Introduction
Osteosarcoma is a commonly occurring primary
malignant bone tumor in children and young adolescents, with a
secondary incidence peak in individuals who are ≥50 years old
(1). Clinically, the incidence of
osteosarcoma is ~1–3 cases/million individuals (2). Osteosarcoma most commonly occurs in the
long tubular bones, including the femur, tibia and humerus and,
less commonly, in the skull, jaw or pelvis; ≤1% of osteosarcoma
cases are localized in the hands or feet (3). Similar to the majority of cancer types,
osteosarcoma development is a multistep process and the precise
etiology and pathogenesis remain to be determined, although a
previous study indicated that ionizing radiation, with or without
chemotherapy, was associated with a higher risk of developing
osteosarcoma (4). Despite amputation
having been frequently utilized over previous years in order to
treat patients with osteosarcoma, therapeutic methods or
alternative methods to treat osteosarcoma remained limited
(1). This did not change until the
early 1970s, and with the introduction of adjuvant chemotherapy, in
particular the use of high-dose methotrexate, facilitating the
surgical resection of patients with osteosarcoma has become
possible (5). Advancement in the
early detection and treatment of osteosarcoma has been made in
previous years; however, the survival rate of patients with
osteosarcoma has remained virtually unchanged over the last 30
years (5). Previous studies revealed
that the five-year survival rate remains at ~20% (5,6). Thus,
more specific biomarkers for the early diagnosis and targeted
therapy of osteosarcoma are required.
One approach for the development of targeted therapy
and specific biomarkers is to focus on microRNAs (miRNA/miR), which
are a class of small non-coding RNAs ~17–24 nucleotides in length
(7). miRNAs are able to regulate the
expression level of target genes by base-pairing to the
3′-untranslated region (3′-UTR) of mRNA, thereby repressing protein
translation or inducing mRNA degradation. For example, Friedman
et al (8) demonstrated that
~60% of human genes are regulated by manifold miRNAs in a
cooperative manner. Accumulating evidence has suggested that
dysregulated miRNA expression may contribute to cancer development
and progression (9). The elucidation
of miRNA expression and function in human cancer is important for
the understanding of cancer pathogenesis, as miRNAs may affect cell
proliferation, differentiation, metastasis, apoptosis and
sensitivity to chemotherapy and radiotherapy (10). Given their pleiotropic actions to
repress multiple target genes simultaneously (11), miRNAs may be ideal candidates to
provide a comprehensive understanding of osteosarcoma pathogenesis.
Thus, the present study investigated a particular miRNA, miR-130b,
in paired osteosarcoma and adjacent normal tissues and cell lines.
The miR-130b gene is localized at chromosome 22q11, and has been
proposed as a novel tumor-associated miRNA (12). Previous studies have demonstrated
altered miR-130b expression in promoting or suppressing
tumorigenesis; for example, in papillary thyroid carcinomas
(13), endometrial cancer (14) and pancreatic cancer (15), miR-130b was downregulated. However,
mR-130b expression was upregulated in melanoma (16), bladder cancer (17) and metastatic renal carcinoma (18). A previous study revealed an
association between miR-130 expression levels and osteosarcoma cell
lines using microarray profiling (19); however, the molecular pathways by
which miR-130b modulates osteosarcoma development and progression
have not been elucidated. The present study also investigated the
effects of miR-130b expression in the regulation of osteosarcoma
cell proliferation, colony formation, migration and invasion in
vitro. Bioinformatic analysis and dual-luciferase reporter
assays were subsequently performed in order to identify the target
gene. The present study aimed to provide an improved understanding
of the effects and function of miR-130b in osteosarcoma, which may
help to develop a novel therapeutic strategy for managing
osteosarcoma.
Materials and methods
Tissues
Osteosarcoma and paired normal tissues from 13
patients were collected between January 2013 and February 2015 from
the Department of Surgery, The Second Xiangya Hospital, Central
South University (Changsha, China). The average of age was 13
(range, 9–19); 6 patients were female and 7 were male. All patients
were histologically diagnosed and verified according to the 2013
World Health Organization classification guidelines (20). None of the patients had received
chemotherapy or radiotherapy prior to surgery. All tissues were
obtained via the surgical resection, snap-frozen in liquid nitrogen
and stored at −80°C. The present study was approved by the Human
Ethics Committee of the Xiangya Medical School of the Central South
University. Written informed consent was obtained from all patients
prior to enrollment in the present study.
Cell lines and culture
U-2OS, Saos-2 and MG-63 human osteosarcoma cell
lines, hFOB normal human osteoblast cells and HEK293T cells were
purchased from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China), and cultured in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% heat-inactivated fetal bovine
serum (FBS), 100 U/ml penicillin and 100 µg/ml streptomycin (all
from Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
at 37°C in a humidified atmosphere of 5% CO2 and 95%
air.
Oligonucleotides, plasmids and cell
transfection
A total of four oligonucleotides (miRNA-130b mimics,
miRNA-130b mimics negative control (mimics NC), miRNA-130b
inhibitor, miRNA-130b inhibitor negative control (inhibitor NC)
were chemically synthesized by Genepharm, Inc. (Sunnyvale, CA,
USA). The open reading frame of human transforming growth factor α
(TGFα; Gen-Bank Accession no., KJ892270.1) was cloned into pcDNA3.1
(+; Invitrogen; Thermo Fisher Scientific, Inc.) and the DNA
sequence was confirmed by PCR kit (PCR master mix; cat. no. K0171;
Invitrogen; Thermo Fisher Scientific, Inc.). The primers for TGFα
were: Forward, 5′-AGCAGTGGTGTCCCATTTTAA-3′ and reverse,
5′-AGCGGTTCTTCCCTTCAGG-3′. The thermocycling conditions were as
follows: Initial denaturation 95°C 1 min, 30 of cycles of
denaturation 95°C 30 sec, annealing 55°C 30 sec and elongation 72°C
1 min; and final extension 72°C 10 min. For gene transfection, the
oligonucleotides and plasmids were mixed with
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions,
and then added to the cell culture and 37°C incubated for 6 h prior
to the growth medium being changed, and subsequently 37°C incubated
for an additional 42 h.
