Introduction
Renal cell carcinoma (RCC) accounts for 2–3% of
solid tumors occurring annually and has a relatively high rate of
cure if detected early while still confined to the kidney. However,
overall mortality is high once the tumor has spread to secondary
sites (1,2). Thus, early detection is key to long-time
survival. Recent studies have focused on microRNAs (miRNAs) as
potential diagnostic and prognostic indicators of several types of
cancer since these small non-coding RNAs often act as either
oncogenes or tumor suppressors and also since the content of a
specific miRNA can vary greatly between a tumor and its tissue of
origin (3–5). Such miRNAs can be measured in several
readily accessible biofluids (serum, saliva, and urine) where they
exist either in a ‘free’ state, bound to specific proteins such as
Argonaute, or enclosed within membrane-delimited vesicles (exosomes
and other microvesicles) released by normal tissues and tumor cells
(5).
To date, several miRNA species have been proposed as
having diagnostic and prognostic value in renal cell carcinoma,
where either as solitary biomarkers or as part of a diagnostic
panel, they appear able to distinguish clear cell renal cell
carcinoma (ccRCC) patients from healthy controls and benign renal
tumors from malignant ones (6). Most
of these panels have been developed from serum samples (7–11), but two
recent studies have focused on urine specimens as a source of
diagnostic information (12,13). Urinary miRNAs are particularly
attractive as RCC biomarkers since urine can be sampled frequently
by non-invasive means.
The 786-O cell line originated from a human ccRCC of
proximal tubule origin, and is frequently employed in in
vitro studies of ccRCC cell behavior (14). Among the characteristics
distinguishing the 786-O line from several other ccRCC cultures is
a stable Von Hippel Lindau (VHL) mutation that results in
overexpression of vascular endothelial growth factor (VEGF)
(14). HK-2 cells are an immortalized
cell line originating from normal human proximal tubule (15).
In the present study we employed 786-O and HK-2 cell
monolayers as the in vitro equivalents of ccRCC tumor and
surrounding renal cortical tissue, respectively, to assess the
correlation between the content of selected miRNAs extracted from
secreted exosomes with miRNA derived from the cell monolayers. Our
results suggest that those miRNAs (e.g., miR-150 and miR-205) that
are of high concentration in exosomes relative to cytoplasmic
concentration and that have the highest levels of differential
expression between tumor and non-tumor tissue could be particularly
useful as biomarkers of RCC in urinary samples.
Materials and methods
Cell culture
786-O cells (ATCC®CRL-1932™)
and HK-2 cells (ATCC®CRL-2190™) were
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and subcultured in a growth medium containing
RPMI-1640 medium, fetal bovine serum (FBS) at a final concentration
of 10%, and penicillin (100 U/ml)-streptomycin (100 µg/ml). For
cell and exosome collection, cultures of HK-2 and 786-O cells grown
to confluence in growth medium in 162 cm2 flasks were
switched to 20 ml serum-free RPMI-1640 for 48 h after which
exosomes were collected and miRNA extracted from those exosome
pellets and from the cell monolayer using the methods described
below.
Exosome isolation
For all studies, purified exosomes were collected
from medium using two steps of ultracentrifugation as described
previously (16). Briefly,
conditioned medium was removed from 786-O and HK-2 cultures and the
supernatant was then frozen at −80°C until exosome isolation. The
cellular monolayer was then washed with PBS, scraped from the flask
and frozen at −80°C until miRNA extraction. To isolate exosomes,
the frozen medium was thawed and centrifuged at 17,000 × g for 18
min to pellet larger organelles and other membrane structures out,
followed by a final centrifugation at 200,000 × g for 1 h and 15
min, and collection of the pellet (exosomal fraction) and
supernatant for miRNA extraction.
miRNA extraction
MiRNA was extracted from both exosome and whole cell
pellets using the miRNeasy Micro kit (Qiagen Inc., Germantown, MD,
USA). Briefly, exosome pellets in polycarbonate ultracentrifuge
tubes were suspended in 25 ul phosphate buffered saline (PBS) and
transferred to 1.5 ml Eppendorf tubes to which 700 ul QIAzol lysis
reagent was added and homogenized by pulling repeatedly through a
hypodermic needle. Following the addition of 140 ul chloroform to
the homogenate and centrifugation (12,000 × g, 15 min), 1.5 volumes
of 100% ethanol was added to the supernatant and the sample was
added to an RNAeasy minElute spin column and centrifuged (8,000 ×
g, 30 sec). The column was then washed with RWT buffer, followed by
RPE buffer, and 80% ethanol, and purified miRNA was eluted with 15
µl RNAse-free dH2O and centrifugation at full speed for
2 min. Total RNA content was determined using the nano-drop
method.
miRNA was extracted from whole cell pellets using a
similar technique with the following modifications: After
dissolving the entire cell pellet in 700 ul lysis buffer, 35 ul of
the homogenate was removed and added to 630 ul lysis buffer (i.e.,
1:20 dilution), and the diluted homogentate was then processed as
described for exosome miRNA extraction. MiRNA was extracted from a
1 ml sample of the supernatant and designated as the ‘exosome-free
supernatant’ (EFS).
miRNA PCR Array
An miRNA PCR array (Cancer PathwayFinder miRNA PCR
Array; Qiagen Inc.) containing a total of 84 miRNA primers that had
previously been shown as differentially expressed in a variety of
tumor types was used to make an initial selection of miRNAs
differently expressed in 786-O cells compared to HK-2 cells.
