Introduction
The piwi like RNA-mediated gene silencing 1 (Hiwi)
gene is a human homolog of the P-element Induced wimpy testis
(Piwi) gene family, which is a class of genes required for stem
cell self-renewal in a range of organisms, including jellyfish,
Caenorhabditis elegans, Drosophila melanogaster, Danio rerio,
Mus musculus and humans (1).
Previously, overexpression of Hiwi was reported to be associated
with poor prognosis in patients with various human malignant
tumors, including seminomas (2),
esophageal squamous cell carcinoma (3), adenocarcinoma of the pancreas (4), gastric adenocarcinoma (5), colorectal cancer (6), soft-tissue sarcoma (7) and endometrial carcinoma (8), indicating that it may represent a
promising biomarker and a potential target for anticancer
treatment.
The function of Hiwi in tumorigenesis is unclear.
Upregulation of Hiwi has been demonstrated to promote tumor cell
growth in breast (9), cervical
(10), endometrial (8) and colorectal cancer (6), as well as in mesenchymal stem cells
(11), while its downregulation has
been noted to suppress the growth, invasion and migration of glioma
(12), gastric (5) and lung cancer (13). These observations indicate that Hiwi
may act as an oncogene during carcinogenesis. However, Sharma et
al (14) reported that
overexpression of Hiwi suppressed proliferation and induced
apoptosis of the acute myeloid leukemia-derived cell line KG1.
Overexpression of Hiwi was reported to inhibit the growth of
chronic myeloid leukemia K562 cells and enhance their
chemosensitivity to daunomycin (15).
These findings indicate that the biological functions of Hiwi may
vary between types of tumor, necessitating its role in each cancer
to be studied individually.
Hepatocellular carcinoma (HCC) is one of the most
common malignancies in China (16).
Elevated levels of Hiwi mRNA and protein have previously been
observed in HCC, and that Hiwi expression is positively associated
with tumor metastasis (17,18). It has been demonstrated that
downregulation of Hiwi using RNA interference (RNAi) significantly
suppressed the proliferation and invasion of HCC cell lines
(18,19). However, whether Hiwi exerts a direct
tumorigenic role in HCC remains unknown. In the present study, an
adenovirus vector was used to overexpress Hiwi in liver cells in
vitro and in vivo, and the effect of Hiwi on cell growth
and migration was evaluated in hepatocellular carcinoma SMMC7721
cells, primary hepatocytes and xenografts. Additionally, the effect
of Hiwi on hepatotoxicant-induced apoptosis was examined.
Materials and methods
Cell culture
The effect of Hiwi overexpression on cell growth was
evaluated in SMMC7721 cells and primary mouse hepatocytes, since
these cells were reported to express low levels of endogenous Hiwi
protein (18). 293 cells were
obtained from the American Type Culture Collection (Manassas, VA,
USA). SMMC7721 cells were established in 1977 by the Second
Military Medicine College (Shanghai, China). These cells were
derived from the liver tissue of a 50-year-old male with a grade
II–III (20) hepatocellular
carcinoma. SMMC7721 cells are epithelial in morphology. The number
of chromosomes varies between 44 and 107; 70% of the cells have
54–58 chromosomes. The cells are tumorigenic in nude mice but with
a low metastatic potential. SMMC7721 and 293 cells were maintained
in Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS, Invitrogen; Thermo Fisher Scientific,
Inc.) and penicillin/streptomycin.
Primary murine hepatocytes were isolated from male
ICR mic (n=6, six weeks old, mean average weight 25 g, purchased
from the experimental animal center of Nanjing medical University,
Nanjing, China) by in situ liver perfusion with collagenase
via the portal vein. Mice were allowed free access to drinking
water and food at room temperature (25°C) with an automatic 12 h
light and 12 h dark cycle. Mice were anesthetized with
pentobarbital sodium, and the livers were then perfused in
situ with 45 ml calcium-free buffer (100 mM HEPES buffer at pH
7.4, 50 mM EGTA), followed by 8 ml liver digest medium (100 mM
HEPES buffer at pH 7.6, with 0.5 mg/ml collagenase IV). Next, the
liver was excised, minced and strained through a steel mesh.
Hepatocytes were obtained by centrifugation 3 times at 50 × g for 5
min at 4°C and washed twice with DMEM. The hepatocytes were
cultured in DMEM supplemented with 10% FBS and
penicillin/streptomycin for further experiments.
Cloning of human cDNA and construction
of plasmids
Human Hiwi cDNA containing the full open reading
frame was synthesized commercially (Shanghai GeneChem Co., Ltd.,
Shanghai, China). The human Hiwi cDNA was sub-cloned into the
pcDNA3.1-myc vector, specifically into the myc epitope sequence
with EcoRI and XhoI sites, which yielded the
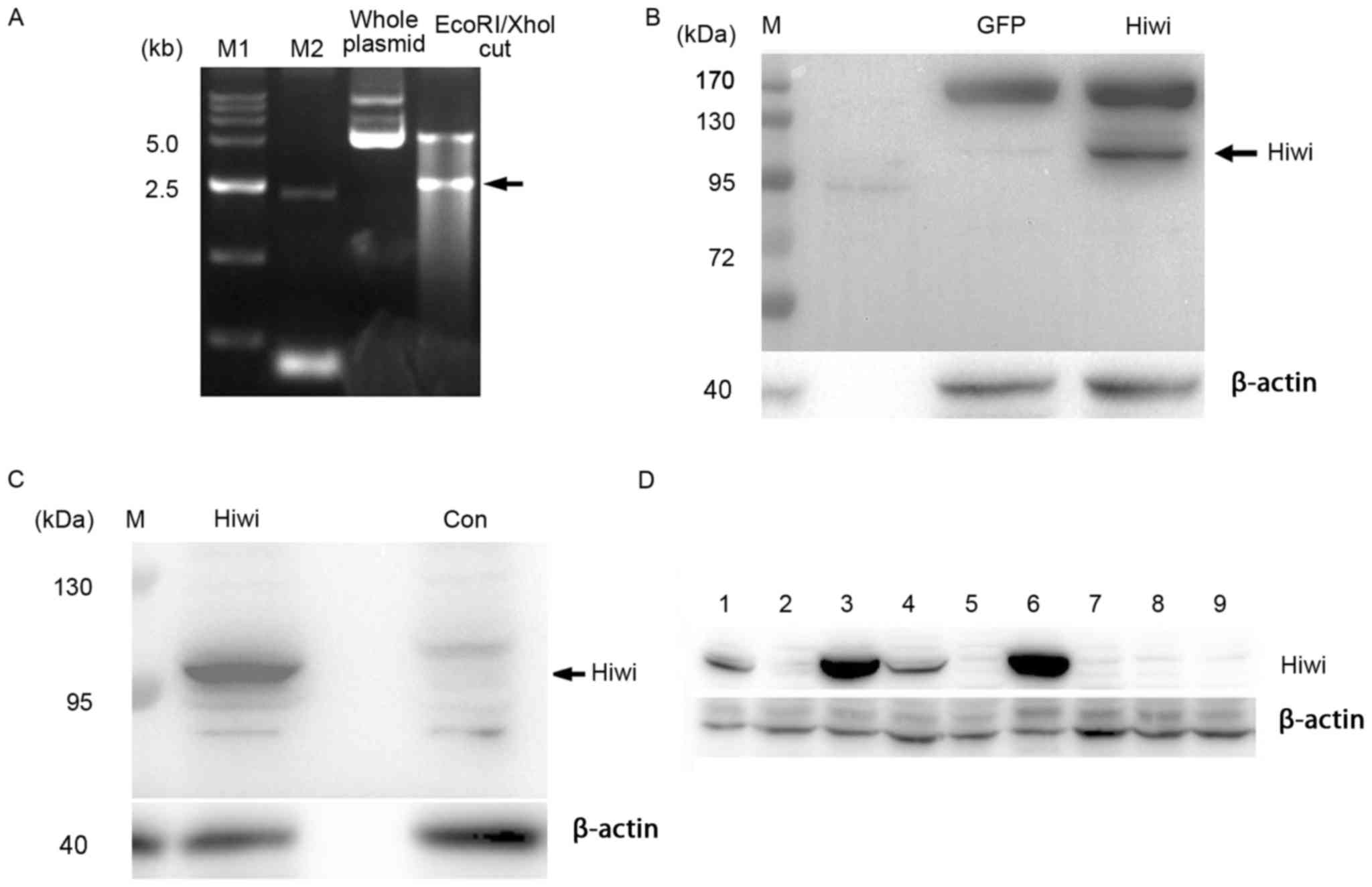
pcDNA3.1-myc-Hiwi plasmid (Fig. 1A).
