Introduction
Breast cancer, one of the most prevalent types of
malignant tumor in females, severely impairs the health of females
worldwide (1,2). Breast cancer is generally classified
into five subtypes, including luminal type A, luminal type B,
normal breast type, human epidermal growth factor receptor-2 (HER2)
overexpression type and basal type (3). For clinical treatment and prognostic
evaluation, it is conventional to assess the expression of three
breast cancer markers, including estrogen receptor (ER),
progesterone receptor (PR) and HER2, by immunohistochemistry, hence
the concept of triple-negative breast cancer (TNBC) (4). TNBC refers to breast cancer with
negative immunohistochemical results for ER, PR and HER2, and acts
as a specific breast cancer subtype, which was proposed in recent
years (5). Specific characteristics
of TNBC include high invasiveness and metastasis, high recurrence
and mortality rates, and a low survival rate. However, the exact
molecular mechanisms of TNBC remain unclear.
Rho-family GTPases, a primary branch of the Ras
superfamily of small GTPases, including RhoA and Ras-related C3
botulinum toxin substrate 1 (Rac1), are identified as crucial
regulators of actin cytoskeleton dynamics and endothelial
contractions (6). Deregulation of
these Rho-GTPases may result in a disorder of the actin
cytoskeleton and may further impact the integrity of the
endothelial barrier. Previous studies have demonstrated that
deregulated Rho-GTPases are associated with the development and
progression of various types of tumor, including testicular cancer
(7), colorectal tumors (8), gastric carcinoma (9) and breast cancer (10). Nevertheless, the precise role of these
signaling mechanisms in the migration and invasion of TNBC cells is
not well understood.
Dedicator of cytokinesis 1 (Dock1), a guanine
nucleotide exchange factor, has been proven to facilitate cell
survival, motility and proliferation via the activation of Rac1
(11,12). Engulfment and cell motility 1 (Elmo1)
serves as a mammalian homolog of Ced-12, which has been
evolutionarily conserved from worm to human. Elmo1 itself has no
intrinsic catalytic activity, but can regulate the activity of
interacting proteins as a scaffold protein (13). Furthermore, Elmo1 also joins multiple
cellular processes, including myoblast fusion, neurite outgrowth,
phagocytosis of apoptotic cells and cell migration (14). Additionally, previous studies have
demonstrated that by interacting with Dock1, Elmo1 acts as an
element of a bipartite guanine nucleotide-exchange factor for Rac1
(15–20). Recently, it was revealed that the
interaction between Dock1 and Elmo1 could modulate tumor
metastasis, which is enhanced by the actin cytoskeleton in breast
cancer (21). Therefore, Dock1 and
Elmo1 were selected as the objects of the present study, in order
to determine their exact roles and mechanisms in the migration and
invasion of TNBC epithelial cells.
The present study analyzed the association between
Dock1, Elmo1 and the migration and invasion of TNBC epithelial
cells. Furthermore, the roles and mechanisms of Dock1 and Elmo1
were investigated, together with those of the RhoA/Rac1 pathway in
the migration and invasion of TNBC epithelial cells.
Materials and methods
Cell culture
The human TNBC epithelial MDA-MB-231 cell line was
obtained from the Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China). Cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), in a 5% CO2
atmosphere at 37°C.
Cell transfection
Dock1 siRNA (targeting sequence:
5′-GGCCTACACTTTGCTTCTGC-3′), Elmo1 siRNA (targeting sequence:
5′-CGACAAUGUAACUCUGCAA-3′) and unspecific scrambled siRNA (50 nM)
(targeting sequence: 5′-ACGUGACACGUUCGGAGAATT-3′) vectors
(Invitrogen; Thermo Fisher Scientific, Inc.) were transfected into
MDA-MB-231 cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37°C for 48 h. Subsequent
experiments were performed 48 h after transfection.
Grouping
In the present study, there were four treatment
groups, including the control group (MDA-MB-231 cells), NC group
(MDA-MB-231 cells transfected with unspecific scrambled siRNA
vector), si-Dock1 group (MDA-MB-231 cells transfected with Dock1
siRNA vector) and si-Elmo1 group (MDA-MB-231 cells transfected with
Elmo1 siRNA vector).