MiRNA was extracted from collected exosomes as
described above, and cDNA was produced from a total of 300 ng RNA
from each sample in a reaction volume of 20 ul using the miScript
II miRNA RT kit (Qiagen Inc.). A total of 19 ul of each reaction
product was added to 200 ul H2O and 100 µl of each
diluted cDNA template was added to a common reaction mix containing
2×QuantiTect SYBR-Green PCR Master Mix, 10× miScript Universal
Primer, RNase-free water, and 25 ul of the total mix was pipetted
into each well of a 96-well Cancer PathwayFinder miRNA PCR Array
plate.
RT-qPCR
Reverse transcription (RT) of extracted miRNA was
performed to confirm the array results using a TaqMan MicroRNA
Reverse Transcription kit (Thermo Fisher Scientific, Inc., Waltham,
MA) and stem-loop oligonucleotides specific for the following
miRNAs (mature miRNA sequences in parentheses): miR-15a-5p
(UAGCAGCACAUAAUGGUUUGUG), miR-15a-3p (CAGGCCAUAUUGUGCUGCCUCA),
miR-16-5p (UAGCAGCACGUAAAUAUUGGCG), miR-150-5p
(UCUCCCAACCCUUGUACCAGUG), miR-34a-5p (UGGCAGUGUCUUAGCUGGUUGU),
miR-210-3p (CUGUGCGUGUGACAGCGGCUGA), miR-155-5p
(UUAAUGCUAAUCGUGAUAGGGGU), and miR-205-5p (UCCUUCAUUCCACCGGAGUCUG)
purchased from Thermo Fisher Scientific, Inc. Briefly, RT reactions
contained 20 ng of purified miRNA in a volume of 5 ul, and 0.15 ul
100 M dNTP mix w/dTT, 1.5 ul 10X RT buffer, 0.19 µl RNase inhibitor
(20 U/ul), 1 µl MultiScribe RT enzyme (50 U/µl), and 3 µl 5X
specific primer in a total volume of 15 ul. Reverse transcription
was performed in a programmable thermal cycler as follows: 16°C-30
min, 42°C 30 min, and 85°C-5 min, after which 1.33 ul of RT
reaction product was added to 10 ul master mix, 7.7 ul
H2O, and 1 µl 20X specific primer in a total volume of
20 ul that was amplified by 50 cycles of 15 sec denature/60 sec
anneal/extend in a real-time thermal cycler.
MiR-16 was selected as a control due to the relative
constancy of its expression in various cultured cell lines in the
literature (17), and RT-qPCR for
this miRNA was performed in tandem with target miRNAs to determine
the optimal normalization procedure.
Data analysis
Statistical analysis was performed on studies
repeated at least three times (with separate cultures of confluent
cells) using GraphPad Prizm version 7 (GraphPad Software, La Jolla,
CA, USA). One-way ANOVA was performed, followed by Tukey's multiple
comparisons test, and P<0.05 was considered to indicate a
statistically significant difference.
Results
miRNA PCR Array
Although not comprehensive, this array contained
primarily miRNAs that have demonstrated an association with human
cancers in the literature, many of which have been identified as
being up-or downregulated in RCC, specifically. As of this writing,
most of the 84 miRNAs in this assay have been identified in
published studies as playing a role in RCC, either as oncogenes or
tumor suppressors in cultured cells, or as biomarkers in
vivo (18–23). Therefore, the probability of finding
differences in expression levels of miRNA in the two cell lines was
assumed to be high. Since the PCR array consisted of 84 miRNAs that
were roughly evenly distributed between oncogenes (upregulated in
cancer) and tumor suppressors (downregulated in cancer), expression
levels of individual miRNAs were determined by normalization of
specific Ct values to the mean Ct for all 84 miRNAs.
We selected for further study several miRNAs that
were increased by >2-fold in 786-O exosomes compared with HK-2
exosomes in the array, and whose difference in expression from
control was consistent with previous citations of upregulation in
either RCC tumors, RCC patient serum/plasma, and/or RCC patient
urine compared with control. Using both criteria, four miRNAs were
chosen for further study using Taqman RT-qPCR: miR-150-5p,
miR-34a-5p, miR-155-5p, and miR-210, increased by at 8-fold,
7-fold, 5-fold and 2-fold, respectively, in the 786-O cells
compared with HK-2 in the PCR array. In addition, miR-15a-5p was
examined due to its prior identification as a urinary marker of RCC
(12). MiR-205 was also of interest
because of its previous citations as a markedly up- or
downregulated miRNA in urogenital cancers (24), and RCC cell lines (25) compared to controls.
