Introduction
Lung cancer is the primary cause of
cancer-associated mortality worldwide and non-small cell lung
cancer (NSCLC) is the most common histological type of lung cancer.
Despite advances in treatment, the 5-year survival rate of patients
with NSCLC remains poor, at <15% (1).
Over the past few decades, epidermal growth factor
receptor-tyrosine kinase inhibitors (EGFR-TKIs), such as gefitinib,
have been the most widely used targeted therapy to treat patients
with lung cancer worldwide and have significantly improved the
overall survival rate of lung cancer (2,3). However,
some patients develop resistance to gefitinib following an initial
period of treatment (4). It has been
determined that acquired gefitinib resistance is primarily linked
to a T790M mutation, hepatocyte growth factor receptor or
insulin-like growth factor 1 receptor gene amplifications or
abnormalities of the phosphatase and tensin homolog (PTEN) and
mechanistic target of rapamycin (mTOR) proteins (5). Other reasons responsible for the
development acquired gefitinib resistance remain unknown; the
underlying mechanisms behind what causes this resistance to develop
are yet to be elucidated.
Exosomes are small extracellular membrane vesicles
40–100 nm in diameter, which are secreted by a wide range of cells
(6). Exosomes contain a substantial
amount of RNA and may be transferred from one cell to another,
thereby contributing to tumor growth, metastasis, angiogenesis and
drug resistance (7,8). Exosomes from drug-resistant breast
cancer cells (MCF-7/DOC) may transfer resistance to sensitive cells
(MCF-7/S) (9). In the presence of
exosomes extracted from drug-resistant cells (DOC/exo), MCF-7/S
developed drug resistance, which did not occur in cells exposed to
their own exosomes (S/exo) (9). It
has been reported that, following the release of exosomes by NSCLC
A549 cells during cisplatin stimulation, the sensitivity of A549
cells to cisplatin was decreased; this process may have been
mediated by the exchange of microRNAs (miRNA) and mRNAs between
exosomes via cell-to-cell communication (10).
To the best of our knowledge, the involvement of
exosomes in the development of resistance to gefitinib in lung
cancer cells remains unknown. In the present study, the effect of
exosomes on the transmission of gefitinib resistance from
gefitinib-resistant HCC827 lung cancer cells to their
gefitinib-sensitive counterparts was investigated and the potential
underlying mechanisms by which this may occur was explored.
Materials and methods
Reagents and cell culture
Gefitinib (Iressa®) was purchased from
AstraZeneca plc. (Cambridge, UK). The human NSCLC cell line HCC827,
which is sensitive to gefitinib and referred to in the current
study as H827S, containing an EGFR exon19 deletion (DelE746-A750)
was obtained from the Cell Resource Center of the Shanghai
Institutes for Biological Sciences of the Chinese Academy of
Sciences (Shanghai, China). A gefitinib-resistant cell line (H827R)
was generated in the laboratory and individual H827R-7-1 clones
were isolated from single cell clone of parental H827R cells using
the method of maximum dose dilution as described previously
(11).
Cells were cultured in RPMI-1640 medium (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 10%
exosome-depleted fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 50 µmol/l
β-mercaptoethanol (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
All cell cultures were maintained in an incubator containing 5%
CO2 at 37°C. The resistant HCC827 cells were passed ≤15
times in the absence of gefitinib to maintain their resistance.
Exosome isolation and
confirmation
Exosomes from the supernatants of H827S and
H827R-7-1 cells were designated as S/exo and R/exo, respectively.
The cell supernatant was centrifuged at 1,000 × g (10 min), 10,000
× g (30 min at 4°C), and 100,000 × g (120 min at 4°C) using a
Beckman ultracentrifuge (AvantiJ-30I; Beckman Coulter, Inc., Brea,
CA, USA). The final pellets were resuspended in ExoQuick-TC exosome
precipitation solution (System Biosciences, Mountain View, CA, USA)
overnight at 4°C and were centrifuged at 1,500 × g (30 min at 4°C).
The extracted pellets were diluted in 200 µl of PBS and stored at
−80°C. Exosomes were observed using a JEM-2100 transmission
electron microscope (TEM; JEOL, Ltd., Tokyo, Japan). Exosomes were
quantified using an enhanced BCA protein assay kit (Beyotime
Institute of Biotechnology, Haimen, China).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Exosomal RNAs were isolated using a
mirVana™ miRNA Isolation kit (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol. Total
cellular RNA was extracted using TRIzol (Life Sciences; Thermo
Fisher Scientific, Inc.).
The Bulge-Loop™ hsa-miR-21 RT-qPCR primer
set and U6 small nuclear (sn)RNA RT-qPCR primer sets (both 100T)
were purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
miRNA was stem-loop reverse transcribed using a
PrimeScript™ 1st Strand cDNA Synthesis kit (Takara Bio,
Inc., Otsu, Japan). qPCR was performed using a TaqMan™
miRNA assay kit according to the manufacturer's protocol on an ABI
7300 detection system (Applied Biosystems, Foster City, CA, USA). A
gene-specific probe mix (miR-21, 000397) was utilized. U6 was used
as the internal control. The 2−ΔΔCt method was used to
determine the fold change of miR-21 (12).
Experimental groups
To investigate whether R/exo induced
gefitinib-resistance in susceptible cells, 6 groups were prepared
as follows: Control (H827S without exosome and gefitinib
treatment), H827S+gefitinib (H827S incubated with 50 nM gefitinib),
H827S+S/exo (H827S pretreated with S/exo for 24 h),
H827S+S/exo+gefitinib (H827S pretreated with S/exo for 24 h and
incubated with 50 nM gefitinib for a further 48 h), H827S+R/exo
(H827S pretreated with R/exo for 24 h), H827S+R/exo+gefitinib
(H827S pretreated with R/exo for 24 h and incubated with 50 nM
gefitinib for a further 48 h). The viability and apoptotic rates of
cells in each group were measured following the aforementioned
treatments to assess the effect of R/exo on the cells.