Cell viability analysis
The viability of MDA-MB-231 cells was assessed by
the Cell Counting kit-8 (CCK-8; Beyotime Institute of
Biotechnology, Haimen, China). MDA-MB-231 cells (~6×104
cells/ml) in the logarithmic phase were plated into the wells of
96-well plates, prior to being incubated in a 5% CO2 atmosphere at
37°C for 12 h. Cells were then treated with one of the four
aforementioned treatments. Subsequently, cells were maintained for
12, 24 or 48 h. Subsequently, 10 µl CCK reagent were added into
each well and cells were maintained for 3 h. A microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to read
the absorbance at 450 nm. Cell viability was evaluated by the
percentage of live cells compared with the control.
Invasion and migration assay
Transwell assays were used to detect the migration
and invasion of cells. Matrigel inserts were used in the invasion
assay, and not the migration assay. Otherwise, the protocols were
identical for both assays. A 24-well Transwell chamber (8 µm;
Corning Incorporated, Corning, NY, USA) coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA) was used. Cultured MDA-MB-231
cells were suspended in serum-free DMEM (100 µl; Gibco; Thermo
Fisher Scientific, Inc.) and maintained for 24 h. Cells
(2×104 cells/ml) were then placed on the upper Transwell
chamber, and the lower chamber was filled with DMEM supplemented
with 10% FBS as chemoattractant. Following incubation for 24 h, the
cells that had invaded into the lower chamber were fixed with 4%
paraformaldehyde (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at
room temperature for 10 min and stained with 0.5% crystal violet
(Beijing Solarbio Science & Technology Co., Ltd.) at room
temperature for 30 min. The number of invaded cells was counted
visually under a fluorescence microscope (magnification, ×200;
Olympus Corporation, Tokyo, Japan).
Plate colony formation assay
Cultured MDA-MB-231 cells in the logarithmic phase
were digested by 0.25% trypsin (Gibco; Thermo Fisher Scientific,
Inc.), and then suspended in DMEM supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.). The cell suspension
(~6×104 cells/ml) was then inoculated in the culture
dishes, which were incubated in a 5% CO2 atmosphere at 37°C for 2
weeks. Subsequently, the supernatant was discarded and cells were
washed twice with PBS. Next, the cells were incubated with 5 ml 4%
paraformaldehyde for 15 min. The supernatant was then discarded and
Giemsa stain (Beijing Solarbio Science & Technology Co., Ltd.)
was added for 30 min at room temperature. Finally, cells were
washed with PBS and air-dried. The number of cell colonies was
counted visually using a fluorescence microscope (magnification,
×200).
Western blot analysis
The protein of cells was extracted using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). The bicinchoninic acid method was applied to assess
the concentration of protein. Protein lysates (35 µg) of cultured
MDA-MB-231 cells were separated by 12% SDS-PAGE. Subsequently, the
separated products were transferred to polyvinylidene difluoride
membranes (EMD Millipore, Billerica, MA, USA). TBST containing 5%
skim milk was used out to block the membranes at 37°C for 60 min.
Western blotting was performed using the following specific
antibodies: Rabbit anti-human anti-Dock1 (dilution, 1:1,000;
catalog no. ab97325; Abcam, Cambridge, UK), rabbit anti-human
anti-Elmo1 (dilution, 1:10,000; catalog no. ab174298; Abcam),
rabbit anti-human anti-FAK (dilution, 1:1,000; catalog no. ab40794;
Abcam), rabbit anti-human anti-Talin (dilution, 1:1,000; catalog
no. ab71333; Abcam), rabbit anti-human anti-Vinculin (dilution,
1:10,000; catalog no. ab129002; Abcam), rabbit anti-human anti-RhoA
(dilution, 1:5,000; catalog no. ab187027; Abcam), rabbit anti-human
anti-Rac1 (dilution, 1:1,000; catalog no. ab155938; Abcam) and
rabbit anti-human anti-actin (dilution, 1:5,000; catalog no.
ab179467; Abcam). A horseradish peroxidase-conjugated secondary
antibody (goat anti-mouse IgG-HRP; catalog no. sc-2005; dilution,
1:6,000; donkey anti-goat IgG-HRP; cat. no. sc-2020; dilution,
1:7,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) were
added, and the membranes were incubated at room temperature for 1
h. Enhanced chemiluminescent reagents (EMD Millipore, Billerica,
MA, USA) were added and an enhanced chemiluminescence system (GE
Healthcare, Chicago, IL, USA) was used for evaluating the results.
The blots were visualized by Gel documentation analyzer (Bio-Rad
Laboratories, Inc.) and densitometry was performed using Quantity
One 1-D software (version 4.62; Bio-Rad Laboratories, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cultured MDA-MB-231
cells using TRIzol reagent (Beijing Solarbio Science &
Technology Co., Ltd.). RNA was reverse transcribed to cDNA using a
Reverse Transcription kit (Beijing Solarbio Science &
Technology Co., Ltd.), according to the manufacturer's protocols.
