Introduction
Aberrant epigenetic regulations have been identified
in various types of cancer and a number of types of human disease
(1). The cancer epigenome is
characterized by genome-wide changes in DNA methylation and altered
histone modification patterns (1).
The global pattern of histone modifications may serve as a
predictor of the risk of recurrence of human cancer (2,3).
Epigenetics has become a common focus in studies investigating the
pathogenesis of many types of human diseases, particularly in
cancer. Hepatocellular carcinoma (HCC) is one of the most common
types of neoplasm, and the most frequent cause of cancer-associated
mortality (4,5). Previous evidence has suggested that
epigenetic alterations are important for the pathogenesis of HCC. A
whole-genome sequencing analysis of 27 HCCs revealed that almost
50% of the tumors analyzed exhibited mutations in chromatin
regulators, including histone methyltransferases (HMTs) (6). Previously, it has been demonstrated that
histone H3 trimethylated at Lys27 and histone H3
trimethylated at Lys4 may serve as prognostic markers
and therapeutic targets in HCC (7).
These results indicate that aberrant histone methylation may be
involved in the carcinogenesis of HCC, but the functions of HMT and
histone demethylase (HDM) in the pathogenesis of HCC require
additional study.
Euchromatic histone-lysine N-methyltransferase (G9A)
is the primary HMT for mono- and dimethylation of histone H3
Lys9 (H3K9) in vivo (8). Among various well-studied histone
methylations, H3K9 methylation is thought to be associated with
gene repression (9). Previously, G9A
has been identified to serve critical roles in various biological
progresses such as behavior plasticity, lymphocyte development,
stem cell differentiation and tumor cell growth (10–15). It
has been suggested that the expression level of G9A is increased in
numerous types of cancer compared with their corresponding normal
tissues, including melanoma, lung cancer, neuroblastoma, leukemia
and HCC (15–17). Specifically decreasing G9A expression
levels or inhibiting its activity may inhibit metastasis in lung
cancer and decrease cell proliferation by inducing autophagy in
colon cancer, breast cancer and neuroblastoma cells (15,18,19).
In the present study, the clinical significance of
G9A expression and its potential function in HCC progression was
investigated. The increased G9A expression level in HCC was
additionally confirmed in 5 publicly available datasets and the HCC
samples. By analyzing GSE14520, it was identified that increased
expression of G9A may serve as an indicator for a poor prognosis in
multinodular HCC. Gene set enrichment analysis (GSEA) was utilized
to predict the potential function of G9A, and the results
demonstrated that G9A may modulate gene sets involving RNA
processing and DNA replication. G9A inhibition significantly
decreased proliferation by arresting cells in G1 phase and
increased microtubule-associated protein light chain 3β (MAP1LC3B)
expression levels in Huh7 and HepG2 cell lines. An inverse
association between the expression of G9A and MAP1LC3B was
identified in HCC tumor samples in GSE14520, which indicated that
G9A also had the potential to regulate MAP1LC3B expression in HCC
tumor tissues. These results demonstrated that G9A was involved in
the pathogenesis of HCC. These data may assist additional
understanding of the crucial role of G9A in tumorigenesis.
Materials and methods
HCC specimens and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
HCC tumor specimens were obtained from the
Department of Pathology, Renmin Hospital of Wuhan University
(Wuhan, China). Informed consent was obtained from each patient
prior to surgery. The research protocol was approved by Wuhan
University School of Basic Medical Sciences Ethics Committee
(Wuhan, China). The total RNA of each sample was extracted using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), according to the manufacturer's protocol. RNA was
reverse-transcribed into cDNA using a RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. RT-qPCR was performed using the Takara
Premix Ex Taq II 820 (Takara Bio, Inc., Otsu, Japan) according to
the manufacturer's protocol on an ABI 7500 instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Primer pairs for human
β-actin and G9A were as follows: β-actin forward,
5′-AGCGCGCATCCCCCAAAGTT-3′ and β-actin reverse,
5′-GGGCACGAAGGCTCATCATT-3′; G9A forward, 5′-AGCCCTGCCCTGAGGATTAC-3′
and G9A reverse, 5′-ATGAGCACGCCTGGTTACACT-3′. An unpaired Student's
t-test was used to compare the expression levels of G9A in HCC
tumor tissues and matched non-cancerous tissues. P<0.05 was
considered to indicate a statistically significant difference.
GEO HCC gene expression data
analysis
The datasets of patients with HCC and corresponding
clinical data were downloaded from the publicly available Gene
Expression Omnibus (GEO) datasets (www.ncbi.nlm.nih.gov/gds). A total of four independent
datasets from GSE6764 (20), GSE14520
(21,22), GSE14323 (23) and GSE50579 (24) were utilized to analyze the expression
level of G9A in HCC. For GSE14520, GSE6764 and GSE14323,
log2 intensity of probe 202326 were used to represent
the expression level of G9A. For GSE50579, log2
intensity of probe A23P214 was used to represent the expression
level of G9A. The log2 intensities of probes 207484 and
208786 were used to represent G9A and MAP1LC3B expression,
respectively, to perform linear regression analysis in HCC tissues
of the GSE14520 testing group (n=225). An unpaired Student's t-test
was used to compare the expression levels of G9A in HCC tumor
tissues and normal tissues. P<0.05 was considered to indicate a
statistically significant difference.
GSEA
A Java program for GSEA (www.broadinstitute.org/gsea) was used to analyze the
potential genes affected by increased G9A expression. The data from
the GSE14520 testing group of patients with HCC were downloaded and
divided into two groups (high and low, determined by comparing with
the median value) according to the expression of G9A and Molecular
Signatures Database c5 (GO gene sets, 1454 gene sets). Gene sets
with a false discovery rate (FDR) <0.25 and nominal P<0.05
after 1,000 permutations were regarded as significant
enrichment.
Cell culture and reagents
Human HCC cell lines Huh7 and HepG2 were obtained
from The China Center for Type Culture Collection (Wuhan, China)
and kept in our lab. Huh7 and HepG2 were cultured in RPMI-1640
medium (cat. no. 11875-085, Gibco; Thermo Fisher Scientific, Inc.)
and Dulbecco's modified Eagle's medium (cat. no. 11965-084, Gibco;
Thermo Fisher Scientific, Inc.) containing 10% fetal bovine serum
(cat. no. 10082147, Gibco; Thermo Fisher Scientific, Inc.),
respectively. The cells were cultured at 37°C in a humidified
atmosphere containing 5% CO2. G9A inhibitor BIX-01294
(cat. no. B9311; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was
dissolved in sterilized water and then added to the medium to a
final concentration of 5 µM. Following incubation for 24, 48 or 72
h, cells were harvested and analyzed in the following
experiments.