Transfection
The Hsa-miR-21 inhibitor,
5′-UCAACAUCA-GUCUGAUAAGCUA-3′ and the negative control (NC),
5′-CAGUACUUUUG-UGUAGUACAA-3′ small interfering (si)RNAs were
obtained from Shanghai GenePharma Co., Ltd. (Shanghai, China).
The following groups were designed to assess the
effect of miR-21 on exosome mediated drug resistance: H827S+NC
siRNA+S/exo (H827S transfected with NC siRNA and pretreated with
S/exo for 24 h without gefitinib treatment), H827S+NC
siRNA+S/exo+gefitinib (H827S transfected with NC siRNA, pretreated
with S/exo for 24 h and stimulated with 50 nM gefitinib for a
further 48 h), H827S+NC siRNA+R/exo+gefitinib (H827S transfected
with NC siRNA, pretreated with R/exo for 24 h and stimulated with
50 nM gefitinib for a further 48 h), H827S+miR-21
siRNA+R/exo+gefitinib (H827S transfected with miR-21 siRNA,
pretreated with R/exo for 24 h and stimulated with gefitinib for a
further 48 h).
Oligonucleotides (50 nM) were transfected into H827S
cells that had reached 70% confluence using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol. Following
24 h transfection, R/exo or S/exo were added to the corresponding
wells. Following another 24 h, cells were exposed to 0 or 50 nM
gefitinib for 48 h at 37°C prior to being harvested for cell
viability assay and flow cytometry analysis, following the
protocols described below.
Cell viability assay
H827S cells were plated into 96-well plates at a
density of ~5,000 cells/well. Following 24 h incubation, S/exo or
R/exo were added to the cells for 24 h at 37°C. The same volume of
PBS without exosomes was added to cells in the control group. Cells
were exposed to 0 or 50 nM gefitinib for 48 h at 37°C and cell
viability was subsequently assessed using a cell counting kit-8
(CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
according to the manufacturer's protocol.
Sensitivity test
Growth inhibition was measured by CCK-8 assay.
Briefly, the cells were plated onto 96-well plates at a density of
approximately 5,000 cells per well and exposed to a concentration
gradient of gefitinib (0, 10, 50, 100, 500 nM, 1, 5 and 10 mM) for
48 h. This test was carried out to confirm the successful
establishment of H827R.
Flow cytometry
H827S cells were seeded and cultured in 6-well
plates at a concentration of 1×106 cells/well. H827S
cells were incubated with S/exo or R/exo for 24 h and subsequently
treated with either 0 or 50 nM gefitinib for a further 48 h at
37°C. The cells were then stained using a fluorescein
isothiocyanate Annexin V apoptosis detection kit (BD Biosciences,
Franklin Lakes, NJ, USA) and BD Accuri C6 flow cytometer (BD
Biosciences) and BD Accuri C6 Software (version 1.1.264.21; BD
Biosciences) was used to discriminate apoptotic cells. At least
10,000 cells were analyzed for each group.
Western blot analysis
Harvested cells were lysed with an ice-cold lysis
buffer containing 50 mmol/l Tris-HCl (pH 7.4); 1% NP-40; 150 mmol/l
NaCl; 1 mmol/l EDTA; 1 mmol/l phenylmethylsulfonyl fluoride and
complete proteinase inhibitor mixture (one tablet per 10 ml; Roche
Molecular Diagnostics, Pleasanton, CA, USA). Protein content was
determined using a DC Protein Assay kit (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) and western blot analysis was performed
following a previously described protocol (10). Individual immunoblots were probed with
antibodies against phosphorylated (p)-protein kinase B (Akt;
Ser473; D9E) XP® rabbit mAb (cat. no. 4060S; dilution
1:1,000), Akt (pan; C67E7) rabbit mAb (cat. no. 4691S; dilution
1:1,000; both Cell Signaling Technology, Inc., Danvers, MA, USA)
and β-actin polyclonal antibodies (cat. no. AP0060; dilution
1:2,000; Bioworld Technology, Inc., St. Louis Park, MN, USA) at 4°C
overnight and finally with a horseradish peroxidase-conjugated
antibody (cat no. 7074S; dilution 1:2,000; Cell Signaling
Technology, Inc.) at room temperature for 1 h. Proteins were
visualized using an enhanced chemiluminescence solution (Life
Sciences; Thermo Fisher Scientific, Inc.).