The cDNA was amplified using SYBR Green qPCR Master Mix
(MedChenExpress, Monmouth Junction, NJ, USA). RT-qPCR analysis was
performed using an ABI 7500 Thermocycler (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The thermocycling conditions were
as follows: 10 min pre-treatment at 94°C, 97°C for 15 sec, 64°C for
45 sec (45 cycles), 97°C for 15 sec, 64°C for 1 min, 94°C for 15
sec, and a final extension at 75°C for 10 min. The primers were
designed by Invitrogen; Thermo Fisher Scientific, Inc.: Dock1
forward, 5′-CCGCCGCAAACTTTTTCCTC-3′ and reverse,
5′-AGATGTGCACAGTGTCTCCG-3′ (product: 222 bp); Elmo1 forward,
5′-GTAGGGACCCTTCTAGCTGC-3′ and reverse, 5′-GCTCAGGTTCTGCAGTTTGC-3′
(product: 202 bp); FAK forward, 5′-GAGCGTCTAATCCGACAGCA-3′ and
reverse, 5′-GGATTTCTTTCCGCCCAATTCT-3′ (product: 209 bp); Talin
forward, 5′-TAGCCTGAAAGGGAACTCGG-3′ and reverse,
5′-CTTCCGTCCTGGGAACGTC-3′ (product: 206 bp); Vinculin forward,
5′-GCAAAAGCTGTGGCTGGAAA-3′ and reverse, 5′-ATGTCATTGCCCTTGCTGGA-3′
(product: 356 bp); actin forward, 5′-GATATTGGCAACGACCCCCA-3′ and
reverse, 5′-CCCAGCCAGGATCTTGAAGG-3′ (product: 174 bp). Actin was
used as the control of the input RNA level. Data were quantified
using the 2−ΔΔCq method (22).
Statistical analysis
The statistical analyses were performed using SPSS
software (version 19.0; IBM Corp, Armonk, NY, USA). The results of
the present study are presented as the mean ± standard error of the
mean of at least three independent experiments. All experimental
