Introduction
Osteosarcoma is the most common malignant bone
tumor, with characteristics that include hematogenous metastasis
and local invasion (1–3). Osteosarcoma primarily occurs in children
aged 10–14 years old (2); the
metaphysis of long bones is the most susceptible area, particularly
the distal femur and proximal tibia. Patients without lung
metastasis have a five-year survival rate range from 50 to 70%
following radical tumor resection, whereas those with lung
metastasis have a <20% five-year survival rate (4). The lung is the most common site of
metastasis, followed by the bones (5).
At present, the standard treatment strategies
include preoperative chemotherapy, surgical resection and
postoperative chemotherapy. Although various chemotherapeutics have
been developed, a large proportion of patients continue to succumb
to disease recurrence and metastasis. The main reasons for
treatment failure are early metastasis and the development of drug
resistance. Therefore, it is necessary to explore novel alternative
treatment options, particularly gene therapy (6,7), to
prevent early metastasis, slow osteosarcoma progression and improve
the quality of life and survival rate for patients with
osteosarcoma.
Circadian rhythm is also known as the ‘biological
rhythm’ and is a cycle of ~24 h. Nearly all living beings have a
biological rhythm, which resembles the Earth's day and night cycle.
This phenomenon is part of the biological evolution and the natural
selection process. Biological rhythm affects not only
macro-organisms, but also in various organs and even single cells,
so biological rhythm is one of the most important basic
characteristics of biological activity. A number of biochemical and
physiological indexes, including body temperature, heart rate,
blood pressure (8) and sleep-wake
cycle (9), are significantly affected
by circadian rhythm. The molecular biological foundation of these
processes are the circadian rhythm genes, which serve important
roles in the maintenance of circadian rhythm, as well as affecting
other gene expression and biochemical processes (10). The disruption of circadian rhythm
leads to the occurrence and development of certain diseases
(11,12). A circadian variation in the onset of
stroke (13) and myocardial
infarction (14) has been
demonstrated, and workers who work night shifts have a greater
chance of developing diabetes (15).
Circadian clock genes, such as Period 2 (per2) and Bmal1, have been
demonstrated to serve important roles in angiogenesis (16).
Per2 is a core circadian clock gene. Per2 protein,
which affects various signaling pathways, including the protein
kinase B (Akt) cascade, (17) to
regulate the biological activities of the body, is expressed
variably in different tissues (18).
Dysregulation of the per2 gene has been identified in various types
of human cancer (12,19), including breast cancer (20), hepatoma (21), colorectal cancer (22) and pancreatic ductal adenocarcinoma
(23), and may be associated with a
poor prognosis. However, there is limited data regarding whether
and how the circadian gene per2 affects the functions of
osteosarcoma cells. As further research is required on this topic,
the present study aimed to investigate the effect of per2 in
osteosarcoma cells. To achieve this, a eukaryotic expression vector
containing per2 or per2 small interfering (si)-RNA was transfected
into MNNG/HOS osteosarcoma cells. Following transfection, the
effect on cell proliferation, migration and apoptosis were
analyzed. Furthermore, in order to investigate the mechanisms of
per2 in MNNG/HOS cells, the effects of per2 overexpression and
knockdown on proliferation and apoptosis-associated proteins and
the Akt signal pathway were examined.
Materials and methods
Cell culture and transfection
The MNNG/HOS Cl #5 human osteosarcoma cell line
(cat. no. R-1059-D) was purchased from the Cell Resource Center of
the Shanghai Institute of Life Sciences, Chinese Academy of
Sciences (Shanghai, China). MNNG/HOS cells were cultured in
Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10%
fetal bovine serum (FBS; both Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), 100 IU/ml penicillin and 100 IU/ml
streptomycin. Cells were cultured at 37°C in a humidified
atmosphere with 5% CO2.
The cells were allocated into five groups, i.e.,
blank control (no treatment), pcDNA3.1-per2, pcDNA3.1-only, per2
siRNA and control siRNA groups. Cells were transfected with the
plasmid and siRNA with Lipofectamine 2000 reagent (Thermo Fisher
Scientific, Inc.). Briefly, according to the manufacturer's
protocol, the transfection complex was prepared based on an
optimized proportion of plasmid or siRNA and
Lipofectamine® 2000 reagent, which was transfected into
MNNG/HOS cells at 70–80% confluence. After 48 h, cells were washed
with PBS and collected for subsequent assays.
Immunofluorescence
Cells were plated on coverslips in six-well plates
and transfected as described, cells were fixed with 4%
paraformaldehyde at room temperature for 10 min and followed by
0.1% Triton X-100 for 10 min, blocked with 1% bovine serum albumin
(cat no. 10270106; Gibco; Thermo Fisher Scientific, Inc.) for 20
min at room temperature, and incubated with rabbit anti-per2
(dilution, 1:250; cat no. ab180655; Abcam, Cambridge, UK) overnight
at 4°C. Subsequent to incubation with an fluorescein
isocyanide-conjugated AffiniPure goat anti-rabbit secondary
antibody (dilution, 1:1,000; cat no. 1095047; Cell Signaling
Technology, Danvers, MA, USA) for 1 h at room temperature, cells
were stained with DAPI (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 15 min at room temperature. Photomicrographs were then
captured from laser confocal microscopy (LCSM, Zeiss KS 400; Zeiss
AG, Oberkochen, Germany).
Cell proliferation assay
Cell proliferation was detected using cell counting
kit-8 (CCK8, cat no. C0038; Beyotime Institute of Biotechnology,
Haimen, China). Following transfection as described, cells were
seeded at a density of 5,000 cells per well in 96-well plates.