Statistical analysis
Statistical analysis was performed with SPSS
software (version 20.0; IBM Corp., Armonk, NY, USA). Comparisons
between pairs were performed using a Student's t-test and multiple
comparisons between the groups were analyzed using one-way analysis
of variance followed by a Student Newman-Keuls test. All
experiments were performed three times and the results are
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Exosomes contribute to gefitinib
resistance in NSCLC H827S cells
The inhibitory concentrations for 50% cell death
(IC50) values of gefitinib were >10 µM for H827R and
0.05 µM for H827S, thus identifying gefitnib resistance in H827R
cells (data not shown). Following exosome isolation from the cell
supernatant of H827S and H827R cells, residual vesicles were
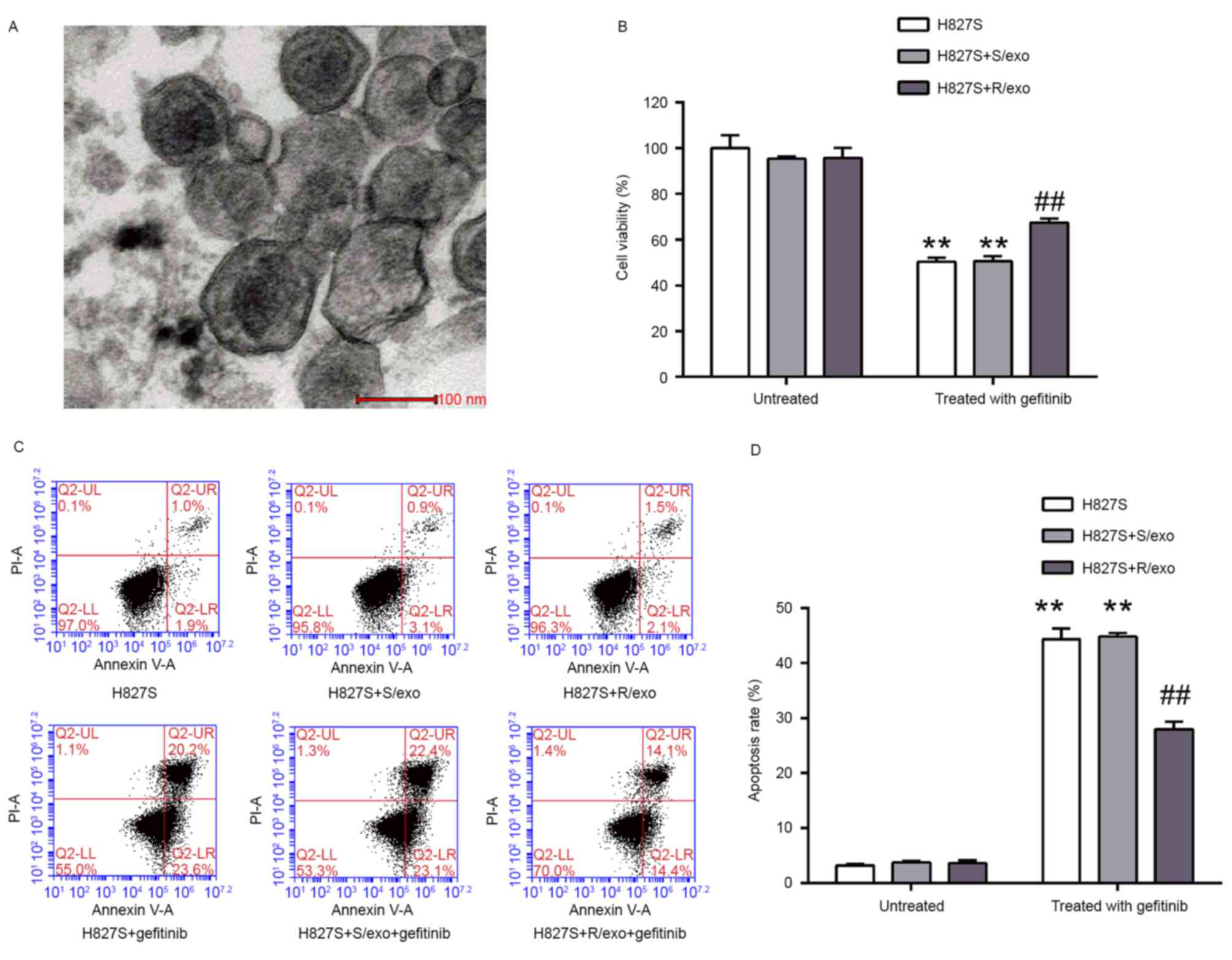
observed using TEM (Fig. 1A). The
diameter of the vesicles was <100 nm, indicating that exosomes
were successfully extracted.
Compared with the control group (100%), cell
viability was 95.3±1.1, 95.7±4.5, 50.4±1.9, 50.7±2.1 and 67.5±1.8%
in the H827S+S/exo, H827S+R/exo, H827S+gefitinib,
H827S+S/exo+gefitinib and H827S+R/exo+gefitinib groups,
respectively (Fig. 1B). Pretreatment
with R/exo significantly increased H827S cell viability following
gefitinib stimulation compared with untreated and S/exo treated
H827A cells. Cells were analyzed using flow cytometry (Fig. 1C) and the results of the apoptosis
assay were similar to those of the CCK-8 assay. The apoptosis rates
were 3.2±0.3, 3.7±0.3, 3.6±0.5, 44.3±2.0, 44.8±0.7 and 27.9±1.4% in
the H827S, H827S+S/exo, H827S+R/exo, H827S+gefitinib,
H827S+S/exo+gefitinib and H827S+R/exo+gefitinib groups,
respectively (Fig. 1D). Treatment
with R/exo prior to gefitinib exposure significantly decreased the
sensitivity of H827S cells to gefitinib compared with the cells
pretreated with S/exo. Cell viability and rates of apoptosis did
not differ significantly between the H827S and H827S+S/exo groups.
These results indicate that exosomes released by H827R cells may
decrease the sensitivity of H827S cells to gefitinib, whereas
exosomes released by H827S cells have little influence on the
sensitivity of H827S cells to gefitinib.
miR-21 expression is increased in
R/exo and R/exo-treated H827S cells
miRNAs are a class of small non-coding RNAs 18–25
nucleotides in length; these molecules post-transcriptionally
inhibit gene expression by inducing degradation or blocking
translation of miR targets (13).
miR-21 is overexpressed in several human malignancies and has been
implicated in various biological processes, including cell
proliferation, apoptosis, invasion and metastasis (14). Evidence has emerged regarding the role
of miR-21 in the regulation of drug resistance (15–17) and
miR-21 overexpression is associated with the development of
gefitinib resistance in NSCLC (18).