Normalization of the target genes in the PCR array
was accomplished by subtracting the mean Ct of all genes in each
plate from the Cq value of individual target miRNAs in the
respective plate (17). However, this
normalization method was not feasible to use with the limited
number of miRNAs selected for TaqMan RT-qPCR. Therefore, endogenous
miR-16, a gene frequently used as a housekeeping gene in miRNA
studies (17), was assessed for
differences in expression between the two cell types, and among the
three pools (exosome, exosome-free, and cytoplasmic) from which
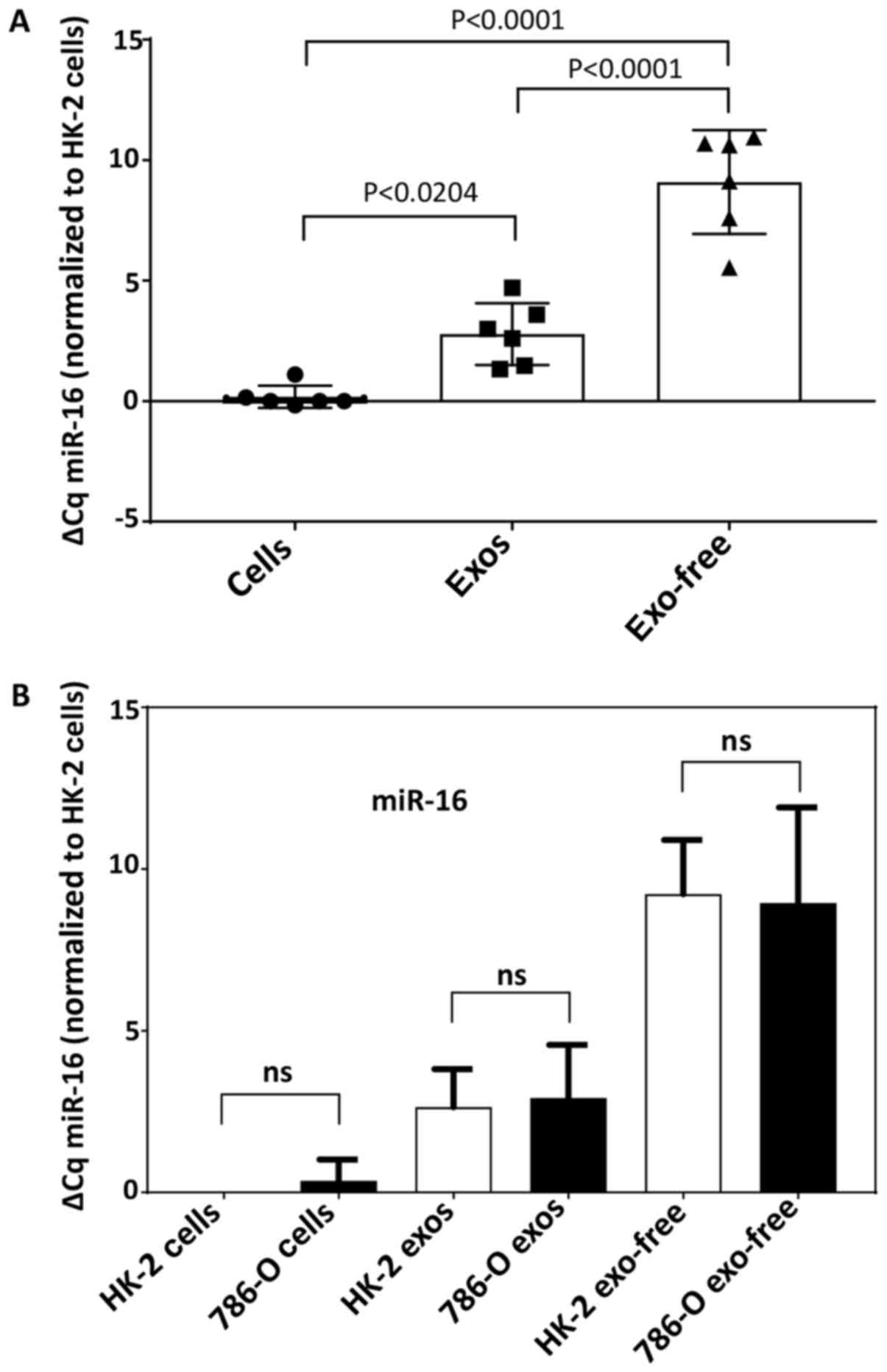
miRNA was extracted (Fig. 1). The
results showed significant differences in miR-16 Cq value among the
three pools, with whole cells exhibiting the lowest mean Cq (i.e.,
the highest concentration) and EFS exhibiting the highest Cq. Mean
miR-16 Cq, however, did not differ between HK-2 and 786-O cells for
any of the three pools (Fig. 1).