Western blotting
Subsequent to treatment, the whole-cell extract was
prepared by scraping the Huh7 and HepG2 cells in
radioimmunoprecipitation buffer (cat. no, G2002, Servicebio Inc.,
Woburn, MA, USA). The protein concentrations were determined using
a BCA protein assay kit (cat. no. P0011; Beyotime Institute of
Biotechnology, Haimen, China). For every sample, 50 µg total
proteins was used and separated by SDS-PAGE (12% gels) and
transferred onto polyvinylidene fluoride membranes. The membranes
were blocked at room temperature with non-fat milk for 1 h and
incubated with primary antibodies (1:1,000) overnight at 4°C.
Secondary antibodies [goat anti-rabbit (cat. no. 926-32211) or
mouse (cat. no. 926-32210) IR Dye 800CW, LI-COR Biosciences,
Lincoln, NE, USA] were used to detect binding of the primary
antibodies by incubation at room temperature for 50 min. The bands
were detected using the Odyssey CLx imaging system (LI-COR
Biosciences) and analyzed by Image Studio Software for Odyssey CLx
(LI-COR Biosciences). The primary antibodies used in the present
study against the following were all from Cell Signaling
Technology, Inc. (Danvers, MA, USA): β-actin (cat. no. 4970), GAPDH
(cat. no. 2118), MAP1LC3B (cat. no. 2775), G2/mitotic-specific
cyclin B1 (cyclin B1; cat. no. 4135) and cyclin-dependent kinase
inhibitor 1B (p27; cat. no. 2552).
Flow cytometry
Following BIX-01294 treatment, Huh7 and HepG2 cells
were trypsinized and washed briefly with PBS once to remove the
residual serum and trypsin. For cell cycle analysis, cells were
fixed in 1 ml ice-cold 70% ethanol overnight at −20°C. Following
fixation, cells were pelleted at 800 × g at 4°C for 5 min, the
supernatant was aspirated carefully so as not to lose the pellet.
The pellets were briefly washed twice with ice-cold PBS. The cells
were resuspended with 500 µl propidium iodide (PI)/RNase A solution
(50 µg/ml PI, 100 µg/ml RNase A and 0.1% Triton X-100 in PBS). The
cells were incubated for 20 min at 37°C. DNA content was measured
using a FACSCalibur instrument (BD Biosciences, Franklin Lakes, NJ,
USA), and Modfit LT v3.3 (Verity Software House, Inc., Topsham, ME,
USA) was used to analyze cell cycle phase distribution. For Annexin
V/PI double staining analysis, following treatment with BIX-01294
as described above, HCC cells were double-stained with Annexin
V-FITC and PI and incubated at room temperature for 5 min in the
dark. A total of 10×104 cells from each group were
analyzed using a FACSCalibur instrument.
Statistical analysis
Experiments were performed at least 3 times. Results
are expressed as the mean ± standard deviation. Statistically
significant differences among the groups were determined by one-way
analysis of variance with Tukey's post hoc tests using SPSS 19.0
(IBM Corp., Armonk, NY, USA) and are presented as *P<0.05,
**P<0.01 and ***P<0.001. P<0.05 was considered to indicate
a statistically significant difference. Linear regression was used