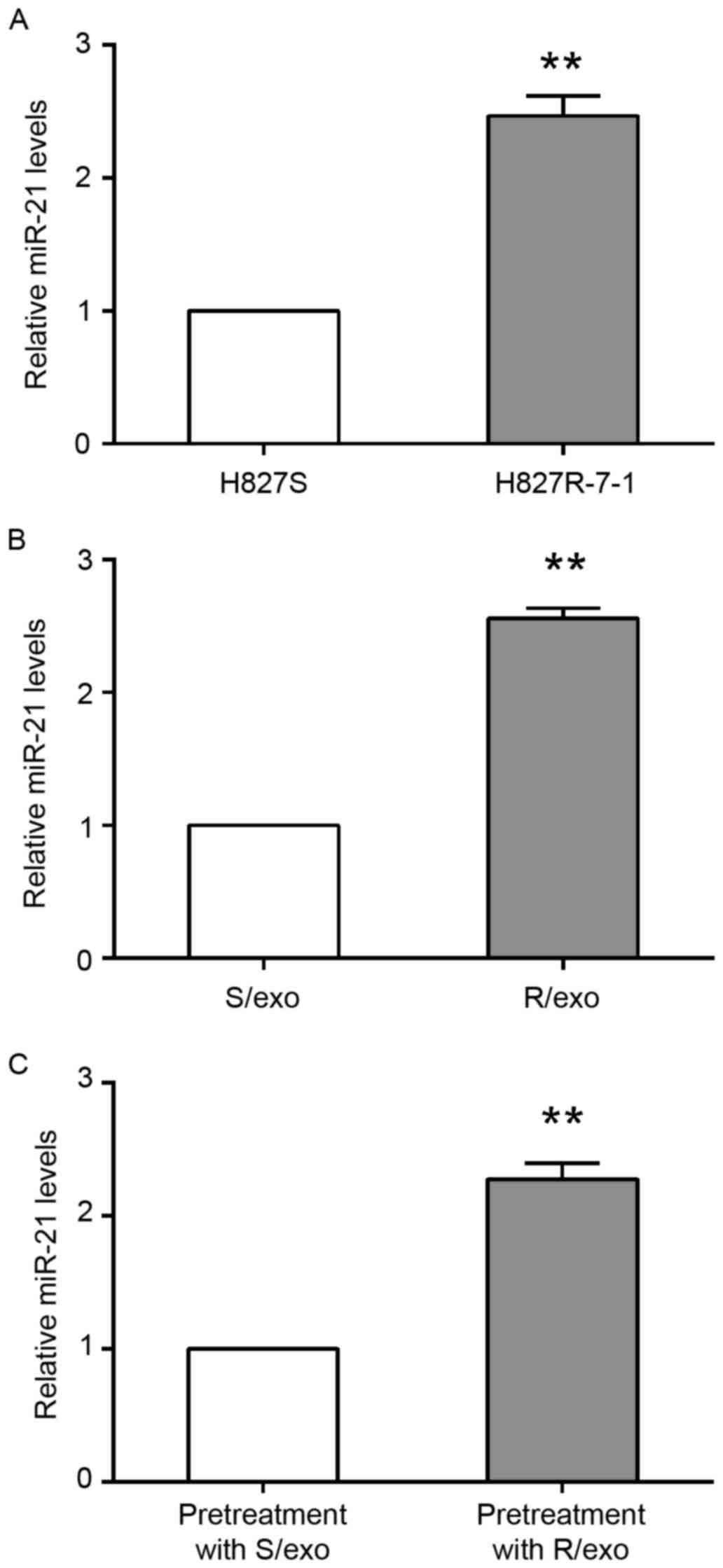
To determine the involvement of miR-21 in exosome-mediated
gefitinib resistance, RT-qPCR was performed to measure the
expression of miR-21 in H827 cells. H827S and H827R-7-1 cells were
seeded and cultured in 6-well plates for 48 h. Cells then underwent
RT-qPCR and the results demonstrated that miR-21 expression was
significantly increased in H827R-7-1 cells by 2.46±0.15-fold
compared with H827S cells (Fig.
2A).
For exosome isolation, H827S and H827R-7-1 cells
were seeded in 15-cm plates for 48 h and exosomes were subsequently
obtained. The H827S cells were incubated with either S/exo or R/exo
for 24 h. miR-21 expression was significantly increased in the
R/exo group by 2.56±0.08-fold compared with the S/exo group
(Fig. 2B). In addition, miR-21
expression was significantly increased in R/exo-treated H827S cells
by 2.28±0.12-fold compared with H827S cells pretreated with S/exo
(Fig. 2C).
