Introduction
Epithelial ovarian cancer (EOC) is the fifth most
common malignancy in women, with a 5-year mortality of >70% in
North America (1). Combined surgery
and cytotoxic therapy have previously been demonstrated to produce
a favorable clinical response in 50–80% of patients (2). The platinum-paclitaxel combination
regimen is utilized as the principal chemotherapy for advanced EOC,
and yields response rates of >80% and complete response rates of
40–60% (3–7). Owing to the symptoms often not being
observed until the cancer has spread extensively, <25% of women
are diagnosed at an early stage of disease (2). Large-scale gene expression analyses have
identified molecular subtypes within high-grade ovarian cancer with
variable survival rates and drug resistance (8,9). These
variations in gene expression may influence the outcomes of
different treatment types. The understanding of gene expression,
the mechanisms underlying cancer processes and drug resistances
have facilitated the development of targeted therapies, such as
small-molecule inhibitors. These agents target the proteins
associating with cell viability/death, metastasis and angiogenesis.
Previous studies have demonstrated breast cancer 1/2 (BRCA1/2) and
DNA repair proteins, as markers for platinum treatment (8–10).
Ribonucleotide reductase subunit M1 (RRM1) and DNA topoisomerase
II-α (TOP2A) have been used as markers to predict the response to
gemcitabine and anthracyclines for patients with ovarian cancer
(11–13).
The far-upstream element (Fuse)-binding protein 1
(FBP1) has been identified as an anti-apoptotic and
pro-proliferative oncoprotein that is overexpressed in a number of
malignancies, including hepatocellular carcinoma (HCC) (14,15), non
small-cell lung cancer (16) and
colorectal cancer (17). Importantly,
nuclear enrichment of FBP1 is associated with poor overall survival
of patients with liver cancer (14).
FBP1 functions as a transcriptional regulator by binding to the
single-stranded DNA element (fuse) and interacting with the basal
transcriptional machinery (18,19). The
study of Rabenhorst et al (15) previously demonstrated that FBP1 serves
a role in hematopoietic development and/or homeostasis. In our
early studies, FBP1 was demonstrated to physically interact with
p53 and suppress p53 transcription activity under radiation-induced
cellular stress and facilitates hepatitis C virus replication in
hepatoma cells (20,21). Silence of FBP1 increases the
sensitivity of ovarian cancer cells to carboplatin (22).
Since FBP1 is always overexpressed in certain
malignancies, the present study aimed to clarify the association
between FBP1 expression and EOC development. We hypothesized that
FBP1 enhances EOC development and that FBP1 is a novel potential
biological marker for EOC diagnosis. The study also attempted to
analyze the potential mechanisms underlying the promotion of FBP1
in EOC development.
Materials and methods
Patients and samples collection
The present study was conducted after informed
consent was obtained from all subjects and the protocol was
approved by the Medical Ethics Committee of Guangzhou Red Cross
hospital of Medical College, Jinan University (Guangzhou, China).
For immunohistochemical analysis, a total of 58 ovarian specimens
were obtained from the Department of Pathology, Guangzhou Red Cross
Hospital from January 2010 to June 2015 with a median age of 47.6
from 17.0–76.0 years old and assigned into three groups, including
normal epithelial ovarian tissues (14 samples), epithelial ovarian
adenoma tissues (25 samples) and epithelial ovarian cancer tissues
(19 samples). None of the patients received preoperative therapies
such as radiation, chemotherapy, or immunotherapy. All tissues were
fixed with 10% formalin and then embedded in paraffin prior to
sectioning.
Antibodies and reagents
The antibodies used for immunohistochemistry and
western blot analysis were as follows: GAPDH (catalog no. 5174;
1:2,000) and FBP1 (cat. no. 72736; 1:1,000) antibodies were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Ki-67 (cat. no. 550609; 1:200) was obtained from BD Biosciences
(Franklin Lakes, NJ, USA). Cyclin D1 (cat. no. sc-70899; 1:500),
cyclin E (cat. no. sc-377100; 1:500), c-Myc (cat. no. sc-398624;
1:500), p21 (cat. no. sc-817; 1:500), p27 (cat. no. sc-1641; 1:500)
and matrix metalloproteinase-2 (MMP-2; cat. no. sc-13594; 1:500)
were obtained from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum
(FBS) and L-glutamine, were obtained from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). The CellTiter 96®
AQueous One Solution Reagent
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium,
inner salt; MTS] was purchased from Promega Corporation (Madison,
WI, USA). Penicillin and streptomycin sulfate were purchased from
GE Healthcare Life Sciences (Logan, UT, USA).
Cell culture and FBP1 knockdown cell
construction
The human ovarian cancer SKOV3 cells (Chinese
Academy of Sciences Cell Bank) were cultured in DMEM with 10% (v/v)
FBS, 2 mM L-glutamine, 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C and 5% CO2 in a humidified
incubator.
FBP1-knockdown lentiviral particles (sc-43760) and
control lentiviral particles (cat. no. sc-108080) were obtained
from Santa Cruz Biotechnology, Inc. SKOV3 cells were plated in
complete DMEM containing 10% (v/v) FBS until they reached 70%
confluence. Subsequently, cells were transfected with 20 µl of
either control lentiviral particles or FBP1-knockdown lentiviral
particles (1×106 IFU) in serum-free medium according to
the manufacturer's protocol. The transfection medium was replaced
by fresh complete DMEM after 6 h and the cells were incubated for
another 48 h at 37°C. Cells were collected after screening with 2.0
µg/ml puromycin for ~2 weeks and termed FBP1-knockdown (FBP1-KD)
and FBP1 control (FBP1-C) SKOV3 cells, respectively.