The following primers were used: Forward
5′-AAAAAGAATTCACTGGGAGAGCCCGAGCCAGAGCC-3′; and reverse,
5′-AAAAAACTCGAGTTAGAGGTAGTAAAGGCGGTTTG-3′. The length of the
plasmid was confirmed by electrophoresis in 0.5× Tris-acetate EDTA
buffer with a 1.5% agarose gel containing 0.1 µg/ml of ethidium
bromide. The DNA bands were visualized and analyzed by Biosense 810
Gel Electrophoresis Image analytic system (Beyotime Institute of
Biotechnology, Haimen, China). The full Hiwi coding region was
verified by sequencing (performed commercially by Generay Co.,
Ltd., Shanghai, China).
Plasmid transfection
The 293 cells and SMMC7721 cells were transfected
with pcDNA3.1-myc-Hiwi using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions, and the controls were transfected with
the control vector. Cells were used for subsequent experiments 48 h
after transfection. For stable transfection, the cells were
selected with G418 (400 µg/ml; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for 2 weeks.
Construction of recombinant
adenoviruses
Recombinant adenoviruses expressing Hiwi were
constructed commercially using an AdEasy Adenoviral Vector system
(Stratagene; Agilent Technologies, Inc., Santa Clara, CA, USA) and
the pAdTrack cytomegalovirus vector (Shanghai GeneChem Co., Ltd.).
To induce adenovirus-mediated overexpression in vitro, 24 h
after plating, the cells were incubated at 37°C for 4 h with
adenovirus at a multiplicity of infection of 10–20 pfu per cell.
Next, the virus was removed, and the cells were cultured at 37°C
for an additional 24–48 h in fresh DMEM supplemented with 10% FBS
and penicillin/streptomycin. Under these conditions, infection
efficiency routinely exceeded 90%.
Confirmation of Hiwi overexpression by
western blotting
The cells were lysed with Radioimmunoprecipitation
Assay buffer containing protease inhibitors (Roche Diagnostics,
Basel, Switzerland) for 15 min on ice. Following centrifugation for
15 min at 20,000 × g (4°C), the protein content of the samples was
determined according to the Bradford method. The protein lysates
(50 µg per lane) were analyzed by SDS-PAGE (8% gel) and transferred
onto methanol-activated polyvinylidene fluoride membranes.
Non-specific binding sites were blocked with 5% non-fat milk in
Tris-buffered saline/0.1% Tween 20 for 2 h at room temperature. The
membranes were incubated overnight with primary antibodies as
follows: Hiwi (cat. no. ab12337; 1:1,000; Abcam, Cambridge, UK) or
myc (cat. no. MABE282; 1:2,000; Sigma-Aldrich; Merck KGaA).
Subsequent to being washed three times, the membranes were blotted
with corresponding horseradish peroxidase-conjugated secondary
antibody (cat. no. A0208 or A0216; 1:1,000; Beyotime Institute of
Biotechnology) for 1 h at room temperature. The membranes were
rinsed three times and then developed with enhanced
chemiluminescence reagent using the ECL chemiluminescence kit
(Thermo Fisher Scientific, Inc.). The two primary antibodies
detected proteins with a molecular mass of ~98 kDa (Fig. 1B and C and data not shown), which is
consistent with the reported molecular weight of human Hiwi
(2). β-actin (cat. no. A5316;
Sigma-Aldrich, USA) was used as the internal control.
Assessment of cell viability by MTT
assay
An MTT assay was performed in 96-well plates.
Briefly, 1×104 primary mouse hepatocytes were seeded per
well and cultured overnight prior to infection with the adenovirus.
At 0, 20, 40, 60, 80 h, 10 µl MTT (5 mg/ml; Sigma-Aldrich, Merck
KGaA) was added to each well, and after 4 h of incubation at 37°C,
the medium was gently decanted, and dimethysulfoxide (100 µl/well)
was added to dissolve the formazan product. The absorbance at 490
nm was measured with a microplate reader according to the
manufacturer's protocol.
Analysis of cell proliferation by
5-ethynyl-2′-deoxyuridine (EdU) assay
A total of 2×104 cells per well were
seeded in 48-well plates and cultured overnight prior to infection
with the adenovirus. At the indicated time, 150 µl of EdU (50 µM;
Guangzhou RiboBio Co., Ltd., Guangzhou, China) was added to each
well. After 4 h of incubation at 37°C, the cells were fixed and EdU
incorporation was visualized using a Leica microscope (Leica
Microsystems, Inc., Buffalo Grove, IL, USA).
Cell cycle analysis by flow
cytometry
SMMC7721 cells were collected and fixed with
ice-cold 70% ethanol at 4°C for 2 h. Next, the cells were washed
twice with PBS and incubated with DAPI in the presence of RNase A
at 37°C for 30 min in the dark. The DNA contents were then measured
using a flow cytometer (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and were analyzed by Modifit software (version
4.0; Verify Software House, Inc., Topsham, ME, USA) for the
proportions of cells in the phases of the cell cycle.
Assessment of cell migration in
culture
Two-dimensional cell migration was analyzed using a
scratch wound assay. SMMC7721 cells were cultured in 6-well plate.