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
Total RNA was isolated from the tissues and cultured
cells using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.), according to manufacturer's instructions, and
quantified using a NanoDrop™ 2000 spectrophotometer
(Thermo Fisher Scientific, Inc.). In order to detect the expression
level of miRNA-130b in tissues and cells, cDNA was reverse
transcribed using a TaqMan® miRNA Reverse Transcription
kit (Applied Biosystems; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol, and subsequently subjected to
RT-qPCR using a SYBR® Premix Ex Taq™ kit (Takara
Biotechnology, Co., Ltd., Dalian, China). The U6 transcript was
used as an internal control. The level of TGFA mRNA was determined
using an ABI Prism® 7900 Sequence Detector system
(Applied Biosystems; Thermo Fisher Scientific, Inc.), and GAPDH was
used as an internal control. miR-103b and TGFA mRNA expression
levels were determined using the 2−ΔΔCq method and
compared as the relative fold changes of internal controls
(21).
Dual-luciferase reporter assay
A fragment of the 3′-UTR of miR-130b containing the
putative binding sites (3344–3351) to TGFA was subcloned into a
firefly luciferase pGL3-control vector (Invitrogen; Thermo Fisher
Scientific, Inc.). All constructs were verified by by PCR kit (PCR
master mix; cat. no. K0171; Invitrogen; Thermo Fisher Scientific,
Inc.)The RVprimer3 (5′-CTAGCAAAATAGGCTGTCCC-3′) and GLprimer2
(5′-CTTTATGTTTTTGGCGTCTTCCA-3′) were used according to the
manufacturer's procol. The thermocycling conditions were as
follows: Initial denaturation 95°C 1 min, 28 of cycles of
denaturation 95°C 30 sec, annealing 55°C 30 sec and elongation 72°C
1 min; and final extension 72°C 10 min. HEK293T cells (in the
logarithmic growth phase) were co-transfected with wild type TGFA
(WT-TGFA) or mutated TGFA (mut-TGFA) in combination with control or
miR-130b-mimic, using Lipofectamine® 2000 according to
the manufacturer's instructions. The cells were harvested 24 h
following transfection and the relative luciferase activities were
determined using the dual-luciferase reporter assay system (Promega
Corporation, Madison, WI, USA) in a GLOMAX20/20 luminometer
(Promega Corporation) and normalized to the Renilla
luciferase activity.
Protein extraction and western blot
analysis
Osteosarcoma cells were harvested and lysed using 1X
radioimmunoprecipitation assay lysis buffer (Upstate Biotechnology,
Inc., Lake Placid, NY, USA). Equal amounts of protein (30 µg) were
separated using 8% SDS-PAGE and transferred onto polyvinylidene
fluoride membranes (Merck Millipore, Darmstadt, Germany). The
membranes were blocked with 5% bovine serum albumin in
Tris-buffered saline supplemented with 0.1% Tween-20 for 1 h at
room temperature, and then incubated overnight at 4°C with a
primary antibody against human TGFA(dilution, 1:1,000; cat. no.,
sc-36; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), protein
kinase B (Akt) (dilution, 1:1,000; cat. no., sc-135829, Santa Cruz
Biotechnology, Inc.), phosphorylated (p)Akt (dilution, 1:1,000;
cat. no., sc-16646-R; Santa Cruz Biotechnology, Inc.), epidermal
growth factor receptor (EGFR; dilution, 1:1,000; cat. no.,
sc-80543; Santa Cruz Biotechnology, Inc.), p-EGFR (dilution,
1:1,000; cat. no., sc-71776; Santa Cruz Biotechnology, Inc.) or
β-actin (dilution, 1:2,000; cat. no., sc-517582 Santa Cruz
Biotechnology, Inc.), and subsequently with a horseradish
peroxidase-conjugated anti-mouse IgG secondary antibody (dilution,
1:10,000; cat. no., sc-516102; Santa Cruz Biotechnology, Inc.). The
signals were visualized using a gel-imaging analyzer (Bio-Rad
Laboratories Inc., Hercules, CA, USA) and the protein band
intensity was quantified using Quantity One software (Version
4.5.0, Bio-Rad Laboratories, Inc.) and normalized to the β-actin
level.
Cell proliferation assay
The transfected cells were seeded into 96-well
plates, in DMEM medium supplemented with 10% FBS, at a density of
2,000 cells/well for Saos-2 and 5,000 cells/well for U-2OS, and
cultured at 37°C for ≤5 days. At the end of each experiment, a
10-µl Cell Counting kit-8 (CCK-8) solution was added to each well
and subsequently incubated for 2 h at 37°C. The relative optical
density (OD) level was evaluated at an absorbance of 450 nm and
normalized to that of the controls using a standard microplate
reader (Thermo Fisher Scientific, Inc.).
Colony-formation assay
Following transfection, 300 Saos-2 or U-2OS cells
were seeded into each well of 6-well plates and incubated at 37°C
for 14 days. The culture medium was replaced twice a week and on
day 14, cells were washed with PBS, fixed with methanol for
approximately 30 min, subsequently stained with 1% crystal violet
solution (Beyotime Institute of Biotechnology, Haimen, China) for
30 min, washed three times with PBS and then air-dried for 2 h.