Cell viability and plate colony
formation assays
FBP1-C and FBP1-KD SKOV3 cells were plated into a
96-well plate at a density of 1×104 cells/well and
cultures for 24 h. Cell viability was measured using MTS in
accordance with the manufacturer's (Promega Corporation) protocol,
and the absorbance at the wavelength of 490 nm was read in an
automated plate reader (Bio Tek Instruments Inc., Winooski, VT,
USA). The experiments were repeated at least three times.
FBP1-C and FBP1-KD SKOV3 cells (2.0×104
cells/plate) were cultured in 5 ml DMEM supplemented with 10% FBS
in 10-cm plates. After 14 days, colonies were washed with PBS,
fixed with 10% methanol at 4°C for 30 min and stained with 1%
crystal violet for 30 min. The colony formation images were
captured by camera.
Immunohistochemical staining
Formalin-fixed paraffin embedded ovarian tissue
sections (5-µm thick) were deparaffinized (using 100% of turpentine
oil, three times for 10 min/time) and hydrated consecutively (100%
of ethanol for 3 min, 95% of ethanol for 3 min, 90% of ethanol for
3 min, 85% of ethanol for 3 min, 80% of ethanol for 3 min, 75% of
ethanol for 3 min and H2O for 3 min). The sections were
incubated with citrate buffer (pH 6.0) to retrieve the antigen
using a microwave cooker. Endogenous peroxidase activity was
quenched using 3% hydrogen peroxide (H2O2).
The sections were blocked with 10% BSA (Wuhan Boster Biological
Technology, Ltd., Wuhan, China) for 30 min at room temperature
prior to incubation with rabbit anti-human FBP1 or Ki-67 antibody
(diluted 1:500) overnight at 4°C, and then incubated with
HRP-conjugated secondary antibody (cat. no. 8114; 1:200; Cell
Signaling Technology) for 1 h at room temperature. After washing
with PBS, the reaction was visualized by incubating the sections
with DAB and hematoxylin for 5 min at room temperature. The
negative control section was incubated with blocking reagent alone
in the absence of primary antibody. Images were captured by an ×40
objective and image processing and analyses were performed using
the Image-Pro Plus 6.0 software (Media Cybernetics, Shanghai,
China). The intensity of the immunohistochemical reaction was
expressed as integral optical density (IOD) of DAB brown reaction
products. The results of 5 separate measurements for each of sample
were expressed as mean ± standard deviation (SD).
Western blot analysis
Cells were lysed for western blotting in modified
RIPA buffer (150 mM NaCl, 1% NP-40, 50 mM Tris-Cl [pH 8.0], 0.1%
SDS) supplemented with PMSF (1 mM) protease and phosphatase
inhibitor. Following rapid homogenization, the homogenates were
incubated in ice for 30 min and centrifuged at 12,000 × g for 15
min at 4°C. Protein concentrations were determined by bicinchoninic
acid protein assay (Pierce; Thermo Fisher Scientific, Inc.). The
protein samples (30–50 µg/lane) were resolved by SDS-PAGE (10–12%
gel) and transferred to polyvinylidene difluoride filter membranes
(Merck KGaA, Darmstadt, Germany). Following transfer, membranes
were blocked with 5% skimmed milk in TBS-Tween (0.05 M Tris, 0.15 M
NaCl, pH 7.5, 0.2% Tween-20) for 1 h at room temperature and then
incubated at 4°C overnight with primary antibodies of GAPDH, FBP1
(Cell Signaling Technology), Cyclin D1, Cyclin E, c-Myc, p21, p27
and MMP-2 (Santa Cruz). Membranes were washed with TBST for three
times and then incubated with anti-rabbit/mouse HRP-labeled
secondary antibodies for 1 h at room temperature, respectively, and
detected with ECL-Plus detection systems (Pierce; Thermo Fisher
Scientific, Inc.). Relative abundance was quantified by
densitometry using Quantity One 4.6.7 software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Cell cycle and cell apoptosis
analysis
Flow cytometry was used for the analysis of cell
cycle distribution and apoptosis. For cell cycle distribution,
FBP1-C and FBP1-KD SKOV3 cells were collected and fixed in 70%
(v/v) ethanol overnight at 4°C, and then incubated with 1 mg/ml
RNase A for 30 min at 37°C. Cells were stained with 50 mg/ml
propidium iodide (PI) (BD Biosciences) in PBS containing 1% Triton
X-100. The data were acquired using a BD FACSCAN flow cytometer
(FACSAria II, BD Biosciences) and analyzed using the BD CellQuest
Pro software (BD Biosciences). For apoptosis, FBP1-C and FBP1-KD
SKOV3 cells were firstly incubated with Annexin V-fluorescein
isothiocyanate (FITC) (Beijing Biosea Biotechnology Co., Ltd.,
Beijing, China) for 30 min at 4°C in the dark, and then incubated
with PI for 5 min. Analysis was immediately performed by FACSAria
II (BD Biosciences) according to the manufacturer's
instructions.
Wound healing and Transwell migration
assays
FBP1-C and FBP1-KD SKOV3 cells were seeded on 6-well
plates at a density of 2×105 cells/well. The cells were
scratched using a sterile 100 µl micropipette tip and washed twice
with PBS. The cells were cultured with complete DMEM with 10% FBS
for an additional 48 h. Images were captured by a microscope at the
0-, 24- and 48-h time-points.
