Introduction
Radiotherapy is used for the treatment of malignant
brain tumors, including high-grade gliomas, brain metastases,
germinoma and medulloblastoma, and metastatic tumors of other
sites, such as lung cancer (1,2). Patients
who received radiation therapy may develop delayed brain injury at
6 months or several years following therapy. Cognitive deficit is
one of the complications of radiation therapy and mainly manifests
as learning and memory disabilities (3). Previous studies demonstrated that glial
cell injury (3,4), cell aplasia (3,5,6), vascular damage (7) and inflammation (8) may lead to radiation-induced brain
injury. A previous study demonstrated that delayed cognitive
deficits may be associated with apoptosis of hippocampal neurons in
immature brain tissues following radiotherapy (9). However, the molecular mechanisms
underlying the cognitive deficits following brain radiotherapy
remain controversial and the clinical prevention and treatment
approaches remain limited.
Calcium channels serve important functions in the
central nervous system. Activation of calcium channels mediates
signal transductions involved in physiological and pathological
neuronal responses. Neurodegeneration induced by acute spinal cord
injury and brain injury may be associated with oxidative stress
induced by apoptosis and calcium influx (10,11).
Calcium channel blockers exhibit a protective effect against brain
injury and cerebral ischemia. The underlying molecular mechanisms
may involve the inhibition of calcium influx, decreased release of
neurotransmitters and inhibition of apoptosis in neurons (12–14).
In the present study, a rat model of whole brain
radiotherapy (WBRT) was established and calcium ion antagonist
nimodipine was administered to evaluate its effects on delayed
cognitive deficits following brain irradiation.
Materials and methods
Animals
A total of 60 male, 10-week-old Wistar rats, with a
mean body weight of 236±10 g were obtained from the Experimental
Animal Center of Hebei Medical University (Hebei, China). Rats were
housed in pairs in a light and temperature-controlled room
(22–24°C, 45–50% humidity and 12/12 h light/dark cycle) and
received food and water ad libitum. Following anesthesia, rats were
randomly divided into the following three groups: Control (rats did
not undergo WBRT and were intraperitoneally injected with 2.5 ml/kg
of saline daily for 7 consecutive days), WBRT (rats underwent WBRT
and were intraperitoneally injected with 2.5 ml/kg of saline daily
for 7 consecutive days) and nimodipine groups [rats underwent WBRT
and were intraperitoneally injected with 2.5 ml/kg of calcium
antagonist nimodipine (0.5 mg/kg/day; Shandong Xinhua
Pharmaceutical Co., Ltd., Shandong, China) daily following
radiotherapy for 7 consecutive days]. Rats (n=10 in each group)
were sacrificed at 7 days or 3 months following WBRT. On day 7,
fresh hippocampal tissues (n=5 in each group) were used for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis of B-cell lymphoma 2 (Bcl-2)-associated X protein (Bax)
and Bcl-2 or for histological examination. For the 3-month
timepoint, rats underwent the Morris water maze test to evaluate
spatial learning and memory performance prior to sacrifice by
decapitation. At three months, rats were randomly divided into two
groups, one group (n=5) was used in PCR of amyloid precursor
protein (APP), brain-derived neurotrophic factor (BDNF) and
growth-associated protein 43 (GAP-43); the other group (n=5) was
used in histological examination following the Morris water maze
test. During the experimental procedures, the hair loss, skin
damage and body weight gain rate [(measured weight value-basal
weight value)/basal weight value] of rats was evaluated.
The procedures involving animals and their care were
conducted in accordance with the Institutional Animal Care and Use
Committee of Hebei Medical University (Hebei, China). The present
study was approved by the Ethics Research Committee of the Second
Hospital of Hebei Medical University (Hebei, China; approval no.
2015215).
Radiation
Rats were anesthetized with an intraperitoneal
injection of chloral hydrate (350 mg/kg), placed on a prone
position and irradiated using a 6-MeV linear accelerator (Varian
Clinac iX; Varian Medical Systems, Palo Alto, CA, USA). Rats were
exposed to 30 Gy radiation. Rats were irradiated from a vertical
direction at a dose rate of 300 cGy/min and at a source-skin
distance of 100 cm. The size of radiation field was 2.0×1.5 cm
(eyes canthus to rear ears), whilst other parts of the body were
sheltered by lead plates to protect against radiation (15). Rats in the control group received
anesthesia only and did not undergo WBRT.
Cognitive function test
The cognitive function was evaluated using the
Morris water maze test. Rats were placed at different starting
positions in a circular pool (diameter, 120 cm; height, 50 cm)
filled with water (depth, 37 cm; 21–22°C, made opaque by adding
powdered milk to prevent the rats seeing the platform under the
water). Rats were trained to locate a platform (diameter, 9 cm;
height, 36 cm), which was submerged 1 cm below the water surface
and located in a quadrant of the pool, using distant visual cues.
These visual cues were present on the four walls surrounding the
pool at a distance of 0.6 m. For the training phase, each rat
performed 5 acquisition trials/day (maximal swimming time 120 sec;
30 sec on platform; inter-trial interval 30 min) for 4 consecutive
days. A different quadrant was selected each time as a point of
entry. The rat from each of the three group rats used the same
starting position for evaluation. All trials were recorded, and
latency time, defined as the time to locate the platform, was used
as a measure for spatial learning. The rats were allowed 60 sec
until they found the platform. If the rat failed to find the
platform within 60 sec, the rat was placed on the platform and
allowed to stay on it for 20 sec. The time taken for the rat to
reach the platform was recorded. During the probe trial, the hidden
platform was removed from the tank and the rats were allowed to
swim freely for 60 sec. Latency to find the maze platform (sec) was
recorded. All the trials were recorded and analyzed by an automated
video-tracking system (JLBehv-M; version 2.1; Shanghai Jiliang
Software Technology Co., Ltd., Xuhui, Shanghai, China).
Histological examination
For the histological examination, the rats were
perfused with saline, followed by fixation with 4%
paraformaldehyde. The brain tissues (thickness, 1–4 mm behind the
optic chiasma) were removed and stored in the same paraformaldehyde
solution for 24 h and embedded in paraffin. Paraffin-embedded
tissue samples were cut into 5-µm thick sections. A total of five
nonconsecutive sections were used for histological examination.