Colonies containing ≥50 cells were identified as a clone. Data are
expressed as the mean ± standard error of the mean (SEM) and
compared with the control cells.
Tumor cell Transwell migration and
invasion assays
Migration and invasion assays were determined using
a BD Transwell assay (pore size 8 µm) with or without Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA), in accordance with the
manufacturer's protocol. Following transfection, Saos-2 or U-2OS
cells (5×104) were seeded into the upper wells on a
filter coated with or without Matrigel in DMEM without serum, and
the lower chambers were filled with DMEM supplemented with 10% FBS,
which acted as a chemoattractant. The cells were cultured for 24 h
at 37°C. Subsequently, the non-migrated or invaded cells on the
upper surface of the filter were removed using cotton swabs. Cells
that had migrated or invaded into the bottom surface were fixed
with 75% ethanol and stained with 1% crystal violet solution at
room temperature for 20 min. The relative degree of cell migration
or invasion was quantified using a light microscope (magnification,
×100) and normalized to the control cells in 5 fields of view.
Statistical analysis
All the data are expressed as the mean ± SEM. Data
were statistically analyzed using a paired-samples t-test using
SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA). Spearman's
correlation analysis was performed using Graphpad prism.6.0
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
The expression of miR-130b is
downregulated in osteosarcoma tissues and cell lines
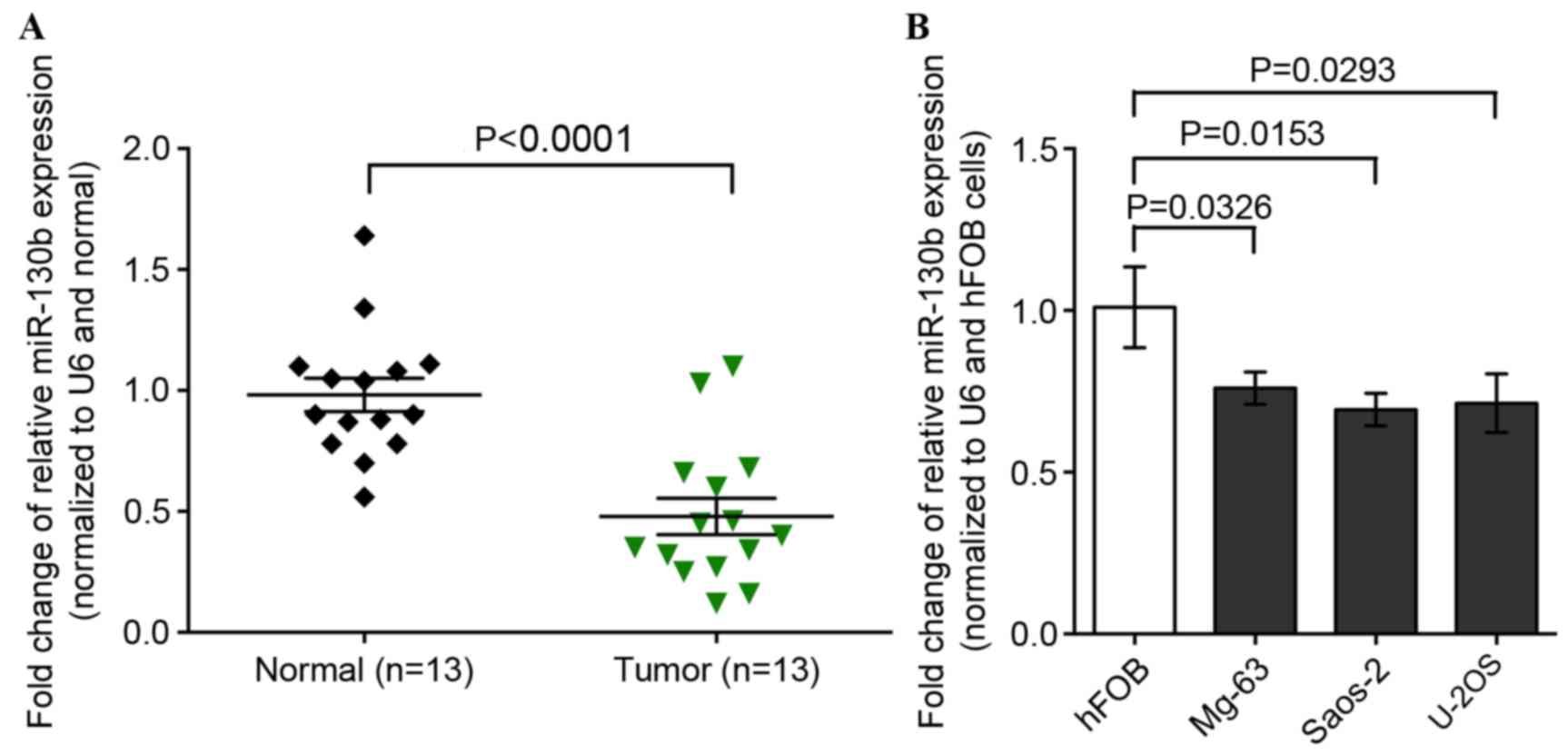
In order to investigate the role of miR-130b in
osteosarcoma, the present study determined the expression levels of
miR-130b in osteosarcoma and paired adjacent normal tissues from 13
patients, using RT-qPCR. It was revealed that miR-130b expression
levels were significantly lower in osteosarcoma tissues compared
with in normal tissues (P<0.0001; Fig.