Following starvation (growth without serum) for 24 h, an aliquot of
10 µl CCK-8 solution was added to the wells and incubated for 2 h
at 37°C. Absorbance was then measured at 450 nm to determine the
optical density (OD) value.
Cell migration assay
The migration assay was performed using an 8-µm pore
size Transwell invasion chamber (24-well; Corning Incorporated,
Corning, NY, USA) according to the manufacturer's protocol.
Briefly, 1×105 cells were transferred to the top chamber
in 200 µl serum-free DMEM, and 500 µl DMEM supplemented with 10%
FBS was added to the lower chambers. After a 24-h incubation, cells
that remained in the top chamber were removed with a cotton swab,
and the migrated cells in the bottom chamber were fixed with 4%
formaldehyde for 10 min at room temperature and stained with
hematoxylin for 10 min at room temperature. Cells were photographed
using an inverted microscope and cell numbers were quantified by
ImageJ software (1.51; National Institutes of Health, Bethesda, MD,
USA).
TUNEL staining
Preconditioned cells were collected for terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling
(TUNEL) apoptosis assay using a In Situ Cell Death Detection
kit (Merck KGaA) according to the manufacturer's protocol. The rate
of apoptosis was calculated as the ratio of TUNEL-positive cells to
total cells as determined under a light microscope (Leica DM 3000;
Leica Microsystems GmbH, Wetzlar, Germany) in five randomly
determined fields.
Western blot analysis
Cells were solubilized in cold
radioimmunoprecipitation lysis buffer (Beyotime Institute of
Biotechnology) to extract protein. The protein concentration was
detected using a BCA kit (Beyotime Institute of Biotechnology).
Cell lysates containing 30 µg of protein were separated with 10%
SDS-PAGE and transferred to polyvinylidene fluoride membranes (GE
Healthcare, Chicago, IL, USA). Subsequent to blocking with 5% skim
milk at room temperature for 1 h, membranes were incubated
overnight at 4°C with the following primary antibodies: Rabbit
anti-per2 (dilution, 1:1,000; cat no. ab180655; Abcam), rabbit
anti-Akt (dilution, 1:3,000; cat no. 4691s), rabbit
anti-phosphorylated (p)-Akt (dilution, 1:3,000; cat no. 4060s),
rabbit anti-Bcl-2 (dilution, 1:500; cat no. 3498s), rabbit anti-p21
(dilution 1:5,000; cat no. 2947s), rabbit anti-p27 (dilution,
