Introduction
Tongue squamous cell carcinoma (TSCC) is one of the
most common malignant tumors of the head and neck worldwide and it
is characterized by its rapid invasion and metastasis (1,2). TSCC has
a number of progression stages, which start with moderate
dysplasia, proceeds to severe dysplasia and eventually invasive
disease (3). TSCC can be cured by
surgical treatment with early detection and diagnosis. Many
therapeutic treatments and efforts have been proposed to prevent
and treat TSCC. However, TSCC mortality rate is still high and
increasing in many countries (4).
Accordingly, identification of new biomarkers or pathways is of
great importance and benefit for the treatment of TSCC.
The microRNAs (miRNAs) are small, 20–24 nucleotides
in length, non-coding RNAs, which regulate the gene expression
post-transcriptionally and have the potential to regulate ~1/3 of
human genome (5–7). The miRNAs modulate the gene expression
by binding to the 3′-UTR of the target mRNA. Most miRNA genes are
solo, and exhibit their functions under the modulation of their own
promoters and regulatory sequences (8).
The miR-184 plays a pivotal role in proliferation in
many cancers. Tung et al found that miR-184 induced
cisplatin resistance in non-small cell lung cancer (NSCLC)
downregulated by E6 oncoprotein (9).
The suppression of miR-184 upregulated SND1 and contributed to
tumor invasion in malignant glioma (10). Foley et al found that the
knockdown of miR-184 could promote neuroblastoma cell
proliferation, increasing the levels of AKT2 (11). The miR-184 also played a role in
epithelia by maintaining the SH2-containing phosphoinositide
5′-phosphatase 2 (SHIP2) level through inhibiting miR-205 (7). In addition, many studies found that
miR-184 also could function as an essential component in
nasopharyngeal carcinoma (12),
hepatocellular carcinoma (HCC) (13),
and TSCC (14).
The Sry-related high-mobility-group box (SOX) genes
are transcription factors, in which the high mobility group (HMG)
box domain is one of the components (15). Sex-determining region Y-box 7 (SOX7)
is grouped into the SOX-F subfamily, which also includes the
sex-determining region Y-box 17 (SOX17) and sex-determining region
Y-box 18 (SOX18) (16). The SOX
proteins have the specific DNA binding sites, which are
5′-(A/T)(A/T)CAA(A/T)G-3′ (17). It
is reported that SOX7 played an important role in various
developmental processes, such as cardiogenesis, endoderm
differentiation, and hematopoiesis (18). The expression of SOX7 was remarkably
downregulated in the breast cancer tissues compared with the
non-tumorous tissues (16). In HCC,
the SOX7 was the direct target of the miR-425, and the
overexpression of the SOX7 could significantly suppress migration
and invasiveness of HCC cells (19).
However, the function of the SOX7 in TSCC progression has not been
determined.
In this study, we found that the miR-184 was
upregulated in TSCC cell lines and tissues compared with the
corresponding adjacent non-tumorous tissues. We further found that
overexpression of miR-184 could promote the migration, and
metastasis of the TSCC cells. Moreover, we identified that SOX7 was
the target of miR-184 and miR-184 could suppress the expression of
SOX7. Taken together, our present study provides a new potential
therapeutic target for TSCC treatment.
Materials and methods
Patient samples
The TSCC tissues and the corresponding non-tumorous
counterparts were obtained from patients of the Affiliated Hospital
of Taishan Medical University (Taian, China). The tissues samples
were frozen in liquid nitrogen and stored at −80°C immediately
after resection from patients who were undergoing surgical
treatment of tongue carcinoma. Written consent of tissue donation
for research purposes was obtained from patients and the research
was approved by the Ethics Committee of the Affiliated Hospital of
Taishan Medical University (Taian, China).
Cell culture
TSCC cell lines Cal27, Tca-8113, and SCC-9 were
purchased from the American Type Culture Collection. All the cell
lines were cultured in RPMI-1640 with 10% fetal bovine serum.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted and purified from the TSCC
tissues and corresponding non-tumorous tissues using the TRIzol
reagent (Invitrogen: Thermo Fisher Scientific, Inc., Carlsbad, CA,
USA) following the manufacturer's instructions. The concentration
and purity of RNA were determined using the NanoDrop 2000
instrument (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
cDNAs were synthesized using the PrimeScript 1st Strand cDNA
Synthesis kit (Takara Biotechnology Co., Ltd., Dalian, China). The
RT-qPCR was carried out using the PrimeScript RT Reagent kit with
gDNA Eraser (Takara Biotechnology Co., Ltd.) on the LightCycler 480
Instrument II (Roche Diagnostics, Basel, Switzerland). The GAPDH
was used as the control for the normalization of expression levels
of genes. The snRNA U6 was used as the internal control for miRNA.