Slices were incubated at 60°C for 30 min, then cooled, cleared with
xylene twice for 5 min each, dehydrated with 100, 95, 90 and 85%
ethanol each for 5 min and then with 80 and 70% ethanol each for 3
min. Furthermore, slices were rinsed with distilled water for 1
min, stained with 0.1% cresyl violet dye in an oven at 37°C for 30
min, rinsed with water for 8 min and rapidly separated with 95%
ethanol. Following this, sections were incubated in anhydrous
alcohol and xylene twice for 5 min each prior to mounting. The
slices were observed and imaged with a Nikon ECLIPSE 50i light
microscope at magnification, ×400 (Nikon Corporation, Tokyo,
Japan). The neuronal density was determined by the average number
of surviving hippocampal Cornu Ammonis (CA)1 in each 1-mm section,
with three sections of bilateral hippocampal slices (16).
Detection of apoptosis of neurons in the dentate
gyrus of the hippocampus was evaluated using immunohistochemical
staining for caspase-3. Prior to staining, the sections were
incubated for 30 min at 65°C and deparaffinized with xylene and
graded series of ethanol (100, 95, 90 and 85% ethanol each for 5
min and then with 80 and 70% ethanol each for 3 min). Antigen
retrieval was performed by boiling the sections in 10 mM sodium
citrate buffer (AM9763; pH 6.0; Thermo Fisher Scientific Inc.,
Waltham, MA, USA) for 10 min at room temperature. The sections were
pre-treated with 10 µg/ml proteinase K (SC-473603; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 10 min at room
temperature prior to incubation with 4% normal goat serum (cat. no.
ab7481; Abcam, Cambridge, USA) at room temperature for 1 h. The
primary antibody against active caspase-3 (1:100; BS1518; Bioworld
Technology, Inc., St. Louis Park, MN, USA) was incubated in a
humidified box at 4°C overnight, followed by an 1 h incubation at
room temperature with a biotinylated secondary goat anti-rabbit
antibody (1:400; SC-2040; Santa Cruz Biotechnology, Inc.).
3,3′-diaminobenzidine (D12384; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) to visualize the antigen; following this, the
sections were counterstained with Hematoxylin (H9627;
Sigma-Aldrich; Merck KGaA) 5 min at room temperature, dehydrated in
ethanol (80, 95, 95, 100 and 100%, each for 3 min) and cleared in
xylene. The sections were mounted and the caspase-3-positive cells
were observed and counted under a Nikon ECLIPSE 50i light
microscope at magnification, ×400.
RT-qPCR
Hippocampal tissues from the rats were removed at
different time points, which included seven days and three months
following WBRT. To extract total RNA, the frozen hippocampi were
homogenized in 1.0 ml TRIzol® (cat. no. 15596-026;
Thermo Fisher Scientific Inc.). Following adding 200 µl chloroform
and placing on ice for 5 min, homogenates were centrifuged (12,000
× g for 15 min at 4°C) and the supernatant was precipitated with
1.0 ml isopropanol, then placed on ice for 10 min and centrifuged
(12,000 × g for 10 min at 4°C). The supernatant fluid was removed,
400 µl 75% ethanol was added and then the tube was centrifuged
(12,000 × g for 5 min at 4°C). Following this the supernatant fluid
was removed and dried on filter paper. The powder was dissolved by
0.1% diethylpyrocarbonate. The RNA was reverse transcribed to cDNA
using a RevertAid First Strand cDNA synthesis kit (K1622; Thermo
Fisher Scientific Inc.), following the manufacturer's protocol.
RT-qPCR reactions were performed using Platinum SYBR-Green qPCR
SuperMix-UDG (CS11733038; Invitrogen; Thermo Fisher Scientific,
Inc.). For the rats in the seven days group, the following primer
pairs were used: Bax forward, GGCGAATTGGCGATGAACTG and reverse,
ATGGTTCTGATCAGCTCGGG; and Bcl-2 forward, GGGGCTACGAGTGGGATACT and
reverse, GACGGTAGCGACGAGAGAAG. The level of expression of these two
tested genes was normalized to the level of expression of the
β-actin gene forward, GGAGATTACTGCCCTGGCTCCTA and reverse,
GACTCATCGTACTCCTGCTTGCTG. For the rats in the three months group,
the following primer pairs were used: APP forward,
TCGTGATTCCTTACCGGTGC and reverse, ATCGATGCTGTCGCTCTCCT; BDNF
forward, GGTCACAGTCCTGGAGAAAG and reverse, GTCTATCCTTATGAACCGCC;
and GAP-43 forward, GCTAGCTTCCGTGGACACAT and reverse,
ACCATCAGCAACGGGAGCAT. GAPDH forward, AGGTGAAGGTCGGAGTCAACG and
reverse, AGGGGTCATTGATGGCAACA was used as the internal control. PCR
reactions were performed with initial denaturation at 95°C for 5
min, followed by 40 cycles at 95°C for 30 sec, 58°C (for Bax), 56°C
(for Bcl-2 and BDNF), 59°C (for GAP-43) and 57°C (for APP and
GAPDH) for 30 sec and 72°C for 45 sec. Real-time PCR was performed
using an Mx3000P Real Time PCR system (Agilent, Technologies, Inc.,
Santa Clara, CA, USA). The quantification cycle (Cq) of target
genes was normalized to that of internal control. mRNA levels in
WBRT or nimodipine group animals were calculated after normalizing
the quantification cycle to internal control expression, and the
2−ΔΔCq method was used to calculate the degree of
expression (17).