data were analyzed using a one-way analysis of variance and the
post-hoc test was Fisher's least significant difference test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Downregulation of Dock1 and Elmo1
inhibits the viability of MDA-MB-231 cells
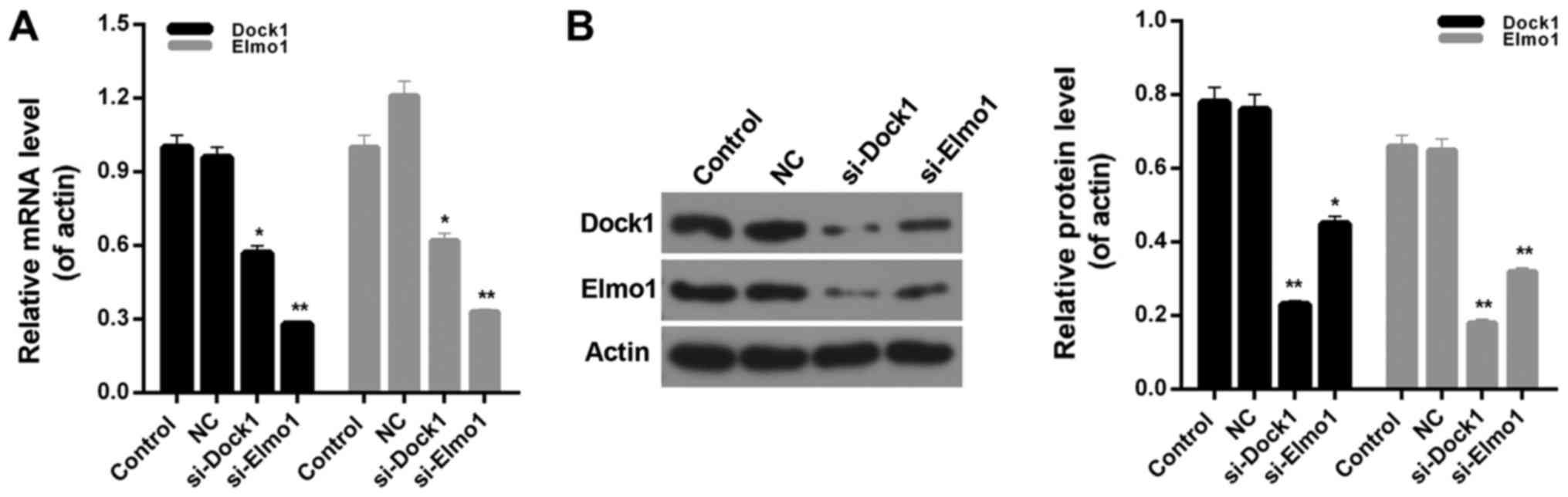
siRNA vectors targeting Dock1 and Elmo1, si-Dock1
and si-Elmo1, were constructed in the present study. The knockdown
efficiency was ~45 and 70% in MDA-MB-231 cells, following stable
transfection with si-Dock1 and si-Elmo1, respectively (Fig. 1A). Additionally, the western blot
analysis results indicated that following transfection with
si-Dock1, the expression levels of Dock1 and Elmo1 proteins in
MDA-MB-231 cells were significantly reduced (P<0.01; Fig. 1B). Furthermore, following transfection
with si-Elmo1, the expression levels of Dock1 and Elmo1 proteins in
MDA-MB-231 cells were significantly downregulated (P<0.05;
Fig. 1B). Therefore, it was confirmed
that interfering with the Dock1 and Elmo1 genes could
simultaneously impact the expression levels of Dock1 and Elmo1 in
MDA-MB-231 cells, which indicated that there is an interaction
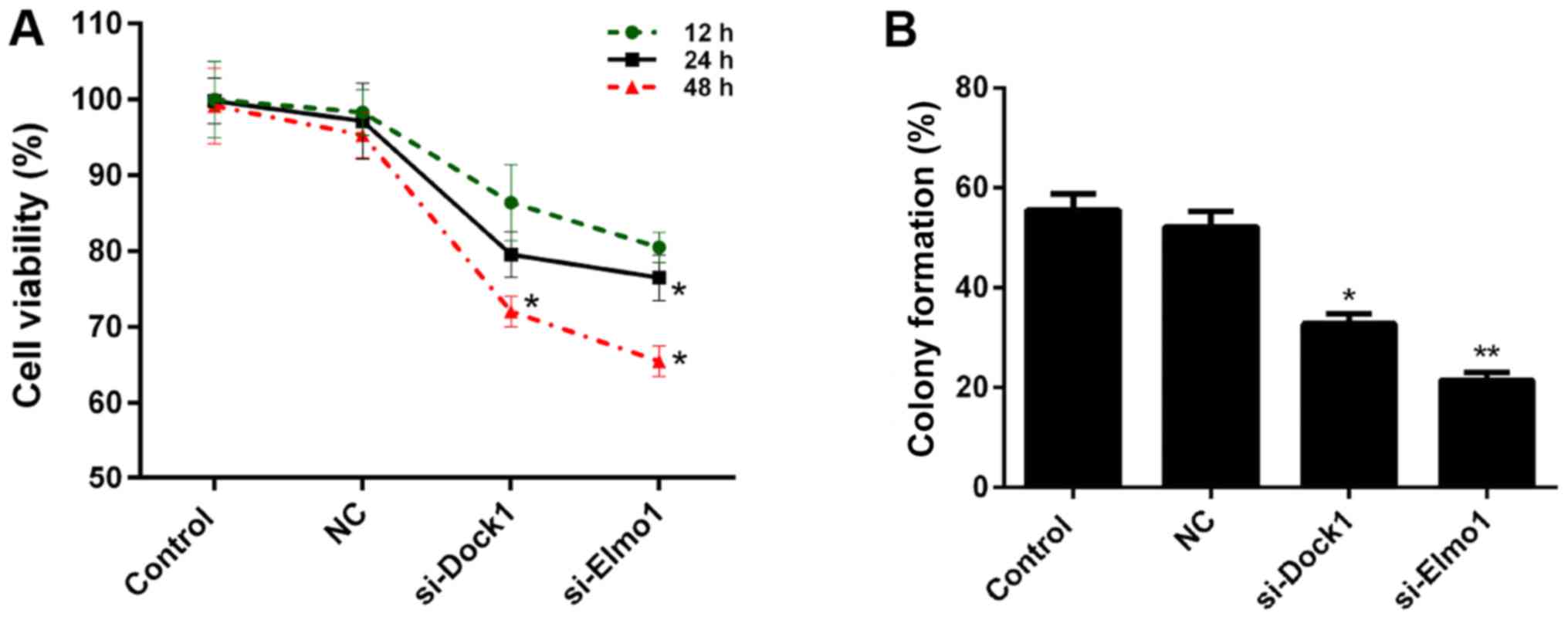
between Dock1 and Elmo1. Therefore, the CCK-8 assay was performed
to measure the viability of MDA-MB-231 cells in the aforementioned
treatment groups. The results demonstrate that, compared with the
control and NC groups, the viability of MDA-MB-231 cells
transfected with si-Dock1 and si-Elmo1, for 24 and 48 h, was
significantly reduced (P<0.05; Fig.