1:500; cat no. 3686s; all Cell Signaling Technology), rabbit
anti-caspase-3 (dilution, 1:250; cat no. ab44976; Abcam), rabbit
anti-cleaved caspase-3 (dilution, 1:500; cat no. 9664s; Cell
Signaling Technology), rabbit anti-CDK4 (dilution, 1:500; cat no.
ab68266; Abcam) and mouse anti-β-actin (dilution, 1:1,000; cat no.
4970s; Cell Signaling Technology). The membranes were washed 3
times in Tris-buffered saline with Tween-20 (TBST) for 10 min, then
incubated with anti-rabbit or anti-mouse horseradish
peroxidase-conjugated IgG (dilution, 1:5,000; cat nos. TA130023 and
TA130003, respectively; OriGene Technologies, Inc., Beijing, China)
at room temperature for 1 h. The membranes were washed 3 times in
TBST for 15 min. Finally, membranes were treated with an enhanced
chemiluminescence reagent (Merck KGaA). Imaging apparatus (AI 600
RGB; GE Healthcare) was used to detect chemiluminescence according
to the manufacturer's protocol.
Statistical analysis
Data were expressed as the mean ± standard
deviation. SPSS 17.0 (SPSS Inc., Chicago, IL, USA) was used to
conduct statistical analysis. A t test was used to compare
differences between two groups, and one-way analysis of variance
followed by a least significant difference post-hoc test was used
for the comparison of multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
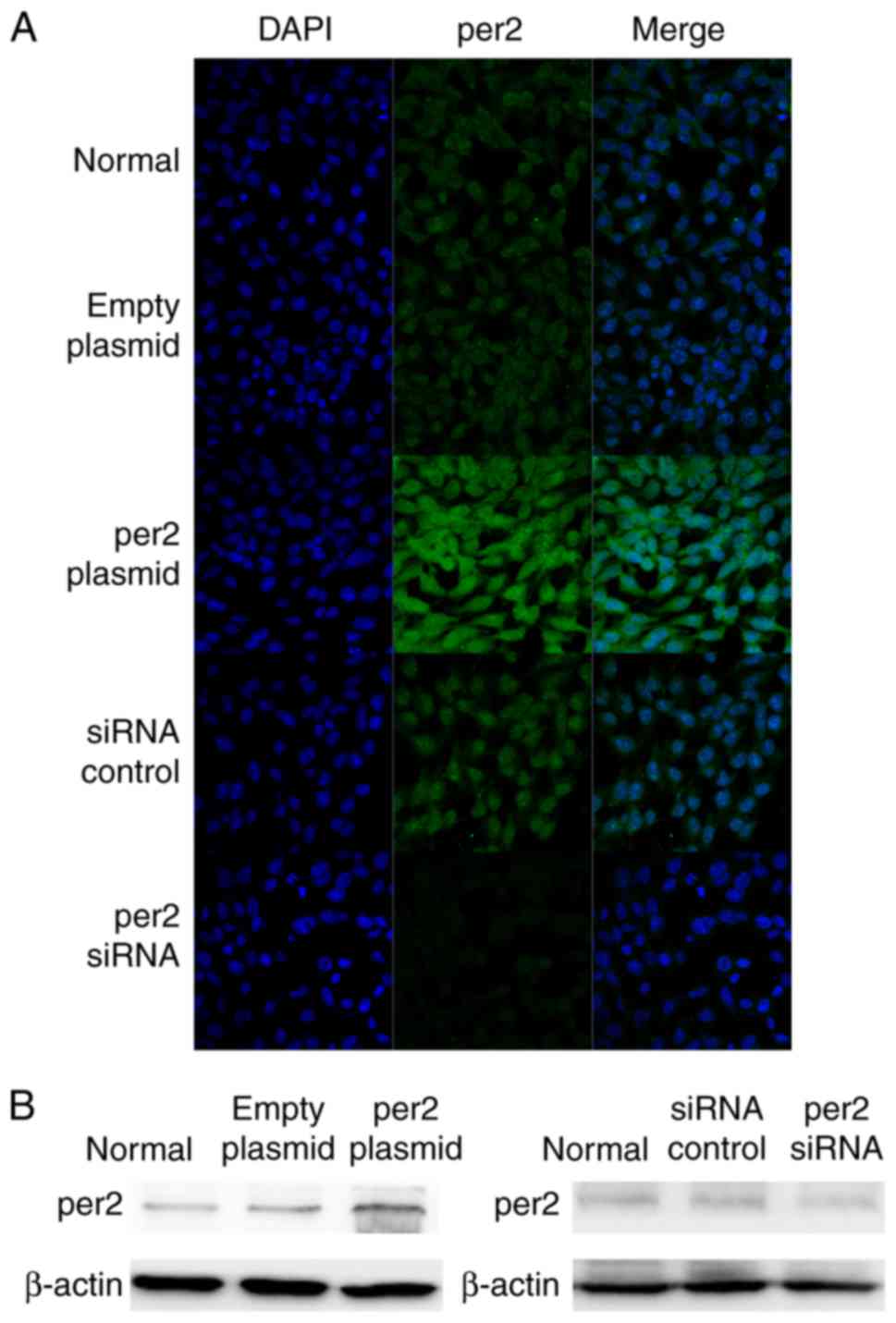
Per2 was successfully overexpressed
and downregulated
Following transfection for 24 h, per2 expression was
detected by the immunofluorescence staining of the five groups of
cells (i.e., Blank control, per2 overexpression, plasmid-only, per2
siRNA and control siRNA groups). The per2 overexpression group
exhibited an increase in per2 fluorescence intensity, while there
was reduced intensity in the per2 siRNA group. There were no
differences between the blank control, plasmid-only and siRNA
control groups (Fig. 1A). Western
blot analysis was also performed to investigate per2 protein
expression. The results of western blotting corresponded with the
results of immunofluorescence.
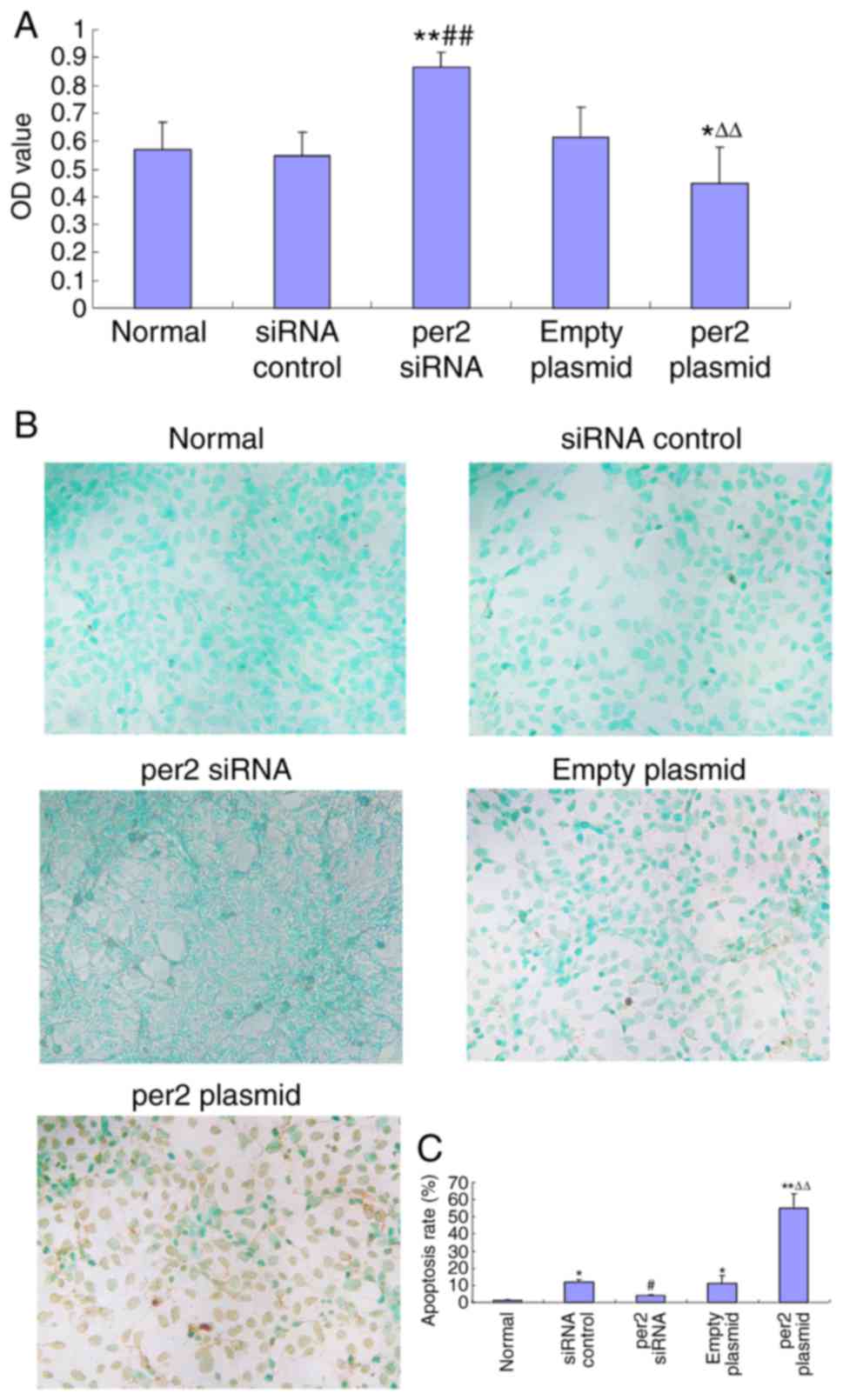
Per2 overexpression decreased
osteosarcoma cell proliferation and prevented apoptosis, in
contrast with per2 knockdown
The CCK8 assay was used to analyze MNNG/HOS cell
proliferation. Per2 overexpression induced a significant decrease
in proliferation (Fig. 2A).