The Transwell inserts (8 µm pore size, Corning
Incorporated, Corning, NY, USA) were coated with 0.1% gelatin for
30 min at 37°C. The bottom chambers of Transwell were filled with
DMEM with 10% FBS, and the top chambers were seeded
1×105 cells per well (pretreated with mitomycin C) in
200 µl DMEM (FBS free). After 24 h of incubation at 37°C, the
migrated cells were stained with 1% crystal violet for 30 min at
room temperature. Images were captured using an Olympus inverted
microscope (Olympus Corporation, Tokyo, Japan) at ×200
magnification.
Statistical analysis
The data were derived from at least three
independent experiments. Values reported are mean ± SD. The data
were analyzed using one-way analysis of variance and
Student-Newman-Keuls post-hoc test by the SPSS 17.0 statistical
package from SPSS Inc. (Chicago, IL, USA). The SPSS non-parametric
Spearman's rank correlation was used to evaluate the correlation
between FBP1 expression and the cell proliferation marker Ki-67.
P<0.05 was considered to indicate a statistically significant
difference.
Results
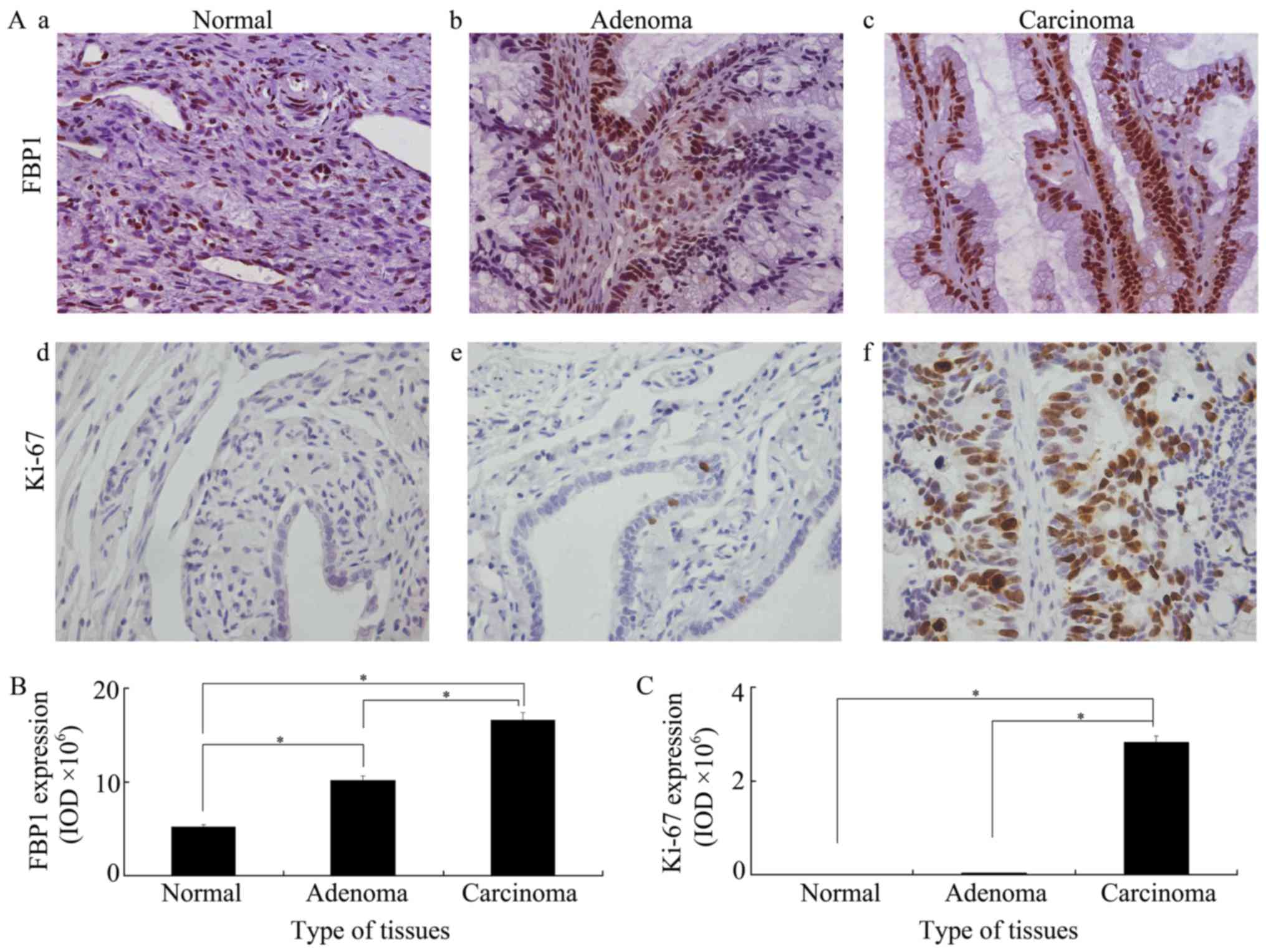
FBP1 overexpression in ovarian
tissues
To identify whether FBP1 is associated with the
development of ovarian cancer, FBP1 expression was analyzed in
epithelial ovarian normal, adenoma and carcinoma tissues by
immunohistochemistry. As shown in Fig.
1A-a/b/c, FBP1 expression was identified in all three groups.