The scratch was performed by using a 200 µl pipette tip to press
firmly against the top of the tissue culture plate and swiftly make
a vertical wound down through the cell monolayer. SMMC7721 cells
were fixed with 70% ethanol and captured using a light microscope
at 48 h after the scratch was made. The farthest distance that the
cells migrated from the wound edge was measured by calculating the
mean of three independent microscope fields in each of the three
independent experiments.
Three-dimensional cell migration was determined
using a Transwell migration assay as per the manufacturer's
instructions (Corning Life Science, Corning, NY, USA). A total of
5,000 cells in serum-free DMEM were added into the top chamber. The
bottom chamber contained 1% or 10% FBS in DMEM as a
chemoattractant. After 24 h, the cells that had not penetrated the
filters were scraped from the upper surface. The membrane was fixed
with formalin, and the cells that migrated to the bottom surface of
the filter were stained with Giemsa at room temperature for 5–10
min (Beyotime Institute of Biotechnology) and counted using a light
microscope (magnification, ×200). An average of four randomly
chosen fields were counted in each of the three independent
experiments.
Effect of Hiwi on genes involved in
the epithelial-mesenchymal transition (EMT) by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total cellular RNA was extracted using TRIzol
reagent (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's instructions. Total RNA (2 µg) was reverse
transcribed at 50°C for 50 min, and 70°C for 10 min using the
PrimeScript RT reagent kit (Takara Bio, Inc.). qPCR was performed
with Power SYBR Green PCR Master Mix (Applied Biosystems; Thermo
Fisher Scientific, Inc.) using a 7500 Real-Time PCR system. The
thermal cycling conditions of the PCR were 94°C for 5 min, followed
by 22–35 cycles for 20 sec at 94°C, 20 sec at 64°C, 1 min at 72°C,
and the final extension at 72°C for 7 min. The primer sequences
used are listed in Table I. The
relative quantities of mRNA were determined using the comparative
cycle threshold method (21) and
normalized against GAPDH mRNA.
 | Table I.Primer sequences for polymerase chain
reaction. |
Table I.
Primer sequences for polymerase chain
reaction.
| Gene | GeneBank accession
number | Species | Primer sequence
(5′-3′) |
|---|
| Hiwi | AF104260.2 | Human |
|
|
Forward | – | – |
GAAGCAGCCTGTCTTGGTCAGC |
|
Reverse | – | – |
GAATCAAAGCTCAAACCCCAGTCTC |
| E-cadherin | AB025106.1 | Human |
|
|
Forward | – | – |
GACACCAACGATAATCCT |
|
Reverse | – | – |
TTTCAGTGTGGTGATTACGACGTTA |
| N-cadherin | M34064.1 | Human |
|
|
Forward | – | – |
GGTGGAGGAGAAGAAGACCAG |
|
Reverse | – | – |
GGCATCAGGCTCCACAGT |
| α-catenin | D13866.1 | Human |
|
|
Forward | – | – |
AGCGAAGATTGCGGAACAGGT |
|
Reverse | – | – |
GCCTTGACCTTGCTGCAGATG |
| Vimentin | NM_003380.3 | Human |
|
|
Forward | – | – |
GAGAACTTTGCCGTTGAAGC |
|
Reverse | – | – |
TCCAGCAGCTTCCTGTAGGT |
| GAPDH | AJ005371.1 | Human |
|
|
Forward | – | – |
TGTTCGTCATGGGTGTGAACC |
|
Reverse | – | – |
GCAGTGATGGCATGGACTGTG |
| E-cadherin | NM_009864.2 | Mouse |
|
|
Forward | – | – |
TCGGAAGACTCCCGATTCAAA |
|
Reverse | – | – |
CGGACGAGGAAACTGGTCTC |
| N-cadherin | NM_007664.4 | Mouse |
|
|
Forward | – | – |
CTCCAACGGGCATCTTCATTAT |
|
Reverse | – | – |
CAAGTGAAACCGGGCTATCAG |
| Bcl2 | NM_009741.5 | Mouse |
|
|
Forward | – | – |
ATGCCTTTGTGGAACTATATGGC |
|
Reverse | – | – |
GGTATGCACCCAGAGTGATGC |
| Bax | NM_007527.3 | Mouse |
|
|
Forward | – | – |
AGACAGGGGCCTTTTTGCTAC |
|
Reverse | – | – |
AATTCGCCGGAGACACTCG |
| P53 | AB021961.1 | Mouse |
|
|
Forward |
|
|
GTCACAGCACATGACGGAGG |
|
Reverse |
|
|
TCTTCCAGATGCTCGGGATAC |
| Mcl1 | NM_008562.3 | Mouse |
|
|
Forward |
|
|
TCCAAGGACTCGAAGCCTCT |
|
Reverse |
|
|
CCAGTTTGTTACGCCATCTTTG |
| GAPDH | AY618568.1 | Mouse |
|
|
Forward |
|
|
AGGTCGGTGTGAACGGATTTG |
|
Reverse |
|
|
GGGGTCGTTGATGGCAACA |
Analysis of the effect of Hiwi or
green fluorescent protein overexpression on apoptosis
Perifosine, an inhibitor of RAC
serine/threonine-protein kinase (22)
and doxorubicin were used to induce apoptosis in primary murine
hepatocytes in vitro. A total of 5 µM Perifosine and 2 µM
doxorubicin (DOX) were separately added to the primary murine
hepatocytes 24 h following infection with recombinant adenovirus
that express Hiwi or green fluorescent protein (GFP). Cell
viability was evaluated at different time points by MTT assays.
Xenograft model to study tumor
growth
Six-week-old male BALB/C nude mice (n=18, purchased
from the experimental animal center of Nanjing medical University,
Nanjing, China) were used to study the effect of Hiwi
overexpression. They were allowed free access to drinking water and
food at room temperature (25°C) with an automatic 12 h light and 12
h dark cycle. All animal protocols and experimental procedures were
approved by the Animal Care and Use Committee of Nanjing Medical
University (Nanjing, China). SMMC7721 cells were transfected with
pcDNA3.1-myc-Hiwi plasmid and selected with 400 µg/ml G418 for 2
weeks, as aforementioned. A total of 9 colonies were selected. The
colonies (1,3,4,6) that stably expressed Hiwi were verified
by western blot analysis (Fig. 1D).
The colonies (2,5,7–9) that did not express Hiwi were used as
blank controls. Hiwi-expressing or blank control SMMC7721 cells
(2×107 for each sample) were suspended in 0.2 ml
serum-free DMEM media and implanted into the flank of the nude mice
mentioned above. After 70 days, the tumors were isolated, and the
volumes were calculated from caliper measurements of tumor
diameter.
