Introduction
Nasopharyngeal carcinoma (NPC), which originates in
the nasopharynx, is the most common type of head and neck
malignancy in Southern China and certain regions of Southeast Asia.
Currently, the standard treatment for NPC consists of concurrent
chemo-radiotherapy, followed by adjuvant chemotherapy (1). Despite marked improvements in clinical
treatments for NPC, neck lymph node metastasis occurs in up to 75%
of NPC patients, which represents an unfavorable prognostic factor
for the disease (2). Radiotherapy
resistance is a cause of local recurrences and distant metastases
(3). A previous study revealed that
colony-stimulating factor-1 receptor (CSF-1R) expression was
4.1-times higher in NPC patients who were resistant to radiation
than in those who were sensitive to radiation (4). Furthermore, the expression of CSF-1R in
NPC tissues was markedly higher than that in the nasopharyngitis
tissues, and patients with moderate or strong-intensity of CSF-1R
expression were more likely to develop metastasis and recurrence
(5). Therefore, a greater
understanding of the biological features of NPC is urgently
required.
CSF-1R, a member of the receptor tyrosine kinase
(RTK) family, is one of the main regulatory factors in the immune
system, and is encoded by the proto-oncogene c-fms. Ligand binding
activates CSF-1R through a process of oligomerization and
trans-phosphorylation, to ultimately activate tyrosine kinase
signaling pathways, which may result in tumor cell proliferation
(6). Growing evidence has indicated
that CSF-1R, together with its ligand colony-stimulating factor 1
(CSF-1/M-CSF), serves an important role in the development of
cancer and may be involved in the process of carcinogenesis, and
tumor cell proliferation and metastasis (7,8). Thus far,
CSF-1R has been revealed to be upregulated in breast cancer
(9), ovarian cancer (10) and head and neck malignancies (11). The expression of CSF-1R in the blood
has been identified as a biomarker in numerous malignant tumors
(12). A previous study demonstrated
that CSF-1R amplification in breast cancer increased cell
proliferation (13). In addition,
another study reported that the CSF-1R inhibitor significantly
reduced the volume of microglia and suppressed the proliferative
and invasive abilities of the cells (14). However, the effects of CSF-1R on NPC
and the possible underlying mechanisms of this have not yet been
evaluated. Therefore, the present study investigated the potential
promotional effects of CSF-1R on the proliferation, migration and
invasion of the 6–10B NPC cell line, and the possible molecular
mechanisms underlying CSF-1R-induced cell proliferation and
metastasis in NPC.
Materials and methods
Cell culture
The human NPC 6–10B cell line was obtained from Sun
Yat-Sen University Cancer Centre (State Key Laboratory of Oncology
in South China, Guangzhou, China). The cells were cultured in
RPMI-1640 medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) in a humidity-controlled 37°C incubator with 5%
CO2. The medium was refreshed every other day.
Transfection
Lentiviral vectors were constructed by Shanghai
GeneChem Co., Ltd. (Shanghai, China). The viruses carried the
enhanced green fluorescent protein (eGFP) gene. CSF-1R (NM 005211;
GeneChem Co., Ltd., Shanghai, China) was transformed into GV358
vector (GeneChem Co., Ltd., Shanghai, China) using ClonExpressTM II
One Step Cloning kit (Vazyme Biotech Co., Ltd., Nanjing, China).
The constructs were then transfected into 293T cells (GeneChem Co.,
Ltd., Shanghai, China) with lentiviral packaging vectors using
Lipofectamine 2000 (Thermo Fisher Scientific, Inc.). Lentivirus was
collected 48 h after transfection and used to infect the 6-10B
cells. The viruses were diluted in sterile phosphate-buffered
saline (PBS) to a final titer of 5E+8 TU/ml. Lentiviruses carrying
the CSF-1R gene (ID, LV-CSF1R) or a control vector were transfected
into 6-10B cells respectively when cell confluence reached 20-30%.
Expression of the eGFP protein was assessed 3 days following
transfection to determine transfection efficiency using an inverted
fluorescence microscope at magnification, ×100. CSF-1R expression
in the 6-10B cells treated with control vector (control group) or
CSF-1R overexpression vector (transfection group) was then verified
using reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blot analysis.
RT-qPCR
Total RNA was extracted from the 6-10B cells using
TRIzol® reagent (Ambion; Thermo Fisher Scientific,
Inc.). cDNA was synthesized using a FastQuant RT kit (Tiangen
Biotech Co., Ltd., Beijing, China) according to the manufacturer's
protocol with the following temperature protocol: 42°C for 15 min,
followed by 95°C for 3 min and then cooling on ice at 4°C for 10
min. Subsequently, 100 ng total cDNA was added per 25-µl reaction
with CSF-1R or GAPDH primers, and quantitative PCR analysis was
performed using a SuperReal PreMix Plus SYBR Green PCR kit (Tiangen
Biotech Co., Ltd.), according to the manufacturer's protocols. The
reactions are typically performed on a PCR machine, and the qPCR
thermo cycling conditions were as follows: 95°C for 15 min, 40
cycles of 95°C for 10 sec and 60°C for 32 sec. Finally, the data
were analyzed using the 2−ΔΔCq method (15) with GAPDH expression as the endogenous
control. All mRNA primers were designed with Primer Premier 5.0
software (Premier Tech Co, Ltd., Quebec, Canada) and synthesized by
Thermo Fisher Scientific, Inc. The qPCR primers used to amplify the
CSF-1R and GAPDH genes are as follows: CSF-1R forward,
5′-TCTGGTCCTATGGCATCCTC-3′ and reverse, 5′-GATGCCAGGGTAGGGATTC-3′;
and GAPDH forward, 5′-AGCCACATCGCTCAGACAC-3′ and reverse,
5′-GCCCAATACGACCAAATCC-3′.