The RT-qPCR data were calculated by the 2−ΔΔCq method
and relative to SnRNA U6 or GAPDH. The conditions for PCR were as
follows: 95°C for 5 min, 40 cycles of denaturation at 95°C (15
sec), 50°C (30 sec) and 72°C (30 sec). The primers were as follows:
miR-184 forward, 5′-TGGACGGAGAACTGATAAGGGT-3′ and reverse,
5′-CCTTATCAGTTCTCCGTCCATT-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; SOX7 forward,
5′-GGGGCCGCCGCCCGAAGCTGATA-3′ and reverse,
5′-CGGGATCCAGAAGGGACCTTGGCTATCA-3′; GAPDH forward,
5′-TTGCCGACAGGATGCAGAAGGA-3′ and reverse,
5′-AGGTGGACAGCGAGGCCAGGAT-3′.
Plasmid construction and small
interfering RNA (siRNA) assay
The miR-184 mimic or inhibitor was constructed for
the overexpression or knockdown of the miR-184. The siRNA was
synthesized and purchased from Takara Biotechnology Co., Ltd.,
which was used for the knockdown of the SOX7. The miR-184 mimic or
inhibitor and siRNA were transfected into Cal27 and Tca-8113 cell
lines using Lipofectamine 2000 reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The sequences
of miR-184 mimics/inhibitor, negative control (NC) were the
following: miR-184 mimic 5′-UGGACGGAGAACUGAUAAGGGU-3′, miR-184
inhibitor 5′-CUGGAGGAAGGGCCCCAGAGG-3′, and the NC is a scrambled
oligonucleotide: 5′-UUCUCCGAACGUGUCACGUTT-3′. miR-184
mimics/inhibitor, NC were designed and synthesized by Guangzhou
RiboBio Co., Ltd. (Guangzhou, China). For SOX7 depletion, siRNA was
synthesized and purified by RiboBio Co., Ltd. (Guangzhou, China).
SOX7-siRNA sequence was as follows: 5′-ACGCCGAGCTGTCGGATGG-3′.
Western blotting
Total protein was extracted from cells using the
radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime
Institute of Biotechnology, Shanghai, China). Protein concentration
was determined using the BCA method. The proteins were separated by
the SDS-PAGE using the Mini-PROTEAN Tetra instrument (Bio-Rad
Laboratories, Inc., Hercules, CA, USA), then transfered onto PVDF
membranes (Bio-Rad Laboratories, Inc.). The membranes were blocked
by 5% BSA and incubated overnight at 4°C with specific primary
antibody rabbit polyclonal anti-SOX7 antibody (ab220293, 1:1,000;
Abcam, Cambridge, UK). After that, the membrane was incubated in
the secondary antibody goat polyclonal anti-rabbit IgG H&L
secondary antibody (ab150077, 1:2,000; Abcam) at room temperature
for 1 h. Anti-GAPDH antibody (sc-25778; 1:3,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) was used as a loading control
at 4°C overnight. Signal was measured using the GelDoc 2000
instrument (Bio-Rad Laboratories, Inc.).
Luciferase reporter assay
A sequence of 3′-UTR of SOX7 containing the
predicted miR-184 binding site was synthesized and inserted into
the p-MIR-reporter plasmid (p-MIR-SOX7-3′-UTR-WT) (Ambion: Thermo
Fisher Scientific, Inc., Foster City, CA, USA). Simultaneously, a
sequence that contains seven mutant nucleotides of the 3′-UTR of
SOX7 was inserted into the the p-MIR-reporter plasmid
(p-MIR-SOX7-3′-UTR-MU). For the luciferase assay, the Cal27 and
Tca-8113 cells were transfected with firefly luciferase reporter
plasmid, miR-184 mimic, or the NC using the Lipofectamine 2000
(Thermo Fisher Scientific, Inc.). The pMIR-REPORT β-galactosidase
vector was used as the control. The luciferase activity was
measured using the ONE-Glo Luciferase Assay instrument (Promega
Corp., Madison, WI, USA).
Cell proliferation assays
In this study, we used the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
as the monitor of the cell proliferation. After transfection, the
cells were cultured for 1–4 days. The cells were incubated for 4 h
at 37°C after adding the MTT. The supernatant was removed and
dissolved the formazan crystals using DMSO (150 µl/well). The
absorbance at 490 nm for each sample was measured using the
Multilabel Plate Reader (PerkinElmer, Inc., Waltham, MA, USA).
Clone formation assay
Cal27 and Tca-8113 cells were seeded on a 24-well
plate at a density of 300 cells/well. Then the cells were
transfected with miR-184 mimic, NC, miR-184 inhibitor using
Lipofectamine 2000. Subsequently, the cells were cultured in a
humidified chamber at 37°C containing 5% CO2 for 14
days. When clones were macroscopic, the culture process was
terminated. Then the cells were fixed by applying neutral methyl
alcohol 4% for 15 min and stained with Wright-Giemsa compound stain
for 30 min. Finally, the number of clones was counted under a
microscope (Olympus, Tokyo, Japan).
Statistical analysis
Student's t-test and Tukey's post hoc test after
ANOVA was used to determine the significant difference between the
groups. All the experiments were performed in three independent
replicates. The results are presented as the mean ± SD and the
differences were considered statistically significant at P<0.05.
Correlation between mRNA and miRNA were estimated using the
Spearman's correlation method. SPSS v.19.0 software (IBM Corp.,
Armonk, NY, USA) was used to perform statistical analyses and
GraphPad Prism 5.02 (GraphPad Software, Inc., La Jolla, CA, USA) to
complete graph presentation.