2A). Subsequently, the proliferation of MDA-MB-231 cells in the
four treatment groups was assessed. Compared with the NC group,
significant decreases in proliferation capacity were observed in
MDA-MB-231 cells transfected with si-Dock1 and si-Elmo1 (P<0.05;
Fig. 2B). Taken together, these
results suggested that the downregulation of Dock1 and Elmo1
inhibited the viability of MDA-MB-231 cells.
Downregulation of Dock1 and Elmo1
suppresses the migration and invasion of MDA-MB-231 cells
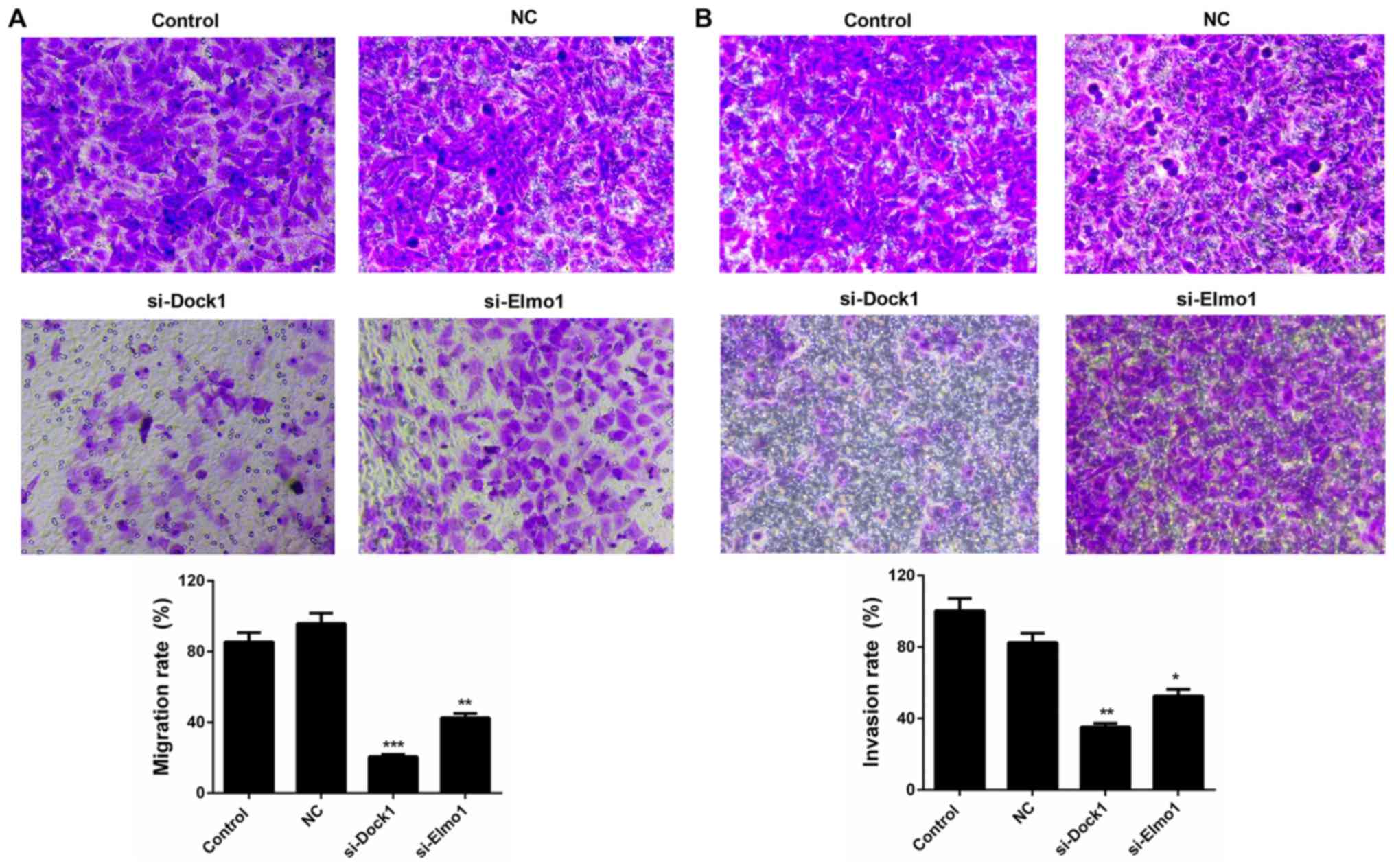
The migration and invasion capacities of MDA-MB-231
cells were also evaluated in the present study. The results of the
migration assay revealed that the migration ability of MDA-MB-231
cells transfected with si-Dock1 and si-Elmo1 was significantly
weaker than that of the NC group, decreasing from 95.84 to 20.51%
and 42.61%, respectively (P<0.01; Fig.