Western blot analysis
Cells were harvested and lysed in
radioimmunoprecipitation assay cell lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China), mixed with protease
inhibitor cocktail (100:1), on ice for ~30 min. The cells were then
pelleted by centrifugation at 4°C for 20 min at 12,000 × g. Total
protein was extracted from the supernatant and the concentration
was determined using a bicinchoninic acid assay kit (Beyotime
Institute of Biotechnology). Equal amounts of protein (40 µg) were
separated on a 12% SDS-PAGE gel and were transferred onto
polyvinylidene difluoride membranes. The membranes were
subsequently blocked with 3% bovine serum albumin for 1 h at room
temperature, prior to being incubated overnight at 4°C with the
following antibodies: Polyclonal rabbit anti-CSF-1R (dilution,
1:1,000; catalog no. 3152; Cell Signaling Technology, Inc.,
Danvers, MA, USA), monoclonal rabbit anti-cyclin D1 (dilution,
1:1,000; catalog no. ab134175; Abcam, Cambridge, UK), monoclonal
rabbit anti-Bcl-2 (dilution, 1:1,000; catalog no. 4223; Cell
Signaling Technology, Inc.), monoclonal rabbit anti-Bax (dilution,
1:2,000; catalog no. ab32124; Abcam, Cambridge, UK), monoclonal
rabbit anti-Akt (dilution, 1:1,000; catalog no. 9272; Cell
Signaling Technology, Inc.), monoclonal rabbit anti-phosphorylated
(p)-Akt (dilution, 1:2,000; catalog no. 4060; Cell Signaling
Technology, Inc.) and monoclonal rabbit anti-GAPDH (dilution,
1:1,000; catalog no. ab181602; Abcam). Membranes were subsequently
incubated with a goat anti-rabbit IgG secondary antibody (dilution,
1:3,000; catalog no. 7074; Cell Signaling Technology, Inc.) for 1.5
h at 37°C. Following washing three times with TBST, signals were
detected with an enhanced chemiluminescence detection system
(Pierce; Thermo Fisher Scientific, Inc.).
MTT assay
6-10B cells transfected with the control vector
(control group) or the CSF-1R vector (transfection group) were
seeded onto 96-well plates at a density of 1,000 cells/well.
Following incubation for 24, 48, 72, 96 or 120 h, 20 µl MTT reagent
was added (at a final concentration of 0.5 mg/ml). The cells were
then cultured for another 4 h and 150 µl dimethyl sulfoxide was
added to each well to dissolve the purple formazan. The cells were
subjected to an absorbance reading at 490 nm using a 96-well
microplate reader. All experiments were performed in triplicate.
Cell proliferation was evaluated at 24, 48, 72, 96 or 120 h.
Flow cytometric apoptosis assay
The two groups of 6-10B cells were seeded onto
6-well plates and trypsinized when the cells reached 70-80%
confluence. Annexin V and propidium iodide (PI) staining was
performed using an Annexin V-APC/7-AAD Apoptosis Detection kit
(Nanjing KeyGen Biotech Co., Ltd., Nanjing, China), according to
the manufacturer's protocols. Following incubation for ~15 min in
the dark, apoptotic cells were immediately analyzed by a flow
cytometer (fluorescence-activated cell sorting; BD Biosciences,
Franklin Lakes, NJ, USA). Using Annexin V-APC and 7-AAD staining
enables different stages of cell apoptosis to be distinguished. For
example, APC-positive and 7-AAD-negative staining indicates early
apoptotic cells, while 7-AAD-positive and APC-positive staining
indicates late apoptotic cells. FlowJo 7.6 software (Tree Star
Inc., Ashland, OR, USA) was used to calculate rate of
apoptosis.
Wound healing assay
The two groups of 6-10B cells were cultivated in
6-well plates (5×105 cells/well). When cell confluence
reached 70–80%, a 200 µl pipette tip was used to scrape a
line-shaped wound in the cell monolayer on each plate. Cells were
subsequently cultured for 24 h in serum-free medium. Cell migration
was evaluated at 0, 12 and 24 h by a light microscope at
magnification, ×100 (Olympus Corporation, Tokyo, Japan).
Transwell invasion assay
An invasion assay was performed in a 24-well
Transwell chamber (Corning Incorporated, Corning, NY, USA). A total
of 5×104 cells were seeded onto filters pre-coated with
Matrigel with 150 µl serum-free RPMI-1640 medium. The lower chamber
was supplied with 500 µl medium containing 10% FBS. After culturing
for 24 h, all the medium was removed. The filters were fixed with
95% methanol for 15–20 min at room temperature and stained with
0.05% crystal violet for 20 min at 25°C, prior to being washed
three times with PBS. Finally, the number of cells adhered to the
bottom of the filters was counted using a light microscope at
magnification, ×100.
Statistical analysis
Data are presented as the mean ± standard deviation.