to determine the correlation between two variants with GraphPad
Prism version 5.01 (GraphPad Software, Inc., La Jolla, CA, USA).
The correlation between gene expression and the clinicopathologic
features was analyzed by χ2 test. Kaplan-Meier plots
were constructed, and a log-rank test was used to evaluate
differences in G9A expression levels for overall survival.
Results
G9A expression is increased in HCC
tissues
It has been demonstrated that the expression level
of G9A was increased in a number of types of cancer, including HCC
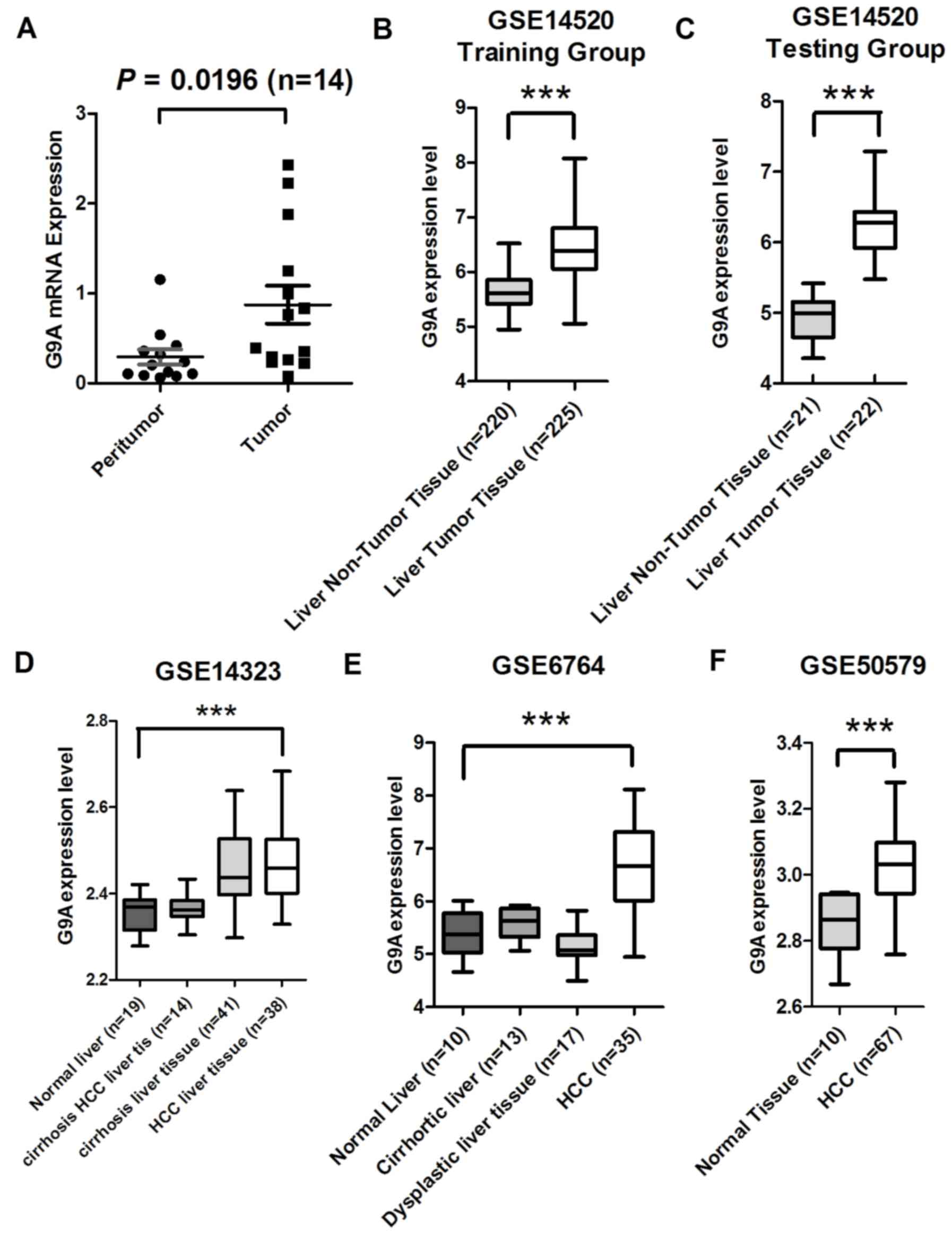
(16). First, in the present study
the relative mRNA expression level was examined in 14 pairs of HCC
tissue samples using RT-qPCR, and identified a significant increase
in G9A mRNA expression level in HCC tumor specimens compared with
their adjacent non-tumorous liver tissue samples (P=0.0196;
Fig. 1A). Subsequently, publicly
available microarray datasets were analyzed to observe whether a
similar phenomenon may be revealed. A total of four independent HCC
microarray datasets containing HCC and normal liver tissues from
the GEO were analyzed, including the GSE14520 training group (n=43)
and testing group (n=445) (21,22); and
GSE6764 (n=75) (20), GSE14323
(n=124) (23) and GSE50579 (n=80)
(24). All four datasets exhibited
significantly increased expression levels of G9A mRNA in HCC tumor
tissues compared with their non-tumor liver tissues (P<0.0001 in
all datasets; Fig. 1B-F).
Collectively, these data additionally confirmed that G9A expression
level was significantly increased in HCC.
Association of G9A expression with the
clinicopathological features of HCC
The association between G9A expression and
clinicopathological features of HCC with GSE14520 were next
analyzed to identify the potential roles of G9A in the pathogenesis
of HCC. The samples pooled in the HCC dataset GSE14520 were divided
into two groups according to the expression level of G9A in tumor
tissues, and examined using a χ2 test. The top 50%
samples with the highest G9A expression were considered the
high-expression group, and the other 50% as the low-expression
group. As summarized in Table I, G9A
expression was significantly associated with the serum
α-fetoprotein (AFP) level (P=0.009). It appeared that patients with
a lower expression level of G9A were more likely to exhibit a lower
serum AFP level. With the exception of a moderate association
between G9A expression and cirrhosis (P=0.054), there was no
significant association between its expression level and the other
clinicopathological features (P>0.05; Table I).
 | Table I.Association between G9A expression
and the clinicopathological features of the hepatocellular
carcinoma cases from the GSE14520 dataset. |
Table I.
Association between G9A expression
and the clinicopathological features of the hepatocellular