Results
miR-184 expression is upregulated and
SOX7 expression is downregulated in TSCC cell lines and
tissues
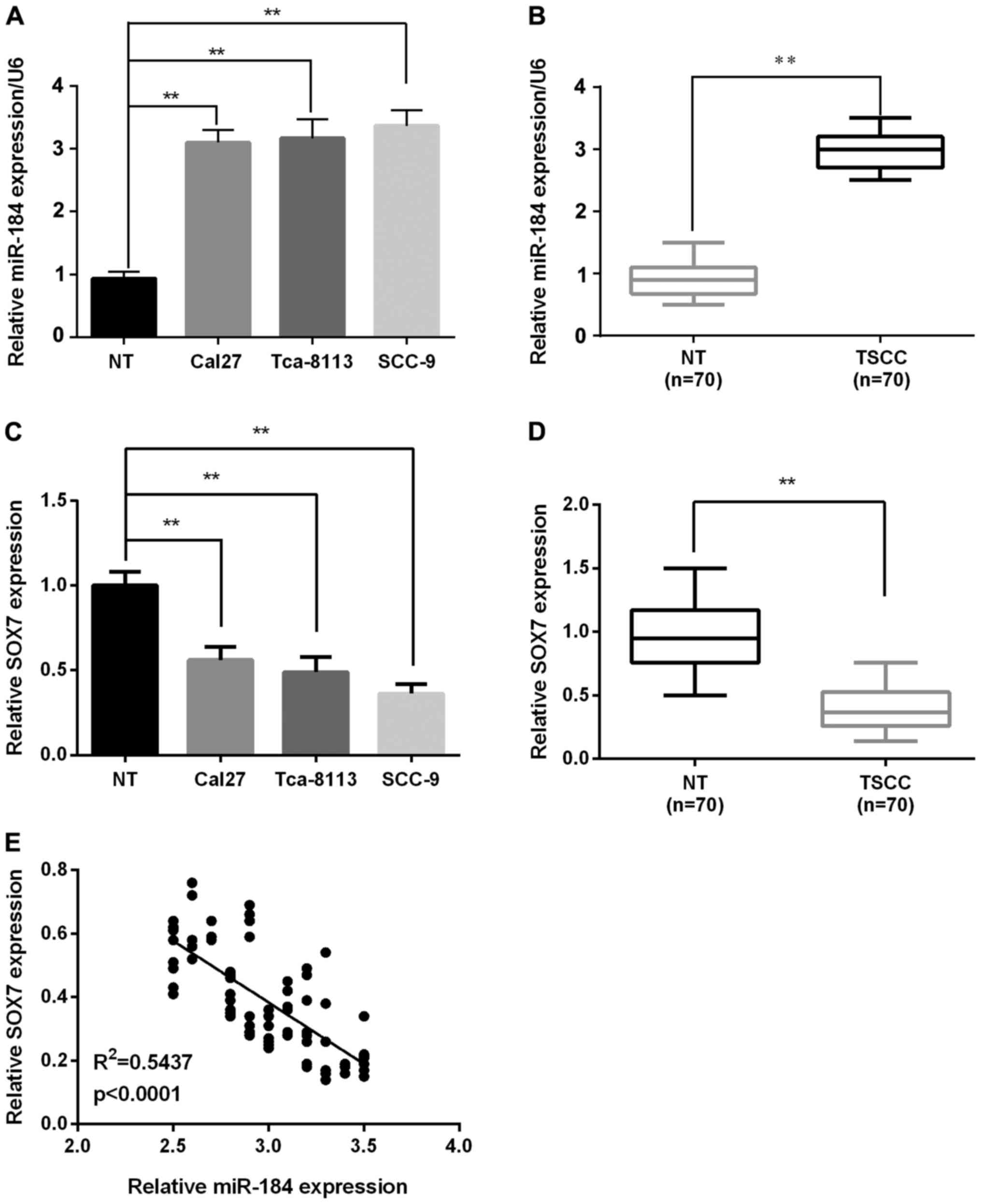
RT-qPCR analysis showed that the miR-184 expression
was significantly increased in all cell lines (Cal27, Tca-8113, and
SCC-9) compared with the normal cell line (Fig. 1A). In TSCC tumors, miR-184 expression
was remarkably upregulated in 70 pairs of TSCC tissues compared
with the corresponding adjacent non-tumorous tissues (Fig. 1B). However, SOX7 expression in TSCC
cells and tissues was decreased significantly compared to normal
tissues by RT-qPCR (Fig. 1C and D).