Since variability was least in the whole cell fraction, the miR-16
content of whole cells chosen for normalization in the studies
described below.
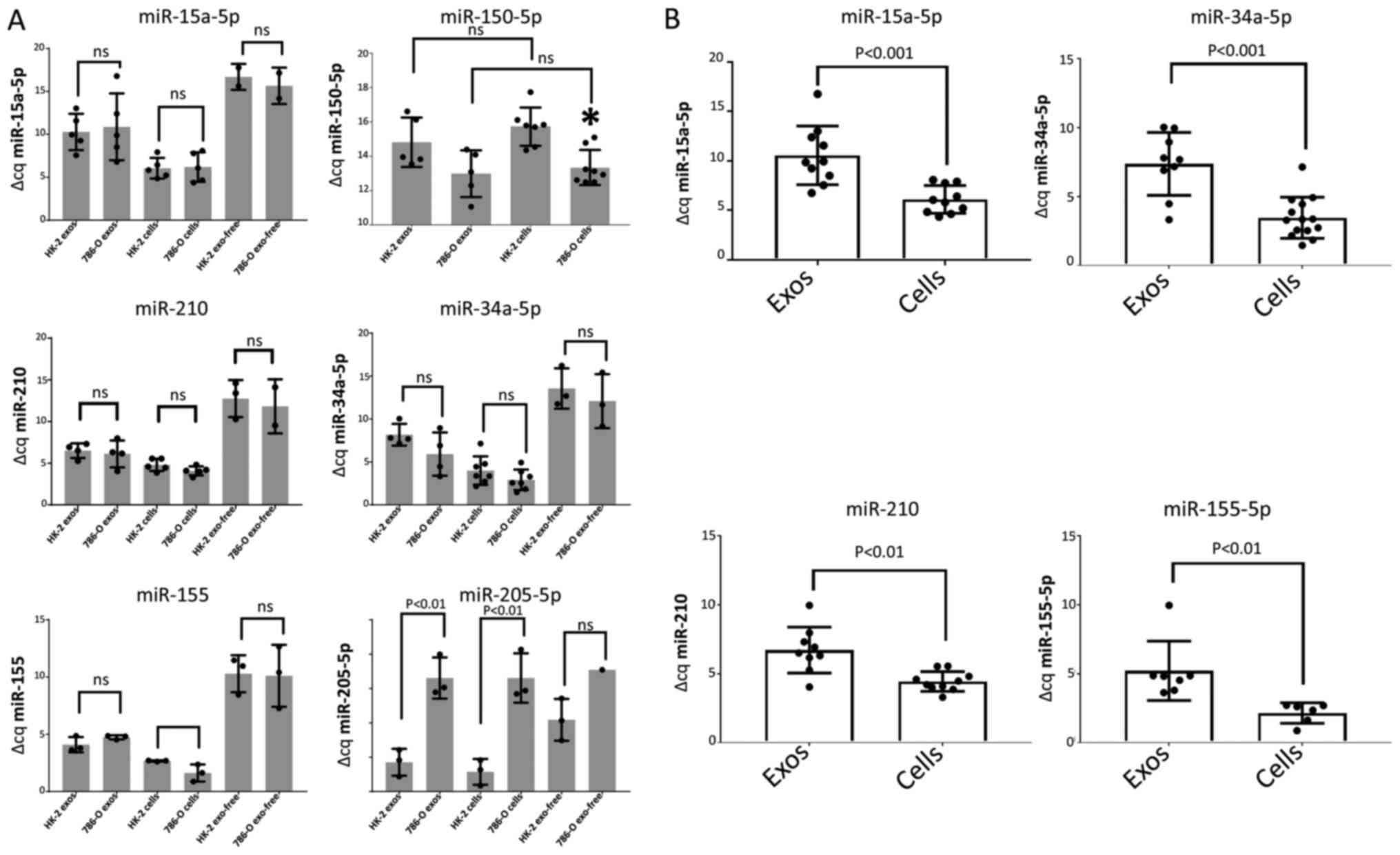
Fig. 2A shows the
results of RT-qPCR for selected mature miRNAs obtained from the
exosomes, exosome-free medium, or whole cells obtained from
confluent cultures of either HK-2 cells or 786-O cells, with all
results expressed as Δ Cq normalized to the miR-16 expression level
in whole cell pellets. MiR-15a-5p levels exhibited no difference
between the HK-2 and 786-O cell lines in either the exosomes,
exosome-free medium, or in the whole cells (Fig. 2A). Of the four miRNAs selected from
the array based on upregulation in 786-O cells and on previous
literature citations, miR-34a-5p, miR-155 and miR-210, showed
elevations in 786-O cells using TaqMan RT-qPCR consistent with the
array results, but the differences did not attain significance
(Fig. 2A). However, significant
differences were obtained between the exosomal concentration of
each of these miRNAs and its respective intracellular content when
results for HK-2 and 786-O cells were pooled (Fig. 2B), indicating that the secreted
exosomes contained only between 5 and 37% of the miRNA
concentration in the parent cells. The content of the target miRNAs
in the exosome and cell free fraction was even lower than this,
with miRNA levels ranging between undetectable (miR-150-5p) and
only 0.5% of the content of the whole cell.