Effects of Hiwi on genes associated
with apoptosis and EMT in vivo
Adenoviruses expressing Hiwi or GFP
(5×109 pfu) were injected into the tail vein of male
BALB/c mice (n=10). Anesthesia was achieved by the intraperitoneal
administration of freshly prepared sodium pentobarbital (60 mg/kg)
5 days later. The mice were sacrificed by cervical dislocation, and
the liver tissues were harvested. RT-qPCR was used to examine the
expression of Hiwi, B-cell lymphoma 2 (Bcl-2), myeloid cell
leukemia 1, Bcl-associated X, p53, E-cadherin and N-cadherin
expression as aforementioned.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistical significance was assessed using one-way analysis of
variance and Scheffe test. In all statistical comparisons,
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of Hiwi overexpression on the
proliferation and migration of liver cells
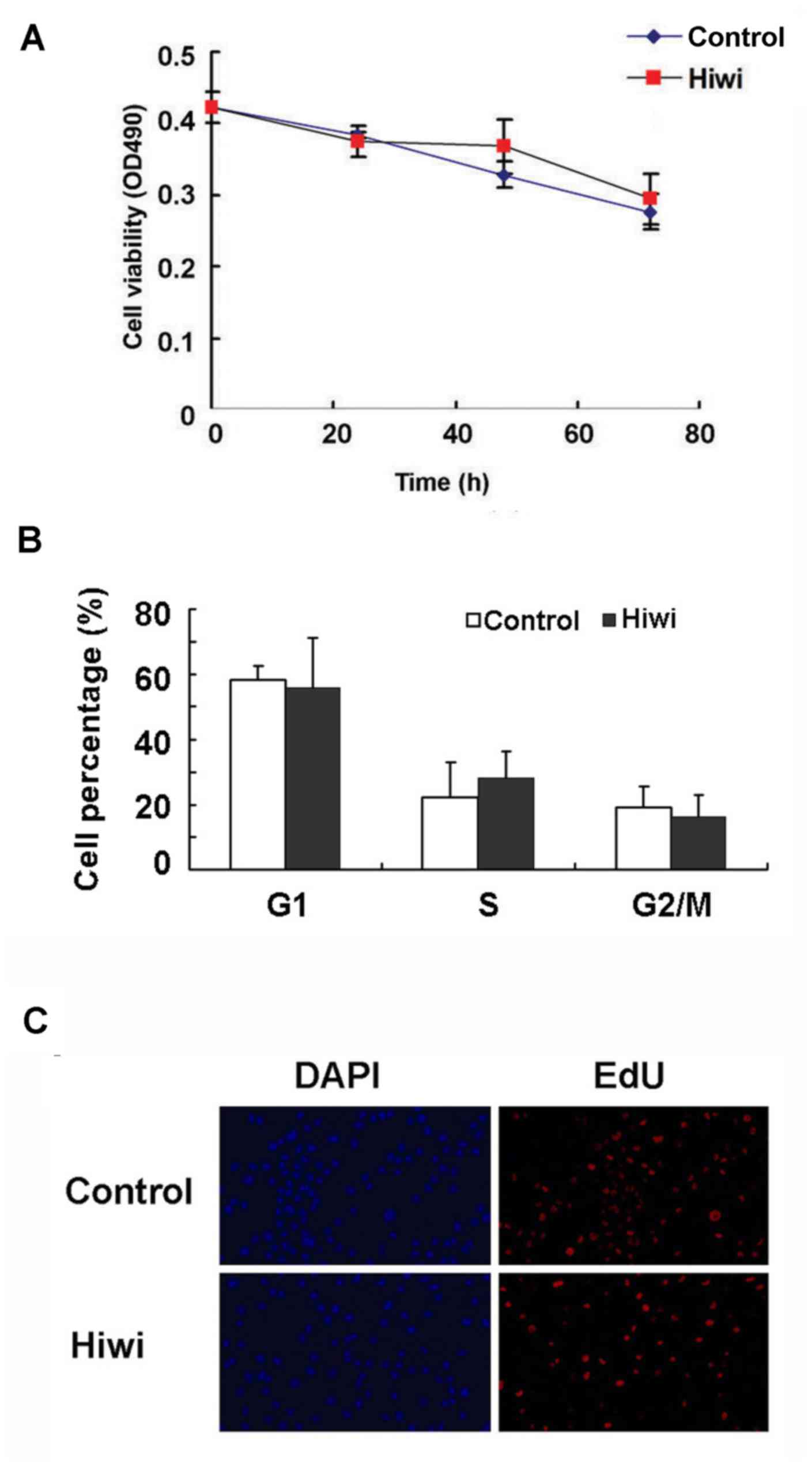
The growth curve of primary mouse hepatocytes that
were infected with Hiwi-expressing adenovirus did not differ
significantly from the control cells (Fig. 2A). Next, the functional role of Hiwi
in cell division and cell cycle of liver cells was analyzed. As
indicated in Fig. 2B, overexpression
of Hiwi did not alter the percentage of SMMC7721 cells in
G1, S and G2/M phases. No significant
difference was observed between the Hiwi-overexpressing and control
SMMC7721 cells in the EdU incorporation assays (Fig. 2C). These results indicate that
overexpression of Hiwi did not affect liver cell proliferation.