Furthermore, to evaluate miR-184 modulation of SOX7 at the gene
expression level, the association between the expression of miR-184
and SOX7 was investigated in these clinical specimens. It was
demonstrated that miR-184 expression was negatively correlated with
SOX7 expression in TSCC (R2=0.5437; P<0.0001;
Fig. 1E), indicating that miR-184
regulates the expression of SOX7.
miR-184 promotes proliferation of the
TSCC cells in vitro
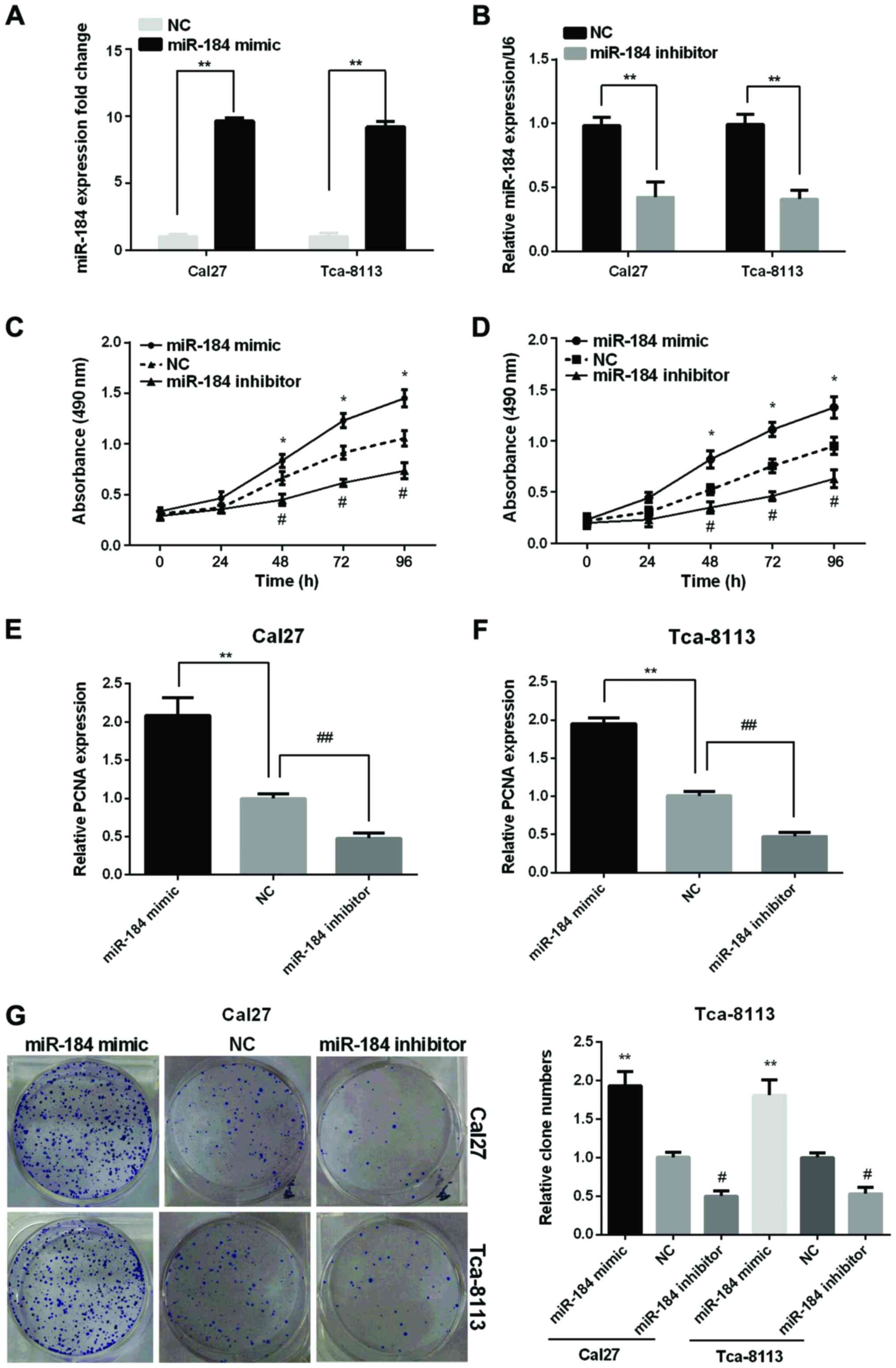
To better understand the function of miR-184 in the
TSCC tissues, we transfected the miR-184 mimic or inhibitor to the
TSCC cell lines (Cal27, Tca-8113). The successful overexpression or
silencing of miR-184 was confirmed by the RT-qPCR (Fig. 2A and B). The results demonstrated that
the miR-184 expression levels were remarkably increased in the two
TSCC cell lines after transfected with miR-184 mimic while
decreased after transfected with miR-184 inhibitor compared with
the NC.
The influence of miR-184 on the proliferation of the
TSCC cell lines was investigated using the MTT, RT-qPCR and clone
formation assays. MTT results showed that the cell viability in the
two cell lines was significantly increased by miR-184 mimic, but
reduced by miR-184 inhibitor (Fig. 2C and
D). RT-qPCR analysis showed that the proliferating cell nuclear
antigen (PCNA) expression was raised remarkably in the two cell
lines after transfected with miR-184 mimic while declined after
transfection with miR-184 inhibitor (Fig.
2E and F). As expected, clone formation assay showed an
increased clone number in miR-184 mimic group while decreased clone
number in miR-184 inhibitor group compared to control group
(Fig. 2G).