Statistical analysis
Data were analyzed using SPSS software (version
20.0; IBM Corp., Armonk, NY, USA). Data are expressed as the mean ±
standard deviation. One-way analysis of variance was performed
followed by the least significant difference test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Evaluation of body weight following
WBRT
A total of 60 rats were divided into three groups:
control, WBRT and nimodipine groups and the body weight gain was
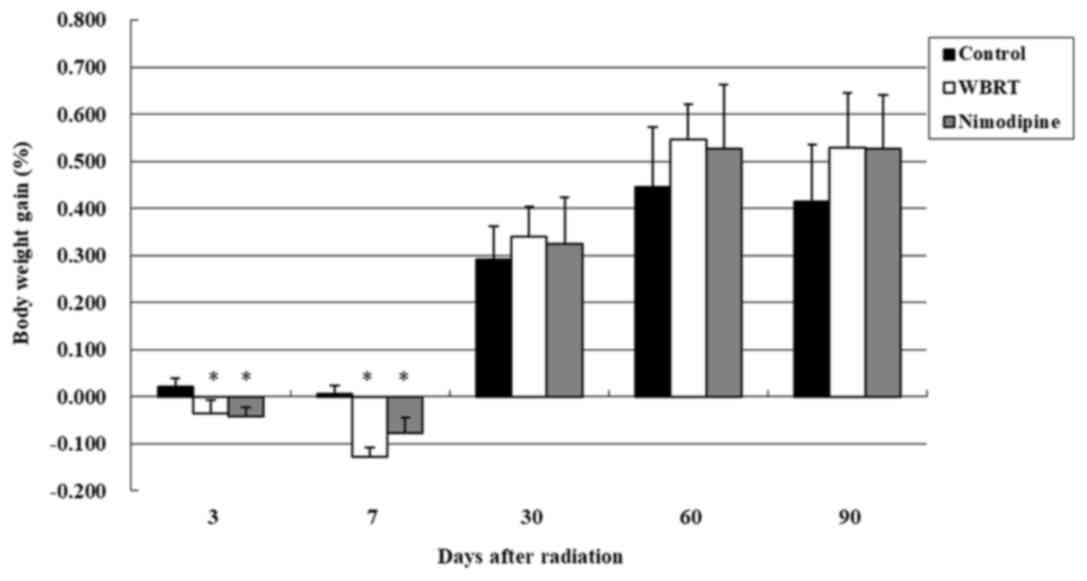
evaluated on day 3, 7 30, 60 and 90 following irradiation (Fig. 1). On day 3, body weight gain rates
were 0.02±0.02, −0.04±0.03 and −0.04±0.02 in the control, WBRT and
nimodipine groups, respectively. One-way ANOVA revealed that the
differences in the body weight gain were statistically significant
(P=0.001) among the three groups. The results demonstrated WBRT and
nimodipine groups lost weight, compared with control groups, three
days after brain radiation and the differences were statistically
significant (P=0.001). However, no differences in the body weight
gain were observed between the nimodipine and WBRT groups on day 3
(P=0.649). There remained a significant difference in body weight
gain between rats in the nimodipine (P=0.007) or WBRT (P<0.001)
groups, compared with the control group, following seven days;
however, no differences in the body weight gain were observed among
the three groups following 1, 2 and 3 months of radiotherapy
(Fig. 1).
Treatment with nimodipine improves
cognitive function following WBRT
During irradiation, the physiological appearance and
behavior of rats was monitored. Rats of WBRT and nimodipine groups
exhibited hair loss and skin damage one week after brain
radiation.
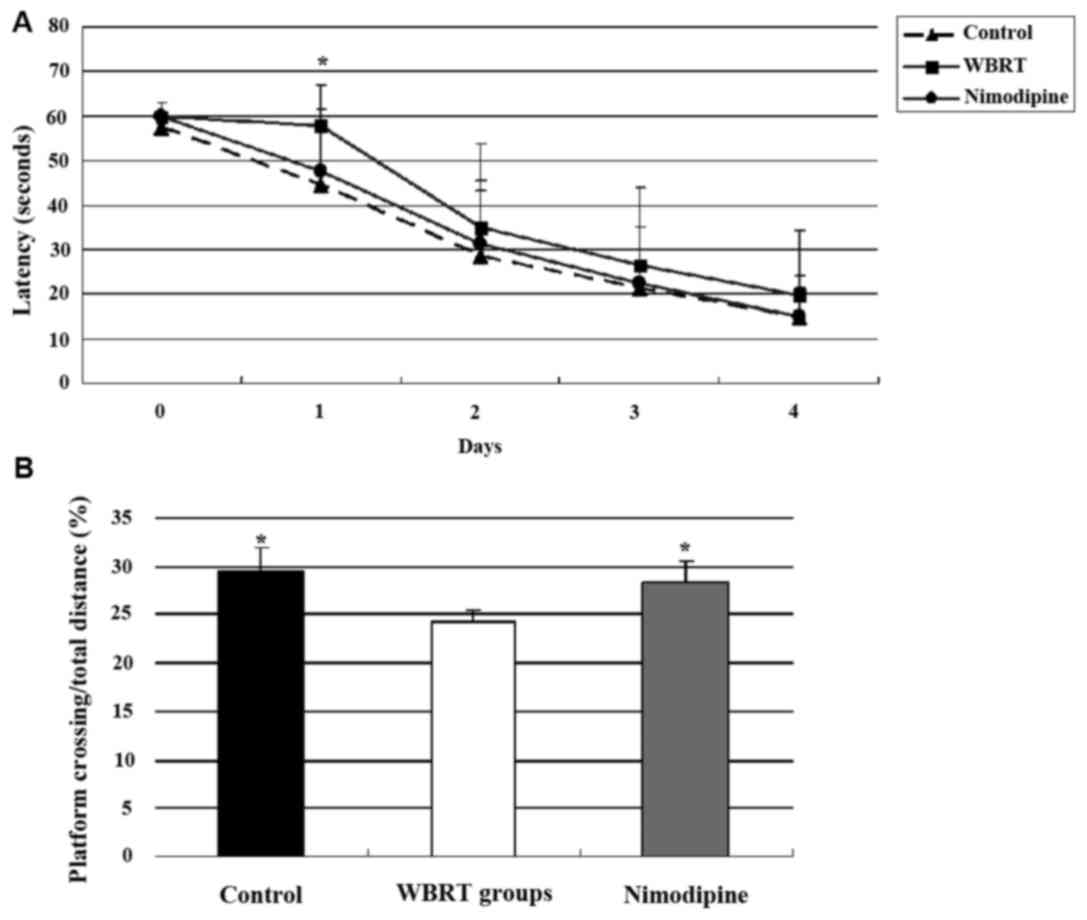
The cognitive function of rats was evaluated using a
Morris water maze test three months after brain radiation. The
results of the pre-training (Day 0) did not differ among the three
groups (P=0.397). On day 1, there was a significant difference in
latency to the platform among the three groups (P<0.05). Rats
from the control (P=0.003) and nimodipine groups (P=0.019) have an
improved performance, compared with the WBRT group. On days 2, 3
and 4, no significant differences in latency to the platform were
observed among the three groups (P>0.05) (Fig. 2A). The average latency was
28.80±18.13, 35.76±20.94 and 30.48±17.83 for control, WBRT and
nimodipine groups, respectively.
During the probe trial, the ratio of platform
crossing/total distance in the WBRT (24.33±1.17%) group was
smaller, compared with the control (29.63±2.25%; P=0.001) and
nimodipine (28.24±2.36%; P=0.009) groups. The data revealed that
the differences in platform crossing times among the three groups
were statistically significant (P=0.003). These results
demonstrated that the cognitive function of rats in the WBRT group
was poorer compared with that in the control and treatment with
nimodipine improved the cognitive function of rats (Fig. 2B).
