Introduction
Colorectal cancer (CRC), which includes colon cancer
and rectal cancer, is the third most prevalent cancer and the
fourth most common cause of cancer-associated mortality in males
and females worldwide in 2016, with >1 million cases diagnosed
and 600,000 deaths recorded each year (1,2). Multiple
risk factors for CRC have been identified, including age, being of
the male sex, consumption of alcohol and red meat, obesity, insulin
resistance and metabolic syndrome (3). At present, the standard treatment for
CRC is surgical resection followed by chemotherapy and radiotherapy
(4). Despite the advances that have
been achieved in the diagnosis and therapy of CRC, however, the
prognosis of patients with CRC remains poor (5). Overall, ~30-50% of patients with CRC
develop local tumor recurrence or distant metastasis, even
following curative resection of the primary lesion (6,7).
Therefore, it is crucial to fully understand the underlying
mechanisms involved in CRC development and progression, and
identify novel, efficient and specific therapeutic targets for the
treatment of patients with CRC.
MicroRNAs (miRNAs/miRs) are a group of
evolutionarily conserved, non-coding, short RNA molecules found in
plants, animals and certain viruses (8). miRNAs serve an important function in the
regulation of gene expression by interacting with complementary
sites in the 3′-untranslated regions (3′-UTRs) of target mRNAs,
inducing translational repression or mRNA cleavage (9). Multiple studies have reported that
miRNAs regulate a number of physiological and pathological
processes, which include cell proliferation, the cell cycle,
differentiation, apoptosis, metastasis, angiogenesis and metabolism
(10–12). miRNAs are involved in tumorigenesis
and cancer development, with >50% of miRNAs located at
cancer-associated genomic regions (13,14).
Aberrant expression of multiple miRNAs has been observed in various
types of human cancer, including CRC (15–17).
Furthermore, these abnormally expressed miRNAs may act as tumor
suppressors or oncogenes in different types of human cancer
depending on whether they specifically target oncogenes or tumor
suppressor genes (18,19). Thus, the targeting of miRNAs may
result in novel anti-cancer therapies, as a result of the close
associations between miRNAs and carcinogenesis.
miR-539 has been reported to be aberrantly expressed
in several types of cancer including prostate cancer (20), nasopharyngeal carcinoma (21) and osteosarcoma (22). However, the expression levels,
biological functions and underlying mechanisms of miR-539 in CRC
remain unclear. Therefore, the aim of the present study was to
detect miR-539 expression and to investigate the biological
functions of miR-539 in CRC, as well as the potential mechanisms
underlying these functions.
Materials and methods
Clinical samples
The present study was approved by the Ethics
Committee of Liaoning Cancer Hospital and Institute (Shenyang,
China), and written informed consent was obtained from all patients
enrolled in the study. A total of 49 pairs of CRC tissues and
corresponding normal adjacent tissues were collected from patients
with CRC who underwent surgical resection at the Liaoning Cancer
Hospital and Institute between June 2012 and October 2015. All
these tissues were derived from initial surgery with neither
preoperative chemotherapy or radiotherapy. Tissue samples were
promptly frozen in liquid nitrogen at the time of surgery and
stored at −80°C until further use. Clinicopathological features of
the patients are listed in Table
I.
 | Table I.Associations between miR-539
expression and clinicopathological characteristics of patients with
colorectal cancer. |
Table I.
Associations between miR-539
expression and clinicopathological characteristics of patients with
colorectal cancer.
|
|
| miR-539
expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | Cases | Low | High | P-value |
|---|
| Sex |
|
|
| 0.320 |
|
Male | 30 | 17 | 13 |
|
|
Female | 19 | 8 | 11 |
|
| Age, years |
|
|
| 0.628 |
|
<60 | 18 | 10 | 8 |
|
|
≥60 | 31 | 15 | 16 |
|
| Tumor location |
|
|
| 0.458 |
|
Colon | 21 | 12 | 9 |
|
|
Rectum | 28 | 13 | 15 |
|
| Tumor size, cm |
|
|
| 0.316 |
|
<5 | 24 | 14 | 10 |
|
| ≥5 | 25 | 11 | 14 |
|
|
Differentiation |
|
|
| 0.117 |
| Well
and moderate | 26 | 16 | 10 |
|
|
Poor | 23 | 9 | 14 |
|
| Lymph node
metastasis |
|
|
| 0.015 |
| No | 26 | 9 | 17 |
|
|
Yes | 23 | 16 | 7 |
|
| TNM stage |
|
|
| 0.007 |
|
I–II | 23 | 7 | 16 |
|
|
III–IV | 26 | 18 | 8 |
|
Cell lines and transfection
A total of five human CRC cell lines (LoVo, HCT116,
HT29, SW480 and SW620) and the normal human colon epithelium cell
line FHC were purchased from the American Type Culture Collection
(Manassas, VA, USA). Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml
streptomycin. All cell lines were maintained at 37°C in a
humidified atmosphere of 5% CO2.
miR-539 mimics and corresponding negative control
miRNA (miR-NC) were acquired from Shanghai GenePharma Co., Ltd.
(Shanghai, China). The miR-539 mimics sequence was
5′-GGAGAAAUUAUCCUUGGUGUUGU-3′ and the miR-NC sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. pcDNA3.1-SRY-box 4 (SOX4) plasmids and
empty vector pcDNA3.1 plasmids were synthesized by Guangzhou
RiboBio Co., Ltd (Guangzhou, China). For transfection, cells were
seeded into 6-well plates in DMEM medium containing 10% FBS at
60–70% confluence. Following incubation overnight at 37°C with 5%
CO2, cells were transfected with miRNA mimics (100 pmol)
or plasmids (4 g) using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.), following the manufacturer's protocol.
The culture medium was replaced with fresh DMEM supplemented with
10% FBS 8 h after transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the cell lines and
tissue samples using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), in accordance with the manufacturer's protocol.
The concentration of total RNA was examined using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). For analysis of
miR-539 expression, cDNA was synthesized using the TaqMan MicroRNA
Reverse Transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. qPCR
was performed using a TaqMan MicroRNA PCR kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.) on an ABI Prism 7500 Sequence
Detection System (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The cycling conditions for qPCR were as follows: 50°C for 2
min, 95°C for 10 min; 40 cycles of denaturation at 95°C for 15 sec;
and annealing/extension at 60°C for 60 sec. To quantify SOX4 mRNA,
reverse transcription was conducted using a PrimeScript RT Reagent
kit (Takara Biotechnology Co., Ltd., Dalian, China) according to
the manufacturer's protocol, and qPCR was performed using SYBR
Premix Ex Taq™ (Takara Biotechnology Co., Ltd., Dalian, China). The
cycling conditions for qPCR were as follows: 5 min at 95°C,
followed by 40 cycles of 95°C for 30 sec and 65°C for 45 sec.