SOX7 is a direct target of miR-184 and
miR-184 regulates the expression of SOX7
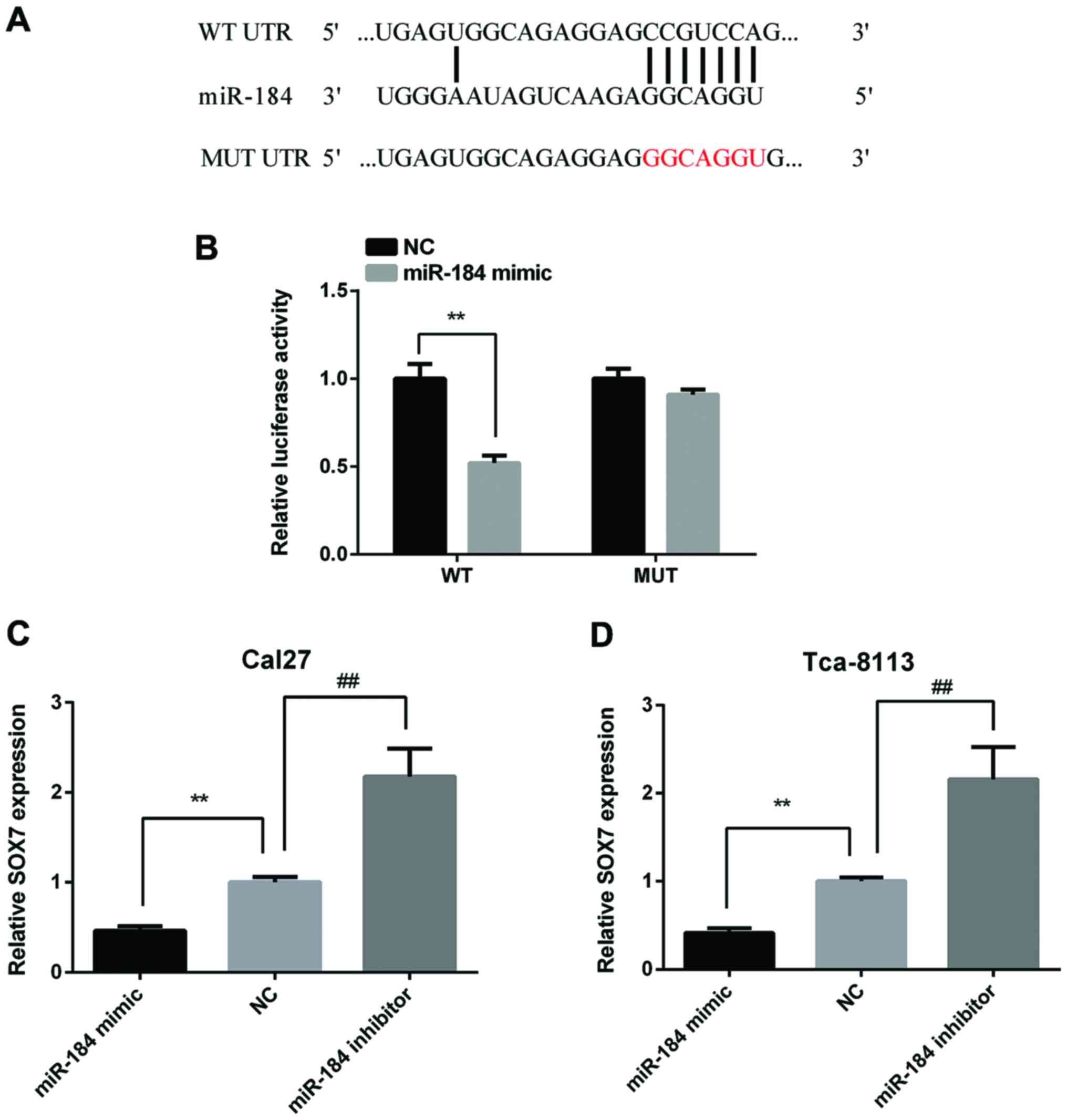
In order to discover the target gene of miR-184,
bioinformatical prediction was performed using the TargeScan
website (http://www.targetscan.org/mamm_31/). As Fig. 3A shows, SOX7 has a binding site at the
36–42 bp of the 3′-UTR for miR-184. To verify this prediction,
Cal27 cells were transfected with the p-MIR-SOX7-3′-UTR-WT or
p-MIR-SOX7-3′-UTR-MT luciferase reporter and miR-184 mimic,
respectively. The relative luciferase activity was significantly
reduced in Cal27 cells co-transfected with p-MIR-SOX7-3′-UTR-WT
luciferase reporter and miR-184 mimic than the NC cells. However,
there were no significance between Cal27 cells co-transfected with
p-MIR-SOX7-3′-UTR-MU luciferase reporter and miR-184 mimic and the
NC cells (Fig. 3B). Then, we detected
SOX7 expression in Cal27 and Tca-8113 cells after transfected with
miR-184 mimic or inhibitor. The results showed that miR-184 mimic
reduced SOX7 expression and miR-184 inhibitor increased SOX7
expression in both two cell lines (Fig.
3C and D).