FBP1 expression increased as cancer development and highest FBP1
expression was found in epithelial ovarian carcinoma tissues. A
significant higher expression of FBP1 was identified in adenoma
than in normal, or in carcinoma than in adenoma, or in carcinoma
than in normal tissues (Fig. 1B). The
expression of Ki-67, a key marker used to show cell proliferation,
was also examined in all three groups. Mimicking the changes
identified in FBP1, Ki-67 expression was increased in cancer
development and the highest Ki-67 expression was found in
epithelial ovarian cancer tissues. However, Ki-67 expression was
very low in normal and adenoma tissues (Fig. 1C). Furthermore, the correlation
between the expression of FBP1 and Ki-67 was analyzed by Spearman's
rank correlation. The correlation coefficient was 0.782 at the 0.01
P-value level. This meant that a significant positive correlation
was present between FBP1 and Ki-67. These data indicated that FBP1
is tightly associated with EOC development and cell
proliferation.
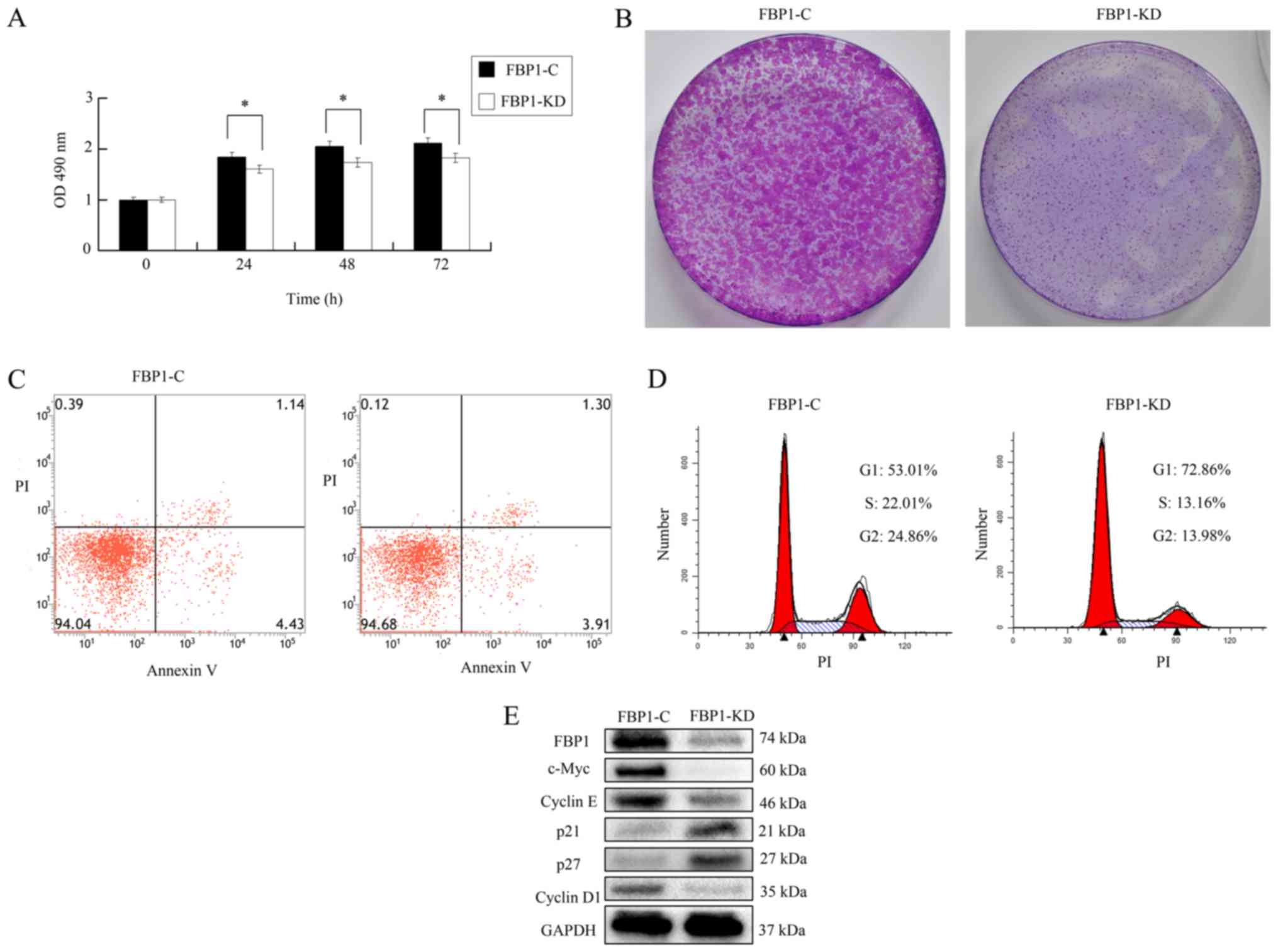
Knockdown of FBP1 influenced cell
proliferation
SKOV3 cells were transfected with control lentiviral
particles or FBP1 knockdown lentiviral particles obtained from
Santa Cruz, Biotechnology, Inc. screened with puromycin (2.0 µg/ml)
for 2 weeks and polyclonal control or FBP1 knockdown SKOV3 cells
were generated and named as FBP1-C and FBP-KD SKOV3 cells. As shown
in Fig. 2A, the proliferation of
FBP1-KD cells was significantly slower than that of the FBP1-C
cells according to MTS assay. The colonies of FBP1-KD were
significantly smaller than that of FBP1-C (Fig. 2B). In FBP1-C and FBP1-KD cells, the
total number of apoptotic cells was ~5.0%, as analyzed by
Annexin-V-FITC/PI flow cytometry (Fig. 2C
and D). As shown in Fig. 2E, the
expression of FBP1 in FBP1-KD cells was ~15% of that in FBP1-C
cells; this meant that the knockdown of FBP1 was successful. This
data implied that that FBP1-knockdown did not influence apoptosis
in SKOV3 cells. Based on these data, we believe that FBP1-knockdown
influences cell proliferation; however, it does not influence
apoptosis.