carcinoma cases from the GSE14520 dataset.
|
|
| G9A expression |
|
|
|---|
|
|
|
|
|
|
|---|
|
Characteristics | Case size | High | Low | χ2 value | P-value |
|---|
| Sex |
|
|
|
|
|
|
Male | 190 | 98 | 92 |
|
|
|
Female | 30 | 11 | 19 | 2.305 | 0.129 |
| α-fetoprotein
level, ng/ml |
|
|
|
|
|
| High
(>300) | 99 | 58 | 41 |
|
|
| Low
(≤300) | 118 | 48 | 70 | 6.909 | 0.009a |
| Age, years |
|
|
|
|
|
|
>50 | 108 | 51 | 57 |
|
|
|
≤50 | 114 | 59 | 55 | 0.456 | 0.500 |
| ALT level, U/l |
|
|
|
|
|
| High
(>50) | 91 | 46 | 45 |
|
|
| Low
(≤50) | 129 | 63 | 66 | 0.063 | 0.802 |
| Tumor size, cm |
|
|
|
|
|
| Large
(>5) | 80 | 42 | 38 |
|
|
| Small
(≤5) | 139 | 66 | 73 | 0.512 | 0.474 |
| Multinodular |
|
|
|
|
|
|
Yes | 45 | 23 | 22 |
|
|
| No | 175 | 86 | 89 | 0.055 | 0.814 |
| Cirrhosis |
|
|
|
|
|
|
Yes | 202 | 104 | 98 |
|
|
| No | 18 | 5 | 13 | 3.716 | 0.054 |
|
Tumor-node-metastasis stage |
|
|
|
|
|
| I | 93 | 46 | 47 |
|
|
| II | 77 | 36 | 41 |
|
|
|
III | 48 | 25 | 23 | 0.345 | 0.841 |
| Barcelona clinic
liver cancer staging |
|
|
|
|
|
| A | 148 | 69 | 79 |
|
|
| B | 22 | 12 | 10 |
|
|
| C | 28 | 14 | 14 | 0.535 | 0.765 |
| Cancer liver
Italian program staging |
|
|
|
|
|
| 0 | 97 | 42 | 55 |
|
|
| 1 | 74 | 38 | 36 |
|
|
| 2 | 35 | 20 | 14 |
|
|
| 3 | 9 | 5 | 4 |
|
|
| 4 | 3 | 2 | 1 |
|
|
| 5 | 1 | 0 | 1 | 4.601 | 0.467 |
Association between G9A expression and
survival of patients with HCC
To investigate the association between G9A
expression and survival of patients with HCC, Kaplan-Meier
estimator analysis was used to evaluate the effect of its
expression level on the survival of patients with HCC by using the
GSE14520 datasets stratified by clinical features included age,
sex, active hepatitis B virus status (chronically active viral
replication), AFP serum level, nodular type, cirrhosis, tumor size
(>5 cm, large; ≤5 cm small) and various tumor staging methods
such as Barcelona Clinic Liver Cancer, Cancer Liver Italian Program
or Tumor Node Metastasis classification (21,22). The
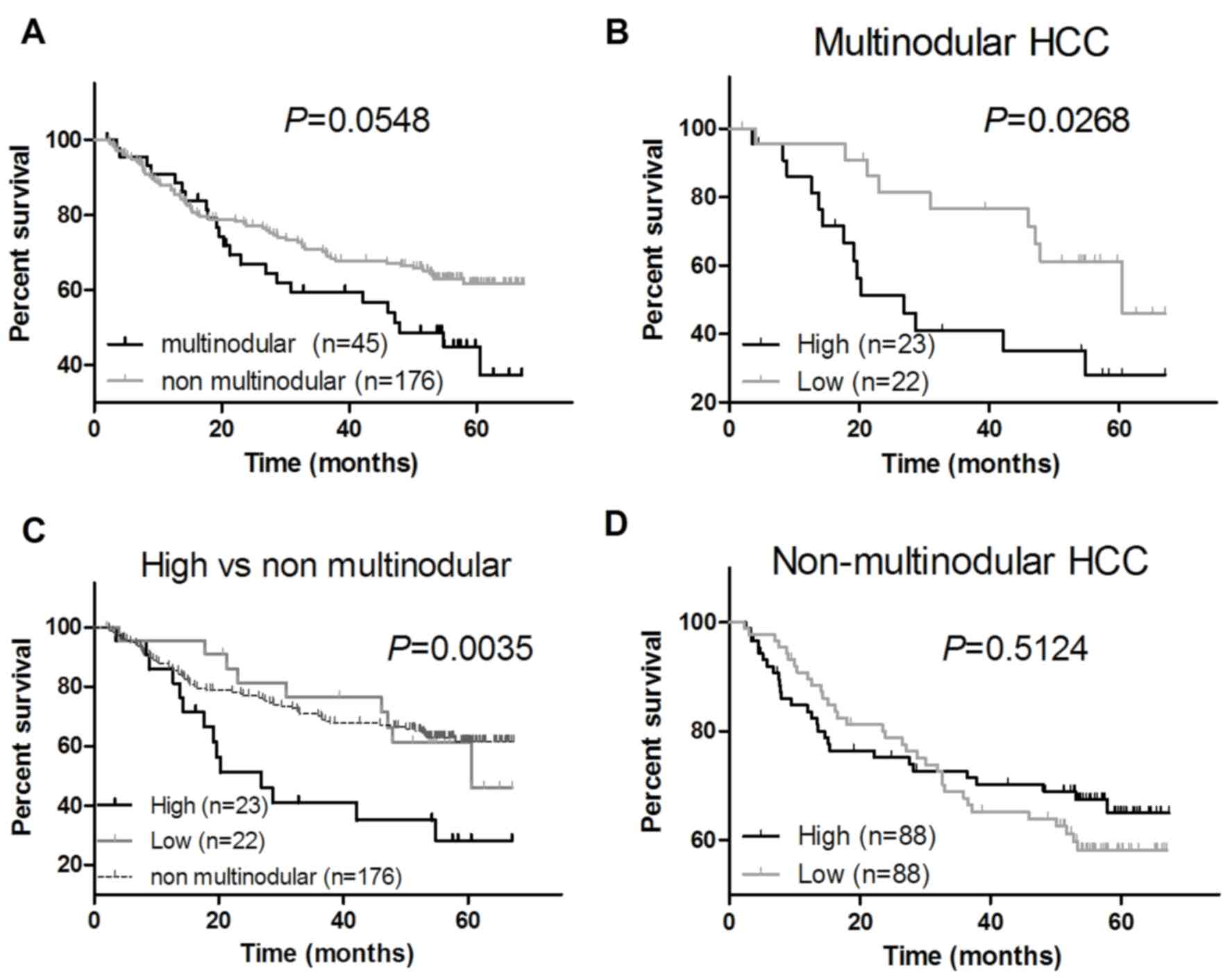
result of survival analysis demonstrated a weak difference in the
survival between patients with multinodular HCC and patients with
non-multinodular HCC (P=0.0548; Fig.
2A). Furthermore, the results of the survival analysis
indicated that the survival rates of patients with multinodular HCC
whose tumor tissues exhibited an increased G9A expression level was
significantly decreased compared with the survival rates of
patients with multinodular HCC with a decreased G9A expression
level (P=0.0268; Fig. 2B), and also
significantly decreased compared with that of the patients with
non-multinodular HCC (P=0.0035; Fig.
2C). Additionally, there was no marked association between the
survival and G9A expression level in patients with non-multinodular
HCC (P=0.5124; Fig. 2D) or HCC
patients with any other clinical features (data not shown).