1A). Furthermore, the expression level of miR-130b was also low
in the three osteosarcoma cell lines, Saos-2, MG-63 and U-2OS, as
compared with hFOB normal human osteoblast cells (Fig. 1B).
Restoration of miR-130b expression
levels inhibits osteosarcoma cell proliferation, colony formation,
migration and invasion
The present study restored and suppressed miR-130b
expression levels in U-2OS and Saos-2 cells via transfection of
miRNA-130b mimics and an miRNA-130b inhibitor, respectively. The
results revealed that miRNA-130b mimics and miRNA-130b inhibitors
induced or suppressed miR-130b expression levels respectively, in
U-2OS and Saos-2 cells compared with the negative control
transfection (Fig. 2A).
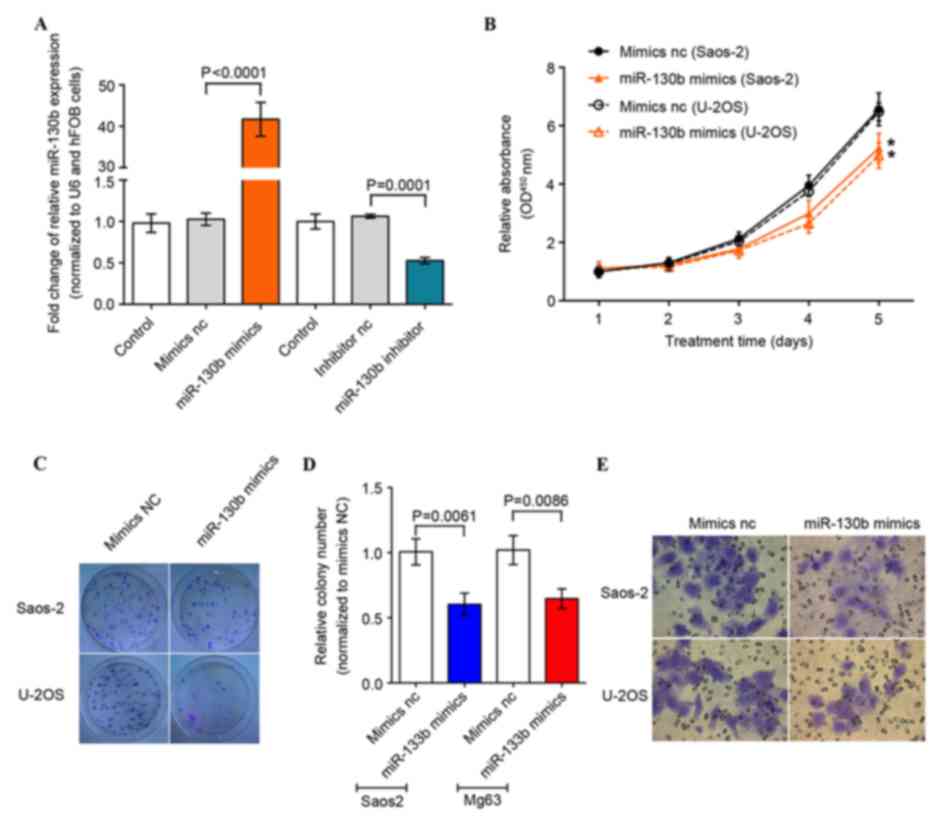 | Figure 2.Inhibition of osteosarcoma cell
proliferation, migration and colony formation by overexpression of
miR-130b. (A) A total of four oligonucleotides (miR-130b mimics,
miR-130b mimics NC, miR-130b inhibitor, miR-130b inhibitor NC) were
transfected into osteosarcoma cell lines and miR-130b expression
level was evaluated using RT-qPCR. (B) Tumor cell proliferation
CCK-8 assay. The data demonstrates the effect of miR-130b on the
proliferation of U-2OS and Saos-2 cells at 24, 48, 72, 96 and 120 h
following transfection. The absorbance was determined at 450 nm.
(C-E) Colony formation and Transwell assays performed 48 h
following transfection in order to evaluate the effects of miR-130b
on cell colony formation, migration and invasion in Saos-2 and
U-2OS cells. Results are expressed as the mean ± standard error of
the mean of three independent experiments. RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; NC, negative
control; miR, microRNA; CCK-8, cell counting kit-8; OD, optical
density. |
Following alteration of the miR-130b expression
level, osteosarcoma cells were subjected to cell proliferation and
colony formation assays. The cell proliferation CCK-8 assay
demonstrated that miR-130b-mimics reduced tumor cell proliferation,
compared with the negative control, from 1–5 days (Fig. 2B). Furthermore, the colony formation
assay revealed that the miR-130b-mimics reduced tumor cell colony
formation (Fig. 2C); the tumor cell
Transwell migration and invasion assays also demonstrated that the
miR-130b-mimic reduced migration and invasion capacity of
osteosarcoma cells (Fig. 2D and E).
These results indicate that the upregulation of miR-130b expression
suppressed osteosarcoma cell malignant behavior.