Knockdown of FBP1 inhibited cell cycle
transition
Since FBP1 knockdown influenced cell proliferation,
the effect of FBP1 on cell cycle progression was examined by flow
cytometry. As shown in Fig. 2D, the
percentage of cells in G1 phase was increased from 53.01% in FBP1-C
cells to 72.86% in FBP1-KD cells. In contrast, the percentage of
cells in S and G2 phase was decreased from 22.01 and 24.86% in
FBP1-C cells to 13.16 and 13.98% in FBP1-KD cells. These data
implied that FBP1 knockdown significantly inhibited cell cycle
transition from G1 to S and G2 phase. Additionally, the expressions
of proteins associated with cell cycle progression were analyzed.
The proteins promoting cell cycle progression, including c-Myc,
cyclin D1/E, were inhibited in FBP1-KD cells. However, proteins
inhibiting cell cycle progression, such as p21 and p27, were
increased in FBP1-KD cells (Fig. 2E).
These data demonstrated that FBP1 influences cell cycle transition
by adjusting the expression of cell cycle associated proteins.
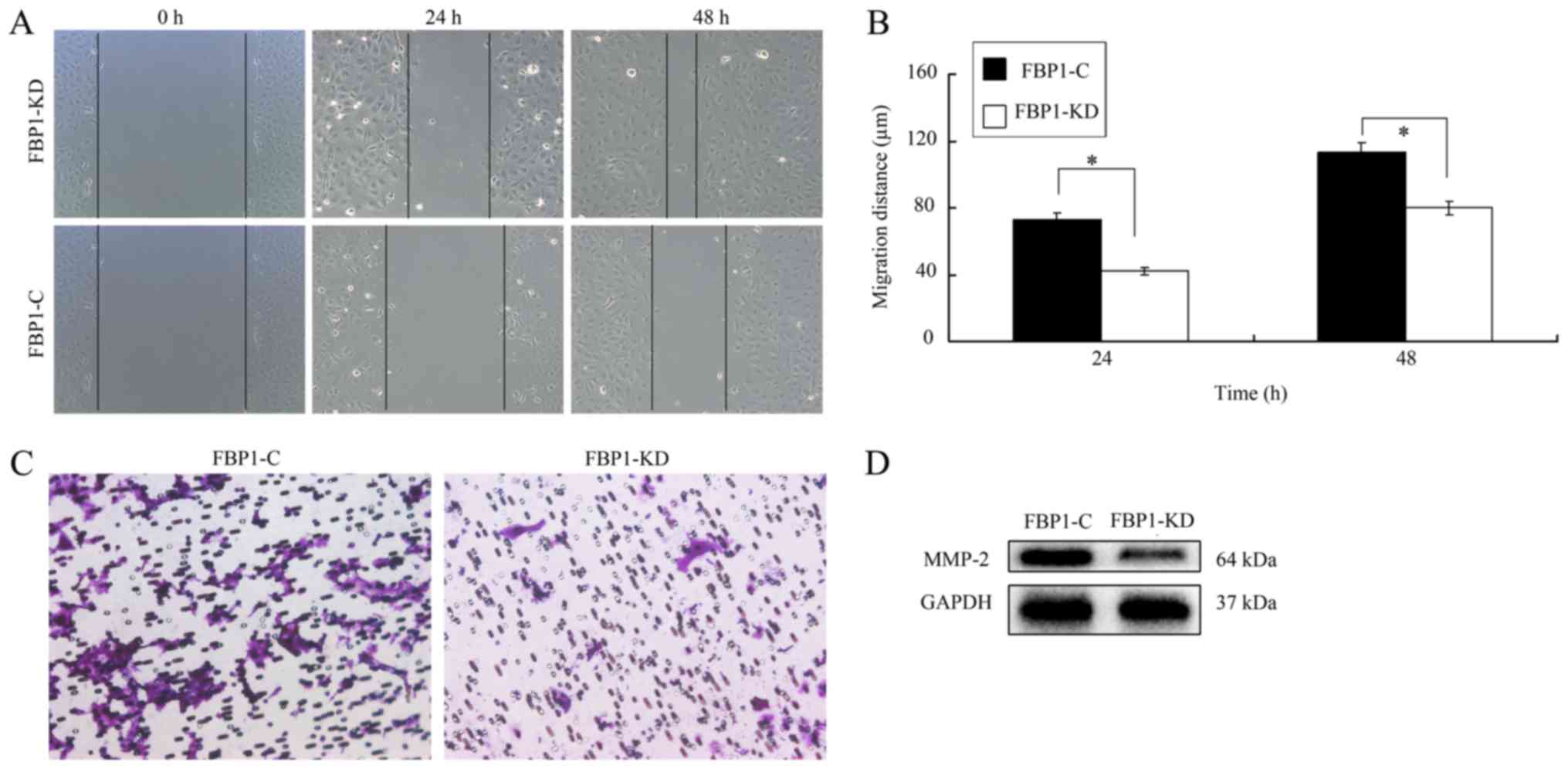
FBP1 was associated with cell
migration and metastasis
Metastasis is a key process that is required for
cancer development (23). An improved
understanding of the genetic evolution of metastasis can provide
insight into the biology of cancer and contribute to the
development of novel therapeutics. Therefore, the influence of FBP1
on ovarian cell migration and invasion was investigated. In the
wound-healing assay, the migration of FBP1 knockdown SKOV3 cells
was slower than that of FBP1 control cells (Fig. 3A). At 24 or 48 h later, the migration
distance of FBP1-C cells was significant higher than that of
FBP1-KD cells (Fig. 3B).