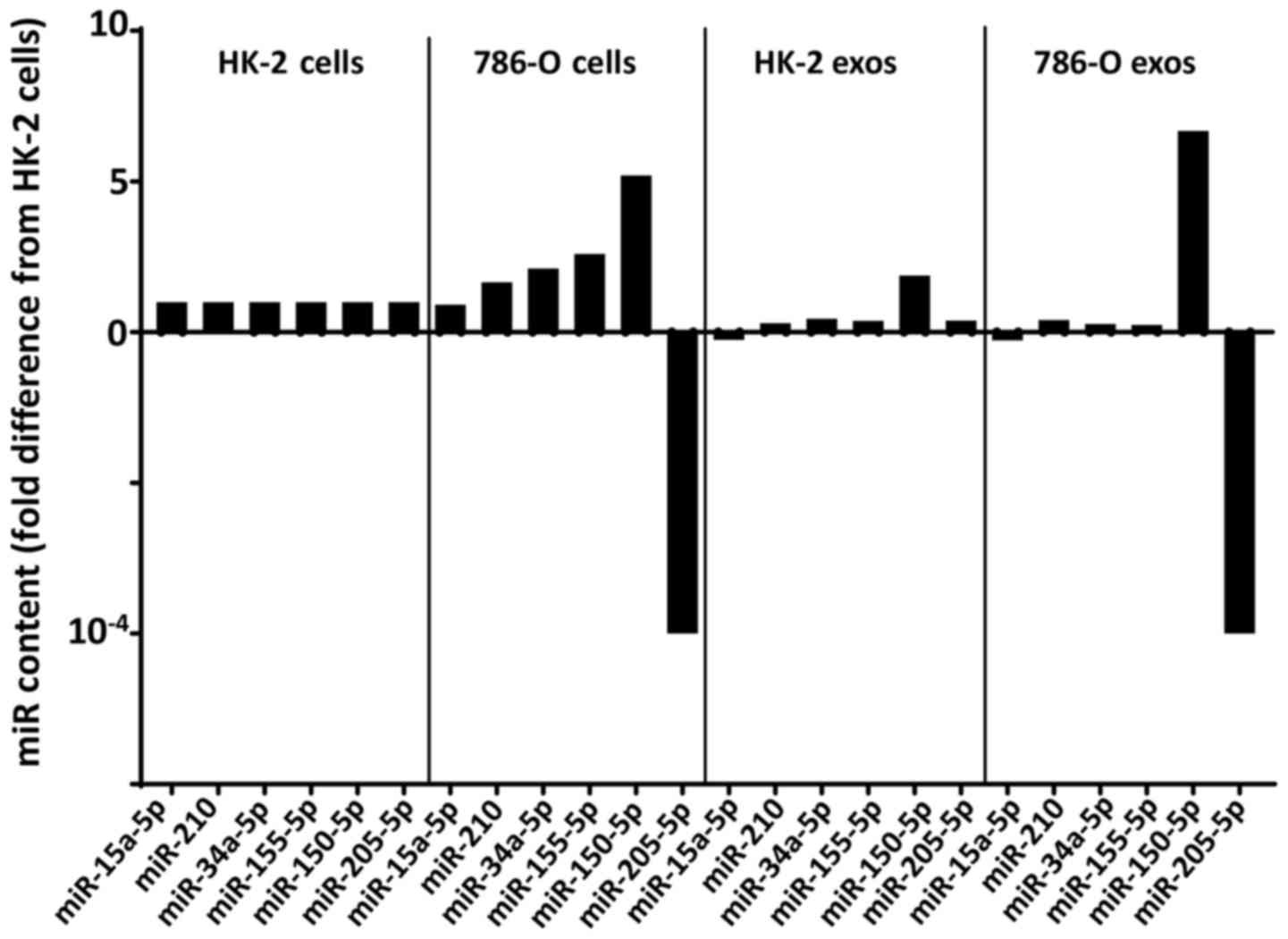
In contrast to these four miRNA species, both
miR-150 and miR-205 exhibited a significant difference in
intracellular expression levels between the two cell lines, with
miR-150 showing a 5.2-fold upregulation and miR-205 showing a
10,000-fold downregulation in 786-O cells relative to HK-2
(Figs. 2A and 3). Also distinguishing miR-150 from the
others was its higher content in the exosome fraction than in the
whole cell fraction in both HK-2 and 786-O cells. These two factors
may have contributed to the 3.6-fold increase of miR-150 in 786-O
exosomes compared to HK-2 cells, although the P-value for this
difference (P=0.10) did not reach significance (Fig. 3).
miR-205 was unlike all other miRNAs used in this
study in showing a significantly lower (10,000-fold) expression
level in 786-O cells than in HK-2 (P=0.0043). Most importantly, the
reduction in cellular miR-205 content was reflected in the exosomal
miR-205 content of the two cell lines with a 10,000-fold (P=0.0040)
lower level in 786-O cells than in HK-2.