Overexpression of SOX7 inhibits
proliferation of TSCC cells in vitro
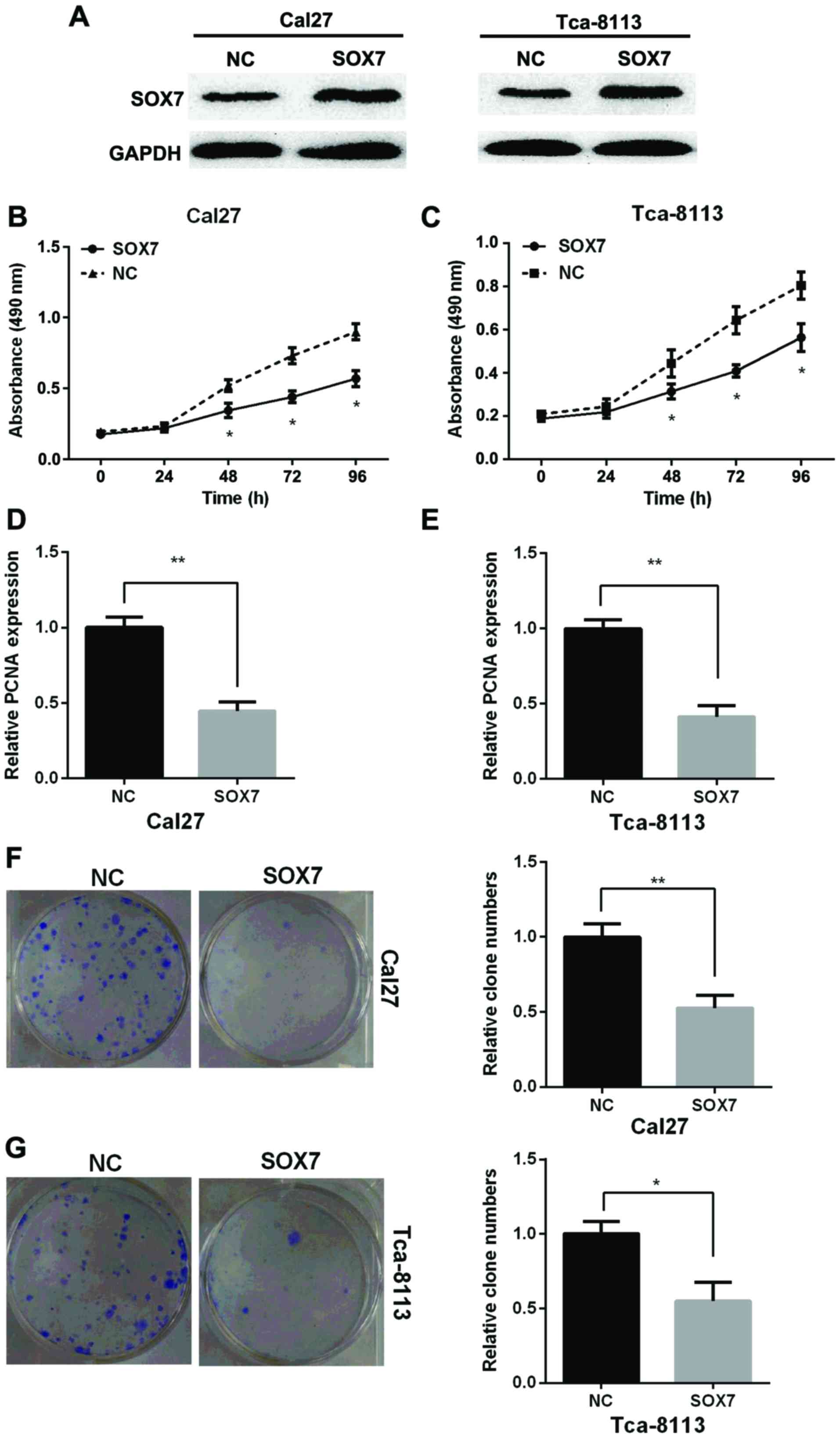
To detect the role of SOX7 on the proliferation of
TSCC, Cal27 and Tca-8113 cell lines were transfected with SOX7
vector or the NC to overexpress of SOX7. SOX7 expression in protein
level was measured by western blotting assay in Cal27 and Tca-8113
cells and the results showed that SOX7 expression was increased
after overexpression of SOX7 compared with NC (Fig. 4A). MTT assay showed that
overexpression of SOX7 inhibited the proliferation of Cal27 and
Tca-8113 cells (Fig. 4B and C). PCNA
expression was decreased after cells were transfected with SOX7
vector in the two TSCC cell lines by RT-qPCR (Fig. 4D and E). Clone formation assay showed
that overexpression of SOX7 could suppress the relative clone
number in the two TSCC cell lines (Fig.
4F). Taken together, overexpression of SOX7 inhibited
proliferation of TSCC cells in vitro.
Discussion
TSCC is the most common type of oral squamous cell
carcinoma and it is well known for its high rate of lymph nodal
metastasis. Although huge efforts have been made to prevent and
cure TSCC, the number of deaths increased during the past 5 years
(20). It is meaningful to study the
related gene expression and miRNA alteration. Dysregulation of
miRNAs is a distinct feature of cancers, which derive from
dysregulated gene expression (14).