Decreased numbers of neurons following
WBRT
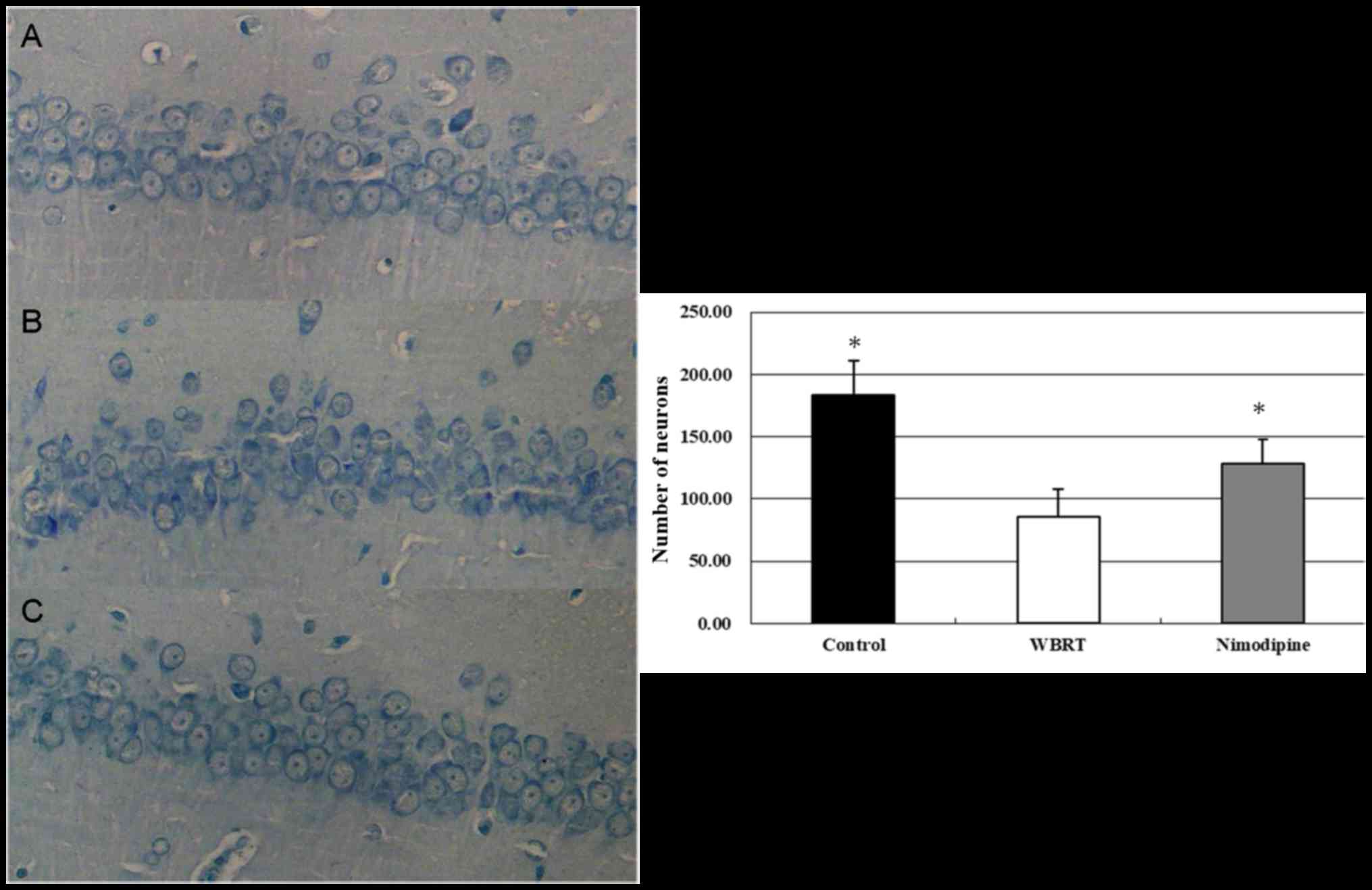
Neuronal survival was assessed using Nissl staining.
The results revealed that the neurons in the CA1 region of the
hippocampus exhibited intact morphology in the control group
(Fig. 3A). The neurons in the WBRT
group appeared to be decreased, exhibited a ‘fuzzy’ appearance and
degeneration (Fig. 3B). The numbers
of neurons was decreased in the CA1 region in the WBRT group
(85.3±22.9/mm2) compared with that in the control group
(183.2±27.6/mm2). The number of neurons in CA1 region
was increased in the nimodipine group (Fig. 3C; 128±19.5/mm2) compared
with that in the WBRT group. The differences in the numbers of
neurons among these three groups were statistically significant
(P<0.01; Fig. 3). These results
suggest that the morphology and survival number of neurons in the
hippocampus were affected following WBRT (Fig. 3).
Cell apoptosis is increased following
WBRT
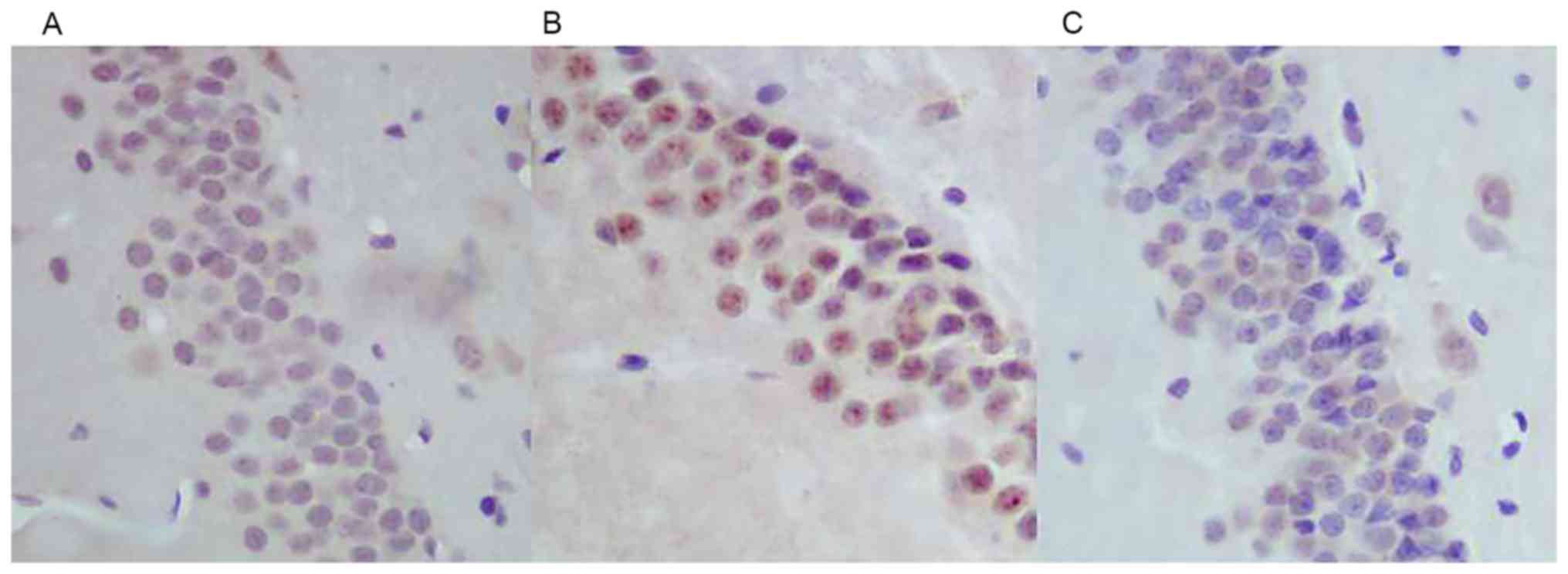
Immunohistochemical staining of caspase-3 (Fig. 4), revealed that the expression of
caspase-3 was increased in the neurons in the hippocampal dentate
gyrus area in the WBRT group (Fig.
