tohtong R:
Expression of CD24 in cholangiocarcinoma cells is associated with
disease progression and reduced patient survival. Int J Oncol.
39:873–881. 2011.PubMed/NCBI
|
|
3
|
Agrawal S, Kuvshinoff BW, Khoury T, Yu J,
Javle MM, LeVea C, Groth J, Coignet LJ and Gibbs JF: CD24
expression is an independent prognostic marker in
cholangiocarcinoma. J Gastrointest Surg. 11:445–451. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kwon MJ, Han J, Seo JH, Song K, Jeong HM,
Choi JS, Kim YJ, Lee SH, Choi YL and Shin YK: CD24 overexpression
is associated with poor prognosis in luminal A and triple-negative
breast cancer. PLoS One. 10:e1391122015. View Article : Google Scholar
|
|
5
|
Tang MR, Wang YX, Guo S, Han SY, Li HH and
Jin SF: CD24 expression predicts poor prognosis for patients with
cutaneous malignant melanoma. Int J Clin Exp Med. 7:4337–4341.
2014.PubMed/NCBI
|
|
6
|
Wu JX, Zhao YY, Wu X and An HX:
Clinicopathological and prognostic significance of CD24
overexpression in patients with gastric cancer: A meta-analysis.
PLoS One. 9:e1147462014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tanaka T, Terai Y, Kogata Y, Ashihara K,
Maeda K, Fujiwara S, Yoo S, Tanaka Y, Tsunetoh S, Sasaki H, et al:
CD24 expression as a marker for predicting clinical outcome and
invasive activity in uterine cervical cancer. Oncol Rep.
34:2282–2288. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Overdevest JB, Thomas S, Kristiansen G,
Hansel DE, Smith SC and Theodorescu D: CD24 offers a therapeutic
target for control of bladder cancer metastasis based on a
requirement for lung colonization. Cancer Res. 71:3802–3811. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sagiv E, Starr A, Rozovski U, Khosravi R,
Altevogt P, Wang T and Arber N: Targeting CD24 for treatment of
colorectal and pancreatic cancer by monoclonal antibodies or small
interfering RNA. Cancer Res. 68:2803–2812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Smith SC, Oxford G, Wu Z, Nitz MD, Conaway
M, Frierson HF, Hampton G and Theodorescu D: The
metastasis-associated gene CD24 is regulated by Ral GTPase and is a
mediator of cell proliferation and survival in human cancer. Cancer
Res. 66:1917–1922. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su D, Deng H, Zhao X, Zhang X, Chen L,
Chen X, Li Z, Bai Y, Wang Y, Zhong Q, et al: Targeting CD24 for
treatment of ovarian cancer by short hairpin RNA. Cytotherapy.
11:642–652. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suyama K, Onishi H, Imaizumi A, Shinkai K,
Umebayashi M, Kubo M, Mizuuchi Y, Oda Y, Tanaka M, Nakamura M and
Katano M: CD24 suppresses malignant phenotype by downregulation of
SHH transcription through STAT1 inhibition in breast cancer cells.
Cancer Lett. 374:44–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ke J, Wu X, Wu X, He X, Lian L, Zou Y, He
X, Wang H, Luo Y, Wang L and Lan P: A subpopulation of CD24 cells
in colon cancer cell lines possess stem cell characteristics.
Neoplasma. 59:282–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Salnikov AV, Bretz NP, Perne C, Hazin J,
Keller S, Fogel M, Herr I, Schlange T, Moldenhauer G and Altevogt
P: Antibody targeting of CD24 efficiently retards growth and
influences cytokine milieu in experimental carcinomas. Br J Cancer.
108:1449–1459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parker AL, Turner N, McCarroll JA and
Kavallaris M: βIII-tubulin alters glucose metabolism and stress
response signaling to promote cell survival and proliferation in
glucose-starved non-small cell lung cancer cells. Carcinogenesis.
37:787–798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zoni E, Chen L, Karkampouna S, Granchi Z,
Verhoef EI, La Manna F, Kelber J, Pelger RCM, Henry MD,
Snaar-Jagalska E, et al: CRIPTO and its signaling partner GRP78
drive the metastatic phenotype in human osteotropic prostate
cancer. Oncogene. 17:4739–4749. 2017. View Article : Google Scholar
|
|
17
|
Ogawa H, Kaira K, Takahashi K, Shimizu A,
Altan B, Yoshinari D, Asao T and Oyama T: Prognostic role of
BiP/GRP78 expression as ER stress in patients with gastric
adenocarcinoma. Cancer Biomark. 7:273–281. 2017. View Article : Google Scholar
|
|
18
|
Kawiak A, Domachowska A, Jaworska A and
Lojkowska E: Plumbagin sensitizes breast cancer cells to
tamoxifen-induced cell death through GRP78 inhibition and Bik
upregulation. Sci Rep. 13:437812017. View Article : Google Scholar
|
|
19
|
Lizardo MM, Morrow JJ, Miller TE, Hong ES,
Ren L, Mendoza A, Halsey CH, Scacheri PC, Helman LJ and Khanna C:
Upregulation of glucose-regulated protein 78 in metastatic cancer
cells is necessary for lung metastasis progression. Neoplasia.