Student's t-test was used to compare the diversity between the two
groups, using SPSS statistical software version 22.0 (IBM Corp.,
Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Overexpression of CSF-1R in 6–10B
cells
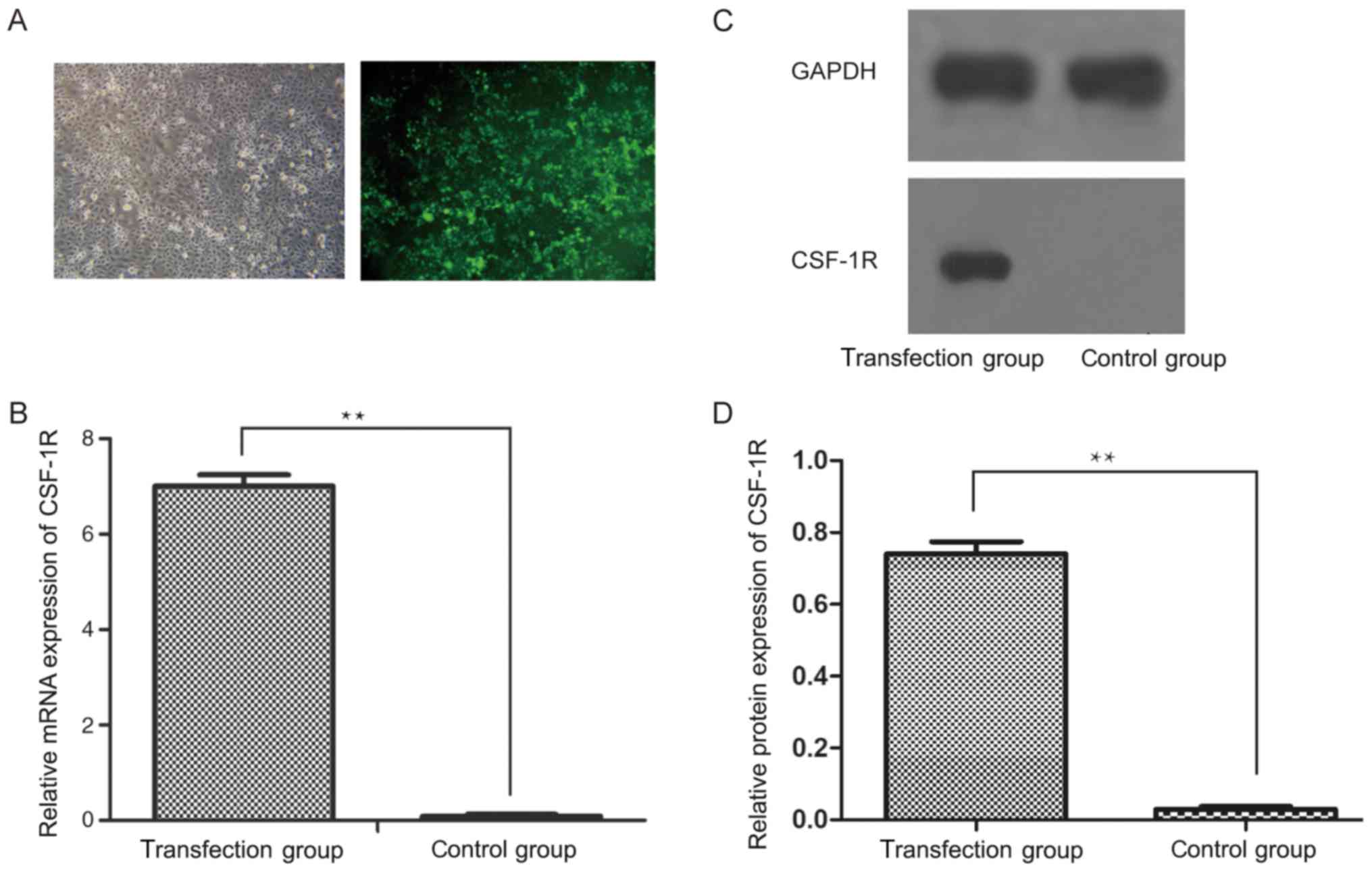
Following the transfection of a CSF-1R
overexpression vector into 6–10B cells, the transfection efficiency
was analyzed by inverted fluorescence microscopy (Fig. 1A). Subsequently, RT-qPCR analysis
revealed that the mRNA expression of CSF-1R was significantly
upregulated in the transfection group, but was downregulated in the
control group (Fig. 1B; P<0.01).
Similarly, the protein expression of CSF-1R was markedly increased
in the transfection group (Fig. 1C;
P<0.01), which indicated that CSF-1R was successfully
overexpressed in the 6–10B cells.
CSF-1R overexpression induces the
proliferation of 6–10B cells
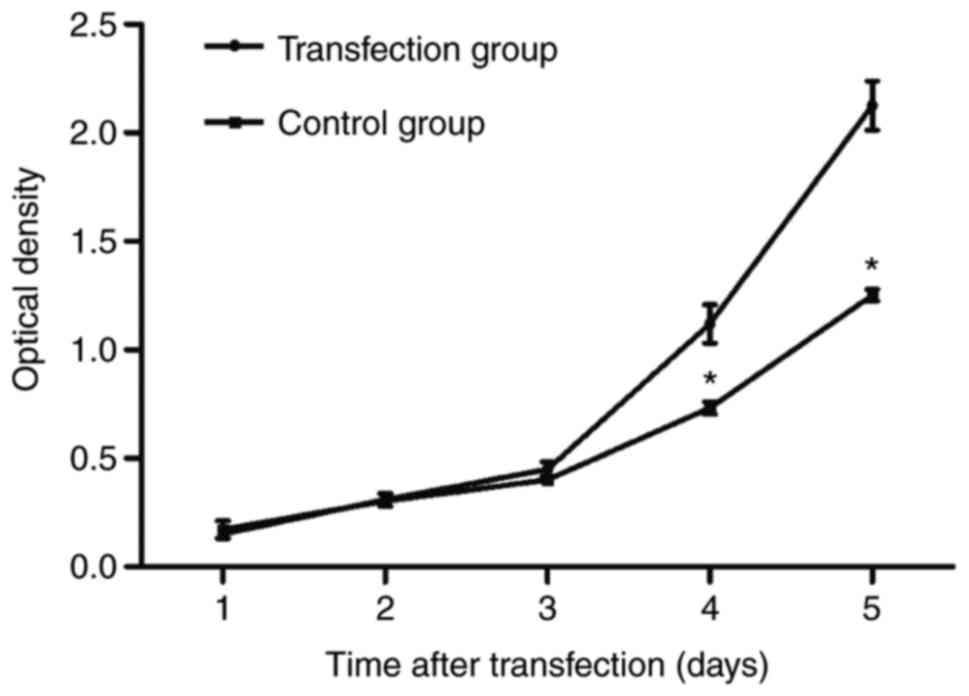
6-10B cell proliferation was measured via an MTT
assay. As demonstrated in Fig. 2, the
number of 6–10B cells in the transfection group significantly
increased from day 4, compared with that in the control group
(P<0.05), which suggested that CSF-1R overexpression promoted
6–10B cell proliferation.