Additionally, the results from the Transwell assay also revealed
that FBP1 knockdown inhibited cell migration to the bottom chambers
(Fig. 3C).
As MMPs are key regulators of cellular matrix
degradation and promote cell migration (24), the expression of MMP-2 in FBP1-C and
FBP1-KD SKOV3 cells was investigated and it was demonstrated that
FBP1 knockdown inhibited the expression of MMP-2 (Fig. 3D). These data implied that FBP1 could
promote cell migration and facilitate cell metastasis.
Discussion
In 2015, ovarian cancer was present in 1.2 million
women and resulted in 0.16 million mortalities worldwide (24). The evolution of surgical techniques
and chemotherapy regimens over the past three decades has resulted
in improvements in the survival of patients with ovarian cancer
(3). Despite high initial response
rates for patients with chemotherapy, the majority of patients
ultimately suffer from recurrence due to chemotherapy resistance
(3). Therefore, novel approaches for
the diagnosis and the treatment of ovarian cancer are urgently
required. Improvements in the understanding of cancer biology and
the underlying mechanisms governing the cancer processes have
facilitated the development of targeted therapies, including
small-molecule inhibitors and monoclonal antibodies (3). Certain ovarian cancers may be caused by
mutations, such as the BRCA1 or BRCA2 mutation (25,26) and
dysregulation in the mitogen-activated protein kinase (MAPK)
pathway (27). However, the majority
of ovarian cancers can be attributed to a growing number of somatic
aberrations (28). In the present
study, the progression of ovarian cancer was positively associated
with FBP1 expression. As the epithelial ovarian cancer progressed,
the expression of FBP1 increased significantly. Furthermore, FBP1
was demonstrated to not only enhance cell proliferation but also
promote cell metastasis; despite FBP1 not affecting the occurrence
of apoptosis. Therefore, we hypothesized that FBP1 is a potential
target for novel treatment development and may serve as a
diagnostic marker for ovarian cancer early screen. In our previous
studies, FBP1 was demonstrated as a potential target of platinum
and knockdown of FBP1 increased the sensitivity of ovarian cancer
cells to carboplatin (22).
Decreased cell viability may be induced by cell
cycle transition suppression and cell death activation. In the
colony formation experiment of the present study, the knockdown of
FBP1 was demonstrated to significantly inhibit colony formation and
an increased number of cells were restrained in G1 phase and fewer
cells were restrained in the S and G2 phases due to FBP1 knockdown.
The cell cycle transition inhibiting proteins, such as p21 and p27,
were induced by FBP1 knockdown. Conversely, cell cycle transition
promoting proteins, including cyclin D1/E, c-Myc, were suppressed
by FBP1 knockdown, and no influence of FBP1 knockdown on apoptosis
was identified. When combined with the results of a previous study
in which caspase 3 was not demonstrated to be activated in ovarian
cancer cells (22), the results of
the present study suggest that FBP1 knockdown significantly
suppresses cell cycle transition; however, does not affect cell
death in ovarian cancer cells.
Metastasis, the process by which a localized cancer
becomes a systemic disease, is a complex, multistep process that
requires cancer cells to detach from the primary tumor, travel,
survive and proliferate in distant organs (29,30).
Metastatic tumors can occur in the early and late stages in the
primary tumor (29,30). An improved understanding of the
genetic evolution of metastasis may provide insight into the
biology of cancer and contribute to the development of novel
therapeutics. MMPs have been traditionally considered to regulate a
number of processes such as cell migration and metastasis (31,32). MMPs
are generally expressed at low levels; however, can be upregulated
during tissue remodeling, inflammation, wound healing, and cancer
progression (33,34). It has been reported that the
expression of MMP-2 was significantly higher in advanced ovarian
cancer than in their benign or premalignant counterparts (35). In the present study, the knockdown of
FBP1 was demonstrated to inhibit the migration of EOC cells,
metastasis and the expression of MMP-2; an important member of the
MMP family. Suggesting that FBP1 knockdown decreases the migration
and metastasis through the inhibition of MMP expression.
The diagnosis of ovarian cancer has been challenging
due to a lack of biomarkers in the early stage of disease, and the
majority of patients eventually experience a median
progression-free survival of 18 months (36). The current gold-standard chemotherapy,
a platinum-paclitaxel combination regimen, is still far from
optimum even though it is a significant improvement from previously
administered treatments (3). A
previous study demonstrated that FBP1 is a target protein of
platinum-based treatment (37). In
the present study, the progression of ovarian cancer was
demonstrated to be positively associated with FBP1 expression. FBP1
enhanced cell cycle transition and metastasis, leading to the
conclusion that FBP1 is a potential diagnostic marker for ovarian
cancer in the early stage of disease.