It is reported that miRNA alteration could induce carcinogenesis
(21) and miR-184 played an important
role in many cancers, such as NSCLC (9), neuroblastoma (11), nasopharyngeal carcinoma (12), and HCC (13). In our study, we found that miR-184 was
significantly upregulated in the TSCC tissues compared with the
adjacent non-tumorous tissues and promoted the proliferation of
TSCC cells. PCNA is a DNA polymerase sliding clamp that serves in
DNA replication and repair (22). A
previous study demonstrated that a high PCNA index resulted in an
increase in cyclin E2 and proliferative activity in pancreatic
cancer cells (23). The results of
the current study demonstrated that the PCNA expression was raised
remarkably in two cell lines after transfected with miR-184 mimic
while declined after transfected with miR-184 inhibitor. This
further indicates that miR-184 promoted the proliferation by
dysregulating the expression of PCNA, moreover, the absence of cell
cycle analysis may be a limitation of our study. In human HCC,
miR-184 was upregulated in HCC cell lines and overexpression of
miR-184 could increase cell proliferation through targeting SOX7
(13). In contrast to our research,
miR-184 expression was downregulated in human malignant glioma
tumor tissues compared with their normal counterparts (10). This exhibits the complicated functions
of miR-184 in the regulation of gene expression. Using the
TargetScan and the luciferase reporter assay, we predicted and
confirmed that miR-184 bound to the 3′-UTR of SOX7. The luciferase
activity was reduced significantly in cell lines co-transfected
with p-MIR-SOX7-3′-UTR-WT luciferase reporter and miR-184 mimic
than the NC cells.
SOX7 has been found to be involved in regulation of
many cancers and to functioned as a suppressor. In the human breast
cancer cells, miR-492 promoted the cell cycle and proliferation
through inhibiting SOX7 (24). Liu
et al found that SOX7 expression was remarkably reduced in
the ovarian cancer tissues compared with the non-cancerous tissues
and played this role through Wnt/β-catenin signaling pathway
(25). In the gastric cancer, miR-935
increased tumorigenesis and cell proliferation by targeting SOX7
(26). Also, SOX7 played an important
role in acute myeloid leukemia, and functioned as a tumor
suppressor (27). SOX7 contains HMG
box, and other important functional domains at the carboxyl and
amino terminal regions. The HMG box plays an important role in DNA
bending and binding, protein transportation and interaction
(15,28). These important domains may explain why
the SOX7 could function as tumor suppressor in a number of cancers.
Consistent with these identifications, our study showed that
overexpression of the SOX7 inhibited the proliferation of TSCC
cells and miR-184 could suppress the expression of SOX7 using the
luciferase reporter assays.
In conclusion, our results demonstrated that miR-184
was significantly upregulated in the TSCC tissues compared with the
non-tumorous tissues and miR-184 promoted TSCC cell proliferation.
We found that miR-184 bound to the 3′-UTR of SOX7 and suppressed
SOX7 expression. Therefore, our results demonstrated that the
miR-184/SOX7 axis might provide a promising therapeutic target for
TSCC treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DC and JL contributed to the conception of the
study. SL and PH contributed significantly to the data analysis and
study preparation. YW performed the data analyses and wrote the
study. SD, PH and NL helped perform the analysis with constructive
discussions and conducted the experiment. All authors have read and
approved the final study.
Ethics approval and consent to
participate
Written consent of tissue donation for research
purposes was obtained from patients and the research was approved
by the Ethics Committee of the Affiliated Hospital of Taishan
Medical University (Taian, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang W, Liu Z, Zhao L, Sun J, He Q, Yan W,
Lu Z and Wang A: Hexokinase 2 enhances the metastatic potential of
tongue squamous cell carcinoma via the
SOD2-H2O2pathway. Oncotarget. 8:3344–3354.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rosebush MS, Rao SK, Samant S, Gu W,
Handorf CR, Pfeffer LM and Nosrat CA: Oral cancer: Enduring
characteristics and emerging trends. J Mich Dent Assoc. 94:64–68.
2012.PubMed/NCBI
|
|
3
|
Chen C, Méndez E, Houck J, Fan W,
Lohavanichbutr P, Doody D, Yueh B, Futran ND, Upton M, Farwell DG,
et al: Gene expression profiling identifies genes predictive of
oral squamous cell carcinoma. Cancer Epidemiol Biomarkers Prev.
17:2152–2162. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fang Z, Wu L, Wang L, Yang Y, Meng Y and
Yang H: Increased expression of the long non-coding RNA UCA1 in
tongue squamous cell carcinomas: A possible correlation with cancer
metastasis. Oral Surg Oral Med Oral Pathol Oral Radiol. 117:89–95.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pillai RS, Bhattacharyya SN and Filipowicz
W: Repression of protein synthesis by miRNAs: How many mechanisms?
Trends Cell Biol. 17:118–126. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu J, Ryan DG, Getsios S,
Oliveira-Fernandes M, Fatima A and Lavker RM: MicroRNA-184
antagonizes microRNA-205 to maintain SHIP2 levels in epithelia.