Conversely, per2 knockdown significantly upregulated proliferation
(Fig. 2A). There were no
statistically significant differences between the blank control,
plasmid-only and siRNA control groups. A TUNEL assay was also used
to analyze the rate of MNNG/HOS cell apoptosis. The results
indicated that the per2 overexpression group demonstrated an
increase in apoptosis whereas per2 siRNA prevented apoptosis.
Plasmid-only and control siRNA groups did exhibited a significantly
increased rate of apoptosis compared with blank control
(P<0.05). The reason may be that damage was caused by
Lipofectamine 2000. Per2 Plasmid was significantly increased
compared with the normal control (P<0.01) and the empty plasmid
(P<0.01). Per2 siRNA was significantly decreased compared with
siRNA control (P<0.05) (Fig. 2B and
C).
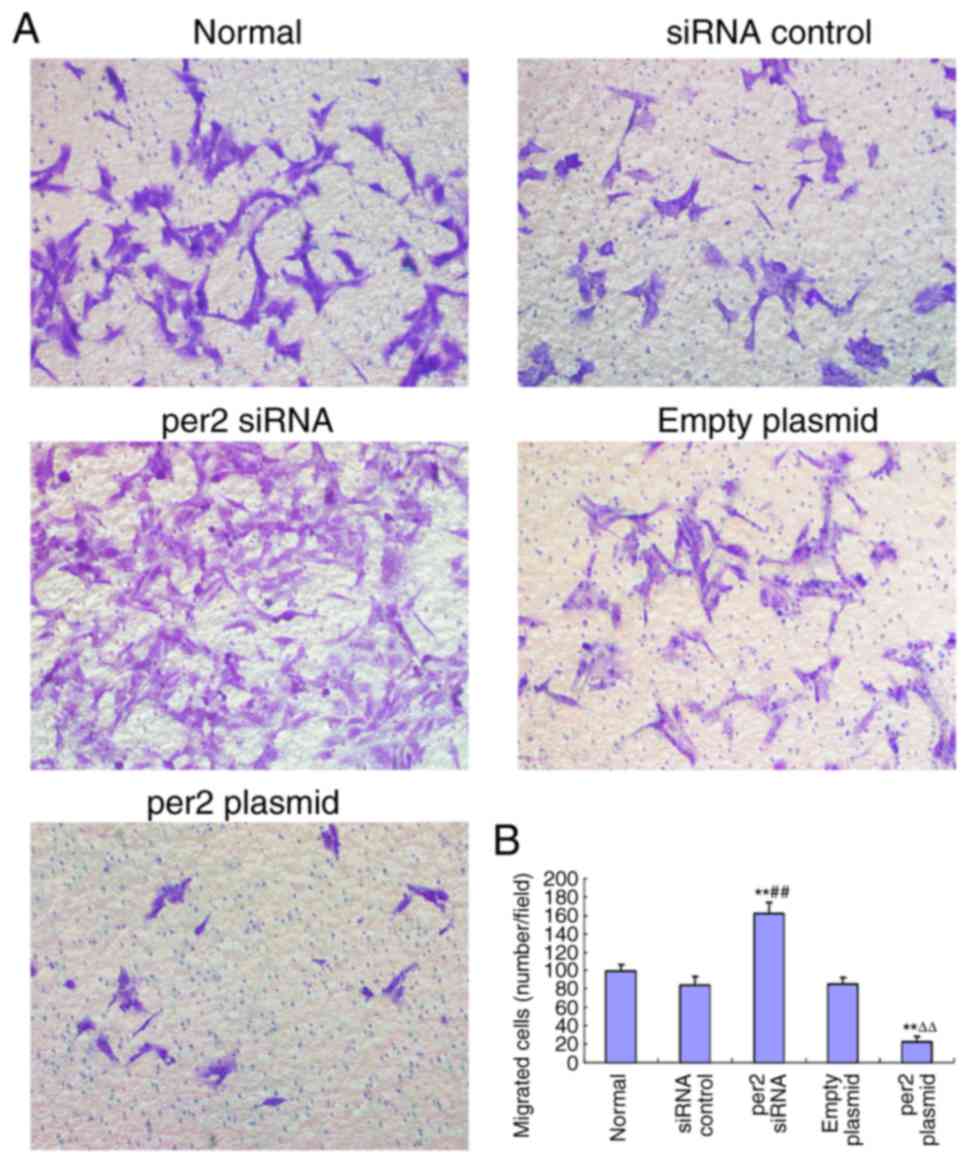
Per2 overexpression decreases MNNG/HOS
cell migration, in contrast with per2 knockdown
The migration ability of MNNG/HOS cells was
evaluated with a Transwell assay. It was demonstrated that per2
overexpression decreased MNNG/HOS cell migration, whereas per2
siRNA increased MNNG/HOS cell migration. There were no
statistically significant differences between the blank control,
plasmid-only and siRNA control groups (Fig. 3A and B).