U6 and GAPDH served as internal controls for the
detection of miR-539 and SOX4 mRNA, respectively. The primers were
designed as follows: miR-539, 5′-GAAGAGGCTAACGTGAGGTTG-3′ (forward)
and 5′-CACCATGACCAAGCCACGTAG-3′ (reverse); U6,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and
5′-CGCTTCACGAATTTGCGTGTCAT-3′ (reverse); SOX4,
5′-CTTGACATGATTAGCTGGCATGATT-3′ (forward) and
5′-CCTGTGCAATATGCCGTGTAGA-3′ (reverse); and GAPDH,
5′-CCAAAATCAGATGGGGCAATGCTGG-3′ (forward) and
5′-TGATGGCATGGACTGTGGTCATTCA-3′ (reverse). The 2−ΔΔCq
method was used for relative quantification (23).
MTT assay
Cell proliferation was determined using MTT assays.
Transfected cells were harvested, resuspended in DMEM medium
containing 10% FBS and seeded in 96-well plates at a density of
3×103 cells/well, and cultured for 0, 24, 48, and 72 h.
At these specified times, MTT assays were performed. Briefly, 20 µl
MTT solution (5 mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was added into each well and incubated at 37°C with 5%
CO2 for additional 4 h. The culture medium was then
removed and dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was used
to dissolve the formazan crystals at 37°C for 10 min. Finally, the
absorbance at 490 nm was detected using a microplate reader
(iMark™; Bio-Rad Laboratories, Inc., Hercules, CA, USA). All
experiments were performed in triplicate and repeated at least
three times.
Cell invasion assay
Cell invasion assays were performed using Transwell
chambers (8 mm pores; Costar, Corning Incorporated, NY USA) coated
with Matrigel solution (40 µl per chamber; Matrigel: FBS-free
medium ratio 1:10; BD Biosciences, San Jose, CA, USA). Following
transfection for 48 h, 5×104 cells cultured with
FBS-free DMEM medium were placed into the upper chamber, and the
lower chamber was filled with 500 µl DMEM with 20% FBS. After 48 h
of incubation at 37°C, cells on the upper surface of the membrane
were carefully removed with cotton swab. Cells that invaded through
the membrane surface were fixed in 100% methanol at room
temperature for 15 min, stained with 0.1% crystal violet at room
temperature for 15 min, and photographed under a light microscope
(Olympus Corporation, Tokyo, Japan) at ×200 magnification in five
randomized fields.
Bioinformatic analysis and luciferase
reporter assay
Target genes of miR-539 were searched using
TargetScan (24) (http://www.targetscan.org) and miRBase (14,25–27)
(http://www.mirbase.org/).
For the luciferase reporter assay,
pMIR-SOX4-3′-UTR-wild type (WT) and pMIR-SOX4-3′-UTR-mutant (MUT)
were synthesized by Shanghai GenePharma Co., Ltd. Cells were seeded
into 24 well plates at 60–70% confluence, and transfected with
miR-539 mimics or miR-NC with pMIR-SOX4-3′-UTR-WT or
pMIR-SOX4-3′-UTR-MUT using Lipofectamine 2000, according to the
manufacturer's protocol. The activity of Renilla luciferase
and firefly luciferase was determined 48 h after transfection using
a Dual-Luciferase Reporter Assay system (Promega Corporation,
Madison, WI, USA), according to the manufacturer's protocol. The
relative luciferase activity was normalized to Renilla
luciferase activity.
Western blot
Protein was extracted from tissues and cells using a
Total Protein Extraction kit (Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China) supplemented with a protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA), according to the manufacturer's
protocol. Protein concentration was detected using a BCA kit
(Beyotime Institute of Biotechnology, Haimen, China). Equal amounts
of protein (30 µg) were resolved on 10% sodium dodecyl
sulfate-polyacrylamide gels and transferred to polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA).
Subsequently, the membranes were blocked at room temperature with
5% non-fat milk in TBS/0.1% Tween (TBST) for 1 h, and incubated
with the following primary antibodies overnight at 4°C: Mouse
anti-human monoclonal SOX4 (1:1,000; cat. No. sc-130633; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and mouse anti-human
monoclonal GAPDH (1:1,000; cat. no. sc-51907; Santa Cruz
Biotechnology, Inc.). Following three washes with TBST, the
membranes were incubated with a goat-anti-mouse horseradish
peroxidase-conjugated secondary antibody (1:3,000; cat. no.
sc-2005; Santa Cruz Biotechnology, Inc.) at room temperature for 1
h. Protein bands were developed using an enhanced chemiluminescent
reagent (GE Healthcare Life Sciences, Little Chalfont, UK) and
analyzed with Image Lab software (version 3.0; Bio-Rad
Laboratories, Inc.). GAPDH was used as a loading control.
Statistical analysis
Each assay was repeated three times, and data were
expressed as the mean ± standard deviation. All statistical
analyses were performed using Spearman's correlation test, paired
Student's t-test or one-way analysis of variance (ANOVA) with SPSS
18.0 software (SPSS, Inc., Chicago, IL, USA). Student-Newman-Keuls
test was used as a post hoc test following ANOVA. The association
between miR-539 expression and the clinicopathological factors in
CRC was analyzed by the χ2 test. P<0.05 was
considered to indicate a statistically significant difference.