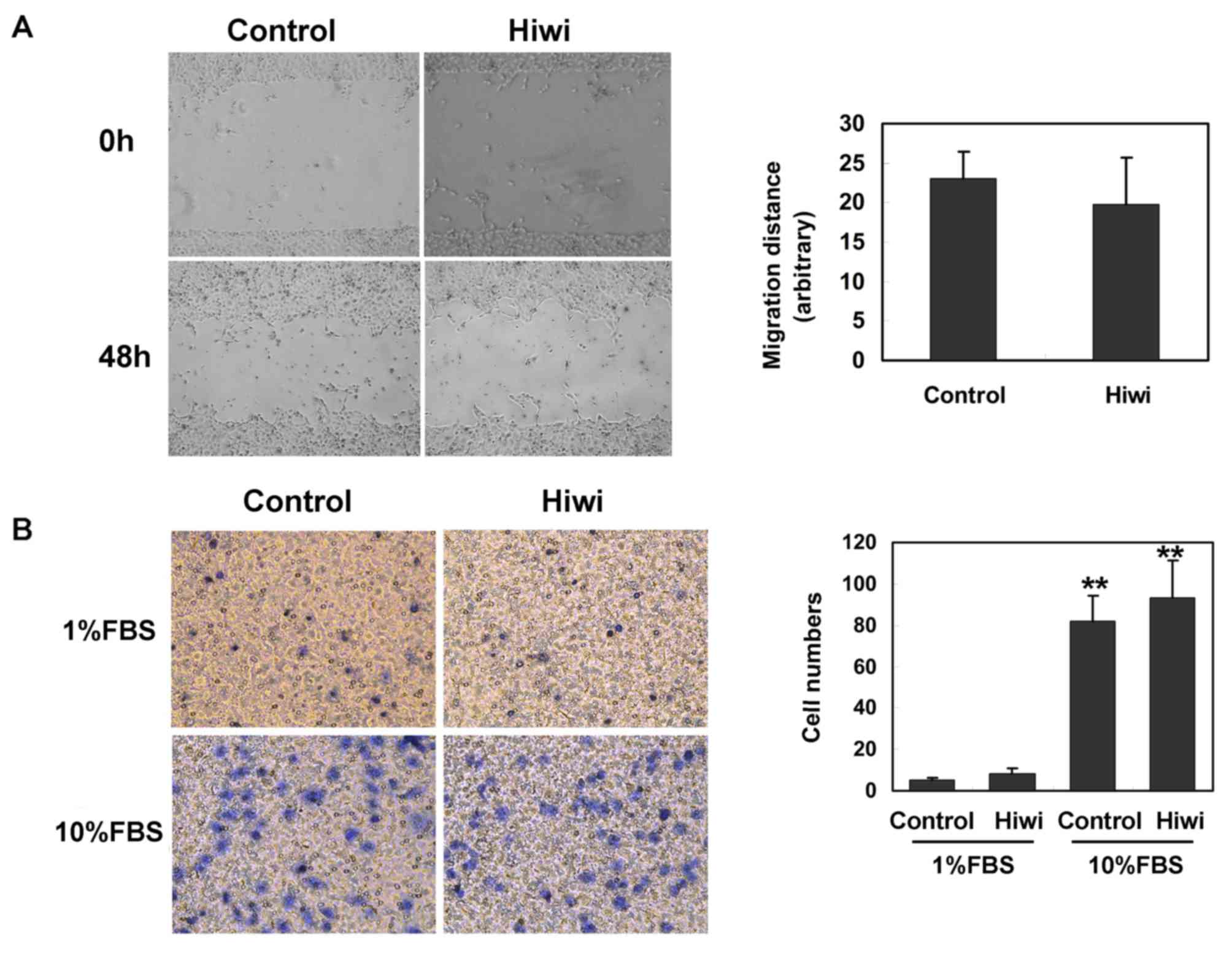
Subsequently, the effect of Hiwi on the invasion of
liver cells was examined. As indicated by the scratch migration
assay, the average distance that SMMC7721 cells overexpressing Hiwi
migrated at 48 h after the cell scratch did not differ
significantly from the control cells (Fig. 3A). The effect of Hiwi expression on
SMMC7721 cell migration was also assessed by Transwell assay using
a modified Boyden chamber. The fraction of SMMC7721 cells
overexpressing Hiwi that migrated through a gelatin-coated membrane
in response to 1% or 10% FBS did not differ significantly from the
control cells (Fig. 3B). These data
indicated that overexpression of Hiwi did not affect the migration
of liver cells.
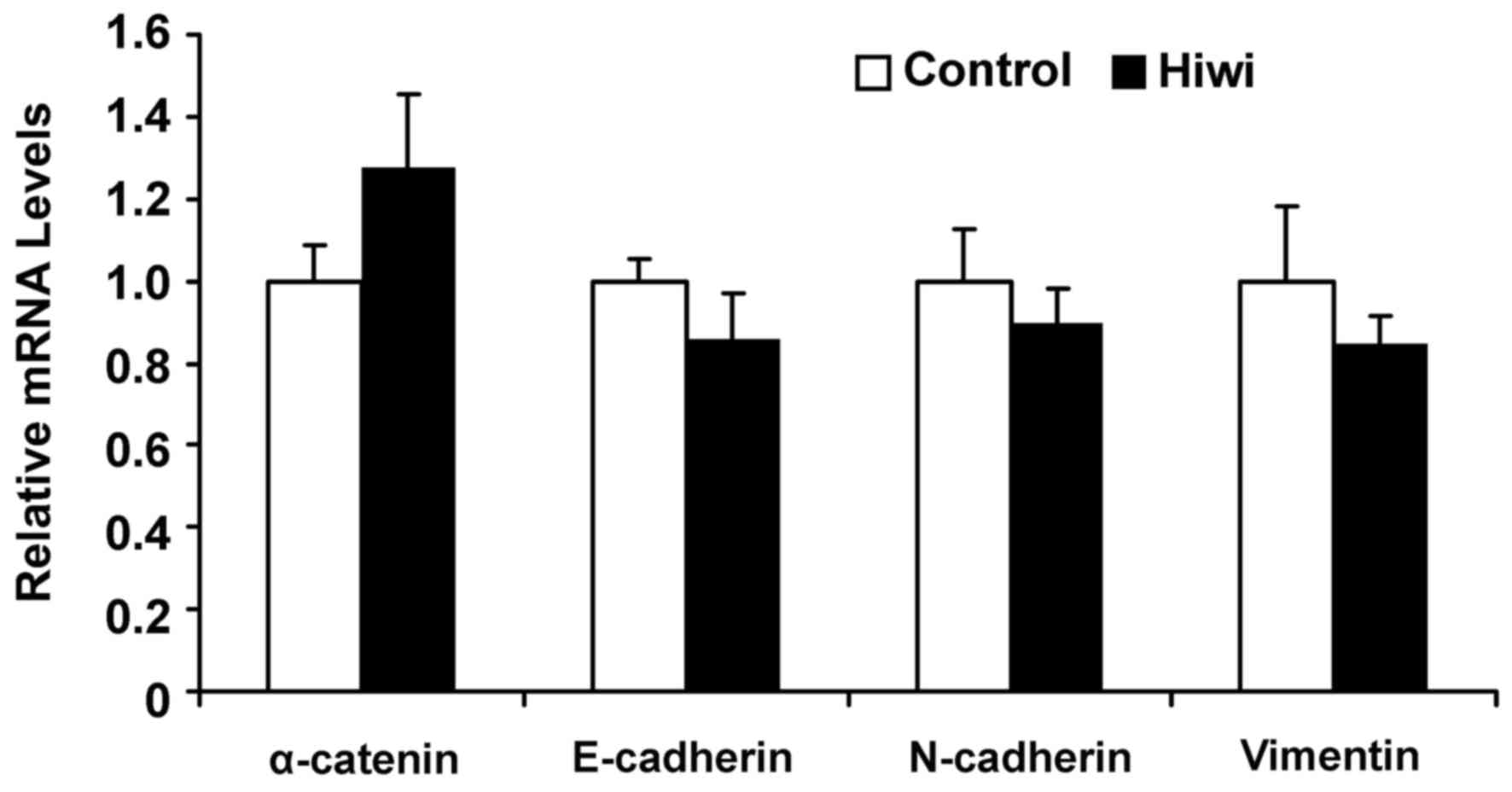
To assess the effect of Hiwi on genes involved in
EMT, levels of α-catenin, E-cadherin, N-cadherin and vimentin mRNA
were measured by RT-qPCR. Consistently, levels of α-catenin,
E-cadherin, N-cadherin and vimentin mRNA did not differ
significantly between Hiwi-overexpressing cells and control cells
(Fig. 4).
Effect of Hiwi on drug-induced
apoptosis in vitro
Oncoproteins, such as Bcl-2, contribute to
neoplastic cell growth primarily by promoting cell survival via
interfering with apoptosis (23).