CSF-1R overexpression reduces the
apoptosis of 6–10B cells
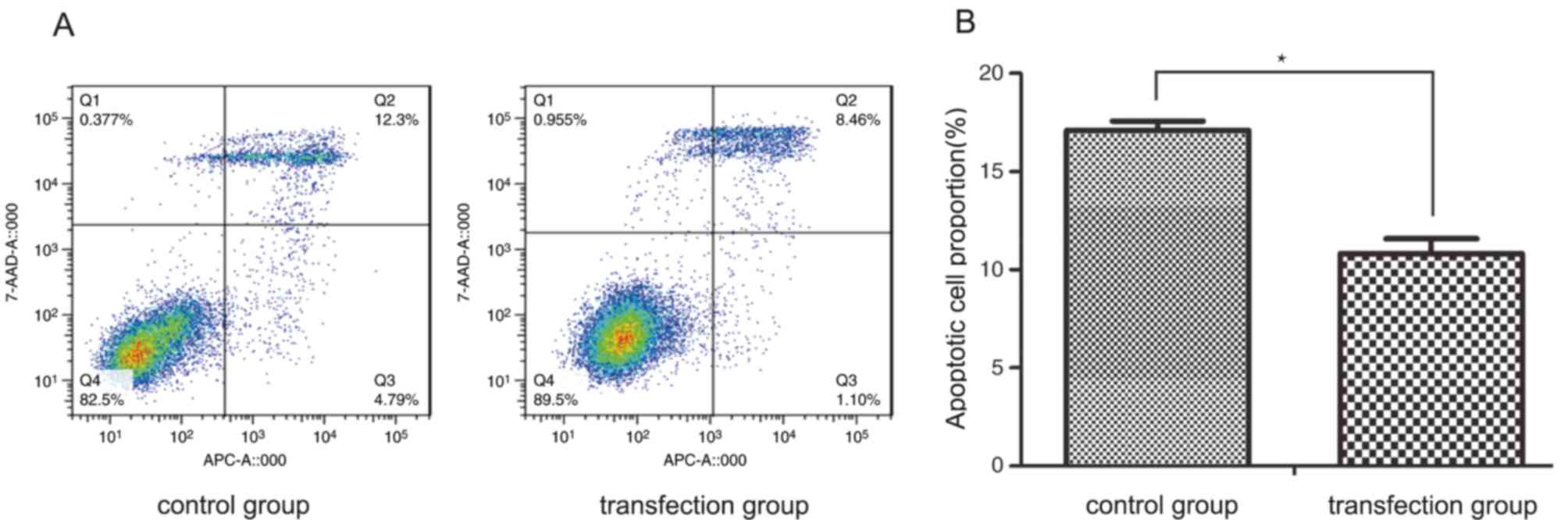
In order to assess whether CSF-1R was able to
inhibit the apoptosis of 6–10B cells, staining with Annexin V-APC
and 7-AAD was performed. As demonstrated in Fig. 3, a lower rate of cell apoptosis was
observed in the transfection group compared with that in the
control group, with regards to early-stage and late-stage apoptosis
(10.82±0.75 vs. 17.11±0.46%; P<0.05). This result revealed that
CSF-1R overexpression significantly inhibited the apoptosis of
6–10B cells.
CSF-1R overexpression promotes 6–10B
cell migration and invasion
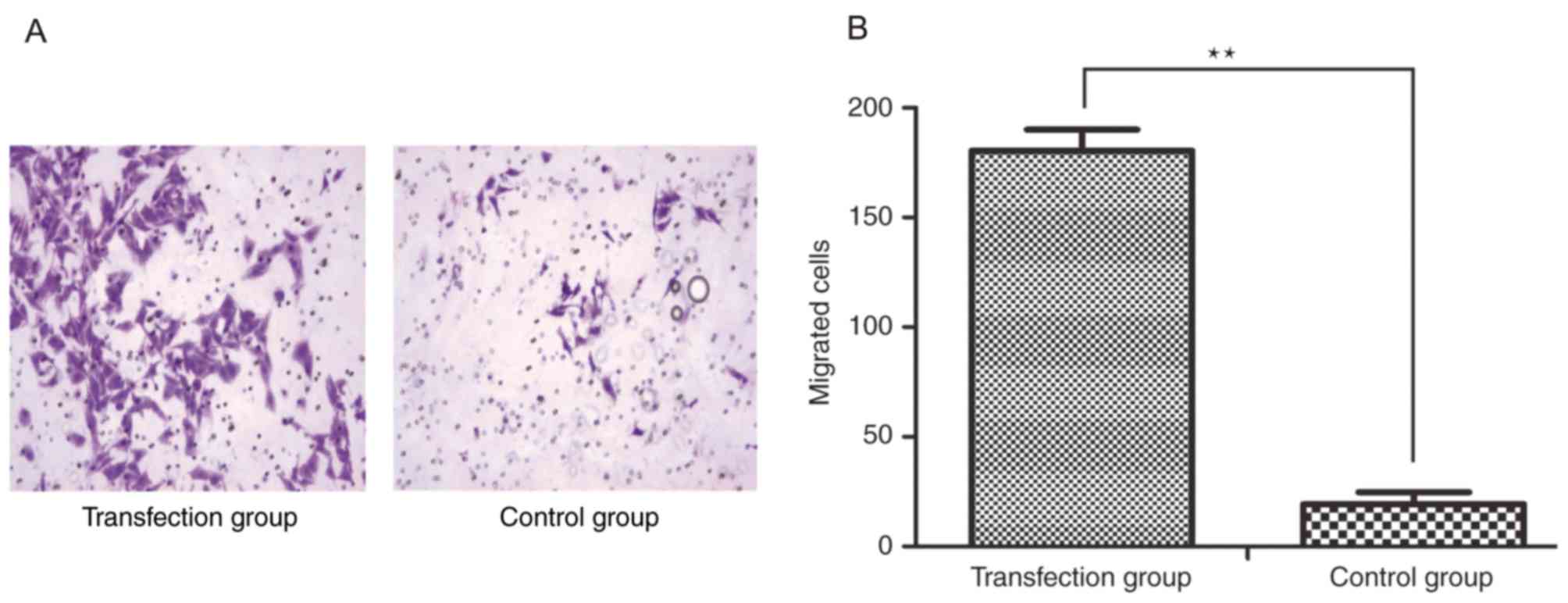
In order to demonstrate the effects of CSF-1R on
cell invasion, an invasion assay was performed, in which cells
passing through the polycarbonate membrane of a Transwell invasion
chamber were counted. Five visual fields were randomly selected to
calculate the average number of migrated cells in each group, which
was determined to be 180.400±9.633 per field in the transfection
group and 19.200±5.541 per field in the control group
(magnification, ×100; Fig. 4A).