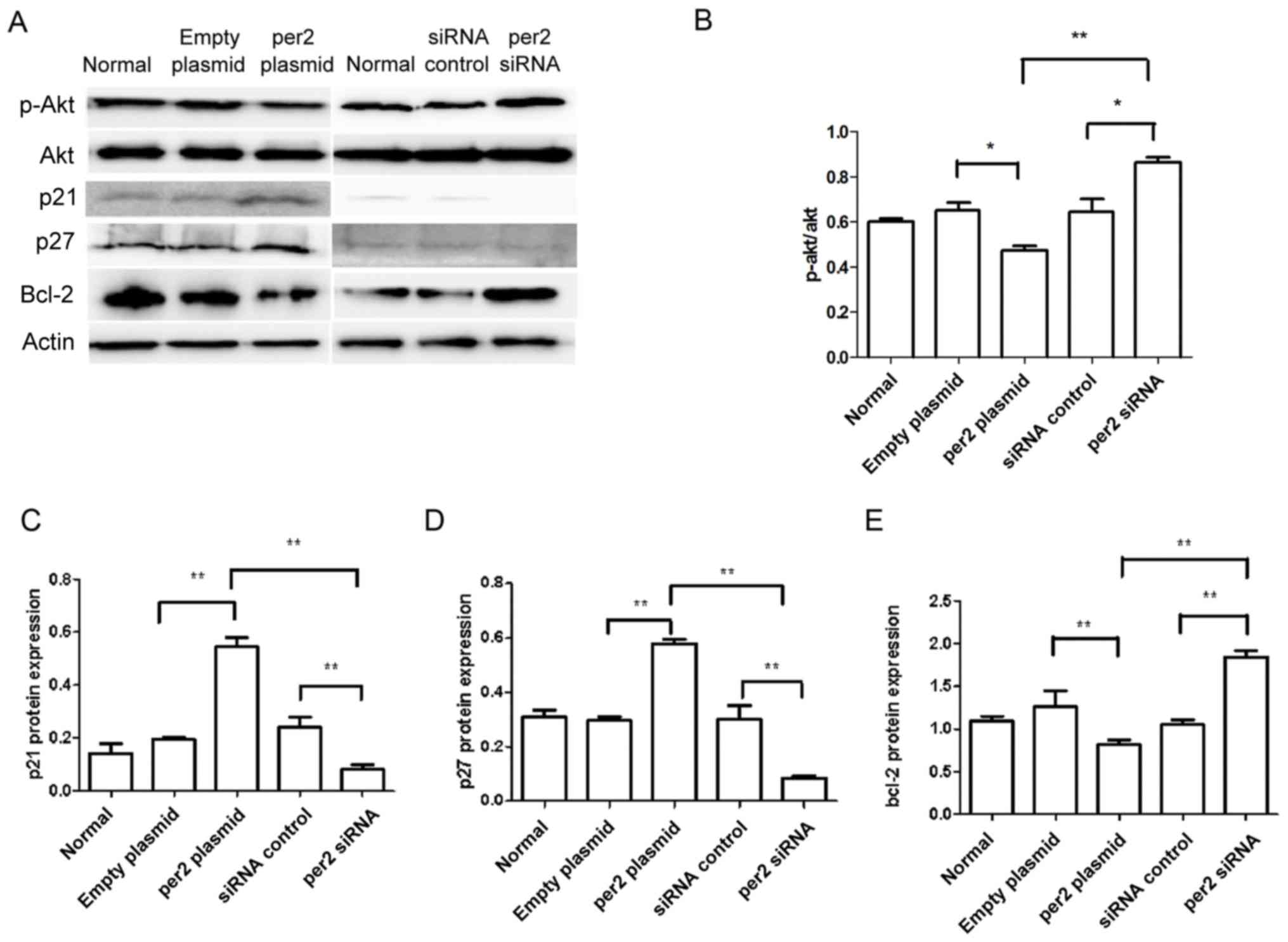
Affects of the alteration of per2 on
the expression of proliferation and apoptosis-associated
proteins
The activation of Akt in tumor cells enhances cell
migration and growth (24). Western
blot analysis was used to detect proteins from the Akt signaling
pathway, and other proteins associated with proliferation and
apoptosis. As demonstrated in Fig. 4,
decreased protein levels of p-Ser473 Akt were observed in MNNG/HOS
cells with per2 overexpression, whereas per2 siRNA increased the
p-Ser473 Akt level (Fig. 4A and B).