3A). A similar trend was also observed in the invasion ability
of MDA-MB-231 cells transfected with si-Dock1 and si-Elmo1, which
decreased from 82.61 to 35.22% and 52.64%, respectively (P<0.05;
Fig. 3B). Therefore, it was confirmed
that the downregulation of Dock1 and Elmo1 decreased the migration
and invasion abilities of MDA-MB-231 cells.
Downregulation of Dock1 and Elmo1
reduced Rac1 activity and the expression of migration-associated
proteins in MDA-MB-231 cells
In the present study, the associated migration
mechanisms and Rac1 activity in MDA-MB-231 cells were also
assessed. The results revealed that Rac1 activity in MDA-MB-231
cells was significantly reduced by transfecting with si-Dock1 and
si-Elmo1 (P<0.01; Fig. 4A).
Additionally, the expression levels of migration-associated
proteins, including focal adhesion kinase (FAK), Talin, and
Vinculin, in MDA-MB-231 cells were measured. The RT-qPCR results
demonstrated that Dock1 and Elmo1 silencing significantly decreased
the expression levels of FAK, Talin and Vinculin in MDA-MB-231
cells (P<0.05; Fig. 4B).
Furthermore, western blotting results also revealed similar trends
in the levels of migration-associated proteins in MDA-MB-231 cells
from each treatment group (Fig. 4C).
Based on these results, it was concluded that the downregulation of
Dock1 and Elmo1 decreased Rac1 activity and reduced the migration
and invasion abilities of MDA-MB-231 cells by downregulating the
expression of FAK, Talin and Vinculin.
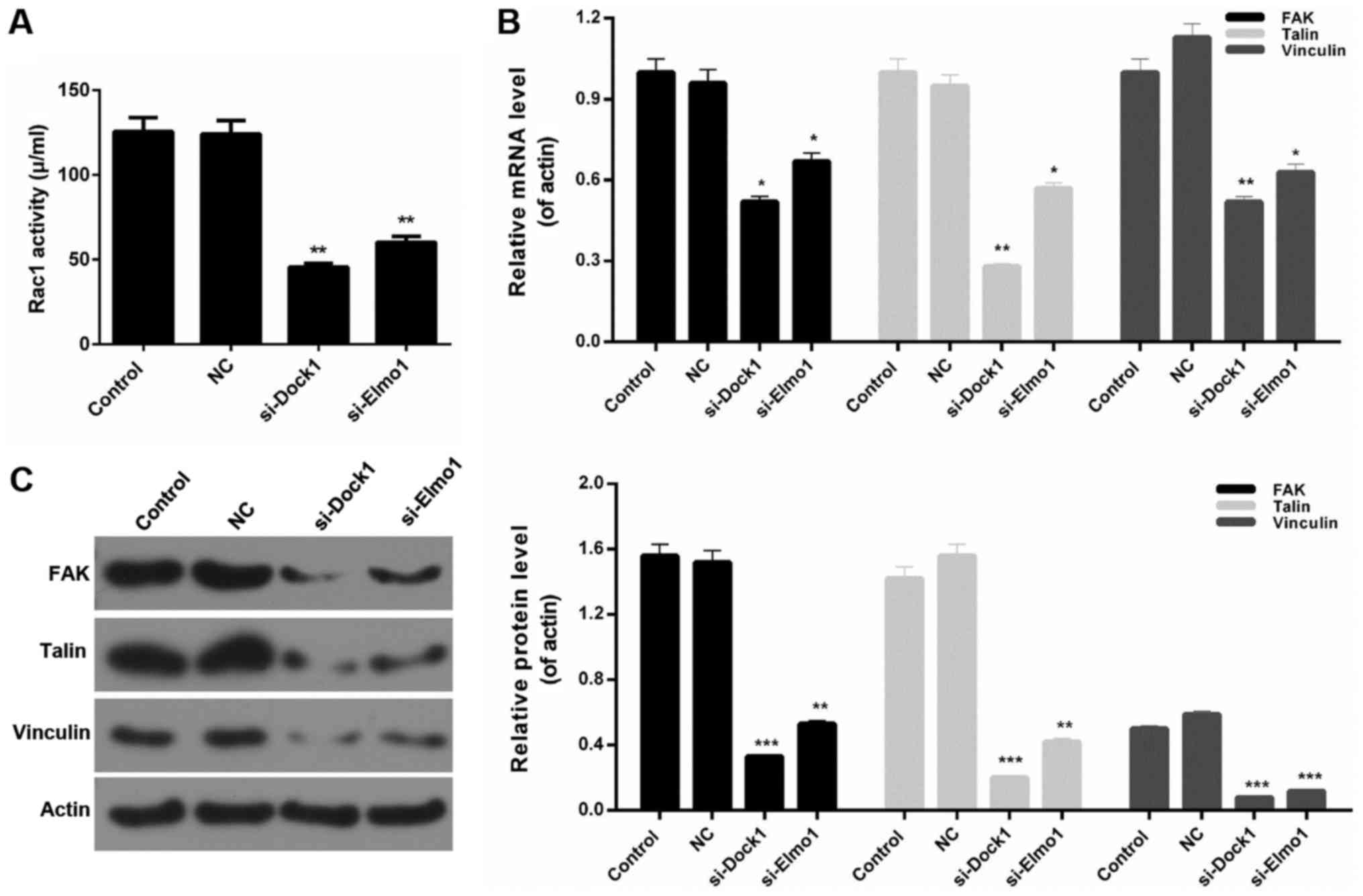 | Figure 4.Downregulation of Dock1 and Elmo1