4B) compared with that in the control group (Fig. 4A). However, the expression of
caspase-3 decreased in the hippocampal dentate gyrus area in the
nimodipine group (Fig. 4C) compared
with that in the WBRT group (Fig.
4B).
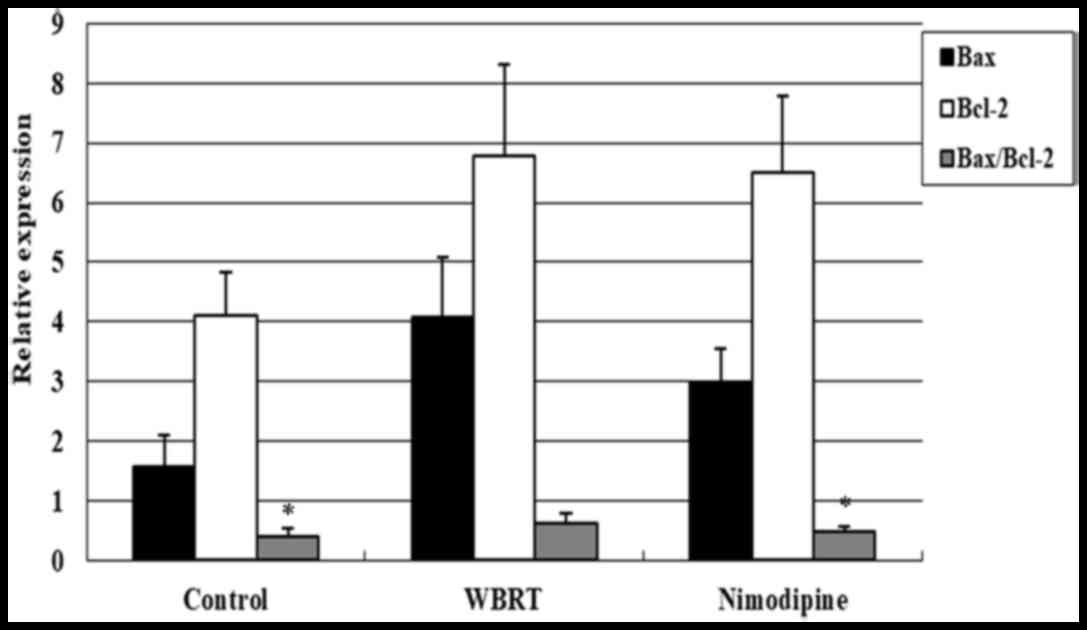
RT-qPCR analysis was performed to assess the
Bax/Bcl-2 ratio among the three groups. The differences in the
Bax/Bcl-2 ratio among these three groups were statistically
significant (P=0.002). The results demonstrated that the Bax/Bcl-2
ratio was upregulated in the WBRT group (0.62±0.16), compared with
that in the control group (0.39±0.14; P=0.001; Fig. 5), suggesting that cell apoptosis may
increase in response to irradiation. Additionally, the Bax/Bcl-2
ratio in the nimodipine group (0.47±0.10, P=0.192) was marginally
increased, compared with control group but significantly decreased,
compared with the WBRT group (P=0.016; Fig. 5).
Expression levels of BDNF, APP and
GAP-43 following WBRT
The expression of genes associated with cognitive
function, including BDNF, GAP-43 and APP (18–21) was
assessed using RT-qPCR. The results demonstrated that the
expression levels of BDNF (5.13±0.46) and GAP-43 (4.57±0.35) were
decreased in the hippocampus of the WBRT group, compared with those
in the control group [6.93±0.61 (P=0.002); 5.89±1.20 (P=0.024),
respectively]. However, the expression levels of BDNF increased in
the nimodipine group (7.18±1.00) compared with that in the WBRT
group (P<0.01; Fig. 6). The
expression levels of GAP-43 between the nimodipine (5.05±0.63) and
WBRT group were not statistically significant (P=0.360).
Additionally, the expression levels of APP (4.74±1.43) were
significantly increased in the WBRT group compared with that in the
control group (3.02±0.30; P=0.016). However, nimodipine treatment
did not affect significantly the expression levels of APP
(4.25±0.82) compared with that in the WBRT group (P=0.441; Fig. 6).