Results
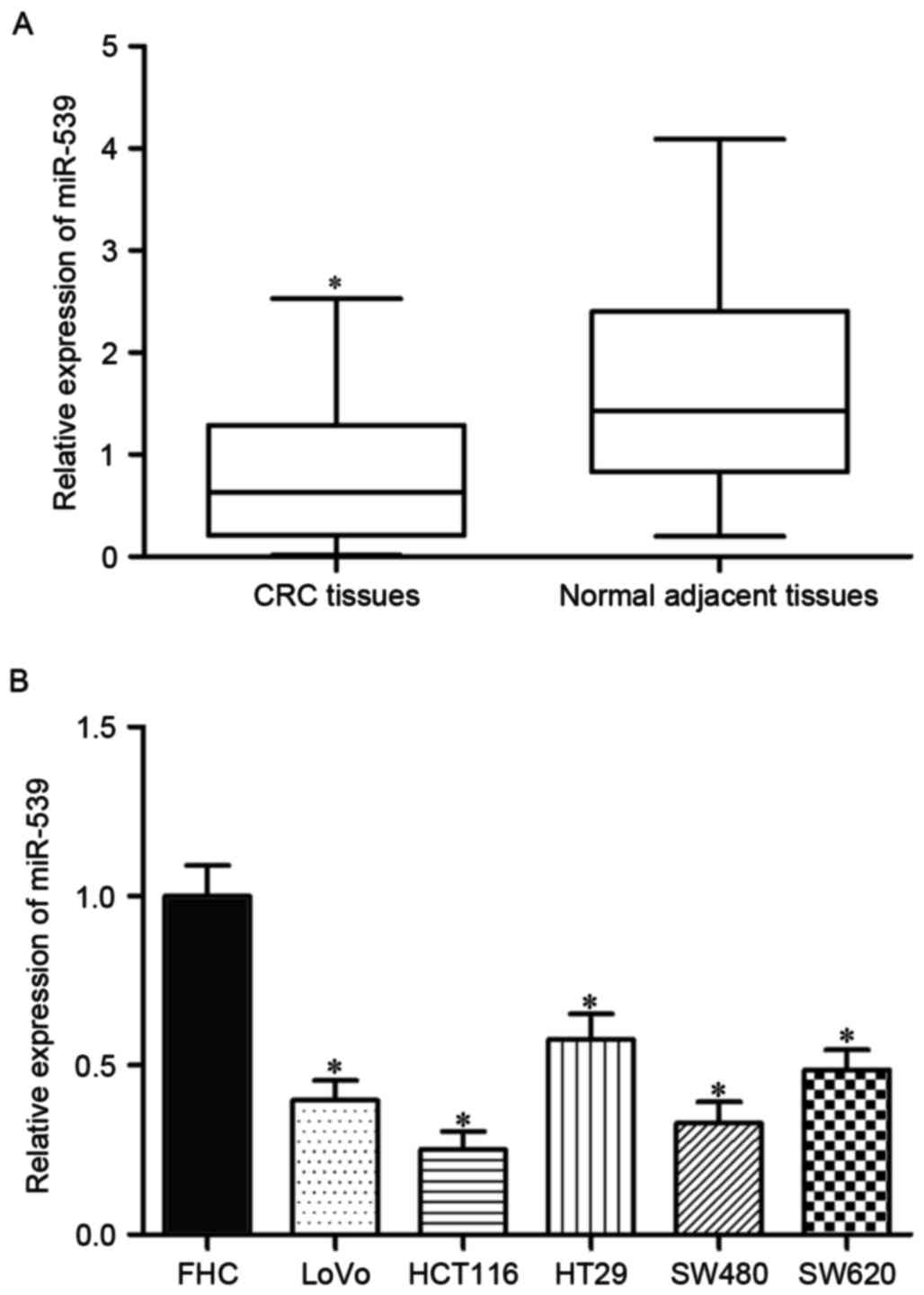
miR-539 expression is downregulated in
CRC tissues and cell lines
To investigate the expression patterns of miR-539 in
CRC, miR-539 expression was examined in 49 pairs of CRC tissues and
corresponding normal adjacent tissues by RT-qPCR. The results
demonstrated that, compared with normal adjacent tissues, miR-539
was significantly downregulated in CRC tissues (Fig. 1A; P<0.05). To analyze the
associations between miR-539 expression and the clinicopathological
characteristics of patients with CRC, the median miR-539 expression
level (0.629) was regarded as the cut-off to divide all patients
into either the miR-539 low-expression group (n=25) or the miR-539
high-expression group (n=24). As presented in Table I, a low expression level of miR-539
was associated with lymph node metastasis (P=0.015) and
tumor-node-metastasis (TNM) stage (28) (P=0.007). However, no significant
associations were observed between miR-539 levels or any other
clinicopathological factor, which included sex (P=0.320), age
(P=0.628), tumor location (P=0.458), tumor size (P=0.316) or
differentiation (P=0.117).
Subsequently, miR-539 expression was measured in
five CRC cell lines and a normal human colon epithelium cell line
by RT-qPCR. As presented in Fig. 1B,
the expression levels of miR-539 were decreased in all five CRC
cell lines relative to FHC cells (P<0.05). CT116 and SW480 cell
lines, which expressed relatively lower miR-539 expression level,
were selected for further experiments. These results suggested that
miR-539 may be involved in CRC initiation and progression.