Acknowledgements
The authors would like to thank the staff of the
Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics
and Gene Regulation, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen
University (Guangzhou, China) for support with regard to the flow
cytometry experiments.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81272222), the Science and
Technology Program Foundation of Guangzhou (grant no.
2014J4100075), the Medical Science and Technology Research
Foundation of Guangdong (grant no. B2016018) and the Medical and
Health Science and Technology Project of Guangzhou (grant no.
20161A010019).
Availability of data and materials
All data generated or analyzed during this study are
included in this article.
Authors' contributions
XX, ZL performed initial experimental design. XX,
JZ, WC and SQ performed the experiments. XH, LD and WL prepared the
tissue samples. XX and ZL wrote the paper. ZZ and XL made
contirbutions to the experimental design and the manuscript.
Ethics approval and consent to
participate
This study was conducted after ethics approval was
obtained from the Medical Ethics Committee of Guangzhou Red Cross
hospital of Medical College, Jinan University (Guangzhou, China).
All contributors consented to participate.
Consent for publication
All authors read and approved the final
manuscript.
Competing interests
The authors declared that they have no competing
interests.
References
|
1
|
A JY, Wang GJ, Sun JG, Gu YC, Wu MS and
Liu JH: Identification of phase I and phase II metabolites of
Guanfu base A hydrochloride in human urine. Eur J Drug Metab
Pharmacokinet. 28:265–272. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Berkenblit A and Cannistra SA: Advances in
the management of epithelial ovarian cancer. J Reprod Med.
50:426–438. 2005.PubMed/NCBI
|
|
3
|
Agarwal R and Kaye SB: Ovarian cancer:
Strategies for overcoming resistance to chemotherapy. Nat Rev
Cancer. 3:502–516. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ozols RF, Bundy BN, Greer BE, Fowler JM,
Clarke-Pearson D, Burger RA, Mannel RS, DeGeest K, Hartenbach EM
and Baergen R; Gynecologic Oncology Group: Phase III trial of
carboplatin and paclitaxel compared with cisplatin and paclitaxel
in patients with optimally resected stage III ovarian cancer: A
Gynecologic Oncology Group study. J Clin Oncol. 21:3194–3200. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
du Bois A, Neijt JP and Thigpen JT: First
line chemotherapy with carboplatin plus paclitaxel in advanced
ovarian cancer-a new standard of care? Ann Oncol. 10(Suppl 1):
S35–S41. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Biagi JJ and Eisenhauer EA: Systemic
treatment policies in ovarian cancer: The next 10 years. Int J
Gynecol Cancer. 13(Suppl 2): S231–S240. 2003. View Article : Google Scholar
|
|
7
|
Neijt JP, Engelholm SA, Tuxen MK, Sorensen
PG, Hansen M, Sessa C, de Swart CA, Hirsch FR, Lund B and van
Houwelingen HC: Exploratory phase III study of paclitaxel and
cisplatin versus paclitaxel and carboplatin in advanced ovarian
cancer. J Clin Oncol. 18:3084–3092. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krivak TC, Darcy KM, Tian C, Armstrong D,
Baysal BE, Gallion H, Ambrosone CB and DeLoia JA; Gynecologic
Oncology Group Phase III Trial: Relationship between ERCC1
polymorphisms, disease progression, and survival in the Gynecologic
Oncology Group Phase III Trial of intraperitoneal versus
intravenous cisplatin and paclitaxel for stage III epithelial
ovarian cancer. J Clin Oncol. 26:3598–3606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Krivak TC, Darcy KM, Tian C, Bookman M,
Gallion H, Ambrosone CB and Deloia JA: Single nucleotide
polypmorphisms in ERCC1 are associated with disease progression,
and survival in patients with advanced stage ovarian and primary
peritoneal carcinoma; a Gynecologic Oncology Group study. Gynecol
Oncol. 122:121–126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dann RB, DeLoia JA, Timms KM, Zorn KK,
Potter J, Flake DD II, Lanchbury JS and Krivak TC: BRCA1/2
mutations and expression: response to platinum chemotherapy in
patients with advanced stage epithelial ovarian cancer. Gynecol
Oncol. 125:677–682. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reynolds C, Obasaju C, Schell MJ, Li X,
Zheng Z, Boulware D, Caton JR, Demarco LC, O'Rourke MA, Shaw Wright
G, et al: Randomized phase III trial of gemcitabine-based
chemotherapy with in situ RRM1 and ERCC1 protein levels for
response prediction in non-small-cell lung cancer. J Clin Oncol.
27:5808–5815. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gong W, Zhang X, Wu J, Chen L, Li L, Sun
J, Lv Y, Wei X, Du Y, Jin H and Dong J: RRM1 expression and
clinical outcome of gemcitabine-containing chemotherapy for
advanced non-small-cell lung cancer: A meta-analysis. Lung Cancer.