28:699–710. 2016. View Article : Google Scholar
|
|
20
|
Gonzalez-Gronow M, Selim MA, Papalas J and
Pizzo SV: GRP78: A multifunctional receptor on the cell surface.
Antioxid Redox Signal. 11:2299–2306. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang M, Wey S, Zhang Y, Ye R and Lee AS:
Role of the unfolded protein response regulator GRP78/BiP in
development, cancer, and neurological disorders. Antioxid Redox
Signal. 11:2307–2316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rauschert N, Ndlein SB, Holzinger E,
Hensel F, Müller-Hermelink HK and Vollmers HP: A new tumor-specific
variant of GRP78 as target for antibody-based therapy. Lab Invest.
88:375–386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Papalas JA, Vollmer RT, Gonzalezgronow M,
Pizzo SV, Burchette J, Youens KE, Johnson KB and Selim MA: Patterns
of GRP78 and MTJ1 expression in primary cutaneous malignant
melanoma. Mod Pathol. 23:134–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fu W, Wu X, Li J, Mo Z, Yang Z, Huang W
and Ding Q: Upregulation of GRP78 in renal cell carcinoma and its
significance. Urology. 75:603–607. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xia YZ, Yang L, Xue GM, Zhang C, Guo C,
Yang YW, Li SS, Zhang LY, Guo QL and Kong LY: Combining GRP78
suppression and MK2206-induced Akt inhibition decreases
doxorubicin-induced P-glycoprotein expression and mitigates
chemoresistance in human osteosarcoma. Oncotarget. 7:56371–56382.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yerlikaya A, Erdogan E, Okur E, Yerlikaya
S and Savran B: A novel combination treatment for breast cancer
cells involving BAPTA-AM and proteasome inhibitor bortezomib. Oncol
Lett. 12:323–330. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kong DH, Zhang Q, Meng X, Zong ZH, Li C,
Liu BQ, Guan Y and Wang HQ: BAG3 sensitizes cancer cells exposed to
DNA damaging agents via direct interaction with GRP78. Biochim
Biophys Acta. 1833:3245–3253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Golden EB, Cho HY, Jahanian A, Hofman FM,
Louie SG, Schönthal AH and Chen TC: Chloroquine enhances
temozolomide cytotoxicity in malignant gliomas by blocking
autophagy. Neurosurg Focus. 37:E122014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luvsandagva B, Nakamura K, Kitahara Y,
Aoki H, Murata T, Ikeda S and Minegishi T: GRP78 induced by
estrogen plays a role in the chemosensitivity of endometrial
cancer. Gynecol Oncol. 126:132–139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gifford JB and Hill R: GRP78 influences
chemoresistance and prognosis in cancer. Curr Drug Targets. Jun
15–2017.(Epub ahead of print).
|
|
31
|
Soni P, Qayoom S, Husain N, Kumar P,
Chandra A, Ojha BK and Gupta RK: CD24 and nanog expression in stem
cells in glioblastoma: Correlation with response to chemoradiation
and overall survival. Asian Pac J Cancer Prev. 18:2215–2219.
2017.PubMed/NCBI
|
|
32
|
Kristiansen G, Machado E, Bretz N, Rupp C,
Winzer KJ, König AK, Moldenhauer G, Marmé F, Costa J and Altevogt
P: Molecular and clinical dissection of CD24 antibody specificity
by a comprehensive comparative analysis. Lab Invest. 90:1102–1116.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Zhang Y, Zhao Y, Liang Y, Xiang C,
Zhou H, Zhang H, Zhang Q, Qing H, Jiang B, et al: CD24 promoted
cancer cell angiogenesis via Hsp90-mediated STAT3/VEGF signalling
pathway in colorectal cancer. Oncotarget. 7:55663–55676.
2016.PubMed/NCBI
|
|
34
|
Huang YN, Guo X, You LP, Wang CJ, Liu JQ
and Li YL: Lysosome-associated protein transmembrane4β is involved
in multidrug resistance processes of colorectal cancer. Oncol Lett.
14:5229–5234. 2017.PubMed/NCBI
|