miR-21 inhibition attenuates
exosome-mediated drug resistance
To elucidate the role of miR-21 in the ability of
R/exo to increase cell viability and inhibit cell apoptosis in
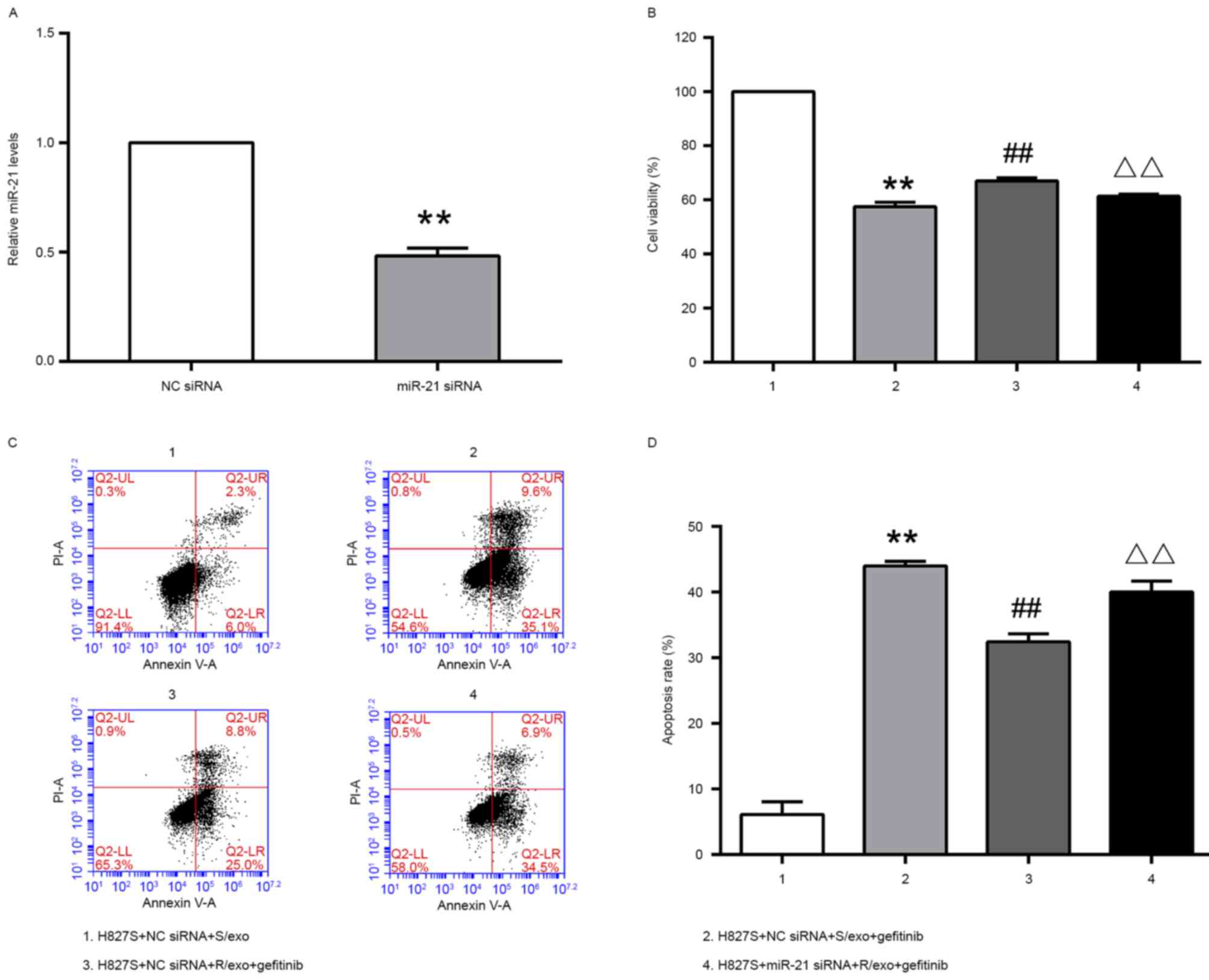
NSCLC cells, miR-21 expression was inhibited (Fig. 3). H827S cells were transfected with
anti-miR-21 or NC siRNA. Following 48 h treatment, miR-21
expression was measured using RT-qPCR. The results demonstrated
that miR-21 expression was significantly downregulated in cells
transfected with the miR-21 siRNA compared with cells transfected
with NC siRNA (Fig. 3A).
As presented in Fig.
3B, compared to H827S+NC siRNA+S/exo group (100%), cell
viability was decreased in H827S+NC siRNA+S/exo+gefitinib group
(57.5±1.6%). However, H827S+NC siRNA+R/exo+gefitinib group had
elevated cell viability (67.0±1.1%), compared to H827S+NC
siRNA+S/exo+gefitinib group. Following miR-21 knockdown, drug
resistance was partially reversed; cell viability decreased in
H827S cells pretreated with R/exo and stimulated with gefitinib
(61.3±0.9%).
The result of apoptosis assay (Fig. 3C and D) was similar with result of
CCK-8 experiment. H827S+NC siRNA+S/exo+gefitinib group exhibited
greater apoptosis (44.0±0.70%) compared with H827S+NC siRNA+S/exo
group (6.1±2.0%). H827S+NC siRNA+R/exo+gefitinib group had a
significantly lower apoptosis rate (32.4±1.21%). Transfection of
the miR-21 inhibitor sensitized H827S cells that were pretreated
with R/exo and underwent stimulation with gefitinib, with an
increased apoptosis rate (40.0±1.7%).