Per2 overexpression also elevated the levels of p27 and p21 protein
expression. Conversely, per2 knockdown decreased the p27 and p21
levels (Fig. 4A, C and D). In
addition, the overexpression of per2 reduced the protein expression
of Bcl-2, whereas the suppression of per2 increased the expression
of Bcl-2 (Fig. 4A and E). Per2
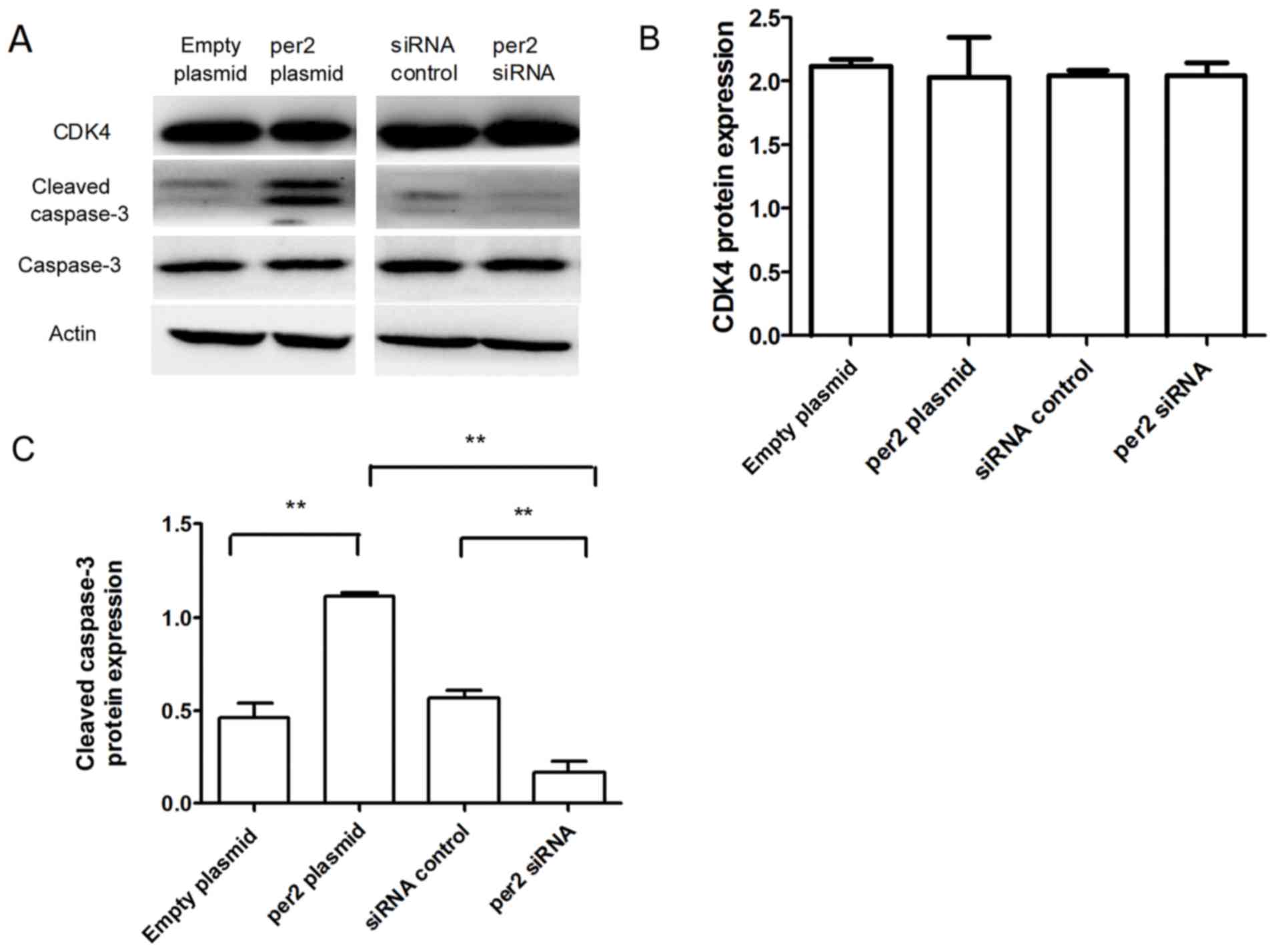
overexpression increased cleaved caspase-3 protein expression,
whereas per2 siRNA decreased cleaved caspase-3 protein expression
(Fig. 5A and C). There was no
alteration of CDK4 expression in per2 overexpression and siRNA
cells (Fig. 5A and B). There were no
evident differences in expression among the control groups.
Discussion
Osteosarcoma continues to present a threat to human
health and survival. Proliferation and metastasis are critical
events in the pathogenesis of cancer (25). Oncogenes and tumor suppressor genes
are associated with cancer cell proliferation and differentiation;
therefore, it is necessary to identify genes which promote
osteosarcoma cell growth, survival and metastasis.
Previous studies have demonstrated that the aberrant
expression or rhythm of circadian clock genes is associated with
carcinogenesis and cancer progression (26–28). The
core clock genes include per1, 2 and 3, CLOCK, cryptochrome1
(CRY1), cryptochrome2 (CRY2), Bmal1, casein kinase1 epsilon
(CSNK1ε), timeless (TIM) and timeless-interacting protein (Tipin)
(23). Among the clock genes, per2
has been demonstrated to serve an important role in cancer
progression, and its altered expression has been identified in a
number of types of cancer (20–23). The
overexpression of per2 has been demonstrated to suppress tumor
growth in vivo (29) and to
induce apoptosis in cancer cells (30). Additionally, mice deficient in per2
exhibited a higher incidence of cancer development (31). It was previously reported that per2
expression level is associated with patient age, tumor histological
grade, invasion depth, lymph node metastasis and
tumor-node-metastasis stage (28). It
was also demonstrated that a reduced expression of per2 accelerated
tumor growth in vitro (32).
Furthermore, Koyanagi et al (33) reported that per2 inhibited tumor
angiogenesis through inhibiting vascular endothelial growth factor
agonist activity.
In the present study, a per2 plasmid and anti-per2
siRNA were constructed, and transfected into MNNG/HOS cells. Per2
expression was detected by western blotting and immunofluorescence.
MNNG/HOS cell proliferation, migration and apoptosis were detected
by CCK-8, Transwell and TUNEL assays. Per2 overexpression inhibited
MNNG/HOS cell proliferation and migration, while apoptosis was
upregulated. In cells with per2 siRNA, the effect was the
opposite.
The results of the present study suggest that the
overexpression of per2 may alter MNNG/HOS cell function through the
Akt cascade, as per2 overexpression inhibited Akt phosphorylation.