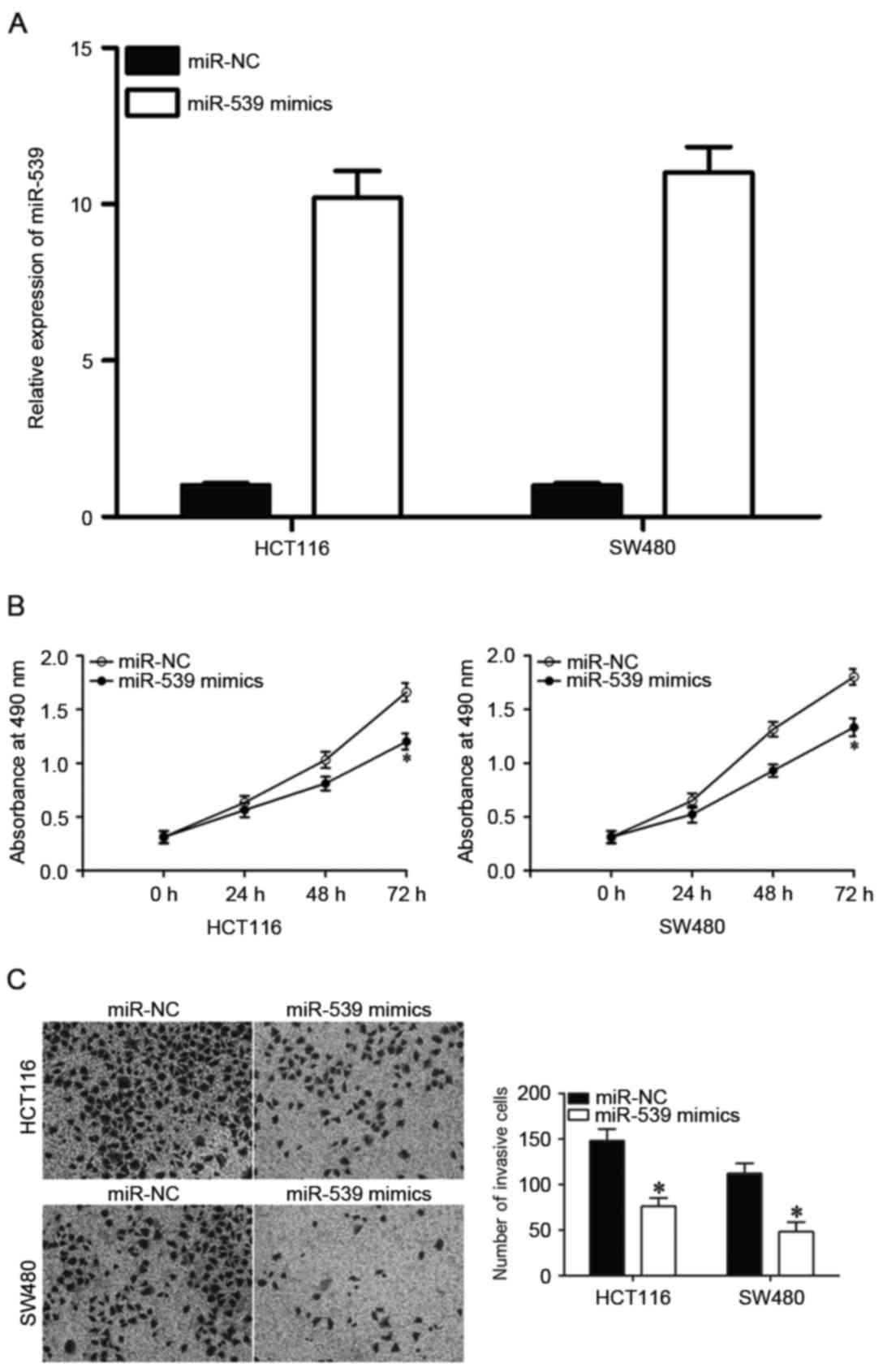
miR-539 inhibits CRC cell
proliferation and invasion in vitro
To observe the potential effects of miR-539 on CRC
cells, miR-539 mimics or miR-NC were transfected into HCT116 and
SW480 cells. Following transfection, RT-qPCR confirmed that miR-539
expression was increased in HCT116 and SW480 cells transfected with
miR-539 mimics compared with those transfected with miR-NC
(Fig. 2A, P<0.05). MTT assays were
utilized to assess whether miR-539 contributed to the suppression
of CRC cell proliferation. Transfection of miR-539 mimics was
revealed to significantly inhibit the proliferation of HCT116 and
SW480 cells at 72 h compared with those transfected with miR-NC
(Fig. 2B; P<0.05). Cell invasion
assays were performed to determine the effect of miR-539 on the
invasiveness of CRC cells. Invasiveness was significantly
suppressed in HCT116 and SW480 cells transfected with miR-539
mimics compared with miR-NC (Fig.
2C). Collectively, these results indicated a potential
tumor-suppressive function of miR-539 in CRC proliferation and
invasion.
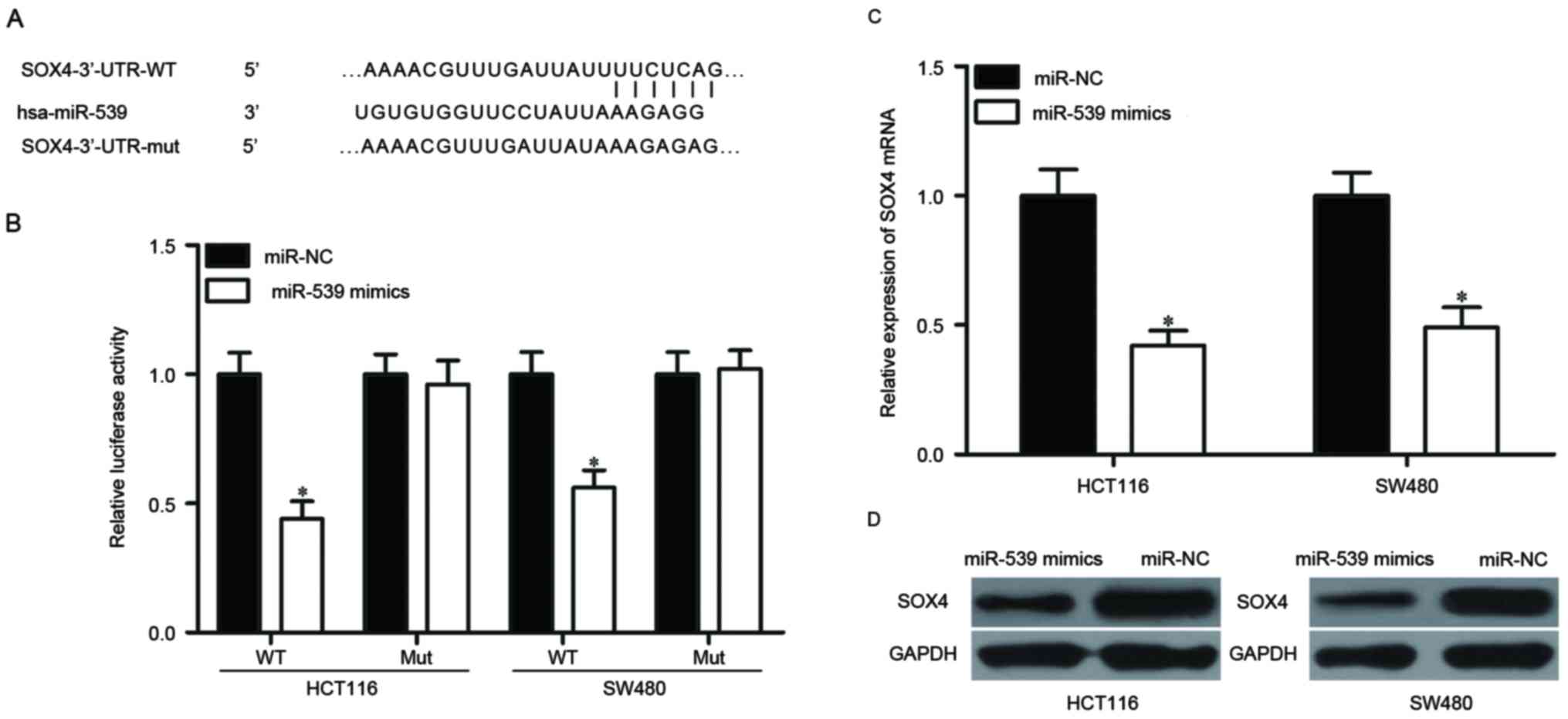
Validation of SOX4 as a direct target
of miR-539 in CRC
To reveal the mechanism by which miR-539 affects the
biological functions of CRC cells, the potential target genes of
miR-539 were explored using bioinformatics. Among these putative
targets, SOX4 was selected for further examination (Fig. 3A), as it is upregulated in CRC and is
involved into CRC progression (29,30). A
luciferase reporter assay was performed to determine whether
miR-539 directly binds to the 3′-UTR of SOX4. HCT116 and SW480
cells were transfected with pMIR-SOX4-3′-UTR-WT or
pMIR-SOX4-3′-UTR-MUT, along with miR-539 mimics or miR-NC. As
presented in Fig. 3B, the luciferase
activity of the reporter containing WT SOX4 3′-UTR was
significantly decreased following co-transfection with miR-539
mimics compared with miR-NC (P<0.05). However, there was no
change in luciferase activity of the reporter containing the mutant
binding site. Furthermore, RT-qPCR and western blotting were
performed to measure the expression of SOX4 in HCT116 and SW480
cells. As predicted, ectopic expression of miR-539 decreased the
mRNA and protein levels of SOX4 in HCT116 and SW480 cells compared
with miR-NC (Fig. 3C and D;
P<0.05). Thus, SOX4 was demonstrated to be a direct target of
miR-539 in CRC.