Exosomes transmit resistance to
gefitinib by activating the Akt signaling pathway
Akt is an important downstream signaling pathway of
EGFR and Akt activation is associated with the prognosis of
patients with lung cancer (19). To
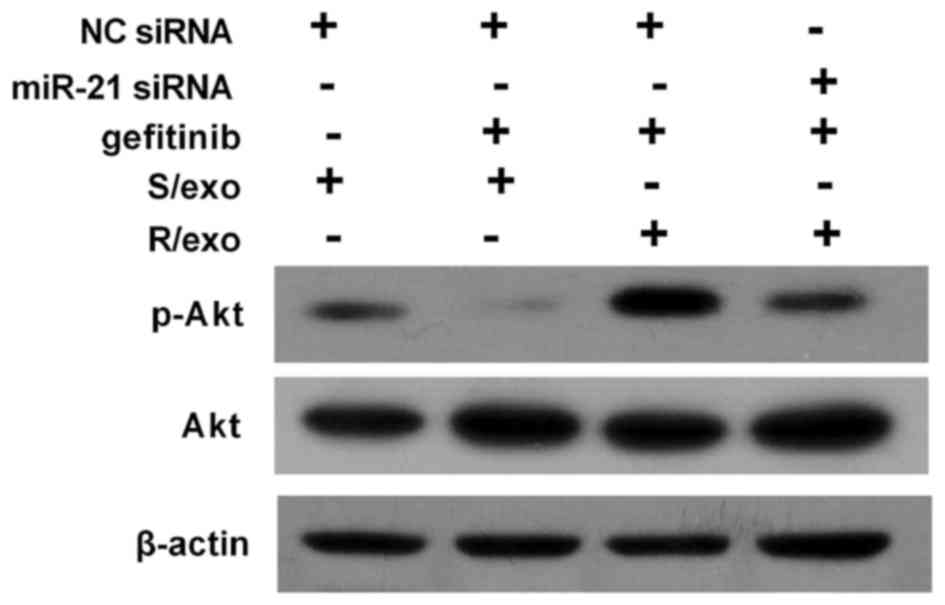
further investigate the underlying mechanism of gefitinib
resistance transmitted by R/exo, the expression of p-Akt was
measured using western blot analysis (Fig. 4). Gefitinib treatment notably
downregulated p-Akt expression in the S/exo treated group compared
with the control. However, R/exo pretreatment elevated p-Akt levels
and promoted Akt activation compared with the S/exo treated group.
However, miR-21 inhibition reduced p-Akt expression and blocked Akt
activation in cells pretreated with R/exo.
Discussion
Gefitinib is a major chemotherapeutic agent used to
treat lung cancer and significantly improves the overall survival
rate of patients with cancer that harbor somatic mutations in the
EGFR gene (2). However, acquired
resistance to gefitinib remains a major problem in cancer treatment
and limits treatment efficacy. At present, the underlying
mechanisms of acquired resistance to gefitinib are not fully
understood.
Exosomes have emerged as important messengers of
cellular communication in normal physiological processes and
diseases, including liver and neurodegenerative diseases, as well
as the development and progression of cancer (20,21). In
addition, studies have identified a close association between
exosomes and drug resistance in various types of cancer, including
breast (9,22), ovarian (23) and prostate (24) cancer, which suggests that drug
resistance in lung cancer cells may be acquired via exosomes.
Cisplatin exposure may increase exosome secretion and the
interaction of these secreted exosomes with other cancer cells may
increase the resistance of A549 cells to cisplatin (10). Exosomes derived from gefitinib-treated
lung cancer cells decreased the anti-tumor effects of cisplatin,
whereas exosomes derived from cisplatin-treated lung cancer cells
did not significantly alter the antitumor effects of gefitinib
(25). These results suggest that the
inhibition of exosome secretion may be a helpful strategy to
overcome drug resistance. In he present study, cell viability was
decreased in the H827S+NC siRNA+S/exo+gefitinib group, indicating
the inhibitory effect of gefitinib. The H827S+NC
siRNA+R/exo+gefitinib group exhibited elevated cell viability and a
decreased apoptosis rate compared to the H827S+NC
siRNA+S/exo+gefitinib group, suggesting that gefitinib resistance
in lung cancer cells may be acquired via exosomes. To the best of
our knowledge, the present study was the first to demonstrate that
exosomes from gefitinib-resistant cells confer drug resistance in
lung cancer cells, as it was observed that R/exo-treated H827S
cells lost their sensitivity to gefitinib.
Exosomes transfer RNAs and proteins to mediate
communication among cancer cells and exosome-mediated miRNA
transfer may be a novel method of gene transfer among cells
(26). Previous in vitro and
in vivo studies have indicated that miR-21 is frequently
overexpressed in various human tumors and in cancer cell lines, and
promotes oncogenesis, suggesting that it is an onco-miR (27,28).
Additionally, the association between miR-21 and drug resistance
has been investigated and high levels of miR-21 were detected in
5-fluorouracil-resistant human pancreatic, cisplatin-resistant
ovarian, doxorubicin-resistant breast and cisplatin-resistant
neuroblastoma cancer cells (15,29–31). In a
previous study, 20 patients with advanced NSCLC with the EGFR 19
deletion were treated with first-line EGFR-TKIs and divided into
two groups: A EGFR-TKI-resistant group and a EGFR-TKI-sensitive
group (32). The expression of plasma
miR-21 was significantly higher in the EGFR-TKI resistant group
compared with the EGFR-TKI-sensitive group as determined by a
TaqMan low-density array (32). Li
et al reported that miR-21 was overexpressed in the EGFR-TKI
resistant cell line PC9R compared with the PC9 non-resistant cell
line, that miR-21 expression was negatively associated with the
expression of PTEN, and that programmed cell death protein 4 was
positively associated with the phosphatidylinositol 3-kinase
(PI3K)/Akt signaling pathway (18).
Additionally, the inhibition of miR-21 induced apoptosis in the
PC9R cell line and inhibited miR-21, while miR-21 antisense oligo
nucleotide (ASO) suppressed tumor growth in nude mice treated with
EGFR-TKI (18). Shen et al
(33) revealed that high miR-21
expression indicated a poor TKI clinical response and a shorter
overall survival rate. miR-21 expression was upregulated in PC-9
gefitinib resistant cells (PC-9/GR) compared with PC-9 cells and
miR-21 knockdown markedly restored gefitinib sensitivity in PC-9/GR
cells (33). Similar to previous
studies, the results of the present investigation revealed that
miR-21 expression was elevated in gefitinib-resistant lung cancer
cells. miR-21 expression was increased in exosomes from H827R-7-1
cells and R/exo-stimulated H827S cells. It was considered that
miR-21 in H827R-7-1 was released into exosomes and transported into
H827S cells via exosomes, thereby changing the sensitivity of H827S
cells to gefitinib. Following miR-21 knockdown, drug resistance was
partially reversed; cell viability decreased and apoptosis rate
increased in H827S cells pretreated with R/exo and stimulated with
gefitinib. These results indicate that miR-21 serves a crucial
function in gefitinib resistance inferred by exosomes.