Analysis of these results indicated that the invasive ability was
significantly increased in the cells overexpressing CSF-1R compared
with that in the control cells (Fig.
4B; P<0.01).
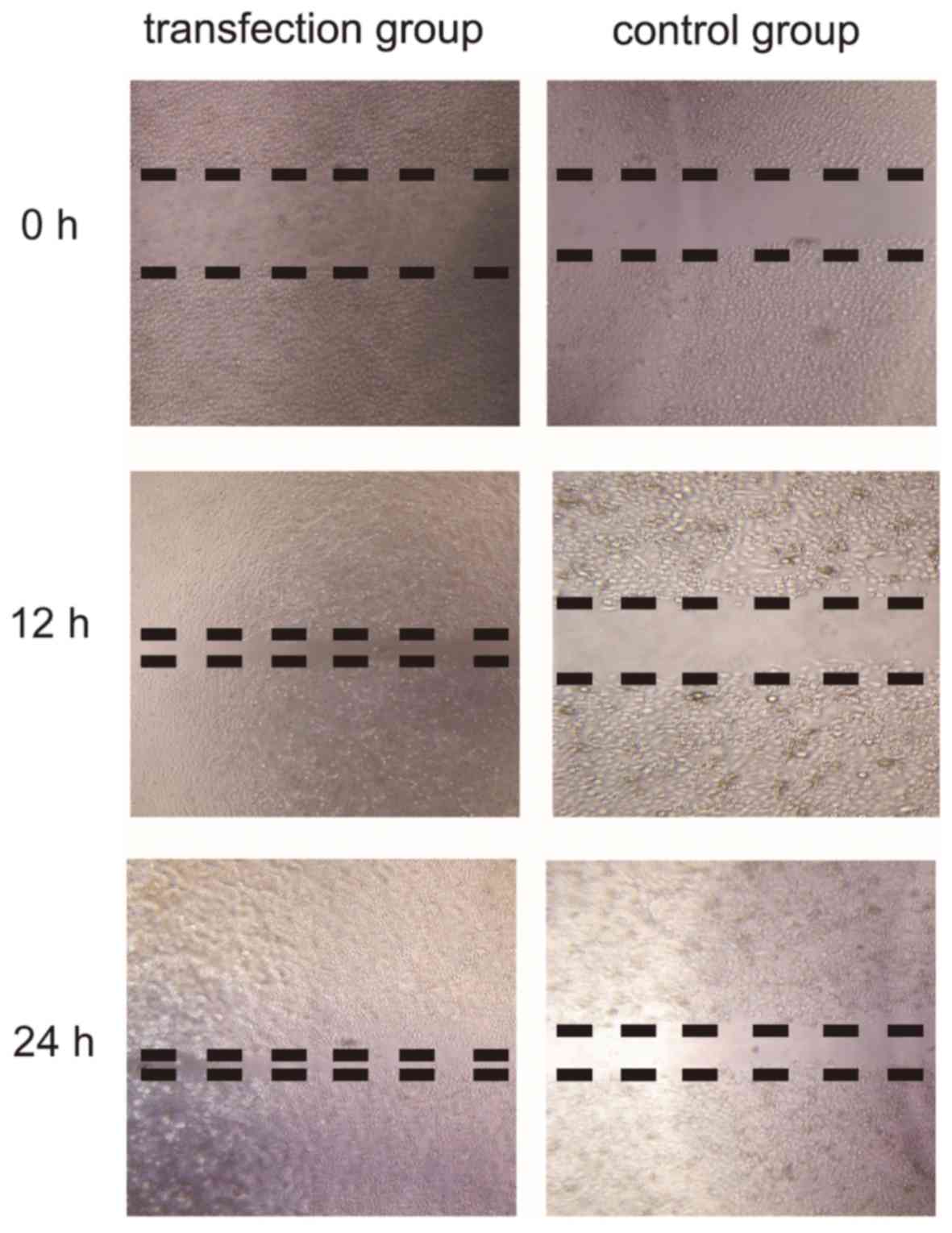
In order to prove that CSF-1R regulates cell
migration, the migration of cells was assessed in a wound healing
assay. As demonstrated in Fig. 5,
CSF-1R expression markedly accelerated cell migration in the
transfection group. Overall, these results suggested that CSF-1R
may induce 6–10B cell invasion and migration.
CSF-1R overexpression enhances the
activity of PI3K/Akt in 6–10B cells
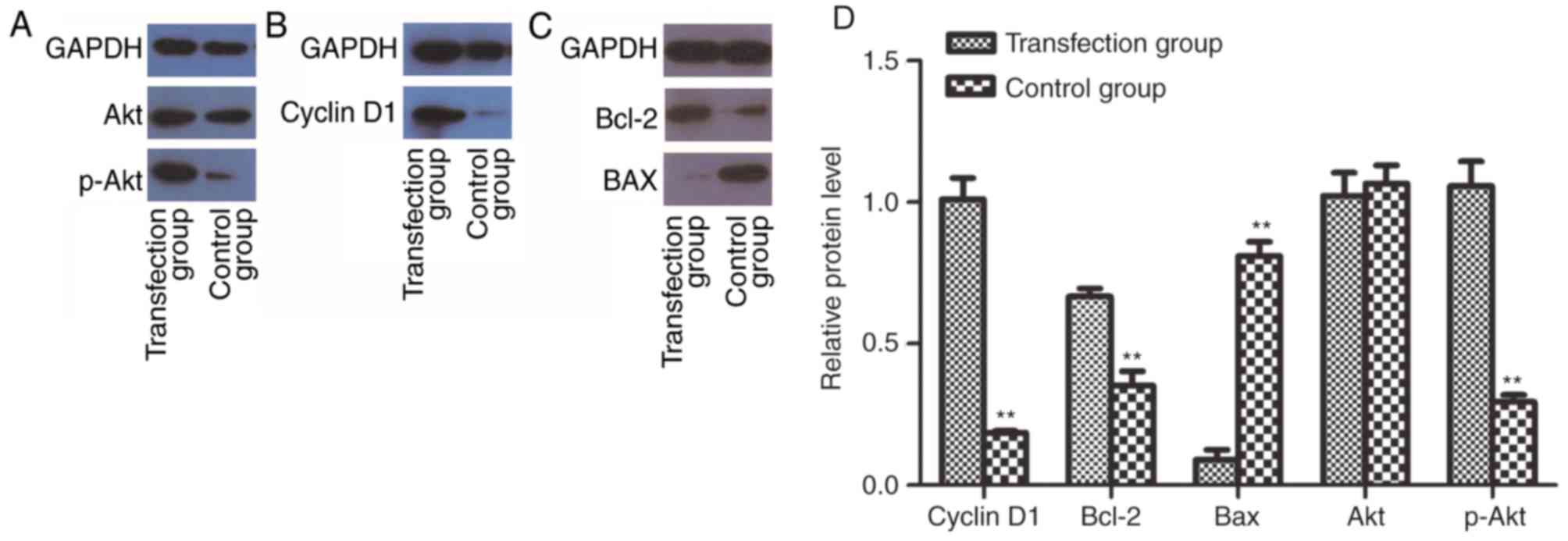
To determine whether CSF-1R may promote cell
proliferation by activating the PI3K/Akt pathway, western blotting
was performed to analyze the protein expression of cyclin D1,
Bcl-2, Bax, Akt and p-Akt. Notably, the expression of p-Akt was
increased in the transfection group (Fig.