G9A inhibitor represses HCC cell
growth
Results of previous studies have suggested that
suppression of G9A may inhibit proliferation in numerous types of
cancer cell (15–17). To investigate the potential function
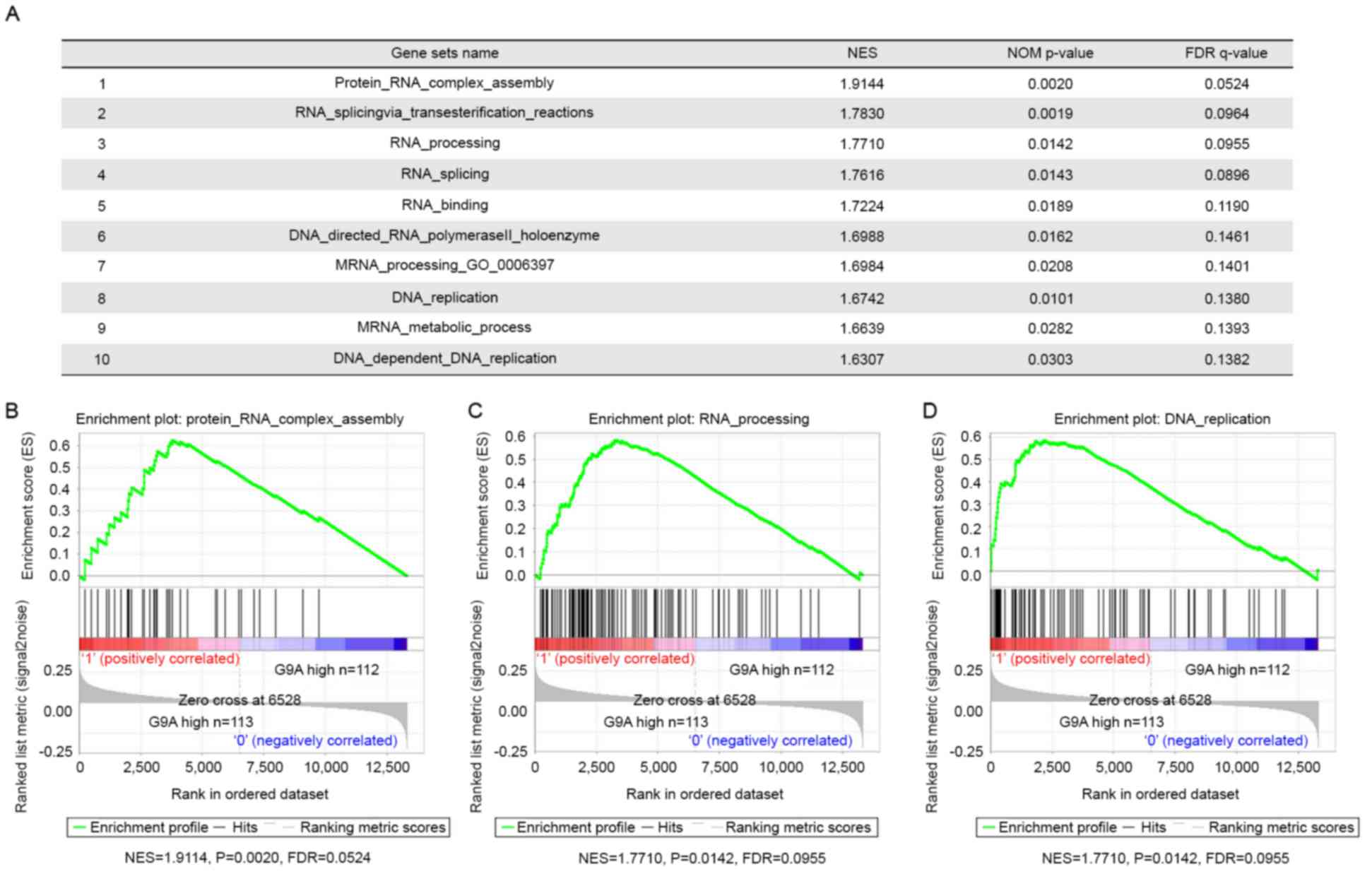
of G9A in HCC pathogenesis, GSEA using HCC tumor gene profiling
data from the GSE14520 group (n=225) was performed. There were 94
gene sets that were significantly upregulated in the ‘G9A high’
group compared with the ‘G9A low’ group (FDR <0.25 and nominal
P<0.05), among which there were 10 gene sets associated with RNA
and DNA processing. These results indicated that G9A-regulated gene
sets that were clearly associated with RNA processing and DNA
replication, and that G9A was involved in tumor cell proliferation
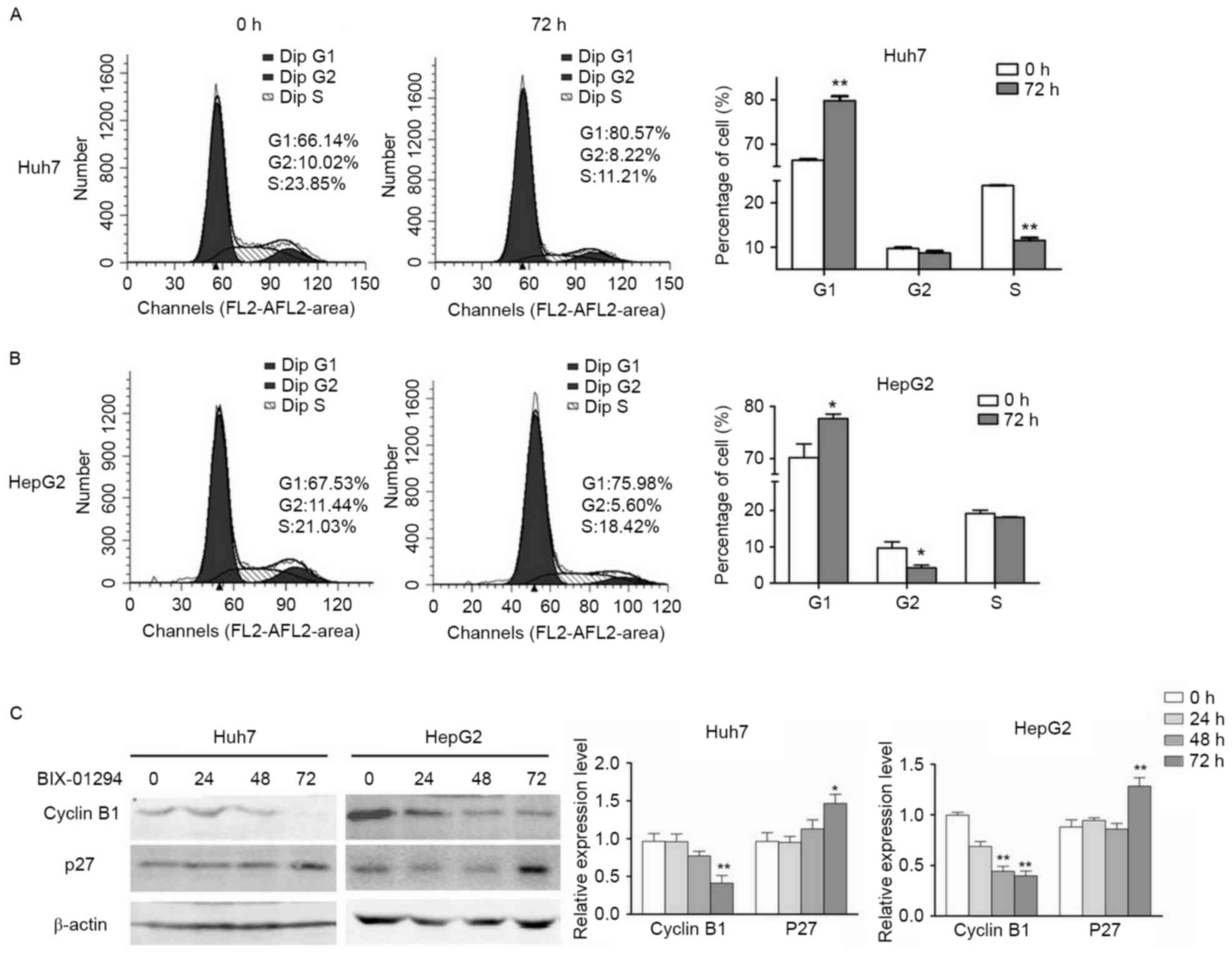
in HCC (Fig. 3). DNA content analysis
was performed and identified that the Huh7 and HepG2 cells were
arrested in G1 phase by G9A-specific inhibition, and that HepG2
also exhibited a significant decrease in the population of cells in
G2/M phase (Fig. 4A and B). In
accordance with the results of the flow cytometry analysis, the
expression levels of cyclin B1 were decreased following G9A
inhibition, and the expression level of p27 was increased in Huh7
and HepG2 cells (Fig. 4C). Taken
together, these results suggested that G9A dysfunction may suppress
HCC cell proliferation.