Akt is a downstream mediator of PI3K, which serves a
central role in tumorigenesis. EGFR-TKIs primarily inhibit the
downstream signaling pathway activity of EGFR via the PI3K/Akt
signaling pathway, thereby inhibiting cell proliferation and
invasion, as well as inducing apoptosis (34). The suppression of the Akt signaling
pathway may preserve gefitinib resistance in NSCLC cell lines
(35). It has been demonstrated that
miR-21 positively regulates the PI3K/Akt signaling pathway
(18,36,37).
miR-21 suppresses tumor cell migration and invasion by reducing
PI3K/Akt signaling and reversing the epithelial-mesenchymal
transition in breast cancer (36).
miR-21 also modulates the radiosensitivity of cervical cancer by
inhibiting autophagy via the Akt-mTOR signaling pathway (37). Furthermore, miR-21 induces Akt
phosphorylation and activates the Akt signaling pathway, thereby
leading to the acquired resistance to EGFR-TKIs in the NSCLC cell
lines (18). In the present study,
p-Akt expression was downregulated by gefitinib treatment and
miR-21 silencing in H827S cells, suggesting that miR-21 knockdown
may ameliorate the exosome-mediated activation of Akt and increase
the sensitivity of H827S cells to gefitinib.
In conclusion, the results of the present study
revealed that exosomes released by gefitinib-resistant lung cancer
cells decreased the sensitivity of gefitinib-sensitive cells. The
present study indicates that miR-21 is a critical mediator in
exosome-induced drug resistance. The inhibition of exosome
formation and release may be developed as a novel therapeutic
strategy of addressing gefitinib resistance in patients with lung
cancer in the future.
Acknowledgements
The present study was supported by the general
program of the Department of Health in Jiangsu province (grant no.
H201410), the National Natural Science Foundation of China (grant
no. 81402483) and by the Natural Science Foundation of Jiangsu
Province (grant no. BK20141016).
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maemondo M, Inoue A, Kobayashi K, Sugawara
S, Oizumi S, Isobe H, Gemma A, Harada M, Yoshizawa H, Kinoshita I,
et al: Gefitinib or chemotherapy for non-small-cell lung cancer
with mutated EGFR. N Engl J Med. 362:2380–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thatcher N, Chang A, Parikh P, Pereira
Rodrigues J, Ciuleanu T, von Pawel J, Thongprasert S, Tan EH,
Pemberton K, Archer V and Carroll K: Gefitinib plus best supportive
care in previously treated patients with refractory advanced
non-small-cell lung cancer: Results from a randomised,
placebo-controlled, multicentre study (Iressa Survival Evaluation
in Lung Cancer). Lancet. 366:1527–1537. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mok TS, Wu YL, Thongprasert S, Yang CH,
Chu DT, Saijo N, Sunpaweravong P, Han B, Margono B, Ichinose Y, et
al: Gefitinib or carboplatin-paclitaxel in pulmonary
adenocarcinoma. N Engl J Med. 361:947–957. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suda K, Murakami I, Katayama T, Tomizawa
K, Osada H, Sekido Y, Maehara Y, Yatabe Y and Mitsudomi T:
Reciprocal and complementary role of MET amplification and EGFR
T790M mutation in acquired resistance to kinase inhibitors in lung
cancer. Clin Cancer Res. 16:5489–5498. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
van der Pol E, Böing AN, Harrison P, Sturk
A and Nieuwland R: Classification, functions, and clinical
relevance of extracellular vesicles. Pharmacol Rev. 64:676–705.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katakowski M, Buller B, Zheng X, Lu Y,
Rogers T, Osobamiro O, Shu W, Jiang F and Chopp M: Exosomes from
marrow stromal cells expressing miR-146b inhibit glioma growth.
Cancer Lett. 335:201–204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vlassov AV, Magdaleno S, Setterquist R and
Conrad R: Exosomes: Current knowledge of their composition,
biological functions, and diagnostic and therapeutic potentials.
Biochim Biophys Acta. 1820:940–948. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lv MM, Zhu XY, Chen WX, Zhong SL, Hu Q, Ma
TF, Zhang J, Chen L, Tang JH and Zhao JH: Exosomes mediate drug
resistance transfer in MCF-7 breast cancer cells and a probable
mechanism is delivery of P-glycoprotein. Tumour Biol.
35:10773–10779. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao X, Yu S, Li S, Wu J, Ma R, Cao H, Zhu
Y and Feng J: Exosomes: Decreased sensitivity of lung cancer A549
cells to cisplatin. PloS One. 9:e895342014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu SW, Cao HX, Wu JZ, Ma R, Jing CW, Wang
Z and Feng JF: Difference of miRNA-7 expression in the non-small
cell lung cancer with different sensitivity to gefitinib and their
clinical significance. Chin J Surg Onco. 7:74–77. 2015.