6A). Accordingly, the expression of cyclin D1 (Fig. 6B) and Bcl-2 (Fig. 6C) was also enhanced. By contrast, Bax,
as a factor that inhibits tumor growth, was downregulated (Fig. 6C). These results indicated that the
activation of PI3K/Akt signaling may mediate the increased
proliferative ability of 6–10B cells overexpressing CSF-1R.
Discussion
Although substantial progress has been achieved in
the treatment of human NPC via radiotherapy, the 5-year local
recurrence rate of patients remains at 30, and 30–60% of patients
experience metastasis (16).
Radiotherapy resistance is a cause of local recurrence and distant
metastases. The involvement of CSF-1R in NPC has recently been a
focus of research. CSF-1R is established as a member of the
receptor tyrosine kinase (RTK) family, and is involved in
regulating the immune system (17,18). To
date, it has been demonstrated that the expression of CSF-1R and
its ligand in neoplastic epithelial cells is correlated with
metastasis, angiogenesis and cell invasion, resulting in poor
prognosis and recurrence (19).
Accumulating evidence has proven that PI3K/Akt pathways are
involved in RTK-induced progression and migration in human cancer
cell lines (20). However, few
studies have focused on the effects of CSF-1R on NPC cells.
Therefore, the present study overexpressed CSF-1R in
the human NPC 6–10B cell line by lentiviral transfection (Fig. 1A), and measured its expression level
by RT-qPCR (Fig. 1B) and western blot
analysis (Fig. 1C). Subsequently, a
series of tests were performed to compare the proliferation,
migration and invasion abilities of the transfection group with
those of a control group (Figs.
2–5). The results demonstrated
that cell proliferation, migration and invasion were increased in
the transfection group when compared with their levels in the
control group. Furthermore, p-Akt protein abundance was increased
following CSF-1R overexpression. As a consequence, the protein
abundance of cyclin D1 and Bcl-2 was also increased, whereas that
of Bax was decreased (Fig. 6), which
suggested that the CSF-1R-induced proliferation of 6–10B cells
involved p-Akt activation. Therefore, it may be hypothesized that
CSF-1R elicits the PI3K/Akt pathway, involving Akt phosphorylation,
to induce cyclin D1 and Bcl-2 expression and reduce Bax expression,
and ultimately promote cell proliferation.
It is well known that cyclin D1 belongs to the
highly conserved cyclin family and primarily functions as a factor
that promotes cell proliferation. Namely, it regulates the
transition of the G1 phase into the S phase by dimerizing with
CDK4/6. Any mutation, amplification or overexpression of this gene
may alter cell cycle progression, which may contribute to
tumorigenesis (21). Cyclin D1
overexpression contributes to increased chemotherapeutic resistance
and protection from apoptosis, resulting in tumor progression.
Furthermore, cyclin D has been widely confirmed as a biomarker in
breast carcinoma (22), with its
overexpression leading to tumorigenesis and malignancy. To date,
cyclin D1 expression has been detected at very high levels in
numerous types of malignant tumor, including head and neck squamous
cell carcinoma (23), breast
carcinoma (24), ovarian cancer
(25), colorectal cancer (26) and gastric cancer (27).
Members of the Bcl-2 family are among the most
established regulators of apoptosis. The Bcl-2 family proteins are
divided into two groups according to their function: One group
includes Bcl-2, B-cell lymphoma-XL and Bcl-2-like protein 2, all of
which promote apoptosis, while the other group includes Bax, Bcl-2
homologous antagonist killer and
phorbol-12-myristate-13-acetate-induced protein 1, which inhibit
apoptosis. These proteins control apoptosis by regulating outer
mitochondrial membrane permeability and the release of cytochrome C
(28). Bcl-2 is generally considered
to be an anti-apoptotic protein that serves an important role in
promoting cellular survival and inhibiting the actions of
pro-apoptotic proteins (29). By
contrast, Bax is identified as a pro-apoptotic member of the Bcl-2
protein family that stimulates apoptosis by heterodimerizing with
Bcl-2 to inhibit its anti-apoptotic function (30). To date, it has been widely reported
that the Bcl-2/Bax ratio is a key factor that reflects the degree
of cell apoptosis resistance (31).
The present study demonstrated that CSF-1R overexpression was able
to increase the expression of Bax and reduce the expression of
Cyclin D1 and Bcl-2. From these results, it may be concluded that
CSF-1R overexpression promotes proliferation and inhibits apoptosis
in NPC 6–10B cells.