reduces Rac1 activity and the expression of migration-associated
proteins in MDA-MB-231 cells. (A) A Rac activity assay was
performed to evaluate the Rac1 activity of MDA-MB-231 cells. (B)
Reverse transcription-quantitative polymerase chain reaction and
(C) western blot analysis assays were performed to measure the
expression levels of FAK, Talin and Vinculin in MDA-MB-231 cells,
MDA-MB-231 cells transfected with an empty vector, MDA-MB-231 cells
transfected with si-Dock1 and MDA-MB-231 cells transfected with
si-Elmo1. *P<0.05, **P<0.01 and ***P<0.001 vs. NC. Dock1,
dedicator of cytokinesis 1; Elmo1, engulfment and cell motility 1;
Rac1, Ras-related C3 botulinum toxin substrate 1; FAK, focal
adhesion kinase; si, small interfering RNA; NC, negative
control. |
Downregulation of Dock1 and Elmo1
affects the RhoA/Rac1 pathway
Finally, the expression of RhoA GTPase, total RhoA,
Rac1 GTPase and total Rac1 was evaluated in MDA-MB-231 cells from
all four treatment groups. Western blotting results indicated that
the expression level of RhoA GTPase in MDA-MB-231 cells was
significantly upregulated by transfecting with si-Dock1 and
si-Elmo1 (P<0.01; Fig. 5A).
Additionally, it was also revealed that the downregulation of Dock1
and Elmo1 significantly reduced the expression level of Rac1 GTPase
in MDA-MB-231 cells (Fig. 5B;
P<0.001). However, there was no significant difference in the
total RhoA and total Rac1 expression in MDA-MB-231 cells from all
treatment groups (Fig. 5A and B).
Therefore, it was concluded that the downregulation of Dock1 and
Elmo1 affected the RhoA/Rac1 pathway in MDA-MB-231 cells.