Therefore, it was hypothesized that Hiwi might promote tumor growth
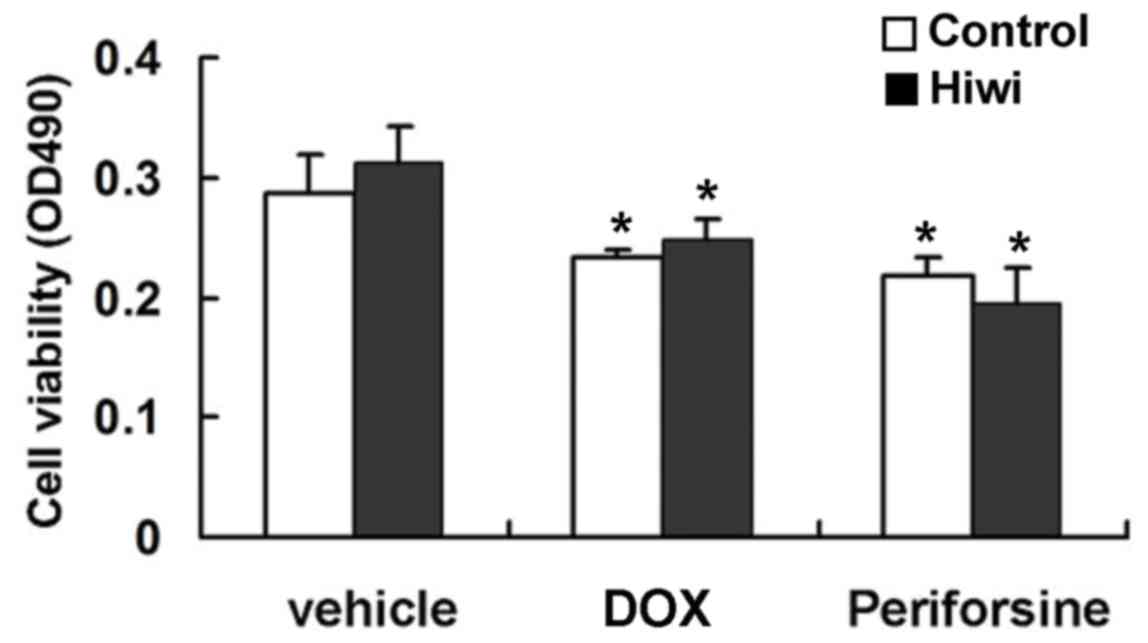
by inhibiting apoptosis. Mouse primary hepatocytes were treated
with perifosine and subsequently a significant reduction in cell
viability was observed (Fig. 5).
However, the overexpression of Hiwi did not enhance the survival of
perifosine-treated cells (Fig. 5).
DOX is a widely used anticancer drug that induces apoptosis
(24). Overexpression of Hiwi had no
detectable effect on DOX-induced hepatotoxicity and apoptosis
(Fig. 5). These results indicated
that overexpression of Hiwi did not inhibit apoptosis.
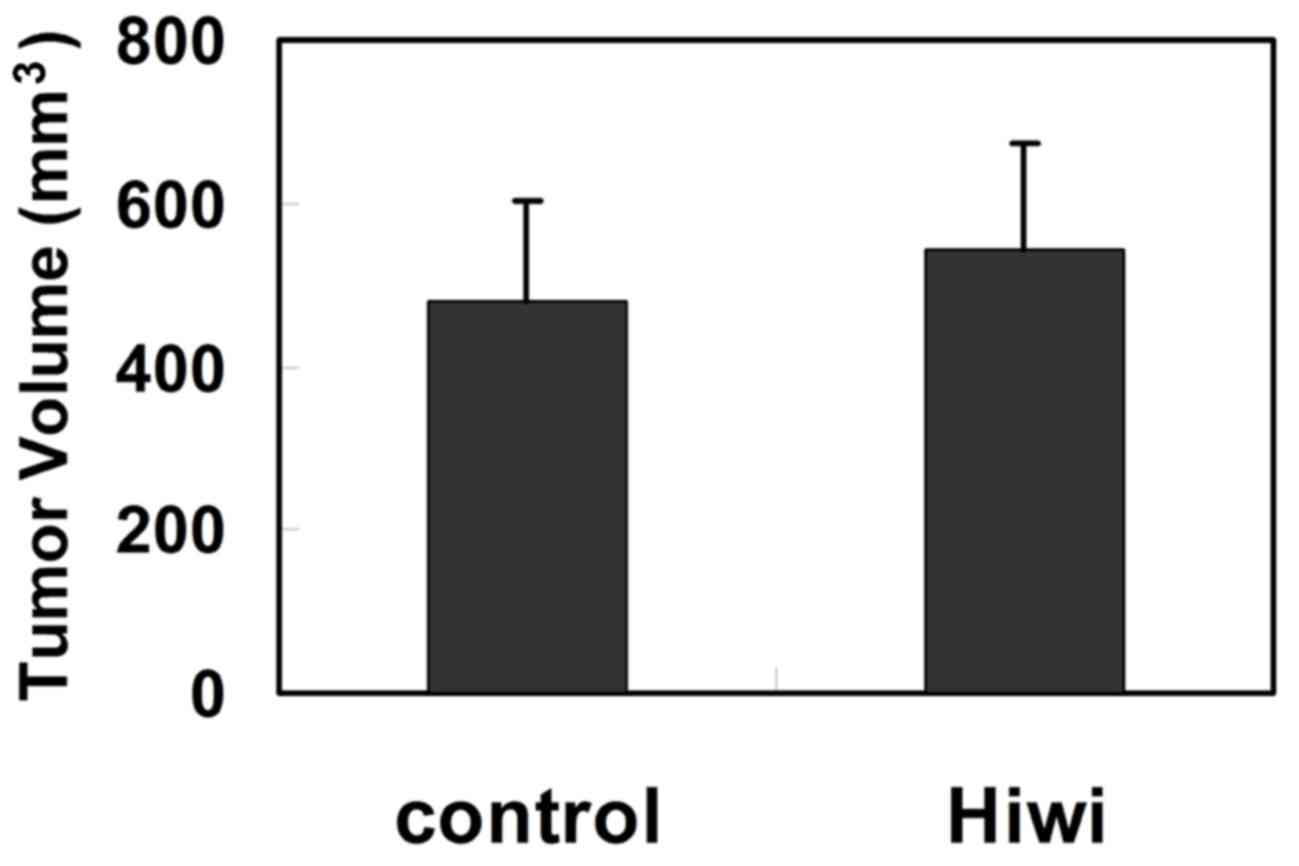
Effect of Hiwi overexpression on the
growth of HCC xenografts
Although exogenous expression of Hiwi did not affect
liver cell growth in vitro, the growth of
Hiwi-overexpressing HCC cells was examined in vivo.
Hiwi-expressing SMMC7721 cells were implanted into the flank of
6-weeks-old BALB/c nude mice. After 70 days, tumor volume was
measured. Consistently, the mean size of tumors derived from
Hiwi-expressing cells did not differ significantly from the tumors
that derived from control cells (Fig.