G9A inhibitor increases MAP1LC3B
expression in HCC cells
It has been suggested that G9A inhibition may induce
autophagy and increase the expression level of a number of
autophagy-associated genes in colon cancer, breast cancer and
neuroblastoma cells (15,19). To assess whether apoptosis was also
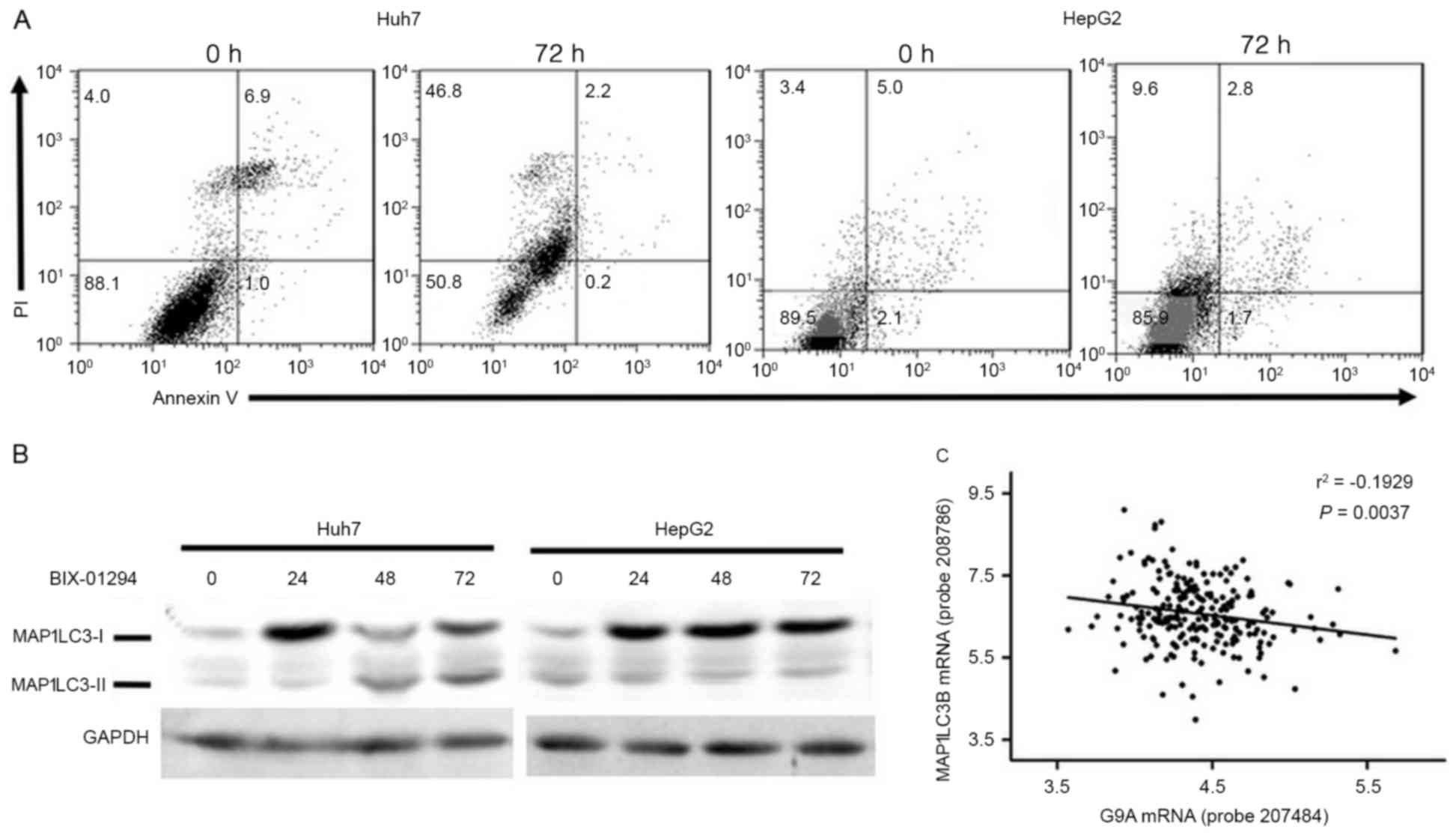
induced, Annexin V/PI double staining was performed and the results
indicated that the number of PI+ cells, instead of
Annexin V+, was significantly increased in Huh7
(Fig. 5A), which indicated that cell
death induced by G9A inhibition was a result of necrosis rather
than apoptosis. The expression of MAP1LC3B was examined by western
blotting, and it was identified that its expression was triggered
within 24 h following G9A inhibition in Huh7 and HepG2 cells, and
it was processed into MAP1LC3B-II subsequent to 48 h treatment in
Huh7 cells (Fig. 5B). A linear
regression analysis was performed using HCC tumor samples of the
GSE14520 dataset to determine whether the association between
expression of G9A and MAP1LC3B existed in HCC tumor tissues. A
significant negative association was identified between G9A and
MAP1LC3B expression in HCC samples of GSE14520 (n=225; r=−0.1929;
P=0.0037; Fig. 5C). The results of
the present study indicated that G9A may also exhibit the potential
to regulate MAP1LC3B expression in HCC tumor tissues.