Proc Natl Acad Sci USA. 105:19300–19305. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tung MC, Lin PL, Cheng YW, Wu DW, Yeh SD,
Chen CY and Lee H: Reduction of microRNA-184 by E6 oncoprotein
confers cisplatin resistance in lung cancer via increasing Bcl-2.
Oncotarget. 7:32362–32374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Emdad L, Janjic A, Alzubi MA, Hu B,
Santhekadur PK, Menezes ME, Shen XN, Das SK, Sarkar D and Fisher
PB: Suppression of miR-184 in malignant gliomas upregulates SND1
and promotes tumor aggressiveness. Neuro Oncol. 17:419–429. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Foley NH, Bray IM, Tivnan A, Bryan K,
Murphy DM, Buckley PG, Ryan J, O'Meara A, O'Sullivan M and
Stallings RL: MicroRNA-184 inhibits neuroblastoma cell survival
through targeting the serine/threonine kinase AKT2. Mol Cancer.
9:832010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhen Y, Liu Z, Yang H, Yu X, Wu Q, Hua S,
Long X, Jiang Q, Song Y, Cheng C, et al: Tumor suppressor PDCD4
modulates miR-184-mediated direct suppression of C-MYC and BCL2
blocking cell growth and survival in nasopharyngeal carcinoma. Cell
Death Dis. 4:e8722013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu GG, Li WH, He WG, Jiang N, Zhang GX,
Chen W, Yang HF, Liu QL, Huang YN, Zhang L, et al: Mir-184
post-transcriptionally regulates SOX7 expression and promotes cell
proliferation in human hepatocellular carcinoma. PLoS One.
9:e887962014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wong TS, Liu XB, Wong BY, Ng RW, Yuen AP
and Wei WI: Mature miR-184 as potential oncogenic microRNA of
squamous cell carcinoma of tongue. Clin Cancer Res. 14:2588–2592.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lefebvre V, Dumitriu B, Penzo-Méndez A,
Han Y and Pallavi B: Control of cell fate and differentiation by
Sry-related high-mobility-group box (Sox) transcription factors.
Int J Biochem Cell Biol. 39:2195–2214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu H, Mastriani E, Yan ZQ, Yin SY, Zeng
Z, Wang H, Li QH, Liu HY, Wang X, Bao HX, et al: SOX7 co-regulates
Wnt/β-catenin signaling with Axin-2: Both expressed at low levels
in breast cancer. Sci Rep. 6:261362016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harley VR, Lovell-Badge R and Goodfellow
PN: Definition of a consensus DNA binding site for SRY. Nucleic
Acids Res. 22:1500–1501. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stovall DB, Cao P and Sui G: SOX7: From a
developmental regulator to an emerging tumor suppressor. Histol
Histopathol. 29:439–445. 2014.PubMed/NCBI
|
|
19
|
Zheng Z, Liu J, Yang Z, Wu L, Xie H, Jiang
C, Lin B, Chen T, Xing C, Liu Z, et al: MicroRNA-452 promotes
stem-like cells of hepatocellular carcinoma by inhibiting Sox7
involving Wnt/β-catenin signaling pathway. Oncotarget.
7:28000–28012. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu X and Li Z: MicroRNA expression and its
implications for diagnosis and therapy of tongue squamous cell
carcinoma. J Cell Mol Med. 20:10–16. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shiomi N, Mori M, Tsuji H, Imai T, Inoue
H, Tateishi S, Yamaizumi M and Shiomi T: Human RAD18 is involved in
S phase-specific single-strand break repair without PCNA
monoubiquitination. Nucleic Acids Res. 35:e92007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Deng J, He M, Chen L, Chen C, Zheng J and
Cai Z: The loss of miR-26a-mediated post-transcriptional regulation
of cyclin E2 in pancreatic cancer cell proliferation and decreased
patient survival. PLoS One. 8:e764502013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen F, Cai WS, Feng Z, Li JL, Chen JW,
Cao J and Xu B: MiR-492 contributes to cell proliferation and cell
cycle of human breast cancer cells by suppressing SOX7 expression.
Tumour Biol. 36:1913–1921. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu H, Yan ZQ, Li B, Yin SY, Sun Q, Kou
JJ, Ye D, Ferns K, Liu HY and Liu SL: Reduced expression of SOX7 in
ovarian cancer: A novel tumor suppressor through the Wnt/β-catenin
signaling pathway. J Ovarian Res. 7:872014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang M, Cui G, Ding M, Yang W, Liu Y, Dai
D and Chen L: miR-935 promotes gastric cancer cell proliferation by
targeting SOX7. Biomed Pharmacother. 79:153–158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Man CH, Fung TK, Wan H, Cher CY, Fan A, Ng
N, Ho C, Wan TS, Tanaka T, So CW, et al: Suppression of SOX7 by DNA
methylation and its tumor suppressor function in acute myeloid
leukemia. Blood. 125:3928–3936. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schepers GE, Teasdale RD and Koopman P:
Twenty pairs of sox: Extent, homology, and nomenclature of the
mouse and human sox transcription factor gene families. Dev Cell.
3:167–170. 2002. View Article : Google Scholar : PubMed/NCBI
|