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li S, Liang Z, Xu L and Zou F:
MicroRNA-21: A ubiquitously expressed pro-survival factor in cancer
and other diseases. Mol Cell Biochem. 360:147–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang ZX, Lu BB, Wang H, Cheng ZX and Yin
YM: MicroRNA-21 modulates chemosensitivity of breast cancer cells
to doxorubicin by targeting PTEN. Arch Med Res. 42:281–290. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang SM, Huang C, Li XF, Yu MZ, He Y and
Li J: miR-21 confers cisplatin resistance in gastric cancer cells
by regulating PTEN. Toxicology. 306:162–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ziyan W and Yang L: MicroRNA-21 regulates
the sensitivity to cisplatin in a human osteosarcoma cell line. Ir
J Med Sci. 185:85–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li B, Ren S, Li X, Wang Y, Garfield D,
Zhou S, Chen X, Su C, Chen M, Kuang P, et al: MiR-21 overexpression
is associated with acquired resistance of EGFR-TKI in non-small
cell lung cancer. Lung Cancer. 83:146–153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang JM, He QY, Guo RX and Chang XJ:
Phosphorylated Akt overexpression and loss of PTEN expression in
non-small cell lung cancer confers poor prognosis. Lung Cancer.
51:181–191. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masyuk AI, Masyuk TV and Larusso NF:
Exosomes in the pathogenesis, diagnostics and therapeutics of liver
diseases. J Hepatol. 59:621–625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vella LJ, Sharples RA, Nisbet RM, Cappai R
and Hill AF: The role of exosomes in the processing of proteins
associated with neurodegenerative diseases. Eur Biophys J.
37:323–332. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu DD, Wu Y, Zhang XH, Lv MM, Chen WX,
Chen X, Yang SJ, Shen H, Zhong SL, Tang JH and Zhao JH: Exosomes
from adriamycin-resistant breast cancer cells transmit drug
resistance partly by delivering miR-222. Tumour Biol. 37:3227–3235.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yeung Au CL, Co NN, Tsuruga T, Yeung TL,
Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, et al: Exosomal
transfer of stroma-derived miR21 confers paclitaxel resistance in
ovarian cancer cells through targeting APAF1. Nat Commun.
7:111502016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Corcoran C, Rani S, O'Brien K, O'Neill A,
Prencipe M, Sheikh R, Webb G, McDermott R, Watson W, Crown J and
O'Driscoll L: Docetaxel-resistance in prostate cancer: Evaluating
associated phenotypic changes and potential for resistance transfer
via exosomes. PloS One. 7:e509992012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li XQ, Liu JT, Fan LL, Liu Y, Cheng L,
Wang F, Yu HQ, Gao J, Wei W, Wang H and Sun GP: Exosomes derived
from gefitinib-treated EGFR-mutant lung cancer cells alter
cisplatin sensitivity via up-regulating autophagy. Oncotarget.
7:24585–24595. 2016.PubMed/NCBI
|
|
26
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Capodanno A, Boldrini L, Proietti A, Alì
G, Pelliccioni S, Niccoli C, D'Incecco A, Cappuzzo F, Chella A,
Lucchi M, et al: Let-7g and miR-21 expression in non-small cell
lung cancer: Correlation with clinicopathological and molecular
features. Int J Oncol. 43:765–774. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radioresistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wei X, Wang W, Wang L, Zhang Y, Zhang X,
Chen M, Wang F, Yu J, Ma Y and Sun G: MicroRNA-21 induces
5-fluorouracil resistance in human pancreatic cancer cells by
regulating PTEN and PDCD4. Cancer Med. 5:693–702. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Echevarría-Vargas IM, Valiyeva F and
Vívas-Mejia PE: Upregulation of miR-21 in cisplatin resistant
ovarian cancer via JNK-1/c-Jun pathway. PloS One. 9:e970942014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen Y, Tsai YH, Fang Y and Tseng SH:
Micro-RNA-21 regulates the sensitivity to cisplatin in human
neuroblastoma cells. J Pediatr Surg. 47:1797–1805. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang S, Su X, Bai H, Zhao J, Duan J, An T,
Zhuo M, Wang Z, Wu M, Li Z, et al: Identification of plasma
microRNA profiles for primary resistance to EGFR-TKIs in advanced
non-small cell lung cancer (NSCLC) patients with EGFR activating
mutation. J Hematol Oncol. 8:1272015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen H, Zhu F, Liu J, Xu T, Pei D, Wang R,
Qian Y, Li Q, Wang L, Shi Z, et al: Alteration in Mir-21/PTEN
expression modulates gefitinib resistance in non-small cell lung
cancer. PloS One. 9:e1033052014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang F, Lee JT, Navolanic PM, Steelman
LS, Shelton JG, Blalock WL, Franklin RA and McCubrey JA:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: A target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li H, Schmid-Bindert G, Wang D, Zhao Y,
Yang X, Su B and Zhou C: Blocking the PI3K/AKT and MEK/ERK
signaling pathways can overcome gefitinib-resistance in non-small
cell lung cancer cell lines. Adv Med Sci. 56:275–284. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yan LX, Liu YH, Xiang JW, Wu QN, Xu LB,
Luo XL, Zhu XL, Liu C, Xu FP, Luo DL, et al: PIK3R1 targeting by
miR-21 suppresses tumor cell migration and invasion by reducing
PI3K/AKT signaling and reversing EMT, and predicts clinical outcome
of breast cancer. Int J Oncol. 48:471–484. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song L, Liu S, Zhang L, Yao H, Gao F, Xu D
and Li Q: MiR-21 modulates radiosensitivity of cervical cancer
through inhibiting autophagy via the PTEN/Akt/HIF-1α feedback loop
and the Akt-mTOR signaling pathway. Tumour Biol. 37:12161–12168.
2016. View Article : Google Scholar : PubMed/NCBI
|