The Akt signaling pathway is a downstream signal transduction
pathway involving numerous growth factors and cytokines. The Akt
signaling pathway inhibits cell apoptosis, promotes cell
proliferation and is associated with tumor occurrence, while also
affecting the migration and invasion of tumor cells and promoting
tumor progression. The associated PI3K/PKB signaling pathway also
affects glucose metabolism and promotes the growth of tumor cells
(34). It was previously demonstrated
that cells with downregulated per2 expression have sustained high
levels of Akt phosphorylation after growth factor stimulation or
DNA damage (35). A study by Chen
et al (17) also demonstrated
that per2 knockdown increased the activity of the PI3K/AKT/mTOR
signaling pathway, and that the overexpression of per2 reduced
growth and promoted apoptosis in A549 cells.
The Bcl-2 family of proteins is a key regulator of
the mitochondrial response to apoptotic signals. The Bcl-2 gene
family comprises different members that regulate apoptosis either
positively or negatively. In the present study, the expression of
Bcl-2 was investigated. As shown in Fig.
4, there was a decrease in the Bcl-2 protein levels in
per2-overexpressing cells compared with control cells.
It was also identified that per2 over-expression
inhibited p21/p27 protein expression. As key regulators in cell
proliferation, p21 and p27 serve important roles in DNA damage
repair, cell differentiation and senescence (36). Per2 may inhibit MNNG/HOS cell
proliferation through inhibiting p21/p27 expression.
Cyclin-dependent kinases (CDKs) are oncogenes in a
range of cancer types (37).
Components of the CDK pathway are deregulated in the majority of
human tumors (38). In the present
study, there was no alteration of CDK4 expression. The expression
of the apoptosis-associated protein cleaved caspase-3 was also
detected. Per2 overexpression may have increased cleaved caspase-3
expression to induce MNNG/HOS cell apoptosis.
In the present study, the rates of MNNG/HOS cell
proliferation and apoptosis at different per2 expression levels
were observed, confirming that the per2 level serves an important
role in osteosarcoma cell function. In a previous study (34), another group reported that the
overexpression of per2 increased apoptosis and decreased
proliferation in MG-63 cells. However, they did not further explore
the influence of a lower expression level of per2 on MG-63
function, and did not explore the p53-dependent apoptosis signaling
pathway.
In summary, the present study demonstrated that the
overexpression of per2 resulted in reduced proliferation and
increased apoptosis in MNNG/HOS osteosarcoma cells. Overexpressed
per2 altered the expression of apoptosis-associated proteins that
were involved directly or indirectly in apoptosis; however, the
exact mechanisms for the effects of per2 require further
elucidation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81600344, 81500339,
81302939 and 81402218), Shandong Provincial Natural Science
Foundation, China (grant nos. ZR2016HQ29 and ZR2016HQ01), and
Shandong Province Outstanding Young Scientists Research Award Fund
(grant no. BS2015YY026).
Availability of data and materials
The data sets generated and analyzed during the
study are available from the corresponding author, on reasonable
request.
Authors' contributions
All authors read and approved the manuscript. YYS
made substantial contributions to the conception and design. TQ
made substantial contributions to the acquisition of data. XTL,
YGL, YL, WY and NL made substantial contributions to the analysis
and interpretation of data. WY was involved in drafting the
manuscript and revising it critically for important intellectual
content. YYS gave final approval of the version to be
published.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Whelan J, McTiernan A, Cooper N, Wong YK,
Francis M, Vernon S and Strauss SJ: Incidence and survival of
malignant bone sarcomas in England 1979–2007. Int J Cancer.
131:E508–E517. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Admassi D: Osteosarcoma of medial cuniform
bone. Ethiop Med J. 47:305–308. 2009.PubMed/NCBI
|
|
4
|
Heaton TE, Hammond WJ, Farber BA, Pallos
V, Meyers PA, Chou AJ, Price AP and LaQuaglia MP: A 20-year
retrospective analysis of CT-based pre-operative identification of
pulmonary metastases in patients with osteosarcoma: A single-center
review. J Pediatr Surg. 52:115–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gelderblom H, Jinks RC, Sydes M, Bramwell
VH, van Glabbeke M, Grimer RJ, Hogendoorn PC, McTiernan A, Lewis
IJ, Nooij MA, et al: Survival after recurrent osteosarcoma: Data
from 3 European Osteosarcoma Intergroup (EOI) randomized controlled
trials. Eur J Cancer. 47:895–902. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martin JW, Squire JA and Zielenska M: The
genetics of osteosarcoma. Sarcoma. 2012:6272542012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang J and Zhang W: Newmolecular insights
into osteosarcoma targeted therapy. Curr Opin Oncol. 25:398–406.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gubin DG, Weinert D, Rybina SV, Danilova
LA, Solovieva SV, Durov AM, Prokopiev NY and Ushakov PA: Activity,
sleep and ambient light have a different impact on circadian blood
pressure, heart rate and body temperature rhythms. Chronobiol Int.
34:632–649. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bueno C and Menna-Barreto L: Development
of sleep/wake, activity and temperature rhythms in newborns
maintained in a neonatal intensive care unit and the impact of
feeding schedules. Infant Behav Dev. 44:21–28. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kelleher FC, Rao A and Maguire A:
Circadian molecular clocks and cancer. Cancer Lett. 342:9–18. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Partch CL, Green CB and Takahashi JS:
Molecular architecture of the mammalian circadian clock. Trends
Cell Biol. 24:90–99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Greene MW: Circadian rhythms and tumor
growth. Cancer Lett. 318:115–123. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kelly-Hayes M, Wolf PA, Kase CS, Brand FN,
McGuirk JM and D'Agostino RB: Temporal patterns of stroke onset.