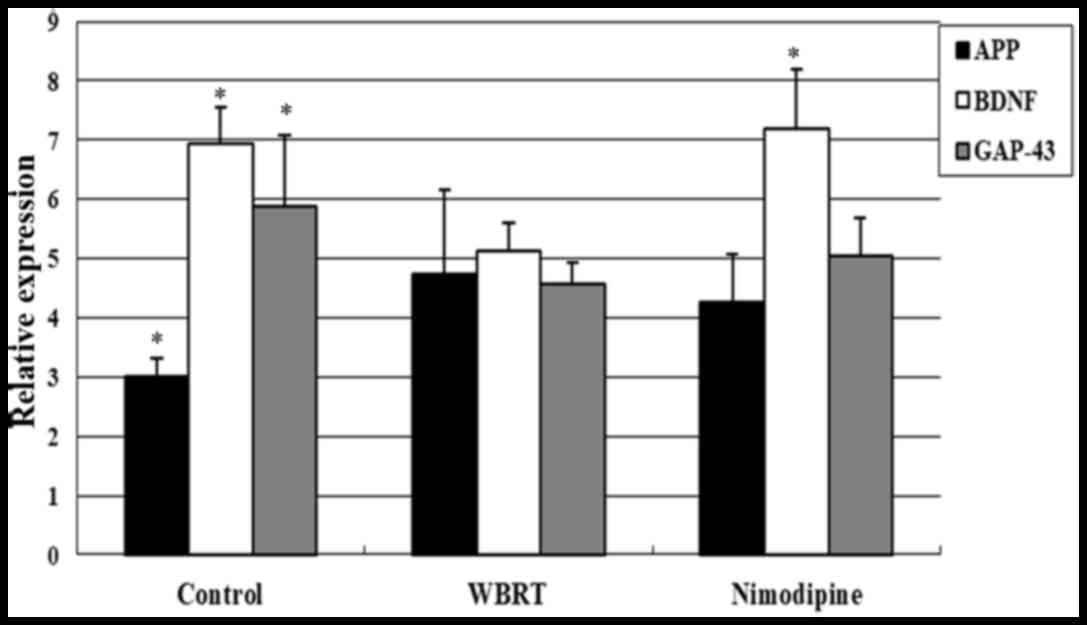 | Figure 6.Expression levels of APP, BDNF and
GAP-43 following WBRT. Expression levels of BDNF (P=0.002) and
GAP-43 (P=0.024) decreased in the hippocampus of rats in the WBRT
group compared with that in the control group. The expression level
of BDNF increased in the nimodipine group, compared with the WBRT
group (P<0.01). Expression of APP was upregulated in the WBRT
group, compared with that in the control group; however, nimodipine
treatment did not significantly downregulate APP expression,
compared with the WBRT group (P=0.441). WBRT, whole brain
radiotherapy; APP, amyloid precursor protein; BDNF, brain-derived
neurotrophic factor; GAP-43, growth-associated protein 43.
*P<0.05, vs. WBRT. |
Discussion
The improvement of the comprehensive treatment of
tumors prolonged the survival time of patients with malignant brain
tumors. Cognitive defects caused by radiotherapy have attracted
increased attention (22,23). In the present study, a rat model of
WBRT was established. Additionally, the molecular mechanism
underlying delayed cognitive deficits and the effects of calcium
antagonists following radiotherapy were investigated.
The results of the present study demonstrated that
rats developed learning and memory dysfunction at 3 months
following a single 30 Gy dose of WBRT. This model may be used to
observe delayed brain injury. Previous studies investigated
strategies to optimize animal models for radiotherapy. Shi et
al (24) used fractionated
radiotherapy with a total of 45 Gy delivered as nine fractions of 5
Gy twice per week for 4.5 weeks and observed for 12 months.
Compared to sham-irradiated rats, the irradiated rats demonstrated
impaired MWM performance. Semmler et al (25) suggested a model whereby rats were
irradiated for four consecutive days with a dose of 5 or 10 Gy and
were observed ≤6 weeks following irradiation. After two weeks
following irradiation, the rats demonstrated decreased open-field
activity but no cognitive deficit as indicated by latencies in the
Morris water maze test. After six weeks following irradiation, no
group demonstrated alterations in histopathology, such as vascular
changes, demyelination or white matter necrosis. An increased
cumulative dosage and dose per fraction was used in this model to
achieve a higher degree of radiation-induced toxicity. In the
present study, rats exhibited learning and memory deficits at 3
months following a single 30 Gy dose of WBRT. Nissl staining
demonstrated that the numbers of neurons in the CA1 region of the
hippocampus decreased and the morphology of the cells changed
following WBRT. These results confirm the efficient establishment
of the delayed brain injury model following WBRT. The animal model
employed in the present study did not require repeated anesthesia
procedures due to a single radiation course and the success rate of
delayed brain injury was increased, compared with animal models
established in previous studies.
Increased apoptosis of neurons in the hippocampus
may be a possible molecular mechanism underlying the impairment in
learning and memory following WBRT (26,27).
Apoptosis serves a key function in several types of brain injury
including ischemic cerebral infarction, traumatic brain injury,
neonatal hypoxic encephalopathy and toxic encephalopathy. Previous
studies have also revealed that the apoptosis of neurons may occur
following brain radiotherapy (28–30). The
present study revealed that the numbers of neurons in the CA1
region of the hippocampus decreased and the morphology of the cells
changed following WBRT. Furthermore, the expression levels of
caspase-3 were upregulated in the dentate gyrus of the hippocampus
following WBRT, suggesting that apoptosis of hippocampal neurons
was increased following WBRT.
According to previous studies, hippocampus may be
associated with the learning and memory function of the brain, and
with spatial memory (31,32). Complete or partial hippocampal damage
may lead to spatial learning and memory impairment (33). A previous study suggested that
increased neurogenesis in the hippocampal dentate gyrus induced by
electrical stimulation of the anterior thalamus may improve memory
function (34). According to these
results, it was hypothesized that the increase in apoptosis and the
decrease in the number of neurons in the hippocampus following WBRT
may lead to learning and memory dysfunction.
Administration of a calcium antagonist nimodipine
may partly inhibit the apoptosis of neurons in the hippocampus and
improve cognitive function following WBRT. Previous studies on
cerebral ischemia, brain injury, chronic alcoholic encephalopathy,
subarachnoid hemorrhage or cerebral hemorrhage demonstrated that
calcium antagonists may exert a protective effect on brain function
(12,35–38). The
expression levels of calcium-associated genes decreased following
cerebral ischemia, leading to inhibition of L-type calcium
channels, which in turn may induce post-ischemic neuronal death.
Calcium antagonists may increase the survival of neurons in the
hippocampal CA1 region and improve learning and memory function
following cerebral ischemia (14,39). The
results of the Morris water maze test demonstrated that rats in the
nimodipine group spent less time finding the hidden platform and
had increased platform crossing, compared with the WBRT group.
These results suggest that nimodipine may improve the learning and
memory function of rats following brain radiotherapy. Nissl
staining and immunohistochemistry analysis revealed that nimodipine
may increase the number of neurons in the CA1 region and partially
downregulated the expression of caspase-3 in the dentate gyrus
region of the hippocampus of rats following radiotherapy. These
results suggested that the application of nimodipine may partially
inhibit apoptosis of neurons in the hippocampus of rats following
brain radiotherapy.