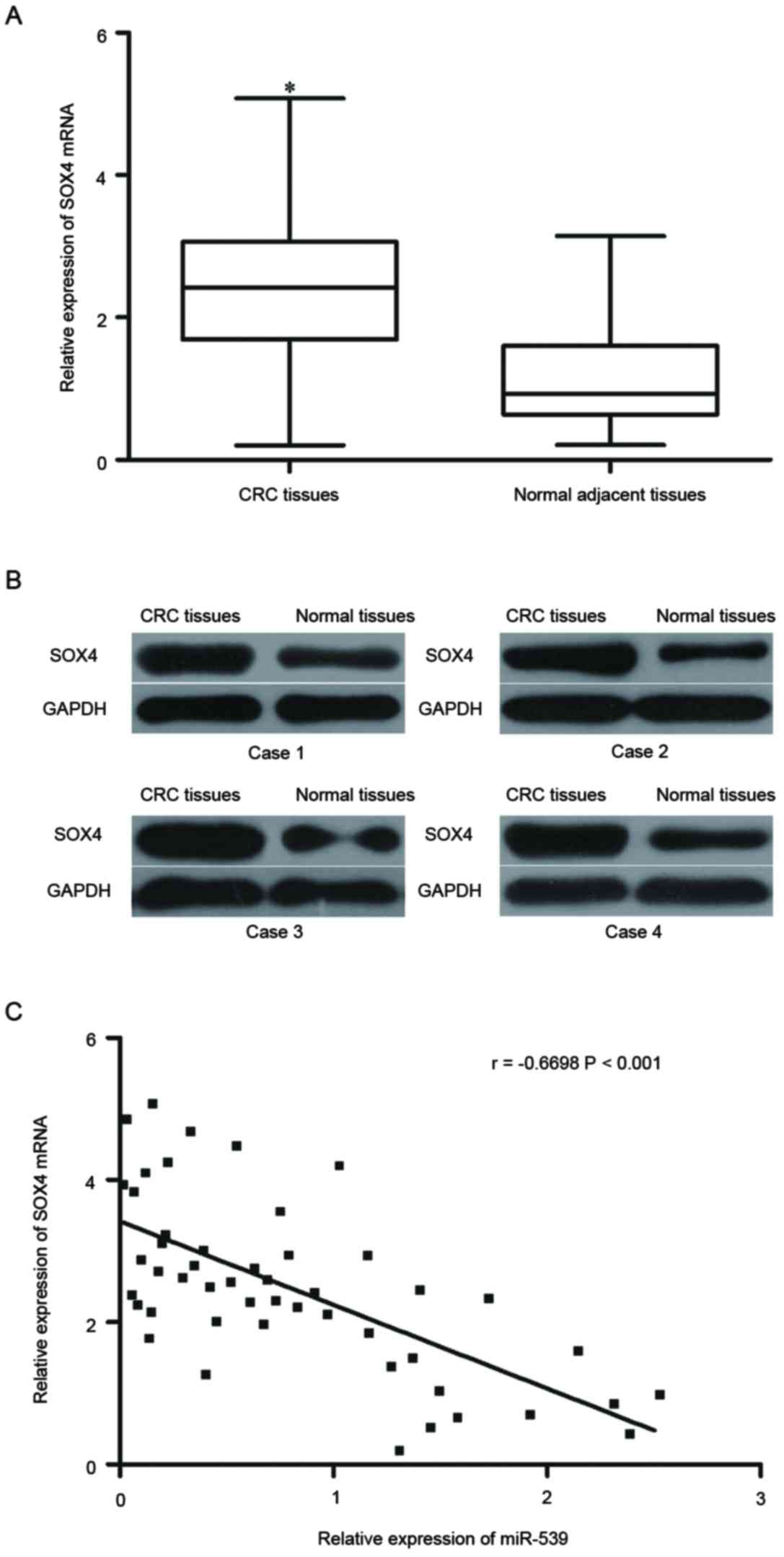
miR-539 is inversely correlated with
SOX4 expression in CRC tissues
To further investigate the relationship between
miR-539 and SOX4, SOX4 mRNA and protein expression in were measured
in CRC tissues and corresponding normal adjacent tissues. The
results revealed that SOX4 mRNA and protein were upregulated in CRC
tissues compared with the corresponding normal adjacent tissues
(Fig. 4A and B; P<0.05).
Spearman's correlation analysis further revealed that miR-539
expression levels were negatively correlated with SOX4 mRNA
expression levels in CRC tissues (Fig.
4C; r=−0.6698, P<0.001). These results suggested that the
downregulation of miR-539 may result in the upregulation of SOX4 in
CRC.