Discussion
In the absence of a single unique biomarker for
early stage ccRCC, attention has recently focused on changes in
serum or urinary levels of multiple miRNA species (i.e., a miRNA
panel) to provide an early indication of the presence of this
cancer (7–13). There is a considerable lack of
uniformity, however, in the specific miRNAs that make up those
diagnostic panels. For example, of three such panels consisting of
2–5 miRNAs each (9–11), only a single miRNA (miR-378) was
present in more than one panel. Similarly, in the few studies
performed thus far of urinary miRNAs as potential biomarkers for
ccRCC, one has pointed to miR-15a as a particularly effective
indicator of disease (12), and the
other (13) has noted that miR-150-5p
is significantly upregulated in the urine of ccRCC patients.
To gain a better understanding of how secreted
miRNAs measured in readily accessible biofluids reflect the status
of those miRNAs in the tumor of origin, we have used in
vitro culture models of ccRCC and of normal renal proximal
tubule (786-O and HK-2 cells, respectively) to measure selected
miRNAs in secreted exosomes and EFS and compare their expression
levels with those in whole cells.
Our results show that even in a simplified in
vitro system where miRNA in secreted exosomes might be expected
to have a higher correlation with cellular miRNA content than serum
or urinary exosomes have with tumors in vivo, only exosomal
miR-150 and miR-205 were strongly correlated with cellular levels,
and of those two, only miR-205 reached significance.
In addition, miR-205 was the only miRNA that was
significantly reduced (~10,000-fold in both cells and exosomes) in
786-O cells compared to control HK-2 cells (Fig. 3). MiR-205 in the exosome-free fraction
was almost undetectable in either cell line. This result indicates
that collection of the exosome fraction and measurement of the
miR-205 content of that fraction can distinguish 786-O cells from
HK-2 cells in culture. However, whether exosomal miR-205 is a
potentially useful biomarker for detecting the presence of ccRCC
in vivo remains unclear. The relationship between the amount
of exosomal miR-205 secreted from a tumor in vivo and its
concentration in a particular biofluid is not as direct as the
relationship between the miR-205 content of cultured cells and the
overlying supernatant, and the detection of changes in biofluid
miR-205 from a growing tumor mass that possesses very little of
that miRNA could pose a challenge, especially in plasma or serum
samples. A recent large-scale study of plasma miRNAs in ccRCC
patients and controls did not list miR-205 as being among the most
differentially expressed, however the assays were performed on
plasma rather than exosome extracts as in the present study
(26).
In regard to the functional significance of this
finding, Hirata et al (25)
had observed the downregulation of miR-205 in both 786-O cells, and
in A-498 RCC cells compared to levels in HK-2 cells. Since both RCC
lines possess a mutated VHL gene (14), it is possible that the mutation could
be related to the reduced miR-205 expression both in the cancer
lines and in RCC tumors bearing the VHL mutation in vivo.
The biological relevance of miR-205 in RCC remains unclear
(27,28), but the targeting of several oncogenic
genes including VEGF-A (27) and
MALAT1 (25) by miR-205 suggests that
it normally acts as a tumor suppressor that is inactivated in
RCC.
MiR-150, the miRNA that was identified in the PCR
microarray assay as displaying the greatest increase (8-fold) in
786-O cells was the only miRNA to show significant upregulation
(5.2-fold) in 786-O cells using RT-qPCR. Moreover, the 3.3-fold
difference in exosomal miRNA content between HK-2 and 786-O cells,
although not reaching significance, reflected the difference in
whole cell miR-150 content between the two cell types. Another
distinctive feature of miR-150 observed in this study was its
approximately two-fold higher concentration in exosomes than in
whole cell cytoplasm. This feature, shared with miR-451 and several
other miRNAs, is its preferential import into microvesicles
compared with other miRNAs (29), a
feature that might favor them as ccRCC biomarkers. The failure to
show a significant difference in exosomal miR-150 in the present
study could be due to the relatively small amounts of miRNA
extractable from exosomes compared to whole cells, and also to the
relatively high Cq value for miR-150-5p compared to the other
miRNAs. Future studies with additional ccRCC cell lines will
address these issues.