6). These results indicated that overexpression of Hiwi did not
alter tumor growth in vivo.
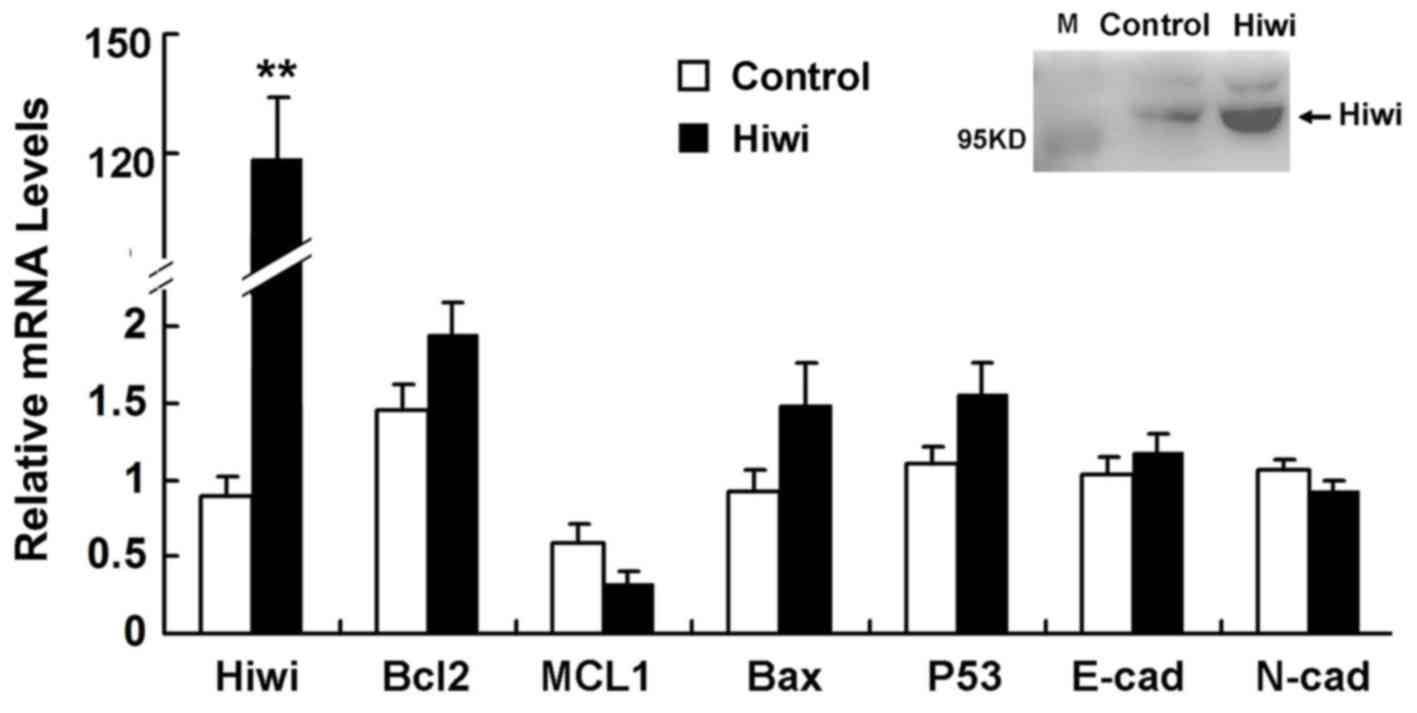
Role of Hiwi overexpression in
altering the expression of genes that are involved in EMT or
apoptosis
The mice were injected with adenovirus expressing
Hiwi or GFP (5×109 pfu) in the tail vein. After 5 days,
Hiwi was detected in hepatocytes via RT-qPCR and western blot
analysis. Expression of genes that are involved in EMT or apoptosis
expressed by these hepatocytes did not differ significantly between
mice that were injected with the Hiwi-expressing vector or the
control adenoviral vector (Fig.
7).
Discussion
Hiwi has been suggested to have a role in several
human malignant tumors (2,3,5,7), and increased expression of the Hiwi gene
has been demonstrated in seminoma cell hyperplasia (2), esophageal squamous cell carcinoma
(3), gastric carcinoma (5), and pancreas adenocarcinoma (4). In the majority of cases, increased
expression of Hiwi was detected in tumors and found to be
associated with poorer patient outcomes (2–8,17,18). These
prior studies indicated that Hiwi expression might be a useful
clinical marker of germ cell malignancies and solid tumors of
epithelial or mesenchymal origin.
Previous studies revealed that significant decreases
in Hiwi expression were observed in renal cell carcinoma, and Hiwi
expression was found to be inversely associated with overall
survival (25,26). Functional analysis of Hiwi protein has
also generated conflicting results. Although overexpression of Hiwi
has been demonstrated to promote the growth of breast or cervical
tumors (9,10), in acute or chronic myeloid leukemia
cell lines Hiwi proteins may suppress proliferation (14,15). These
findings indicate that the function of Hiwi differs in different
cellular contexts, and the biological function of Hiwi remains
poorly understood.
Since the downregulation of Hiwi suppressed
proliferation and invasion of HCC cells (18,19), we
hypothesized that the overexpression of Hiwi may contribute to the
development and progression of HCC. However, overexpression of Hiwi
in SMMC7721 HCC cell lines, primary mouse hepatocytes, xenografts
and an adenovirus-mediated mouse hepatic gene-expression model did
not alter the proliferation, migration or apoptosis of liver cells
in vitro or in vivo. Although Hiwi expression was
elevated in HCC (17,18), it did not appear to function as an
oncoprotein. The RNAi-mediated downregulation of Hiwi was
previously revealed to suppress the proliferation and invasion of
HCC cell lines (18). Therefore, we
hypothesize that Hiwi may be necessary but not sufficient for tumor
genesis of liver cells.
Similar results were previously obtained in gastric
cancer cells. Overexpression of Hiwi in AGS cells did not alter
their proliferative rate, whereas suppression of Hiwi expression
using antisense RNAs or RNAi inhibited cell growth and induced cell
cycle arrest (5). One possible
explanation for this discrepancy would be that, in certain types of
tumors, other cellular factors might be required to interact with
Hiwi to promote tumorigenesis. The existence of additional
signaling pathways that counteract the function of Hiwi in these
tumors cannot be ruled out. Additionally, the findings of the
present study do not necessarily reflect the role of Hiwi in humans
in vivo, and other unknown mechanisms may well affect tumor
growth in humans.
In summary, the findings of the present study
indicate that although expression of Hiwi is associated with the
development and progression of HCC, it does not act as an oncogene
in liver cancer cells. To characterize the contribution of Hiwi to
the progression of HCC in humans further, other factors or pathways
that interact with Hiwi require elucidation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81101800), the 12th
Six Talents Peak Project of Jiangsu Province (grant no.