75:374–380. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Erriquez J, Becco P, Olivero M, Ponzone R,
Maggiorotto F, Ferrero A, Scalzo MS, Canuto EM, Sapino A, Verdun di
Cantogno L, et al: TOP2A gene copy gain predicts response of
epithelial ovarian cancers to pegylated liposomal doxorubicin:
TOP2A as marker of response to PLD in ovarian cancer. Gynecol
Oncol. 138:627–633. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Malz M, Weber A, Singer S, Riehmer V,
Bissinger M, Riener MO, Longerich T, Soll C, Vogel A, Angel P, et
al: Overexpression of far upstream element binding proteins: A
mechanism regulating proliferation and migration in liver cancer
cells. Hepatology. 50:1130–1139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rabenhorst U, Beinoraviciute-Kellner R,
Brezniceanu ML, Joos S, Devens F, Lichter P, Rieker RJ, Trojan J,
Chung HJ, Levens DL and Zörnig M: Overexpression of the far
upstream element binding protein 1 in hepatocellular carcinoma is
required for tumor growth. Hepatology. 50:1121–1129. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Singer S, Malz M, Herpel E, Warth A,
Bissinger M, Keith M, Muley T, Meister M, Hoffmann H, Penzel R, et
al: Coordinated expression of stathmin family members by far
upstream sequence element-binding protein-1 increases motility in
non-small cell lung cancer. Cancer Res. 69:2234–2243. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen M, Zhang J, Li N, Qian Z, Zhu M, Li
Q, Zheng J, Wang X and Shi G: Promoter hypermethylation mediated
downregulation of FBP1 in human hepatocellular carcinoma and colon
cancer. PLoS One. 6:e255642011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Avigan MI, Strober B and Levens D: A far
upstream element stimulates c-myc expression in undifferentiated
leukemia cells. J Biol Chem. 265:18528–18545. 1990.
|
|
19
|
He L, Weber A and Levens D: Nuclear
targeting determinants of the far upstream element binding protein,
a c-myc transcription factor. Nucleic Acids Res. 28:4558–4565.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dixit U, Liu Z, Pandey AK, Kothari R and
Pandey VN: Fuse binding protein antagonizes the transcription
activity of tumor suppressor protein p53. BMC Cancer. 14:9252014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dixit U, Pandey AK, Liu Z, Kumar S,
Neiditch MB, Klein KM and Pandey VN: FUSE binding protein 1
facilitates persistent hepatitis c virus replication in hepatoma
cells by regulating tumor suppressor p53. J Viro. 89:7905–7921.
2015. View Article : Google Scholar
|
|
22
|
Zhang J, Xiong X, Hua X, Cao W, Qin S, Dai
L, Liang P, Zhang H and Liu Z: Knockdown of FUSE binding protein 1
enhances the sensitivity of epithelial ovarian cancer cells to
carboplatin. Oncol Lett. 14:5819–5824. 2017.PubMed/NCBI
|
|
23
|
Steeg PS: Theodorescu: Metastasis: A
therapeutic target for cancer. Nat Clin Pract Oncol. 5:206–219.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodriguez D, Morrison CJ and Overall CM:
Matrix metalloproteinases: What do they not do? New substrates and
biological roles identified by murine models and proteomics.
Biochim Biophys Acta. 1803:39–54. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pal T, Permuth-Wey J, Betts JA, Krischer
JP, Fiorica J, Arango H, LaPolla J, Hoffman M, Martino MA, Wakeley
K, et al: BRCA1 and BRCA2 mutations account for a large proportion
of ovarian carcinoma cases. Cancer. 104:2807–2816. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Risch HA, McLaughlin JR, Cole DE, Rosen B,
Bradley L, Fan I, Tang J, Li S, Zhang S, Shaw PA and Narod SA:
Population BRCA1 and BRCA2 mutation frequencies and cancer
penetrances: A kin-cohort study in Ontario, Canada. J Natl Cancer
Inst. 98:1694–1706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cancer Genome Atlas Research Network:
Integrated genomic analyses of ovarian carcinoma. Nature.
474:609–615. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bast RC Jr, Hennessy B and Mills GB: The
biology of ovarian cancer: New opportunities for translation. Nat
Rev Cancer. 9:415–428. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Massagué J and Obenauf AC: Metastatic
colonization by circulating tumour cells. Nature. 529:298–306.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Talmadge JE and Fidler IJ: AACR centennial
series: The biology of cancer metastasis: historical perspective.
Cancer Res. 70:5649–5669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zitka O, Kukacka J, Krizkova S, Huska D,
Adam V, Masarik M, Prusa R and Kizek R: Matrix metalloproteinases.
Curr Med Chem. 17:3751–3768. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Butler GS and Overall CM: Updated
biological roles for matrix metalloproteinases and new
‘intracellular’ substrates revealed by degradomics. Biochemistry.
48:10830–10845. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: regulators of the tumor microenvironment. Cell.
41:52–67. 2010. View Article : Google Scholar
|
|
34
|
Hadler-Olsen E, Fadnes B, Sylte I,
Uhlin-Hansen L and Winberg JO: Regulation of matrix
metalloproteinase activity in health and disease. FEBS J.
278:28–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sakata K, Shigemasa K, Nagai N and Ohama
K: Expression of matrix metalloproteinases (MMP-2, MMP-9, MT1-MMP)
and their inhibitors (TIMP-1, TIMP-2) in common epithelial tumors
of the ovary. Int J Oncol. 17:673–681. 2000.PubMed/NCBI
|
|
36
|
Greenlee RT, Hill-Harmon MB, Murray T and
Thun M: Cancer statistics, 2001. CA Cancer J Clin. 51:15–36. 2011.
View Article : Google Scholar
|
|
37
|
Pénzváltó Z, Lánczky A, Lénárt J,
Meggyesházi N, Krenács T, Szoboszlai N, Denkert C, Pete I and
Győrffy B: MEK1 is associated with carboplatin resistance and is a
prognostic biomarker in epithelial ovarian cancer. BMC Cancer.
14:8372014. View Article : Google Scholar : PubMed/NCBI
|