Like miR-150, miR-15a had also shown promise as a
urinary biomarker for RCC in a recent clinical study (12). However, our in vitro results
could discern no difference in miR-15a-5p levels between 786-O
cells and HK-2, either in the secreted exosomes or in the parent
cells, suggesting that the reported clinical changes in urinary
miR-15a-5p are unlikely to be due to increased expression in RCC
tumor cells.
In contrast to miR-150, a preferentially exported
miRNA, miR-15a was determined by Guduric-Fuchs et al
(29) to be preferentially retained
within cells rather than exported (29). This observation is supported by our
findings that the miR-15a-5p content of both HK-2 and 786-0 cells
is about 16-fold higher than its content in the exosomes secreted
from the corresponding cell line. MiR-34a, miR-210, and miR-155
also exhibited significantly lower levels in exosomes than in whole
cells (Fig. 3), and none of the four
was demonstrably different in concentration in exosomes between the
two cell lines.
In conclusion, the present study, limited initially
to 84 miRNA species important in cancer pathways and six miRNAs
selected for further study, shows that only two of the miRNAs
(miR-205 and miR-150) were sufficiently altered from control in
786-O cells to serve as exosomal markers of ccRCC, even in the
simplified in vitro model that we use here. One of these,
miRNA-150, is increased in both 786-O cells and its secreted
exosomes, in close agreement with differences in the exosomal
miR-150 content of patient and control urine (13), but opposite to differences in
circulating levels of miR-150 that were observed in earlier
clinical studies (10,26). In vitro observations of
cancer-associated differences in miR-150 (and other miRNA) content
in assays such as the present one could therefore be especially
relevant to the design of urinary biomarker assays. Furthermore,
urinary assays of exosomal miRNA may be more amenable to the
selection of downregulated ccRCC miRNAs as biomarkers owing
to the close contact of the intial tumor to the pathway to final
urine. Many of these downregulated miRNAs, like miR-205, could have
immense differences in expression levels between cancer and
non-cancer cells that would help make RCC easier to detect.
We suggest that comparison of miRNA content of
exosomes with miRNA content of whole cells in homogenous cultures
of specific strains of RCC could help to identify specific miRNAs
that are particularly useful urinary biomarkers for monitoring
changes in primary ccRCC and metastases.
Acknowledgements
Not applicable.
Funding
The present study was performed with the support of
USUHS (grant no. R083406616).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
VCC conceived the project idea and executed
experiments; HL designed and executed experiments; DFS aided in the
design of the project and wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rini BI, Campbell SC and Escudier B: Renal
cell carcinoma. Lancet. 373:1119–1132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Greef B and Eisen T: Medical treatment of
renal cancer: New horizons. Br J Cancer. 115:505–516. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stahlhut Espinosa CE and Slack FJ: The
role of microRNAs in cancer. Yale J Biol Med. 79:131–140.
2006.PubMed/NCBI
|
|
4
|
Wuchty S, Arjona D, Bozdag S and Bauer PO:
Involvement of microRNA families in cancer. Nuc Acids Res.
40:8219–8226. 2012. View Article : Google Scholar
|
|
5
|
Wang J, Zhang KY, Liu SM and Sen S:
Tumor-associated circulating microRNAs as biomarkers of cancer.
Molecules. 19:1912–1938. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sellitti DF and Doi SQ: MicroRNAs in renal
cell carcinoma. Microrna. 4:26–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iwamoto H, Kanda Y, Sejima T, Osaki M,
Okada F and Takenaka A: Serum miR-210 as a potential biomarker of
early clear cell renal cell carcinoma. Int J Oncol. 44:53–58. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wulfken LM, Moritz R, Ohlmann C,
Holdenrieder S, Jung V, Becker F, Herrmann E, Walgenbach-Brünagel
G, von Ruecker A, Müller SC and Ellinger J: MicroRNAs in renal cell
carcinoma: Diagnostic implications of serum miR-1233 levels. PLoS
One. 6:e257872011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang C, Hu J, Lu M, Gu H, Zhou X, Chen X,
Zen K, Zhang CY, Zhang T, et al: A panel of five serum miRNAs as a
potential diagnostic tool for early-stage renal cell carcinoma. Sci
Rep. 5:76102015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Redova M, Poprach A, Nekvindova J, Iliev
R, Radova L, Lakomy R, Svoboda M, Vyzula R and Slaby O: Circulating
miR-378 and miR-451 in serum are potential biomarkers for renal
cell carcinoma. J Transl Med. 10:552012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hauser S, Wulfken LM, Holdenrieder S,
Moritz R, Ohlmann CH, Jung V, Becker F, Herrmann E,
Walgenbach-Brünagel G, von Ruecker A, et al: Analysis of serum
microRNAs (miR-26a-2*, miR-191, miR-337-3p and miR-378) as
potential biomarkers in renal cell carcinoma. Canc Epidemiol.