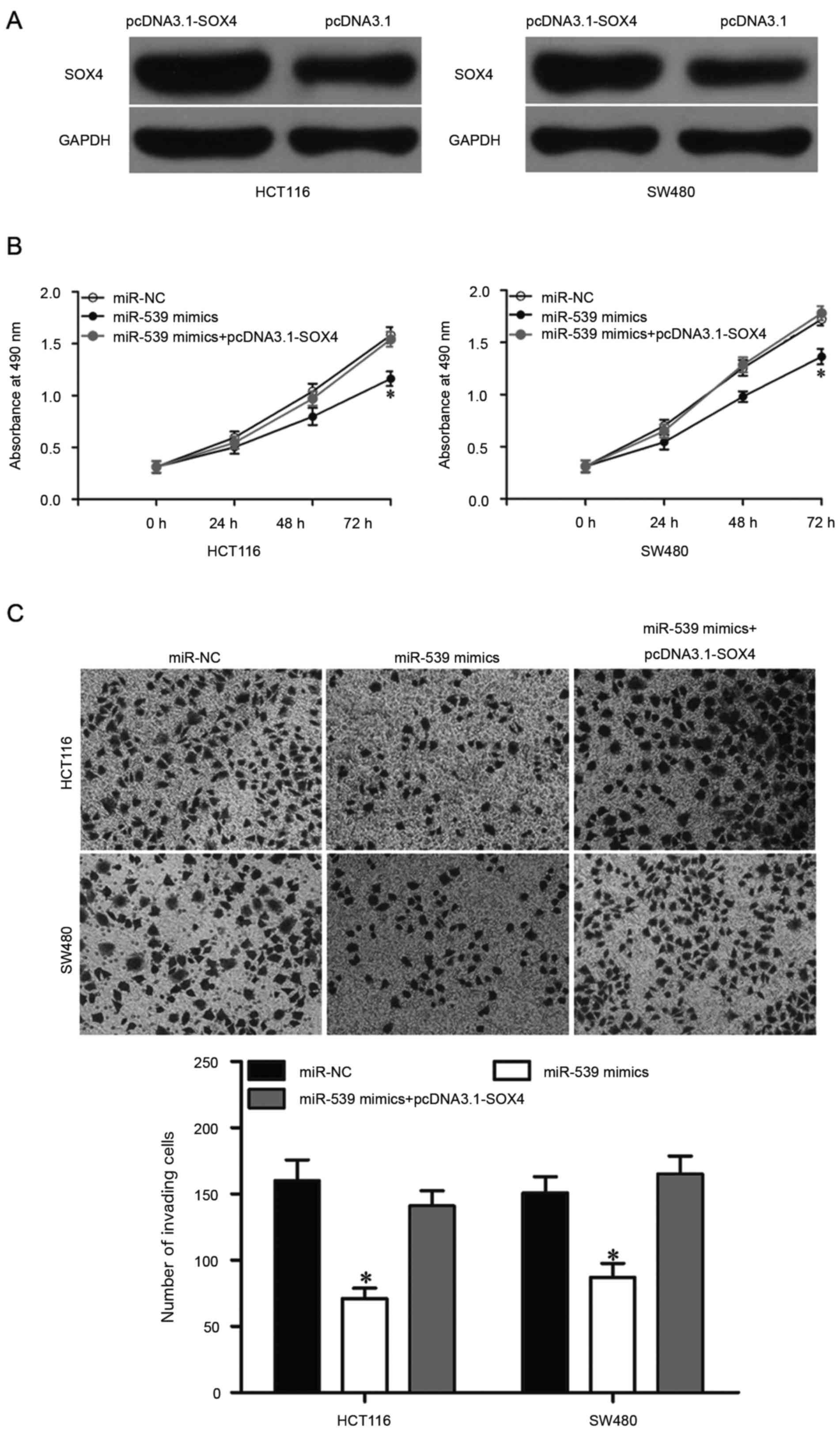
Upregulation of SOX4 reverses the
tumor-suppressive effects of miR-539 in CRC cells
To examine whether the downregulation of SOX4
mediates the tumor suppressive functions of miR-539 in CRC cells,
rescue experiments were performed. HCT116 and SW480 cells were
transfected with pcDNA3.1-SOX4 to increase its expression (Fig. 5A; P<0.05). In addition, further
exploration with MTT and Transwell invasion assays indicated that
upregulation of SOX4 could rescue the suppressive effects on CRC
cell proliferation (Fig. 5B;
P<0.05) and invasion (Fig. 5C;
P<0.05) induced by miR-539 overexpression. Collectively, these
results suggested that miR-539 inhibits CRC cell invasion and
proliferation by negatively modulating SOX4 expression.
Discussion
Tumor progression in CRC is a multi-step process
that involves alterations at the genetic and epigenetic level
(31–33). Accumulated evidence has led to the
view that miRNAs serve important functions in cancer formation and
through the regulation of tumor cell proliferation, cycle,
apoptosis, metastasis and angiogenesis (34–36).
Therefore, miRNAs may be potential therapeutic targets for
miRNA-based diagnosis, prognosis and therapy. The present study
used RT-qPCR to demonstrate that miR-539 expression was
significantly decreased in CRC tissues and cell lines. In addition,
low expression of miR-539 was significantly associated with tumor
progression in patients with CRC. Functional experiments were used
to demonstrate that miR-539 upregulation suppressed CRC cell
proliferation and invasion. Furthermore, SOX4 was identified as a
direct target gene of miR-539 in CRC. In summary, these results
suggested that miR-539 may be used to develop novel strategies
against CRC growth and metastasis in the future.
Abnormal expression of miR-539 has been reported to
in multiple types of human cancer. For example, a previous study
demonstrated that miR-539 was downregulated in an osteosarcoma cell
line compared with an osteoblast cell line (37). Mirghasemi et al (38) reported that miR-539 expression levels
are relatively low in osteosarcoma tissues, and low expression of
miR-539 was associated with TNM stage, metastasis and recurrence.
In addition, Kaplan-Meier survival analysis and log-rank tests
demonstrated that reduced miR-539 expression was associated with
low overall survival in patients with osteosarcoma, and miR-539 was
revealed to be an independent prognostic marker of overall survival
in patients with osteosarcoma (38).