The PI3K/Akt pathway is an intracellular signaling
pathway that is associated, not only with tumor proliferation and
survival, but also with invasion and metastasis (32). Only class I PI3K has been shown to
regulate this pathway and is associated with tumor cell behavior.
By interacting with growth factors or linked proteins that possess
phosphorylated tyrosine residues, PI3K is activated, resulting in
the generation of the second messenger
phosphatidylinositol-3,4,5-triphosphate (PIP3). PIP3 then combines
with Akt and phosphoinositide-dependent kinase-1 (PDK1), thereby
stimulating PDK1 to phosphorylate Thr(308) on the central kinase
catalytic domain of Akt. Subsequently, Akt is activated and
phosphorylates specific downstream proteins, including the
pro-apoptotic proteins of the Bcl-2 family and glycogen synthase
kinase 3 (GSK-3), thereby preventing the phosphorylation and
degradation of cyclin D1 (33).
Several RTKs regulate the activation of the PI3K/Akt pathway
(34). It has been reported that, by
inhibiting the activity of PI3K, cell proliferation may be
significantly reduced and cell cycle progression prohibited;
however, these effects may be prevented when activated Akt (p-Akt)
is expressed (32).
The present study further revealed that one of the
RTK family members, CSF-1R, may promote cell proliferation and
inhibit cell apoptosis via the PI3K/Akt pathway. It was revealed
that CSF-1R activated PI3K-dependent Akt phosphorylation, resulting
in the upregulation of Cyclin D1 and Bcl-2 and the downregulation
of Bax. Consequently, there was an increase in cell cycle
progression and a reduction in cell apoptosis. However, these
observations were based on one cell line, and future studies
involving additional cell lines and more clinical data are required
to confirm these effects in patients with NPC.
In conclusion, the present study demonstrated that
CSF-1R promoted the proliferation, migration and invasion of the
6–10B NPC cell line. The possible underlying mechanisms may involve
activation of the PI3K/Akt pathway. These findings provide further
understanding of the processes underlying NPC development, and
indicate that a CSF-1R inhibitor may be an effective treatment for
NPC.
Acknowledgements
The authors would like to thank Dr Musheng Zeng and
his colleagues at Sun Yat-Sen University Cancer Center for
providing the 6–10B cell line.
Funding
The present study was supported by Guangxi Natural
Science Fund (grant no. 2010037) and the National Natural Science
Fund (grant no. 81260348).
Availability of data and resources
The datasets used or analyzed during the current
study are available from the corresponding authors on reasonable
request.
Authors' contributions
JYC performed experiments and was a major
contributor in writing the manuscript. YH contributed to the
conception and design of the present study. JXC also contributed to
the conception and design of the present study. LH conducted cell
transfection and RT-qPCR. WA conducted western blotting and the
flow cytometric apoptosis assay. JY analyzed and interpreted the
data. WL collected clinical data. LL conducted IHC. JH followed up
the patients and analyzed the clinical data. ZC and XH performed
radiotherapy. WG evaluated the NPC patients during the treatment in
the preliminary study.
Ethics statement and consent to
participate
Not applicable.
Consent for publication
No applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CSF-1R
|
colony stimulating factor-1
receptor
|
|
CSF-1/M-CSF
|
colony stimulating factor 1
|
|
NPC
|
nasopharyngeal carcinoma
|
|
RTK
|
receptor tyrosine kinase
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
BAX
|
Bcl-2-associated X protein
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
Akt/PKB
|
protein kinase B
|
|
PIP3
|
phosphatidylinositol-3,4,5-triphosphate
|
|
PDK1
|
phosphoinositide-dependent
kinase-1
|
|
CAT
|
central kinase catalytic domain
|
|
GSK-3
|
glycogen synthase kinase 3
|
References
|
1
|
Razak AR, Siu LL, Liu FF, Ito E,
O'Sullivan B and Chan K: Nasopharyngeal carcinoma: The next
challenges. Eur J Cancer. 46:1967–1978. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chua DT, Sham JS, Wei WI, Ho WK and Au GK:
The predictive value of the 1997 American Joint Committee on cancer
stage classification in determining failure patterns in
nasopharyngeal carcinoma. Cancer. 11:2845–2855. 2001. View Article : Google Scholar
|
|
3
|
Guo Y, Zhu XD, Qu S, Li L, Su F, Li Y,
Huang ST and Li DR: Identification of genes involved in
radioresistance of nasopharyngeal carcinoma by integrating gene
ontology and protein-protein interaction networks. Int J Oncol.
40:85–92. 2012.PubMed/NCBI
|
|
4
|
Yang S, Chen J, Guo Y, Lin H, Zhang Z,
Feng G, Hao Y, Cheng J, Liang P, Chen K, et al: Identification of
prognostic biomarkers for response to radiotherapy by DNA
microarray in nasopharyngeal carcinoma patients. Int J Oncol.
40:1590–1600. 2012.PubMed/NCBI
|
|
5
|
Huang L, Xu X, Hao Y, Chen J, Li L, Cheng
J, Chen Z, Liang W, Yang J and Ao W: Overexpression of CSF-1R in
nasopharyngeal carcinoma. Rom J Morphol Embryol. 56:1279–1283.
2015.PubMed/NCBI
|
|
6
|
Yu W, Chen J, Xiong Y, Pixley FJ, Dai XM,
Yeung YG and Stanley ER: CSF-1 receptor structure/function in
MacCsflr-/-macrophages: Regulation of proliferation,
defferetiation, and morphology. J Leukoc Biol. 84:852–863. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Beck AH, Espinosa I, Edris B, Li R,
Montgomery K, Zhu S, Varma S, Marinelli RJ, van de Rijn M and West
RB: The macrophage colony stimulating factor-1 response signature
in breast carcinoma. Clin Cancer Res. 15:778–787. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pixley FJ: Macrophage migration and its
regulation by CSF-1. Int J Biochem Cell B. 2012:5019622012.
|
|
9
|
Swierczak A, Cook AD, Lenzo JC, Restall
CM, Doherty JP, Anderson RL and Hamilton JA: The promotion of
breast cancer metastasis caused by inhibition of CSF-1R/CSF-1
signaling is blocked by targeting the G-CSF receptor. Cancer
Immunol Res. 2:765–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gruessner C, Gruessner A, Glaser K,
Abushahin N, Laughren C, Zheng W and Chambers SK: Biomarkers and
endosalpingiosis in the ovarian and tubal microenvironment of women
at high-risk for pelvic serous carcinoma. Am J Cancer Res. 4:61–72.