TGFA is the target gene of miR-130b in
osteosarcoma cells
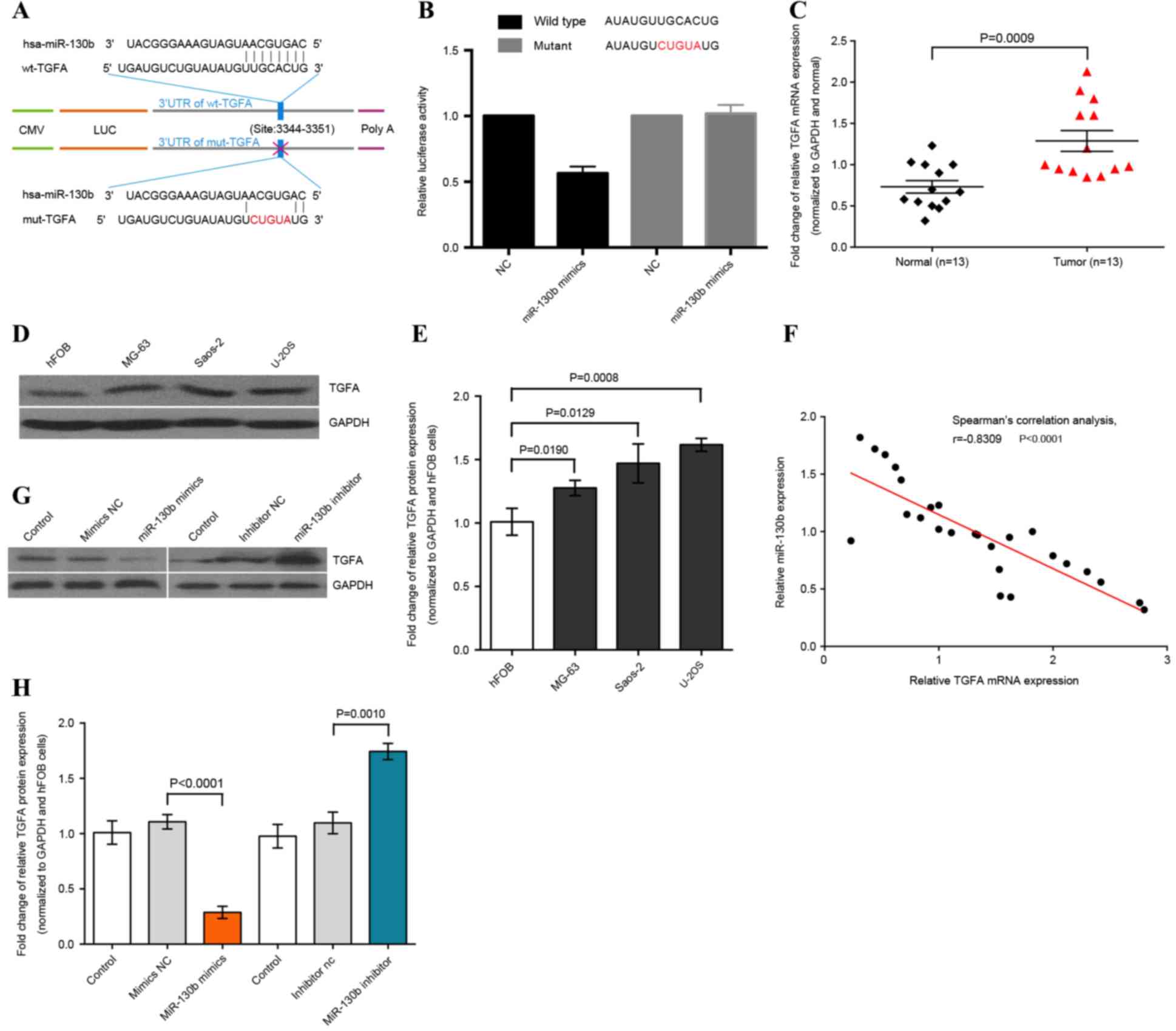
As miRNA serves a role in the inhibition of
targeting gene transcription and translation, a TargetScan search
was performed in order to screen for potential target genes of
miR-130b. It was revealed that miR-130b may suppress TGFA mRNA
translation. Therefore, the present study constructed luciferase
reporter plasmids and conducted a dual-luciferase reporter assay.
The results demonstrated that miR-130b mimics were able to directly
target the 3′-UTR of TGFA mRNA (Fig. 3A
and B); however, there was no significant difference in
luciferase activity detected between the miR-130b mimics and
negative control in the mut-TGFA-transfected U-2OS cells (Fig. 3A and B). Thus, this study determined
that TGFA is the downstream target of miR-130b.
Furthermore, it was verified that miR-130b mimics
induced a significant reduction of TGFA expression in osteosarcoma
cells (Fig. 3G); subsequently, the
expression levels of TGFA were evaluated when compared to normal
tissues (P=0.0009). Spearman's correlation analysis demonstrated
that TGFA expression levels in osteosarcoma tissues were inversely
associated with miR-130b expression levels (Spearman's correlation,
r=−0.8309; P<0.0001; Fig. 3F).
Following this, the relation of miR-130b and TGFA expression in
osteosarcoma cells was further confirmed by transfecting the
miR-130b mimics, miR-130b inhibitor and matched negative control
into osteosarcoma cells. The western blot analysis demonstrated
that miR-130b mimics inhibited TGFA expression, whereas the
miR-130b inhibitor upregulated TGFA expression in osteosarcoma
cells (P<0.01; Fig. 3G and H).
Thus, miR-130b may inhibit TGFA expression by targeting TGFA 3′-UTR
in osteosarcoma cells.
TGFA upregulation reverses the effects
of miR-130b on osteosarcoma cells
As aforementioned, miR-130b is able to target TGFA
and inhibit TGFA expression; therefore, the present study
determined whether the upregulation of TGFA expression may reserve
the effects of miR-130b on osteosarcoma cells via transfection of
the TGFA-expression vector with or without co-transfection of
miR-130b mimics or miR-130b mimics NC, into U-2OS and Saos-2
osteosarcoma cells. Tumor cell proliferation and clone formation
assays revealed that osteosarcoma cell proliferation was reduced
following miR-130b mimic transfection, but was reversed by
co-transfection with the pcDNA3.1-TGFA plasmid (Fig. 4A). Subsequently, clone formation
assays demonstrated that TGFA upregulation significantly restored
the clone formation capability of osteosarcoma cells
(P<0.01;Fig. 4C). Overexpression
of TGFA restored miR-130b-inhibited expression levels of pAkt and
EGFR (Fig. 4B), which suggested that
miR-130b may suppress the phosphatidylinositol 3-kinase (PI3K)/Akt
signaling pathway by targeting TGFA.