Discussion
H3K9 methyltransferase G9A is involved in a number
of important developmental events, such as the lineage commitment
of stem cells and early embryonic stem cell differentiation
(14,25). It has also been demonstrated to be
overexpressed in numerous types of cancer, such as breast,
prostate, colon, bladder, ovarian, melanoma, lung and liver cancer
(16–18,26–28).
Despite this, its functions in tumorigenesis require additional
investigation. In the present study, its potential functions in the
progression of HCC were preliminarily explored, and the association
between G9A expression and clinicopathological features of HCC were
analyzed.
The results of the present study from RT-qPCR
analysis of HCC samples and the publicly available microarray
datasets, additionally confirmed that G9A was overexpressed in HCC
tumor tissues. These results suggested that increased G9A
expression was a biomarker of malignant cells. By analyzing the
association between G9A expression and clinicopathological features
using GSE14520, it was identified that its expression level was
significantly associated with serum AFP level, and that the high
expression level of G9A may indicate poor prognoses in patients
with multinodular HCC. As serum AFP level is a primary tumor
serological marker for HCC diagnosis, it was hypothesized that G9A
expression level may be of assistance in diagnosing HCC, and may
also have significant prognostic value in HCC.
The increased level of G9A expression in many types
of tumor tissue clearly indicated that this molecule may promote
the survival of tumor cells (15–17). This
assumption had been confirmed by the GSEA analysis in the present
study, which indicated that G9A may modulate genes that are
associated with RNA processing and DNA replication, suggesting that
G9A was involved in cell proliferation. The results of the present
study, and those of a number of previous studies, have demonstrated
that G9A inhibition may suppress cell proliferation in HCC cells
and numerous other types of cancer cell (15,16,19). As
G9A is important for cell proliferation, it is important to
consider the crucial role of its primary target, H3K9me2. A
previous study indicated that G9A and H3K9me2 were involved in
R-loop formation, efficient transcriptional termination and gene
post-transcriptional regulation (29). Therefore, it was hypothesized that G9A
served a crucial role in cell proliferation, which may be involved
in its role in tumorigenesis.
Autophagy is an evolutionarily conserved
‘self-eating’ process that is characterized by the formation of
autophagosomes (30). MAP1LC3B is
essential for membrane elongation and closure, which is the key
molecular event in autophagy (30).
It has been demonstrated experimentally that G9A is an epigenetic
regulator for MAP1LC3B expression, and that G9A inhibition may
induce autophagy-associated cell death (15,19,31). The
present study demonstrated that G9A inhibition may increase
MAP1LC3B expression in Huh7 and HepG2 cells. Furthermore, by
analyzing the publicly available datasets in the GSE14520 testing
group, a significant inverse association between G9A and MAP1LC3B
expression levels was identified. These results provide evidence
that G9A may have the potential to regulate MAP1LC3B expression in
HCC tumor tissues.
Despite increased G9A expression being demonstrated
as being crucial for tumor cell proliferation and autophagy-induced
cell death (15–17) its clinical significance in these types
of cancer remains obscure. The present study provided results that
the high expression of G9A may serve as an indicator for poor
prognosis in multinodular HCC, and additionally confirmed an
association between G9A and cell proliferation and MAP1LC3B
expression in HCC cells. The results also demonstrated that G9A was
involved in the pathogenesis of HCC, and may also serve as a
potential target for HCC therapy. As aforementioned, its importance
was not only confined to tumor cells. It is vital to disable G9A
specifically in tumor cells without affecting normal cells during
cancer therapy. The underlying molecular mechanisms of its
expression remain obscure and require elucidation, which would
assist in understanding the origin of G9A-associated cancer, and
the developmental events in which G9A is involved.
In conclusion, the present study investigated the
association between G9A expression level and the
clinicopathological features of HCC by using publicly available
datasets, and identified that G9A expression was associated with
cell proliferation and autophagy in HCC cells. These results
provide novel evidence for additional understanding of the crucial
role of G9A in the tumorigenesis of HCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Hubei Province in China (grant nos.
2012FFB04316 and 2013CFB255), The Incubator Project of Renmin
Hospital Wuhan University (grant no. 2013RMFH008), and the National
Natural Science Foundation of China (grant no. 31300609).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
JQ and QL conducted the majority of the experiments.
ZZ, PW, YJ and YH collected the samples. TL contributed to data
analysis and manuscript drafting. XJ and QZ contributed to Flow
Cytometry. JQ and LC contribute to design and manuscript.
Ethics approval and consent to
participate
The research protocol was approved by Wuhan
University School of Basic Medical Sciences Ethics Committee.
Informed written consent was obtained from all participants
involved in the study.
Consent for publication
Informed written consent was obtained from all
participants involved in the study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rodriguez-Paredes M and Esteller M: Cancer
epigenetics reaches mainstream oncology. Nat Med. 17:330–339. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fraga MF, Ballestar E, Villar-Garea A,
Boix-Chornet M, Espada J, Schotta G, Bonaldi T, Haydon C, Ropero S,
Petrie K, et al: Loss of acetylation at Lys16 and trimethylation at
Lys20 of histone H4 is a common hallmark of human cancer. Nat
Genet. 37:391–400. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wood LD, Parsons DW, Jones S, Lin J,
Sjöblom T, Leary RJ, Shen D, Boca SM, Barber T, Ptak J, et al: The
genomic landscapes of human breast and colorectal cancers. Science.
318:1108–1113. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fujimoto A, Totoki Y, Abe T, Boroevich KA,
Hosoda F, Nguyen HH, Aoki M, Hosono N, Kubo M, Miya F, et al:
Whole-genome sequencing of liver cancers identifies etiological
influences on mutation patterns and recurrent mutations in
chromatin regulators. Nat Genet. 44:760–764. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gao SB, Xu B, Ding LH, Zheng QL, Zhang L,
Zheng QF, Li SH, Feng ZJ, Wei J, Yin ZY, et al: The functional and
mechanistic relatedness of EZH2 and menin in hepatocellular
carcinoma. J Hepatol. 61:832–839. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shinkai Y and Tachibana M: H3K9
methyltransferase G9a and the related molecule GLP. Genes Dev.