The molecular mechanism whereby nimodipine may
inhibit the apoptosis of hippocampal neurons following radiotherapy
may involve the regulation of BDNF and Bax/Bcl-2 ratio. BDNF and
Bcl-2 are associated with the survival of neurons (40–42).
Neuronal apoptosis is correlated with the interactions between
apoptosis promoting factors and anti-apoptotic factors. During
apoptosis, Bcl-2 family members may interact with
apoptosis-promoting factors and regulate the release of cytochrome
C from mitochondria, inducing cell death (43). Bax is the main apoptosis-promoting
factor in the nervous system that interacts with the Bcl-2 family
members. Bcl-2-dependent and Bax-dependent apoptosis is associated
with the regulation of caspase-3 in the brain (43,44). A
previous study demonstrated that Bax-deficient mice with brain
injury exhibited increased numbers of neural progenitor cells in
the dentate gyrus and improved remodeling of the hippocampus
(45). In the present study, RT-qPCR
analysis indicated that nimodipine may decrease the ratio of
Bax/Bcl-2 and increase the expression levels of BDNF following
WBRT. These results suggest that the calcium ion antagonist may
downregulate the Bax/Bcl-2 ratio by inhibiting the release of Bax,
upregulating the expression of BDNF, inhibiting apoptosis of
hippocampal neurons following radiotherapy, thus improving
cognitive function.
To conclude, the present study indicated that
delayed cognitive deficits may occur in rats following whole brain
radiation, and the application of calcium ion antagonist may
improve cognitive function following radiotherapy. The molecular
mechanism underlying the effects of nimodipine following WBRT may
involve the inhibition of the apoptosis of neurons in the
hippocampus. Nevertheless, further studies are required to
elucidate the duration and dose of calcium ion antagonist treatment
and provide more reliable and comprehensive data for the study of
cognitive deficits, clinical prevention and treatment.
Acknowledgements
Not applicable.
Funding
The study is supported by a grant of Support Plan
Project sponsored by Hebei Science and Technology Department (grant
no. 122777101) and grant of Project of Science and Technology
activities for returning scientists in Hebei Province (grant no.
C2012003029).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JT, XXZ, BHJ participated in the conception and
design of the study. JL, QSZ, JKY, JWY and XMS conducted the
experiments. LZ, HYL, YZL acquired and analyzed the data. JT and
BHJ drafted and revised the manuscript. All authors read and
approved the final manuscript.
Ethical approval and consent to
participate
The procedures involving animals and their care were
conducted in accordance with the Institutional Animal Care and Use
Committee of Hebei Medical University (Hebei, China). The present
study was approved by the Ethics Research Committee of the Second
Hospital of Hebei Medical University (Hebei, China; approval no.
2015215).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Surma-aho O, Niemelä M, Vilkki J, Kouri M,
Brander A, Salonen O, Paetau A, Kallio M, Pyykkönen J and
Jääskeläinen J: Adverse long-term effects of brain radiotherapy in
adult low-grade glioma patients. Neurology. 56:1285–1290. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blomstrand M, Brodin NP, Munck Af
Rosenschöld P, Vogelius IR, Merino SHnchez G, Kiil-Berthlesen A,
Blomgren K, Lannering B, Bentzen SM and Björk-Eriksson T: Estimated
clinical benefit of protecting neurogenesis in the developing brain
during radiation therapy for pediatric medulloblastoma. Neuro
Oncol. 14:882–889. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Greene-Schloesser D, Moore E and Robbins
ME: Molecular pathways: Radiation-induced cognitive impairment.
Clin Cancer Res. 19:2294–2300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sano K, Morii K, Sato M, Mori H and Tanaka
R: Radiation-induced apoptosis and injury of oligodendrocytes on
neonatal rat brains. Neurol Med Chir (Tokyo). 40:495–500. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Naylor AS, Bull C, Nilsson MK, Zhu C,
Björk-Eriksson T, Eriksson PS, Blomgren K and Kuhn HG: Voluntary
running rescues adult hippocampal neurogenesis after irradiation of
the young mouse brain. Proc Natl Acad Sci USA. 105:14632–14637.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Balentova S and Adamkov M: Molecular,
cellular and functional effects of radiation-induced brain injury:
A review. Int J Mol Sci. 16:27796–27815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brown WR, Blair RM, Moody DM, Thore CR,
Ahmed S, Robbins ME and Wheeler KT: Capillary loss precedes the
cognitive impairment induced by fractionated whole-brain
irradiation: A potential rat model of vascular dementia. J Neurol
Sci. 257:67–71. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jenrow KA, Brown SL, Lapanowski K, Naei H,
Kolozsvary A and Kim JH: Selective inhibition of microglia-mediated
neuroinflammation mitigates radiation-induced cognitive impairment.
Radiat Res. 179:549–556. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huo K, Sun Y, Li H, Du X, Wang X, Karlsson
N, Zhu C and Blomgren K: Lithium reduced neural progenitor
apoptosis in the hippocampus and ameliorated functional deficits
after irradiation to the immature mouse brain. Mol Cell Neurosci.
51:32–42. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vosler PS, Brennan CS and Chen J:
Calpain-mediated signaling mechanisms in neuronal injury and
neurodegeneration. Mol Neurobiol. 38:78–100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nazırog˘lu M, Senol N, Ghazizadeh V and
Yürüker V: Neuroprotection induced by N-acetylcysteine and selenium
against traumatic brain injury-induced apoptosis and calcium entry
in hippocampus of rat. Cell Mol Neurobiol. 34:895–903. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi SK, Lee GJ, Choi S, Kim YJ, Park HK
and Park BJ: Neuroprotective effects by nimodipine treatment in the
experimental global ischemic rat model: Real time estimation of
glutamate. J Korean Neurosurg Soc. 49:1–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bullock R, Zauner A, Woodward J and Young
HF: Massive persistent release of excitatory amino acids following
human occlusive stroke. Stroke. 26:2187–2189. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu HH, Li SJ, Wang P, Yan HC, Cao X, Hou
FQ, Fang YY, Zhu XH and Gao TM: An L-Type Calcium Channel Agonist,
Bay K8644, Extends the Window of intervention against ischemic
neuronal injury. Mol Neurobiol. 47:280–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Y, Sun AM, Chen ZX, Liu Y, Chen LH
and Yuan YW: Ginsenoside Rg1 protects rat hippocampal neurons from
radiation injury by regulating NOS activity. Nan Fang Yi Ke Da Xue
Xue Bao. 30:1522–1555. 2010.(In Chinese). PubMed/NCBI
|
|
16
|
Zhang DS, Liu YL, Zhu DQ, Huang XJ and Luo
CH: Point application with Angong Niuhuang sticker protects
hippocampal and cortical neurons in rats with cerebral ischemia.