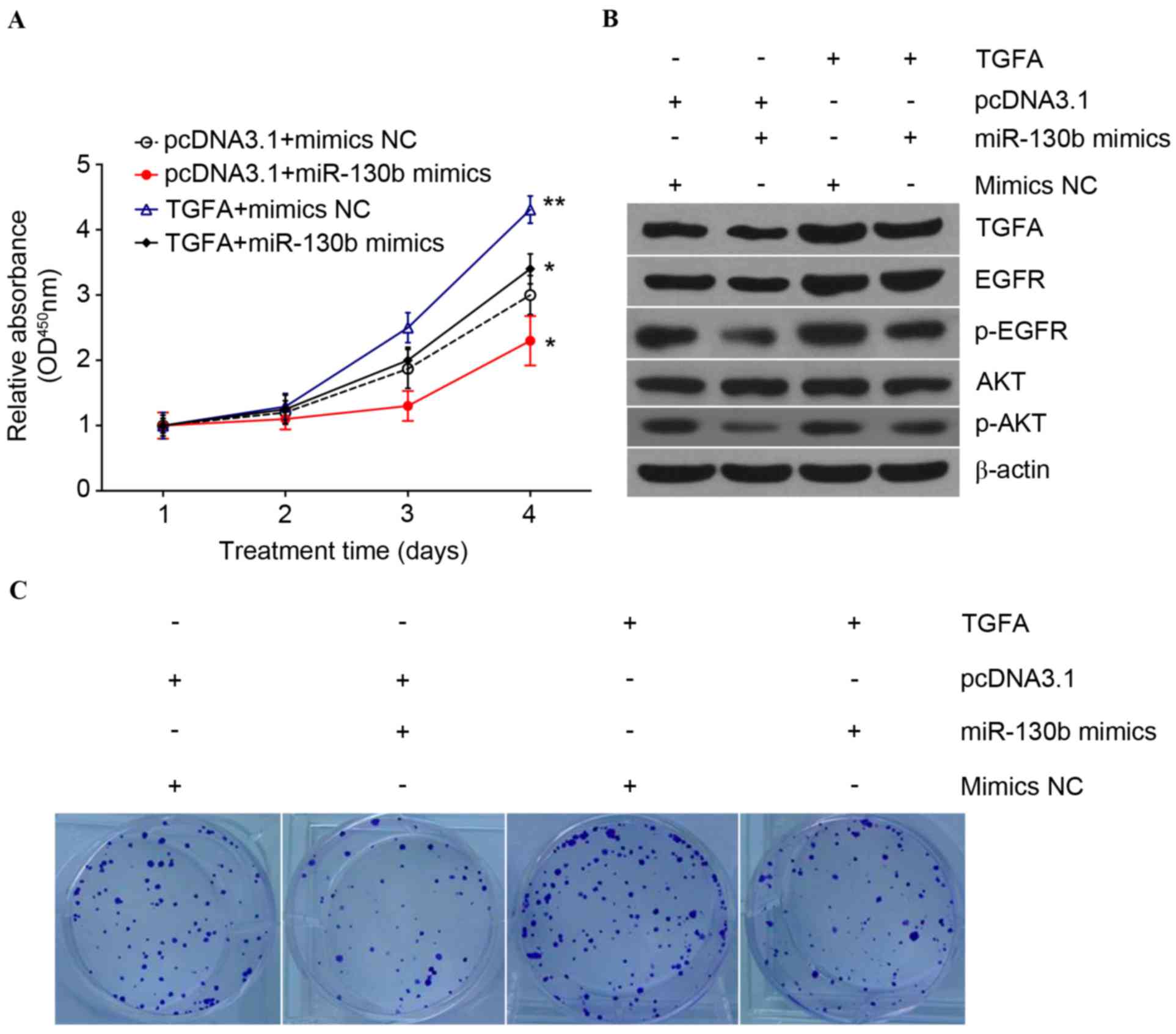 | Figure 4.TGFA expression reverses the effects
of miR-130b on the proliferation of osteosarcoma cells. (A) U-2OS
osteosarcoma cells transfected with the eukaryotic expression
plasmid (pcDNA3.1-TGFA) or an empty control plasmid
(pcDNA3.1-Ctrl), followed by second transfection with miR-130b
mimics or miR-130b mimics NC, and then subjected to a CCK-8 assay
in order to evaluate cancer proliferation. All groups were compared
with the control group (pcDNA3.1+mimicsNC) *P<0.01 and
**P<0.001. (B) Western blot analysis performed in order to
determine the levels of TGFA, EGFR and AKT proteins. (C) TGFA
expression reverses the effects of miR-130b on the clone formation
capability of osteosarcoma cells. U-2OS osteosarcoma cells
transfected with the eukaryotic expression plasmid (pcDNA3.1-TGFA)
or an empty control plasmid (pcDNA3.1-Ctrl), followed by second
transfection with miR-130b mimics or miR-130b mimics NC, and then
subjected to a clone formation assay in order to evaluate colony
formation capacity. TGFA, transforming growth factor α; ctrl,
control; NC, negative control; CCK-8, cell counting kit-8; EGFR,
epidermal growth factor receptor; AKT, protein kinase B; miR,
microRNA; OD, optical density; p, phosphorylated. |
Discussion
Over the previous decades, the knowledge and
understanding of the criteria and classification of osteosarcoma
have progressed; however, certain aspects of this disease remain
controversial (19). Developments in
the study of miRNAs have provided a more complete understanding of
how miRNAs contribute to cancer development and progression
(22). It is estimated that miRNAs
are able to regulate the expression of 30–60% of all human genes
(8) via inhibiting the expression of
their target genes by binding to the 3′-UTR of mRNA (23). A previous study revealed that miR-130b
expression is significantly reduced in endometrial cancer tissues,
and that mutant tumor protein 53 accelerates tumor progression and
metastasis via the regulation of miR-130b expression (14). miR-130b is downregulated in multidrug
resistant ovarian cancer cells, and the restoration of miR-130b
expression is concomitant with increased sensitivity of tumors to
anticancer drugs (24). Conversely,
miR-130b is significantly overexpressed in esophageal squamous cell
carcinoma cells, which increases the viability of tumor cells and
their ability to migrate and invade in vitro by targeting
phosphatase and tensin homolog (25).