2015-WSN-028), the Priority Academic Program Development of Jiangsu
Higher Education Institutions (PAPD) (grant no. JX10231801) and the
Research Project of Chinese Medical Association and Chinese High
Education Association (grant no. 2016B-KC019).
Availability of data and materials
The datasets analyzed during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
CXS and HaL performed the cloning and transfection
of plasmids, and generated SMMC7721 cells stably expressing Hiwi.
HuL and HaL performed the apoptosis assays and generated xenograft
model. HHZ and HMS assisted in the flow cytometry analysis. MDS,
JLC and SFX studied the expression of genes and analyzed the data.
HaL and JXJ designed the study and wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal protocols and experimental procedures
were approved by the Animal Care and Use Committee of Nanjing
Medical University (Nanjing, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
Glossary
Abbreviation
Abbreviations:
References
|
1
|
Suzuki R, Honda S and Kirino Y: PIWI
expression and function in cancer. Front Genet. 3:2042012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qiao D, Zeeman AM, Deng W, Looijenga LH
and Lin H: Molecular characterization of hiwi, a human member of
the piwi gene family whose overexpression is correlated to
seminomas. Oncogene. 21:3988–3999. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He W, Wang Z, Wang Q, Fan Q, Shou C, Wang
J, Giercksky KE, Nesland JM and Suo Z: Expression of HIWI in human
esophageal squamous cell carcinoma is significantly associated with
poorer prognosis. BMC Cancer. 9:4262009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grochola LF, Greither T, Taubert H, Möller
P, Knippschild U, Udelnow A, Henne-Bruns D and Würl P: The stem
cell-associated Hiwi gene in human adenocarcinoma of the pancreas:
Expression and risk of tumour-related death. Br J Cancer.
99:1083–1088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu X, Sun Y, Guo J, Ma H, Li J, Dong B,
Jin G, Zhang J, Wu J, Meng L and Shou C: Expression of hiwi gene in
human gastric cancer was associated with proliferation of cancer
cells. Int J Cancer. 118:1922–1929. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raeisossadati R, Abbaszadegan MR, Moghbeli
M, Tavassoli A, Kihara AH and Forghanifard MM: Aberrant expression
of DPPA2 and HIWI genes in colorectal cancer and their impacts on
poor prognosis. Tumour Biol. 35:5299–5305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taubert H, Greither T, Kaushal D, Würl P,
Bache M, Bartel F, Kehlen A, Lautenschläger C, Harris L, Kraemer K,
et al: Expression of the stem cell self-renewal gene Hiwi and risk
of tumour-related death in patients with soft-tissue sarcoma.
Oncogene. 26:1098–1100. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Z, Che Q, He X, Wang F, Wang H, Zhu
M, Sun J and Wan X: Stem cell protein Piwil1 endowed endometrial
cancer cells with stem-like properties via inducing
epithelial-mesenchymal transition. BMC Cancer. 15:8112015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang DW, Wang ZH, Wang LL, Song Y and
Zhang GZ: Overexpression of hiwi promotes growth of human breast
cancer cells. Asian Pac J Cancer Prev. 15:7553–7558. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu W, Gao Q, Chen K, Xue X, Li M, Chen Q,
Zhu G and Gao Y: Hiwi facilitates chemoresistance as a cancer stem
cell marker in cervical cancer. Oncol Rep. 32:1853–1860. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Siddiqi S, Terry M and Matushansky I: Hiwi
mediated tumorigenesis is associated with DNA hypermethylation.
PLoS One. 7:e337112012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Tong X, Gao H, Yan X, Xu X, Sun S,
Wang Q and Wang J: Silencing HIWI suppresses the growth, invasion
and migration of glioma cells. Int J Oncol. 45:2385–2392. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang D, Fang Z, Dong M, Liang C, Xing C,
Zhao J and Yang Y: Effect of RNA interference-related HiWi gene
expression on the proliferation and apoptosis of lung cancer stem
cells. Oncol Lett. 4:146–150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sharma AK, Nelson MC, Brandt JE, Wessman
M, Mahmud N, Weller KP and Hoffman R: Human CD34(+) stem cells
express the hiwi gene, a human homologue of the Drosophila gene
piwi. Blood. 97:426–434. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Y, Jiang Y, Ma N, Sang B, Hu X, Cong
X and Liu Z: Overexpression of Hiwi inhibits the growth and
migration of chronic myeloid leukemia cells. Cell Biochem Biophys.
73:117–124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kao JH and Chen DS: Changing disease
burden of hepatocellular carcinoma in the Far East and Southeast
Asia. Liver Int. 25:696–703. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang J, Zhang H, Tang Q, Hao B and Shi R:
Expression of HIWI in human hepatocellular carcinoma. Cell Biochem
Biophys. 61:53–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao YM, Zhou JM, Wang LR, He HW, Wang XL,
Tao ZH, Sun HC, Wu WZ, Fan J, Tang ZY and Wang L: HIWI is
associated with prognosis in patients with hepatocellular carcinoma
after curative resection. Cancer. 118:2708–2717. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie Y, Yang Y, Ji D, Zhang D, Yao X and
Zhang X: Hiwi downregulation, mediated by shRNA, reduces the
proliferation and migration of human hepatocellular carcinoma
cells. Mol Med Rep. 11:1455–1461. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Webber C, Gospodarowicz M, Sobin LH,
Wittekind C, Greene FL, Mason MD, Compton C, Brierley J and Groome
PA: Improving the TNM classification: Findings from a 10-year
continuous literature review. Int J Cancer. 135:371–378. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fei HR, Chen G, Wang JM and Wang FZ:
Perifosine induces cell cycle arrest and apoptosis in human
hepatocellular carcinoma cell lines by blockade of Akt
phosphorylation. Cytotechnology. 62:449–460. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miyashita T and Reed JC: Bcl-2 oncoprotein
blocks chemotherapy-induced apoptosis in a human leukemia cell
line. Blood. 81:151–157. 1993.PubMed/NCBI
|
|
24
|
Mizutani H, Tada-Oikawa S, Hiraku Y,
Kojima M and Kawanishi S: Mechanism of apoptosis induced by
doxorubicin through the generation of hydrogen peroxide. Life Sci.
76:1439–1453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Al-Janabi O, Wach S, Nolte E, Weigelt K,
Rau TT, Stöhr C, Legal W, Schick S, Greither T, Hartmann A, et al:
Piwi-like 1 and 4 gene transcript levels are associated with
clinicopathological parameters in renal cell carcinomas. Biochim
Biophys Acta. 1842:686–690. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iliev R, Stanik M, Fedorko M, Poprach A,
Vychytilova-Faltejskova P, Slaba K, Svoboda M, Fabian P, Pacik D,
Dolezel J and Slaby O: Decreased expression levels of PIWIL1,
PIWIL2, and PIWIL4 are associated with worse survival in renal cell
carcinoma patients. Onco Targets Ther. 9:217–222. 2016.PubMed/NCBI
|