36:391–394. 2012. View Article : Google Scholar
|
|
12
|
von Brandenstein M, Pandarakalum JJ, Kroon
L, Loeser H, Herden J, Braun G, Wendland K, Dienes HP, Engelmann U
and Fries JW: MicroRNA 15a, inversely correlated to PKCα, is a
potential marker to differentiate between benign and malignant
renal tumors in biopsy and urine samples. Am J Pathol.
180:1787–1797. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Butz H, Nofech-Mozes R, Ding Q, Khella
HWZ, Szabó PM, Jewett M, Finelli A, Lee J, Ordon M and Stewart R:
et alExosomal microRNAs are diagnostic biomarkers and can
mediate cell-cell communication in renal cell carcinoma. Eur Urol
Focus. 2:210–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brodaczewska KK, Szczylik C, Fiedorowicz
M, Porta C and Czarnecka AM: Choosing the right cell line for renal
cancer research. Mol Canc. 15:832016. View Article : Google Scholar
|
|
15
|
Ryan MJ, Johnson G, Kirk J, Fuerstenberg
SM, Zager RA and Torok-Storb B: HK-2: An immortalized proximal
tubule epithelial cell line from normal adult human kidney. Kidney
Int. 45:48–57. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vlasov P, Doi SQ and Sellitti DF: FRTL-5
rat thyroid cells release thyroglobulin sequestered in exosomes: A
possible novel mechanism for thyroglobulin processing in the
thyroid. J Thyroid Res. 2016:92764022016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schwarzenbach H, da Silva AM, Calin G and
Pantel K: Data normalization strategies for microRNA
quantification. Clin Chem. 61:1333–1342. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang Y, Dai Y, Yang J, Chen T, Yin Y,
Tang M, Hu C and Zhang L: Microarray analysis of microRNA
expression in renal clear cell carcinoma. Eur J Surg Oncol.
35:1119–1123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Juan D, Alexe G, Antes T, Liu H,
Madabhushi A, Delisi C, Ganesan S, Bhanot G and Liou LS:
Identification of a microRNA panel for clear-cell kidney cancer.
Urology. 75:835–841. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jung M, Mollenkopf HJ, Grimm C, Wagner I,
Albrecht M, Waller T, Pilarsky C, Johannsen M, Stephan C, Lehrach
H, et al: MicroRNA profiling of clear cell renal cell cancer
identifies a robust signature to define renal malignancy. J Cell
Mol Med. 13:3918–3928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakada C, Matsuura K, Tsukamoto Y,
Tanigawa M, Yoshimoto T, Narimatsu T, Nguyen LT, Hijiya N, Uchida
T, Sato F, et al: Genome-wide microRNA expression profiling in
renal cell carcinoma: Significant down-regulation of miR-141 and
miR-200c. J Pathol. 216:418–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Petillo D, Kort EJ, Anema J, Furge KA,
Yang XJ and The BT: MicroRNA profiling of human kidney cancer
subtypes. Int J Oncol. 35:109–114. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weng L, Wu X, Gao H, Mu B, Li X, Wang JH,
Guo C, Jin JM, Chen Z, Covarrubias M, et al: MicroRNA profiling of
clear cell renal cell carcinoma by whole-genome small RNA deep
sequencing of paired frozen and formalin-fixed, paraffin-embedded
tissue specimens. J Pathol. 222:41–51. 2010.PubMed/NCBI
|
|
24
|
Huang X, Liang M, Dittmar R and Wang L:
Extracellular microRNAs in urologic malignancies: Chances and
challenges. Int J Mol Sci. 14:14785–14799. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hirata H, Hinoda Y, Shahryari V, Deng G,
Nakajima K, Tabatabai ZL, Ishii N and Dahiya R: Long noncoding RNA
MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and
interacts with miR-205. Canc Res. 75:1322–1331. 2015. View Article : Google Scholar
|
|
26
|
Chanudet E, Wozniak MB, Bouaoun L, Byrnes
G, Mukeriya A, Zaridze D, Brennan P, Muller DC and Scelo G:
Large-scale genome-wide screening of circulating microRNAs in clear
cell renal cell carcinoma reveals specific signatures in late-stage
disease. Int J Cancer. 141:1730–1740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Orang AV, Safaralizaden R and Hosseinpour
Feizi MA: Insights into the diverse roles of miR-205 in human
cancers. Asian Pac J Cancer Prev. 15:577–583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qin AY, Zhang XW, Liu L, Yu JP, Li H, Wang
SZE, Ren XB and Cao S: Mir-205 in cancer: An angel or a devil? Eur
J Cell Biol. 92:54–60. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guduric-Fuchs J, O'Conner A, Camp B,
O'Neill CL, Medina RJ and Simpson DA: Selective extracellular
vesicle-mediated export of an overlapping set of microRNAs from
multiple cell types. BMC Genomics. 13:3572012. View Article : Google Scholar : PubMed/NCBI
|