Therefore, miR-130b may serve disparate tumor-associated roles
depending on the tumor type and targeted genes. Furthermore, the
present study demonstrated that miR-130b expression is
downregulated in osteosarcoma tissues and cell lines, whereas
restoration of miR-130b expression inhibits osteosarcoma cell
proliferation, colony formation, migration and invasion in
vitro. miR-130b is able to directly target TGFA mRNA and
inhibit TGFA expression in osteosarcoma cells. TGFA expression also
reverses the effect of miR-130b on osteosarcoma cells.
The association between miR-130b expression and
osteosarcoma has been demonstrated in a previous study by global
microarray analyses of a panel of 19 human osteosarcoma cell lines
and normal bone tissues (19). To the
best of our knowledge, there have been no further studies
investigating the role or the molecular mechanisms underlying
miR-130b in osteosarcoma. In the present study, the expression
level of miR-130b was determined in osteosarcoma tissues and three
osteosarcoma cell lines, and compared with adjacent tissues and
normal osteoblast cells, respectively. In contrast to a previous
study by Wu et al (18), the
present study revealed that miR-130b expression is downregulated in
osteosarcoma tissues and cell lines, which is consistent with that
in ovarian cancer, papillary thyroid carcinoma and hepatocellular
carcinoma (13,24,26).
Furthermore, miR-130b expression in osteosarcoma cells suppresses
tumor cell proliferation, colony formation, migration and invasion
in vitro. The present study further confirmed that miR-130b
may function as a tumor suppressor in osteosarcoma development and
progression.
TGFA encodes a growth factor that is a ligand for
the EGFR (27). The latter regulates
cell growth signaling and mediates cell proliferation in a number
of types of cancer, including lung, breast and prostate cancer
(28). Accumulating evidence has
demonstrated that EGFR activation via amphiregulin, epiregulin and
TGFA, triggers numerous biological responses, including cell
survival, proliferation, invasion, differentiation and migration
(28,29). It has previously been suggested that
there is an association between EGFR activation and other signaling
pathways, including the RAS-activated Akt signaling pathway
(30). PI3K/Akt is a major signaling
pathway that serves a role in the regulation of cell division in
malignant cells (31). Previous
studies have suggested that this signaling pathway may also have a
role in tumorigenesis and cancer progression by affecting the
activation state of a range of downstream effector molecules
(32,33). In the present study, it was
demonstrated that miR-130b is able to bind directly to the TGFA
3′-UTR and inhibit TGFA expression in osteosarcoma cells.
Furthermore, it was also revealed that the overexpression of TGFA
restored miR-130b-inhibited levels of pAkt and EGFR, which suggests
that miR-130b suppress the PI3K/Akt signaling pathway by targeting
TGFA. The data of the current study indicate that targeting the
miR-130b-mediated gene pathway may be a strategy for the
development of osteosarcoma treatments. In addition, the
tumor-suppressive effects of miR-130b upregulation on the
proliferation and migration of osteosarcoma cells were reversed
following transfection with TGFA. Further studies evaluating the
downstream genes of miR-130b, including Akt, may improve
understanding of the involvement of the PI3K/Akt signaling pathway
in osteosarcoma development or progression.
The present study is a proof-of-principle
investigation. Further research is required in order to confirm the
downregulation of miR-130b expression and its association with TGFA
expression in a larger number of tissues from patients with
osteosarcoma. In conclusion, the data from the current study
demonstrate that miR-130b is a novel regulator in osteosarcoma
development and progression, and that miR-130b-regulated TGFA
expression contributes to osteosarcoma cell malignant behavior.
Investigation of the underlying mechanisms of miR-130b and its
associated signaling pathways may assist the development of novel
therapeutic strategies to treat patients with osteosarcoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Science Foundation of China (grant no. 81201503).
Availability of data and materials
The datasets used/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW was responsible for the study design and the
acquisition of data, WS undertook data analysis, YK and BL
performed the functional experiments. MZ helped to design the
experiments and interpret the data. WW undertook project design and
manuscript revisions.
Ethics approval and consent to
participate
The present study was approved by the Human Ethics
Committee of the Xiangya Medical School of the Central South
University.
Consent for publication
Written informed consent was obtained from all
patients prior to enrollment in the present study.
Competing interests
The authors declare that they no financial conflicts
of interest.
References
|
1
|
Biazzo A and De Paolis M:
Multidisciplinary approach to osteosarcoma. Acta Orthop Belg.