25:781–788. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dawson MA and Kouzarides T: Cancer
epigenetics: From mechanism to therapy. Cell. 150:12–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maze I, Covington HE III, Dietz DM,
LaPlant Q, Renthal W, Russo SJ, Mechanic M, Mouzon E, Neve RL,
Haggarty SJ, et al: Essential role of the histone methyltransferase
G9a in cocaine-induced plasticity. Science. 327:213–216. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thomas LR, Miyashita H, Cobb RM, Pierce S,
Tachibana M, Hobeika E, Reth M, Shinkai Y and Oltz EM: Functional
analysis of histone methyltransferase g9a in B and T lymphocytes. J
Immunol. 181:485–493. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Katoh K, Yamazaki R, Onishi A, Sanuki R
and Furukawa T: G9a histone methyltransferase activity in retinal
progenitors is essential for proper differentiation and survival of
mouse retinal cells. J Neurosci. 32:17658–17670. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ueda J, Ho JC, Lee KL, Kitajima S, Yang H,
Sun W, Fukuhara N, Zaiden N, Chan SL, Tachibana M, et al: The
hypoxia-inducible epigenetic regulators Jmjd1a and G9a provide a
mechanistic link between angiogenesis and tumor growth. Mol Cell
Biol. 34:3702–3720. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen X, Skutt-Kakaria K, Davison J, Ou YL,
Choi E, Malik P, Loeb K, Wood B, Georges G, Torok-Storb B and
Paddison PJ: G9a/GLP-dependent histone H3K9me2 patterning during
human hematopoietic stem cell lineage commitment. Genes Dev.
26:2499–2511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ke XX, Zhang D, Zhu S, Xia Q, Xiang Z and
Cui H: Inhibition of H3K9 methyltransferase G9a repressed cell
proliferation and induced autophagy in neuroblastoma cells. PloS
One. 9:e1069622014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang J, Dorsey J, Chuikov S, Pérez-Burgos
L, Zhang X, Jenuwein T, Reinberg D and Berger SL: G9a and Glp
methylate lysine 373 in the tumor suppressor p53. J Biol Chem.
285:9636–9641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kondo Y, Shen L, Suzuki S, Kurokawa T,
Masuko K, Tanaka Y, Kato H, Mizuno Y, Yokoe M, Sugauchi F, et al:
Alterations of DNA methylation and histone modifications contribute
to gene silencing in hepatocellular carcinomas. Hepatol Res.
37:974–983. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen MW, Hua KT, Kao HJ, Chi CC, Wei LH,
Johansson G, Shiah SG, Chen PS, Jeng YM, Cheng TY, et al: H3K9
histone methyltransferase G9a promotes lung cancer invasion and
metastasis by silencing the cell adhesion molecule Ep-CAM. Cancer
Res. 70:7830–7840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim Y, Kim YS, Kim DE, Lee JS, Song JH,
Kim HG, Cho DH, Jeong SY, Jin DH, Jang SJ, et al: BIX-01294 induces
autophagy-associated cell death via EHMT2/G9a dysfunction and
intracellular reactive oxygen species production. Autophagy.
9:2126–2139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wurmbach E, Chen YB, Khitrov G, Zhang W,
Roayaie S, Schwartz M, Fiel I, Thung S, Mazzaferro V, Bruix J, et
al: Genome-wide molecular profiles of HCV-induced dysplasia and
hepatocellular carcinoma. Hepatology. 45:938–947. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roessler S, Long EL, Budhu A, Chen Y, Zhao
X, Ji J, Walker R, Jia HL, Ye QH, Qin LX, et al: Integrative
genomic identification of genes on 8p associated with
hepatocellular carcinoma progression and patient survival.
Gastroenterology. 142:957–966. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roessler S, Jia HL, Budhu A, Forgues M, Ye
QH, Lee JS, Thorgeirsson SS, Sun Z, Tang ZY, Qin LX and Wang XW: A
unique metastasis gene signature enables prediction of tumor
relapse in early-stage hepatocellular carcinoma patients. Cancer
Res. 70:10202–10212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mas VR, Maluf DG, Archer KJ, Yanek K, Kong
X, Kulik L, Freise CE, Olthoff KM, Ghobrial RM, McIver P and Fisher
R: Genes involved in viral carcinogenesis and tumor initiation in
hepatitis C virus-induced hepatocellular carcinoma. Mol Med.
15:85–94. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neumann O, Kesselmeier M, Geffers R,
Pellegrino R, Radlwimmer B, Hoffmann K, Ehemann V, Schemmer P,
Schirmacher P, Bermejo Lorenzo J and Longerich T: Methylome
analysis and integrative profiling of human HCCs identify novel
protumorigenic factors. Hepatology. 56:1817–1827. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Melcer S, Hezroni H, Rand E,
Nissim-Rafinia M, Skoultchi A, Stewart CL, Bustin M and Meshorer E:
Histone modifications and lamin A regulate chromatin protein
dynamics in early embryonic stem cell differentiation. Nat Commun.
3:9102012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kondo Y, Shen L, Ahmed S, Boumber Y,
Sekido Y, Haddad BR and Issa JP: Downregulation of histone H3
lysine 9 methyltransferase G9a induces centrosome disruption and
chromosome instability in cancer cells. PloS One. 3:e20372008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cho HS, Kelly JD, Hayami S, Toyokawa G,
Takawa M, Yoshimatsu M, Tsunoda T, Field HI, Neal DE, Ponder BA, et
al: Enhanced expression of EHMT2 is involved in the proliferation
of cancer cells through negative regulation of SIAH1. Neoplasia.
13:676–684. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vedadi M, Barsyte-Lovejoy D, Liu F,
Rival-Gervier S, Allali-Hassani A, Labrie V, Wigle TJ, Dimaggio PA,
Wasney GA, Siarheyeva A, et al: A chemical probe selectively
inhibits G9a and GLP methyltransferase activity in cells. Nat Chem
Biol. 7:566–574. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Skourti-Stathaki K, Kamieniarz-Gdula K and
Proudfoot NJ: R-loops induce repressive chromatin marks over
mammalian gene terminators. Nature. 516:436–439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Barth S, Glick D and Macleod KF:
Autophagy: Assays and artifacts. J Pathol. 221:117–124. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Narvajas Artal-Martinez A, Gomez TS,
Zhang JS, Mann AO, Taoda Y, Gorman JA, Herreros-Villanueva M, Gress
TM, Ellenrieder V, Bujanda L, et al: Epigenetic regulation of
autophagy by the methyltransferase G9a. Mol Cell Biol.
33:3983–3993. 2013. View Article : Google Scholar : PubMed/NCBI
|