2014.PubMed/NCBI
|
|
11
|
Kuropkat C, Dünne AA, Plehn S, Ossendorf
M, Herz U, Renz H and Werner JA: Macrophage colony-stimulating
factor as a tumor marker for squamous cell carcinoma of the head
and neck. Tumour Biol. 24:236–240. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gazdar AF and Minna JD: Deregulated EGFR
signaling during lung cancer progression: Mutations, amplicons, and
autocrine loops. Cancer Prev Res (Phila). 1:156–160. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kirma N, Luthra R, Jones J, Liu YG, Nair
HB, Mandava U and Tekmal RR: Overexpression of the colony
stimulating factor (CSF-1) and/or its receptor c-fms in mammary
glands of transgenic mice results in hyperplasia and tumor
formation. Cancer Res. 64:4162–4170. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Torti D and Trusolino L: Oncogene
addiction as a foundational rationale for targeted anticancer
therapy: Promises and perils. EMBO Mol Med. 3:623–636. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kogan M, Haine V, Ke Y, Wigdahl B,
Fischer-Smith T and Rappaport J: Macrophage colony stimulating
factor regulation by nuclear factor kappa B: A relevant pathway in
human immunodeficiency virus type 1 infected macrophages. DNA Cell
Biol. 31:280–289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu W, Xu GZ, Jiang CH and Tian J:
Macrophage colony-stimulating factor and its receptor signaling
augment glycated albumin-induced retinal microglial inflammation in
vitro. BMC Cell Biol. 12:52011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Achkova D and Maher J: Role of the
colony-stimulating factor (CSF)/CSF-1 receptor axis in cancer.
Biochem Soc Trans. 44:333–341. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Foster P, Yamaguchi K, Hsu PP, Qian F, Du
X, Wu J, Won KA, Yu P, Jaeger CT, Zhang W, et al: The selective
PI3K inhibitor XL147 (SAR245408) inhibits tumor growth and survival
and potentiates the activity of chemotherapeutic agents in
preclinical tumor models. Mol Cancer Ther. 14:931–940. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li XR, Liu M, Zhang YJ, Wang JD, Zheng YQ,
Li J, Ma B and Song X: Evaluation of ER, PgR, HER-2, Ki-67, cyclin
D1, and nm23-H1 as predictors of pathological complete response to
neoadjuvant chemotherapy for locally advanced breast cancer. Med
Oncol. 28(Suppl 1): S31–S38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He Y, Liu Z, Qiao C, Xu M, Yu J and Li G:
Expression and significance of Wnt signaling components and their
target genes in breast carcinoma. Mol Med Rep. 9:137–143. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kanazawa T, Misawa K, Misawa Y, Uehara T,
Fukushima H, Kusaka G, Maruta M and Carey TE: G-protein-coupled
receptors: Next generation therapeutic targets in head and neck
cancer? Toxins (Basel). 7:2959–2984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Czapiewski P, Wełnicka-Jaśkiewicz M,
Seroczyńska B, Skokowski J, Sejda A, Szade J, Wiewiora C, Biernat W
and Żaczek A: CD99 correlates with low cyclin D1, high
topoisomerase 2 status and triple negative molecular phenotype but
is prognostically irrelevant in breast carcinoma. Pol J Pathol.
66:269–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Q, Chen X, Zhang X, Zhan J and Chen
J: Knockdown of TMEM14A expression by RNAi inhibits the
proliferation and invasion of human ovarian cancer cells. Biosci
Rep. 36:e002982016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen Y, Jiang T, Shi L and He K: hcrcn81
promotes cell proliferation through Wnt signaling pathway in
colorectal cancer. Med Oncol. 33:32016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao Z, Wang L, Song W, Cui H, Chen G,
Qiao F, Hu J, Zhou R and Fan H: Reduced miR-29a-3p expression is
linked to the cell proliferation and cell migration in gastric
cancer. World J Surg Oncol. 13:1012015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee MS, Ha JH, Yoon HS, Lee CK and Chi SW:
Structural basis for the conserved binding mechanism of
MDM2-inhibiting peptides and anti-apoptotic Bcl-2 family proteins.
Biochem Biophys Res Commun. 445:120–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Edison N, Curtz Y, Paland N, Mamriev D,
Chorubczyk N, Haviv-Reingewertz T, Kfir N, Morgenstern D,
Kupervaser M, Kagan J, et al: Degradation of Bcl-2 by XIAP and ARTS
promotes apoptosis. Cell Rep. 21:442–454. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou L, Gao R, Wang Y, Zhou M and Ding Z:
Loss of BAX by miR-365 promotes cutaneous squamous cell carcinoma
progression by suppressing apoptosis. Int J Mol Sci. 18(pii):
E11572017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song S, Jacobson KN, McDermott KM, Reddy
SP, Cress AE, Tang H, Dudek SM, Black SM, Garcia JG, Makino A and
Yuan JX: ATP promotes cell survival via regulation of cytosolic
[Ca2+] and Bcl-2/Bax ratio in lung cancer cells. Am J Physiol Cell
Physiol. 310:C99–C114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singh SS, Yap WN, Arfuso F, Kar S, Wang C,
Cai W, Dharmarajan AM, Sethi G and Kumar AP: Targeting the PI3K/Akt
signaling pathway in gastric carcinoma: A reality for personalized
medicine? World J Gastroenterol. 21:12261–12273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mahajan K and Mahajam NP: PI3K-Independent
AKT activation in cancers: A Treasure trove for novel therapeutics.
J Cell Physiol. 227:3178–3184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tan AC, Vyse S and Huang PH: Exploiting
receptor tyrosine kinase co-activation for cancer therapy. Drug
Discov Today. 22:72–84. 2017. View Article : Google Scholar : PubMed/NCBI
|