82:690–698. 2016.PubMed/NCBI
|
|
2
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the Surveillance, Epidemiology, and End Results Program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Picci P: Osteosarcoma (osteogenic
sarcoma). Orphanet J Rare Dis. 2:62007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Logue JP and Cairnduff F: Radiation
induced extraskeletal osteosarcoma. Br J Radiol. 64:171–172. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Link MP, Goorin AM, Miser AW, Green AA,
Pratt CB, Belasco JB, Pritchard J, Malpas JS, Baker AR, Kirkpatrick
JA, et al: The effect of adjuvant chemotherapy on relapse-free
survival in patients with osteosarcoma of the extremity. N Engl J
Med. 314:1600–1606. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meyers PA, Healey JH, Chou AJ, Wexler LH,
Merola PR, Morris CD, Laquaglia MP, Kellick MG, Abramson SJ and
Gorlick R: Addition of pamidronate to chemotherapy for the
treatment of osteosarcoma. Cancer. 117:1736–1744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambros V: microRNAs: Tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin S and Gregory RI: MicroRNA biogenesis
pathways in cancer. Nat Rev Cancer. 15:321–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nana-Sinkam SP and Croce CM: Clinical
applications for microRNAs in cancer. Clin Pharmacol Ther.
93:98–104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burmistrova OA, Goltsov AY, Abramova LI,
Kaleda VG, Orlova VA and Rogaev EI: MicroRNA in schizophrenia:
Genetic and expression analysis of miR-130b (22q11). Biochemistry
(Mosc). 72:578–582. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yip L, Kelly L, Shuai Y, Armstrong MJ,
Nikiforov YE, Carty SE and Nikiforova MN: MicroRNA signature
distinguishes the degree of aggressiveness of papillary thyroid
carcinoma. Ann Surg Oncol. 18:2035–2041. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong P, Karaayvaz M, Jia N, Kaneuchi M,
Hamada J, Watari H, Sudo S, Ju J and Sakuragi N: Mutant p53
gain-of-function induces epithelial-mesenchymal transition through
modulation of the miR-130b-ZEB1 axis. Oncogene. 32:3286–3295. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leone V, Langella C, D'Angelo D, Mussnich
P, Wierinckx A, Terracciano L, Raverot G, Lachuer J, Rotondi S,
Jaffrain-Rea ML, et al: Mir-23b and miR-130b expression is
downregulated in pituitary adenomas. Mol Cell Endocrinol. 390:1–7.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sand M, Skrygan M, Sand D, Georgas D,
Gambichler T, Hahn SA, Altmeyer P and Bechara FG: Comparative
microarray analysis of microRNA expression profiles in primary
cutaneous malignant melanoma, cutaneous malignant melanoma
metastases, and benign melanocytic nevi. Cell Tissue Res.
351:85–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scheffer AR, Holdenrieder S, Kristiansen
G, von Ruecker A, Muller SC and Ellinger J: Circulating microRNAs
in serum: Novel biomarkers for patients with bladder cancer? World
J Urol. 32:353–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu X, Weng L, Li X, Guo C, Pal SK, Jin JM,
Li Y, Nelson RA, Mu B, Onami SH, et al: Identification of a
4-microRNA signature for clear cell renal cell carcinoma metastasis
and prognosis. PLoS One. 7:e356612012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Namlos HM, Meza-Zepeda LA, Baroy T,
Østensen IH, Kresse SH, Kuijjer ML, Serra M, Bürger H,
Cleton-Jansen AM and Myklebost O: Modulation of the osteosarcoma
expression phenotype by microRNAs. PLoS One. 7:e480862012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kansara M, Teng MW, Smyth MJ and Thomas
DM: Translational biology of osteosarcoma. Nat Rev Cancer.
14:722–735. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang C, Cai J, Wang Q, Tang H, Cao J, Wu L
and Wang Z: Epigenetic silencing of miR-130b in ovarian cancer
promotes the development of multidrug resistance by targeting
colony-stimulating factor 1. Gynecol Oncol. 124:325–334. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu T, Cao R, Li S, Fu M, Ren L, Chen W,
Zhu H, Zhan Q and Shi R: MiR-130b plays an oncogenic role by
repressing PTEN expression in esophageal squamous cell carcinoma
cells. BMC Cancer. 15:292015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin YH, Wu MH, Liao CJ, Huang YH, Chi HC,
Wu SM, Chen CY, Tseng YH, Tsai CY, Chung IH, et al: Repression of
microRNA-130b by thyroid hormone enhances cell motility. J Hepatol.
62:1328–1340. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Singh B, Carpenter G and Coffey RJ: EGF
receptor ligands: Recent advances. F1000Res. 5:pii: F1000. 2016.
View Article : Google Scholar
|
|
28
|
Ferguson KM, Berger MB, Mendrola JM, Cho
HS, Leahy DJ and Lemmon MA: EGF activates its receptor by removing
interactions that autoinhibit ectodomain dimerization. Mol Cell.
11:507–517. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Traish AM and Morgentaler A: Epidermal
growth factor receptor expression escapes androgen regulation in
prostate cancer: A potential molecular switch for tumour growth. Br
J Cancer. 101:1949–1956. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gan Y, Shi C, Inge L, Hibner M, Balducci J
and Huang Y: Differential roles of ERK and Akt pathways in
regulation of EGFR-mediated signaling and motility in prostate
cancer cells. Oncogene. 29:4947–4958. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Costa RLB, Han HS and Gradishar WJ:
Targeting the PI3K/AKT/mTOR pathway in triple-negative breast
cancer: A review. Breast Cancer Res Treat. Feb 18–2018.(Epub Ahead
of Print). View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Engelman JA: Targeting PI3K signalling in
cancer: opportunities, challenges and limitations. Nat Rev Cancer.
9:550–562. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|