Gu et al (39) reported that
miR-539 expression is decreased in thyroid cancer tissues and cell
lines. In addition, consistent with these results, low expression
levels of miR-539 was observed in nasopharyngeal carcinoma
(21) and prostate cancer (20). The high frequency of miR-539
downregulation in these types of human cancer suggested that
miR-539 may be a novel marker for the diagnosis and prognosis of
specific cancers.
Abnormal expression of miR-539 serves an important
function in the initiation and progression of several types of
tumor. For instance, upregulation of miR-539 decreases cell
migration and invasion in thyroid cancer (39). Another previous study demonstrated
that miR-539 serves tumor suppressive functions in the growth and
metastasis of osteosarcoma (22). Lv
et al (21) reported that
enforced expression of miR-539 suppresses the growth of
nasopharyngeal carcinoma cells in vitro and in vivo.
Meanwhile, Zhang et al (20)
demonstrated that restoration of miR-539 expression in prostate
cancer represses cell proliferation, migration and invasion in
vitro, and decreases tumor growth and metastasis in
vivo. These results suggest that miR-539 may serve an important
function in these types of cancer, and it may be a potential target
for the treatment of specific cancers.
Since the potential molecular mechanism underlying
the tumor-suppressive effects of miR-539 in CRC remains to be
elucidated, the present study aimed to explore this mechanism. To
date, few genes have been validated as direct targets of miR-539,
including sperm associated antigen 5 in prostate cancer (20), cyclin dependent kinase 4 in
nasopharyngeal carcinoma (21),
matrix metalloproteinase 8 in osteosarcoma (22) and caspase recruitment domain family
member 11 in thyroid cancer (39). In
the present study, SOX4 was validated as a novel direct and
functional target of miR-539 in CRC. First, bioinformatics
predicated SOX4 as a potential target of miR-539. Secondly,
luciferase reporter assays demonstrated that miR-539 directly
targeted the 3′-UTR of SOX4. Thirdly, overexpression of miR-539
suppressed SOX4 mRNA and protein expression in CRC cells. Fourthly,
SOX4 was upregulated in CRC tissues at the mRNA and protein levels.
A significant negative correlation between miR-539 and SOX4 mRNA
expression levels was also observed in CRC tissues. Finally, rescue
experiments indicated that upregulation of SOX4 partially restored
the tumor suppressive effects of miR-539, by suppressing the
proliferation and invasion of CRC cells. It is essential to
identify miR-539 target genes in order to elucidate the functions
of miR-539 in the initiation and progression of CRC.
SOX4, a member of the SOX family of transcription
factors, serves essential functions during embryonic development
and cell fate specification in virtually all cells, tissues, and
organ systems (40–42). SOX4 is overexpressed in multiple types
of human cancer, including gastric cancer (43), hepatocellular carcinoma (44), prostate cancer (45), osteosarcoma (46) and non-small cell lung cancer (47). Furthermore, multiple studies have
demonstrated that SOX4 serves an oncogenic function in
tumorigenesis and tumor development. For example, inhibition of
SOX4 represses prostate cancer proliferation, migration and
invasion, and promotes the apoptotic process and the
epithelial-mesenchymal transition in vitro (45). In CRC,
SOX4 was upregulated at the mRNA and protein level, and was
significantly associated with recurrence (29). SOX4 knockdown also inhibited the
proliferation and invasion of CRC cells (29,30).
Therefore, SOX4 may be a novel therapeutic target in CRC, due to
its cancer-associated functions.
To conclude, the present study indicated that
miR-539 may function as a novel tumor suppressor in CRC. Our data
provide a novel insight into the underlying mechanisms responsible
for the progression of CRC. Therefore, the miR-539/SOX4 axis should
be further investigated as a promising treatment for patients with
CRC.
Acknowledgements
The present study was supported by the Clinical
Capability Construction Project for Liaoning Provincial Hospitals
(grant no. LNCCC-D42-2015).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abdelsatir AA, Husain NE, Hassan AT,
Elmadhoun WM, Almobarak AO and Ahmed MH: Potential benefit of
metformin as treatment for colon cancer: The evidence so far. Asian
Pac J Cancer Prev. 16:8053–8058. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu H, Gao G, Jiang L, Guo L, Lin M, Jiao
X, Jia W and Huang J: Decreased expression of miR-218 is associated
with poor prognosis in patients with colorectal cancer. Int J Clin
Exp Pathol. 6:2904–2911. 2013.PubMed/NCBI
|
|
5
|
Meyerhardt JA and Mayer RJ: Systemic
therapy for colorectal cancer. N Engl J Med. 352:476–487. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Varol U, Yildiz I, Salman T, Karabulut B
and Uslu R: Markers to predict the efficacy of bevacizumab in the
treatment of metastatic colorectal cancer. Tumori. 100:370–376.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Konda B, Shum H and Rajdev L:
Anti-angiogenic agents in metastatic colorectal cancer. World J
Gastrointest Oncol. 7:71–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen PS, Su JL and Hung MC: Dysregulation
of microRNAs in cancer. J Biomed Sci. 19:902012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou L, Liang X, Zhang L, Yang L, Nagao N,
Wu H, Liu C, Lin S, Cai G and Liu J: miR-27a-3p functions as an
oncogene in gastric cancer by targeting BTG2. Oncotarget.
7:51943–51954. 2016.PubMed/NCBI
|
|
11
|
Tagscherer KE, Fassl A, Sinkovic T,
Richter J, Schecher S, Macher-Goeppinger S and Roth W: MicroRNA-210
induces apoptosis in colorectal cancer via induction of reactive
oxygen. Cancer Cell Int. 16:422016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cui F, Wang S, Lao I, Zhou C, Kong H,
Bayaxi N, Li J, Chen Q, Zhu T and Zhu H: miR-375 inhibits the
invasion and metastasis of colorectal cancer via targeting SP1 and
regulating EMT-associated genes. Oncol Rep. 36:487–493. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Griffiths-Jones S, Grocock RJ, van Dongen
S, Bateman A and Enright AJ: miRBase: microRNA sequences, targets
and gene nomenclature. Nucleic Acids Res. 34:(Database Issue):.
D140–D144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong F, Xu T, Shen Y, Zhong S, Chen S,
Ding Q and Shen Z: Dysregulation of miRNAs in bladder cancer:
Altered expression with aberrant biogenesis procedure. Oncotarget.