Neural Regen Res. 10:286–291. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bekinschtein P, Kent BA, Oomen CA,
Clemenson GD, Gage FH, Saksida LM and Bussey TJ: Brain-derived
neurotrophic factor interacts with adult-born immature cells in the
dentate gyrus during consolidation of overlapping memories.
Hippocampus Aug. 24:905–911. 2014. View Article : Google Scholar
|
|
19
|
Jeon YK and Ha CH: The effect of exercise
intensity on brain derived neurotrophic factor and memory in
adolescents. Environ Health Prev Med. 22:272017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Holahan MR: A shift from a pivotal to
supporting role for the growth-associated protein (GAP-43) in the
coordination of axonal structural and functional plasticity. Front
Cell Neurosci. 11:2662017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Preat T and Goguel V: Role of drosophila
amyloid precursor protein in memory formation. PLoS One.
12:1422016.
|
|
22
|
Ganau L, Paris M, Ligarotti GK and Ganau
M: Management of gliomas: Overview of the latest technological
advancements and related behavioral drawbacks. Behav Neurol.
2015:8626342015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brown PD, Jaeckle K, Ballman KV, Farace E,
Cerhan JH, Anderson SK, Carrero XW, Barker FG II, Deming R, Burri
SH, et al: Effect of radiosurgery alone vs radiosurgery with whole
brain radiation therapy on cognitive function in patients with 1 to
3 brain metastases: A randomized clinical trial. JAMA. 316:401–409.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi L, Adams MM, Long A, Carter CC,
Bennett C, Sonntag WE, Nicolle MM, Robbins M, D'Agostino R and
Brunso-Bechtold JK: Spatial learning and memory deficits after
whole-brain irradiation are associated with changes in NMDA
receptor subunits in the hippocampus. Radiat Res. 166:892–899.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Semmler A, Garbe S, Moskau S, Frisch C,
Eter N, Schlegel U and Linnebank M: An efficient method for
fractionated whole rodent brain radiation. Neurol Res. 35:355–359.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu M, Fan Q, Zhang J, Chen Y, Xu R, Chen
L, Zhao P and Tian Y: NFAT3/c4-mediated excitotoxicity in
hippocampal apoptosis during radiation-induced brain injury. J
Radiat Res. 58:827–833. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li YQ, Cheng Z and Wong S: Differential
apoptosis radiosensitivity of neural progenitors in adult mouse
hippocampus. Int J Mol Sci. 17:E972016.PubMed/NCBI
|
|
28
|
Tada E, Parent JM, Lowenstein DH and Fike
JR: X-irradiation causes a prolonged reduction in cell
proliferation in the dentate gyrus of adult rats. Neuroscience.
99:33–41. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hassan HA, Hafez HS and Goda MS: Mentha
piperita as a pivotal neuro-protective agent against gamma
irradiation induced DNA fragmentation and apoptosis: Mentha extract
as a neuroprotective against gamma irradiation. Cytotechnology.
65:145–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Cheng Z, Wang C, Ma H, Meng W and
Zhao Q: Neuroprotective effects of kukoamine a against
radiation-induced rat brain injury through inhibition of oxidative
stress and neuronal apoptosis. Neurochem Res. 41:2549–2558. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Derdikman D and Moser EI: A manifold of
spatial maps in the brain. Trends Cogn Sci. 14:561–569. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Preston AR and Eichenbaum H: Interplay of
hippocampus and prefrontal cortex in memory. Curr Biol.
23:R764–R773. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Monje ML and Palmer T: Radiation injury
and neurogenesis. Curr Opin Neurol. 16:129–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hamani C, Stone SS, Garten A, Lozano AM
and Winocur G: Memory rescue and enhanced neurogenesis following
electrical stimulation of the anterior thalamus in rats treated
with corticosterone. Exp Neurol. 232:100–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schurr A: Neuroprotection against
ischemic/hypoxic brain damage: Blockers of ionotropic glutamate
receptor and voltage sensitive calcium channels. Curr Drug Targets.
5:603–618. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li H, Yang X, Shi W, Ma Z, Feng G, Wang Q,
Shen L and Xie C: Protective effects of nimodipine on
cerebrovascular function in chronic alcoholic encephalopathy. Int J
Mol Med. 33:201–208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Koskimäki J, Matsui N, Umemori J,
Rantamäki T and Castrén E: Nimodipine activates TrkB neurotrophin
receptors and induces neuroplastic and neuroprotective signaling
events in the mouse hippocampus and prefrontal cortex. Cell Mol
Neurobiol. 35:189–196. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ma B and Zhang J: Nimodipine treatment to
assess a modified mouse model of intracerebral hemorrhage. Brain
Res. 1078:182–188. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li XM, Yang JM, Hu DH, Hou FQ, Zhao M, Zhu
XH, Wang Y, Li JG, Hu P, Chen L, et al: Contribution of
downregulation of L-type calcium currents to delayed neuronal death
in rat hippocampus after global cerebral ischemia and reperfusion.
J Neurosci. 27:5249–5259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Murphy TH, Worley PF and Baraban JM:
L-type voltage-sensitive calcium channels mediate synaptic
activation of immediate early genes. Neuron. 7:625–635. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bito H, Deisseroth K and Tsien RW: CREB
phosphorylation and dephosphorylation: A Ca(2+)- and stimulus
duration-dependent switch for hippocampal gene expression. Cell.
87:1203–1214. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
West AE, Chen WG, Dalva MB, Dolmetsch RE,
Kornhauser JM, Shaywitz AJ, Takasu MA, Tao X and Greenberg ME:
Calcium regulation of neuronal gene expression. Proc Natl Acad Sci
USA. 98:11024–11031. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Roth KA and D'Sa C: Apoptosis and brain
development. Ment Retard Dev Disabil Res Rev. 7:261–266. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Love S: Apoptosis and brain ischaemia.
Prog Neuropsychopharmacol Biol Psychiatry. 27:267–282. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shi J, Miles DK, Orr BA, Massa SM and
Kernie SG: Injury-induced neurogenesis in Bax-deficient mice:
Evidence for regulation by voltage-gated potassium channels. Eur J
Neurosci. 25:3499–3512. 2007. View Article : Google Scholar : PubMed/NCBI
|