The Framingham Study. Stroke. 26:1343–1347. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muller JE, Stone PH, Turi ZG, Rutherford
JD, Czeisler CA, Parker C, Poole WK, Passamani E, Roberts R,
Robertson T, et al: Circadian variation in the frequency of onset
of acute myocardial infarction. N Engl J Med. 313:1315–1322. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hansen AB, Stayner L, Hansen J and
Andersen ZJ: Night shift work and incidence of diabetes in the
Danish Nurse Cohort. Occup Environ Med. 73:262–268. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jensen LD, Cao Z, Nakamura M, Yang Y,
Bräutigam L, Andersson P, Zhang Y, Wahlberg E, Länne T, Hosaka K
and Cao Y: Opposing effects of circadian clock genes bmal1 and
period2 in regulation of VEGF-dependent angiogenesis in developing
zebrafish. Cell Rep. 2:231–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen B, Tan Y, Liang Y, Li Y, Chen L, Wu
S, Xu W, Wang Y, Zhao W and Wu J: Per2 participates in AKT-mediated
drug resistance in A549/DDP lung adenocarcinoma cells. Oncol Lett.
13:423–428. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Storch KF, Lipan O, Leykin I, Viswanathan
N, Davis FC, Wong WH and Weitz CJ: Extensive and divergent
circadian gene expression in liver and heart. Nature. 417:78–83.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ripperger JA and Albrecht U: The circadian
clock component PERIOD2: From molecular to cerebral functions. Prog
Brain Res. 199:233–245. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen ST, Choo KB, Hou MF, Yeh KT, Kuo SJ
and Chang JG: Deregulated expression of the PER1, PER2 and PER3
genes in breast cancers. Carcinogenesis. 26:1241–1246. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin YM, Chang JH, Yeh KT, Yang MY, Liu TC,
Lin SF, Su WW and Chang JG: Disturbance of circadian gene
expression in hepatocellular carcinoma. Mol Carcinog. 47:925–933.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Štorcelová M, Vicián M, Reis R, Zeman M
and Herichová I: Expression of cell cycle regulatory factors hus1,
gadd45a, rb1, cdkn2a and mre11a correlates with expression of clock
gene per2 in human colorectal carcinoma tissue. Mol Biol Rep.
40:6351–6361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Relles D, Sendecki J, Chipitsyna G, Hyslop
T, Yeo CJ and Arafat HA: Circadian gene expression and
clinicopathologic correlates in pancreatic cancer. J Gastrointest
Surg. 17:443–450. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neiva KG, Zhang Z, Miyazawa M, Warner KA,
Karl E and Nör JE: Cross talk initiated by endothelial cells
enhances migration and inhibits anoikis of squamous cell carcinoma
cells through STAT3/Akt/ERK signaling. Neoplasia. 11:583–593. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sgourakis G, Gockel I, Lyros O, Hansen T,
Mildenberger P and Lang H: Detection of lymph node metastases in
esophageal cancer. Expert Rev Anticancer Ther. 11:601–612. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mazzoccoli G, Panza A, Valvano MR, Palumbo
O, Carella M, Pazienza V, Biscaglia G, Tavano F, Di Sebastiano P,
Andriulli A and Piepoli A: Clock gene expression levels and
relationship with clinical and pathological features in colorectal
cancer patients. Chronobiol Int. 28:841–851. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gu X, Xing L, Shi G, Liu Z, Wang X, Qu Z,
Wu X, Dong Z, Gao X, Liu G, et al: The circadian mutation
PER2(S662G) is linked to cell cycle progression and tumorigenesis.
Cell Death Differ. 19:397–405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Karantanos T, Theodoropoulos G, Pektasides
D and Gazouli M: Clock genes: Their role in colorectal cancer.
World J Gastroenterol. 20:1986–1992. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miyazaki K, Wakabayashi M, Hara Y and
Ishida N: Tumor growth suppression in vivo by overexpression of the
circadian component, PER2. Genes Cells. 15:351–358. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hua H, Wang Y, Wan C, Liu Y, Zhu B, Yang
C, Wang X, Wang Z, Cornelissen-Guillaume G and Halberg F: Circadian
gene mPer2 overexpression induces cancer cell apoptosis. Cancer
Sci. 97:589–596. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fu L, Pelicano H, Liu JS, Huang P and Lee
CC: The circadian gene period2 plays an important role in tumor
suppression and DNA damage response in vivo. Cell. 111:41–50. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang X, Wood PA, Oh EY, Du-Quiton J,
Ansell CM and Hrushesky WJ: Down regulation of circadian clock gene
Period 2 accelerates breast cancer growth by altering its daily
growth rhythm. Breast Cancer Res Treat. 117:423–431. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koyanagi S, Kuramoto Y, Nakagawa H,
Aramaki H, Ohdo S, Soeda S and Shimeno H: A molecular mechanism
regulating circadian expression of vascular endothelial growth
factor in tumor cells. Cancer Res. 63:7277–7283. 2003.PubMed/NCBI
|
|
34
|
Elstrom RL, Bauer DE, Buzzai M, Karnauskas
R, Harris MH, Plas DR, Zhuang H, Cinalli RM, Alavi A, Rudin CM and
Thompso CB: Akt stimulates aerobic glycolysis in cancer cells.
Cancer Res. 64:3892–3899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang X, He X, Yang Z and Jabbari E:
Mammalian PER2 regulates AKT activation and DNA damage response.
Biochem Cell Biol. 90:675–682. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pérez-Yépez EA, Saldívar-Cerón HI,
Villamar-Cruz O, Pérez-Plasencia C and Arias-Romero LE: p21
activated kinase 1: Nuclear activity and its role during DNA damage
repair. DNA Repair (Amst). 65:42–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Williams GH and Stoeber K: The cell cycle
and cancer. J Pathol. 226:352–364. 2012. View Article : Google Scholar : PubMed/NCBI
|