8:27547–27568. 2017.PubMed/NCBI
|
|
16
|
Zhou L, Xu Z, Ren X, Chen K and Xin S:
MicroRNA-124 (miR-124) inhibits cell proliferation, metastasis and
invasion in colorectal cancer by downregulating Rho-associated
protein kinase 1 (ROCK1). Cell Physiol Biochem. 38:1785–1795. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang H, Ju H, Zhang L, Lu H and Jie K:
microRNA-577 suppresses tumor growth and enhances chemosensitivity
in colorectal cancer. J Biochem Mol Toxicol. 31:e218882017.
View Article : Google Scholar
|
|
18
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: Oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Porkka KP, Pfeiffer MJ, Waltering KK,
Vessella RL, Tammela TL and Visakorpi T: MicroRNA expression
profiling in prostate cancer. Cancer Res. 67:6130–6135. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Li S, Yang X, Qiao B, Zhang Z and
Xu Y: miR-539 inhibits prostate cancer progression by directly
targeting SPAG5. J Exp Clin Cancer Res. 35:602016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lv LY, Wang YZ, Zhang Q, Zang HR and Wang
XJ: miR-539 induces cell cycle arrest in nasopharyngeal carcinoma
by targeting cyclin-dependent kinase 4. Cell Biochem Funct.
33:534–540. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jin H and Wang W: MicroRNA-539 suppresses
osteosarcoma cell invasion and migration in vitro and targeting
Matrix metallopeptidase-8. Int J Clin Exp Pathol. 8:8075–8082.
2015.PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kozomara A and Griffiths-Jones S: miRBase:
Annotating high confidence microRNAs using deep sequencing data.
Nucleic Acids Res. 42:(Database Issue):. D68–D73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kozomara A and Griffiths-Jones S: miRBase:
Integrating microRNA annotation and deep-sequencing data. Nucleic
Acids Res. 39:(Database Issue):. D152–D157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Griffiths-Jones S, Saini HK, van Dongen S
and Enright AJ: miRBase: Tools for microRNA genomics. Nucleic Acids
Res. 36:(Database Issue):. D154–D158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shia J, Klimstra DS, Bagci P, Basturk O
and Adsay NV: TNM staging of colorectal carcinoma: Issues and
caveats. Semin Diagn Pathol. 29:142–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang B, Li Y, Tan F and Xiao Z: Increased
expression of SOX4 is associated with colorectal cancer
progression. Tumour Biol. 37:9131–9137. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Andersen CL, Christensen LL, Thorsen K,
Schepeler T, Sørensen FB, Verspaget HW, Simon R, Kruhøffer M,
Aaltonen LA, Laurberg S and Ørntoft TF: Dysregulation of the
transcription factors SOX4, CBFB and SMARCC1 correlates with
outcome of colorectal cancer. Br J Cancer. 100:511–523. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guda K, Veigl ML, Varadan V, Nosrati A,
Ravi L, Lutterbaugh J, Beard L, Willson JK, Sedwick WD, Wang ZJ, et
al: Novel recurrently mutated genes in African American colon
cancers. Proc Natl Acad Sci USA. 112:1149–1154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vaiopoulos AG, Athanasoula KCh and
Papavassiliou AG: Epigenetic modifications in colorectal cancer:
Molecular insights and therapeutic challenges. Biochim Biophys
Acta. 1842:971–980. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bardhan K and Liu K: Epigenetics and
colorectal cancer pathogenesis. Cancers (Basel). 5:676–713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar
|
|
35
|
Qu S, Yao Y, Shang C, Xue Y, Ma J, Li Z
and Liu Y: MicroRNA-330 is an oncogenic factor in glioblastoma
cells by regulating SH3GL2 gene. PLoS One. 7:e460102012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li G, Yang F, Xu H, Yue Z, Fang X and Liu
J: MicroRNA-708 is downregulated in hepatocellular carcinoma and
suppresses tumor invasion and migration. Biomed Pharmacother.
73:154–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu H, Zhang Y, Cai XH, Huang JF and Cai L:
Changes in microRNA expression in the MG-63 osteosarcoma cell line
compared with osteoblasts. Oncol Lett. 4:1037–1042. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mirghasemi A, Taheriazam A, Karbasy SH,
Torkaman A, Shakeri M, Yahaghi E and Mokarizadeh A: Down-regulation
of miR-133a and miR-539 are associated with unfavorable prognosis
in patients suffering from osteosarcoma. Cancer Cell Int.
15:862015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gu L and Sun W: miR-539 inhibits thyroid
cancer cell migration and invasion by directly targeting CARMA1.
Biochem Biophys Res Commun. 464:1128–1133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schepers GE, Teasdale RD and Koopman P:
Twenty pairs of sox: Extent, homology, and nomenclature of the
mouse and human sox transcription factor gene families. Dev Cell.
3:167–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wegner M: From head to toes: The multiple
facets of sox proteins. Nucleic Acids Res. 27:1409–1420. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheung M, Abu-Elmagd M, Clevers H and
Scotting PJ: Roles of Sox4 in central nervous system development.
Brain Res Mol Brain Res. 79:180–191. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fang CL, Hseu YC, Lin YF, Hung ST, Tai C,
Uen YH and Lin KY: Clinical and prognostic association of
transcription factor SOX4 in gastric cancer. PLoS One.
7:e528042012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zheng JH, Jian ZX, Jin HS, Chen SC and
Wang GY: Expression of SOX4 gene and early recurrence of
hepatocellular carcinoma: Their relationship and the clinical
significance. Nan Fang Yi Ke Da Xue Xue Bao. 30:818–819. 2010.(In
Chinese). PubMed/NCBI
|
|
45
|
Wang L, Zhang J, Yang X, Chang YW, Qi M,
Zhou Z, Zhang J and Han B: SOX4 is associated with poor prognosis
in prostate cancer and promotes epithelial-mesenchymal transition
in vitro. Prostate Cancer Prostatic Dis. 16:301–307. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bao ZQ, Zhang CC, Xiao YZ, Zhou JS, Tao YS
and Chai DM: Over-expression of Sox4 and β-catenin is associated
with a less favorable prognosis of osteosarcoma. J Huazhong Univ
Sci Technolog Med Sci. 36:193–199. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang D, Hao T, Pan Y, Qian X and Zhou D:
Increased expression of SOX4 is a biomarker for malignant status
and poor prognosis in patients with non-small cell lung cancer. Mol
Cell Biochem. 402:75–82. 2015. View Article : Google Scholar : PubMed/NCBI
|