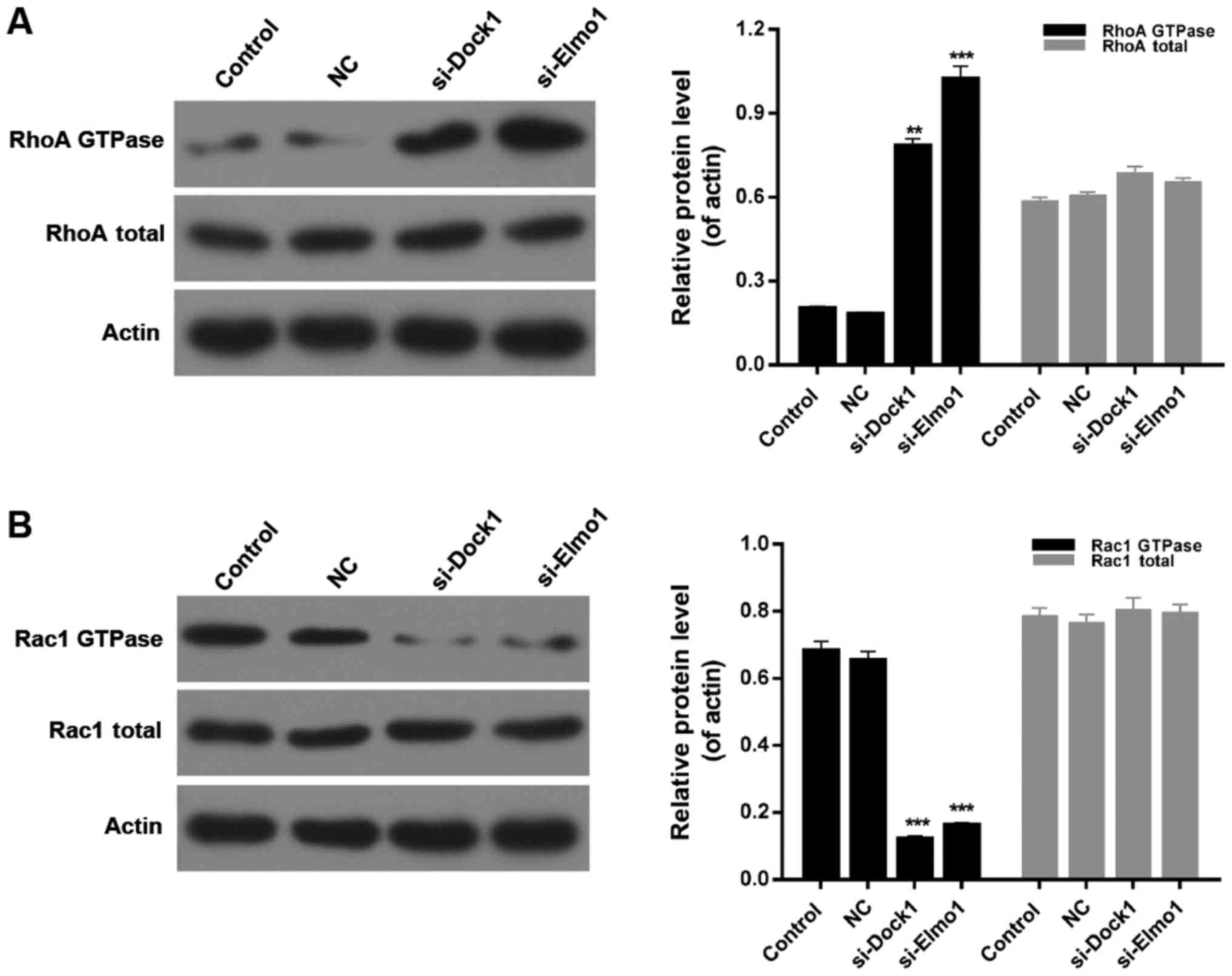 | Figure 5.Downregulation of Dock1 and Elmo1
affected the RhoA/Rac1 pathway. Western blot analyses were
performed to evaluate the expression levels of (A) RhoA QTPase and
RhoA total, and (B) Rac1 GTPase and Rac1 total in MDA-MB-231 cells,
MDA-MB-231 cells transfected with an empty vector, MDA-MB-231 cells
transfected with si-Dock1 and MDA-MB-231 cells transfected with
si-Elmo1. **P<0.01 and ***P<0.001 vs. NC. Dock1, dedicator of
cytokinesis 1; Elmo1, engulfment and cell motility 1; RhoA, Ras
homolog gene family, member A; Rac1, Ras-related C3 botulinum toxin
substrate 1; si, small interfering RNA; NC, negative control. |
Discussion
A previous study has demonstrated that Dock1 and
Elmo1 are highly expressed in invasive cancer cells, accelerating
cancer progression (23). In
addition, an increasing volume of evidence has revealed that the
silencing of Dock1 and Elmo1 can impede tumor development,
including cell proliferation, invasion and migration (17,24,25). To
the best of our knowledge, the roles of Dock1 and Elmo1 in the
prevention and treatment of TNBC have not yet been studied. In the
present study, plasmids cloned with si-Dock1 and si-Elmo1 were
prepared. The knockdown efficiency was ~45 and 70% in the
MDA-MB-231 cells, following stable transfection with si-Dock1 and
si-Elmo1, according to the RT-qPCR and western blot analysis data.
Additionally, an interaction between Dock1 and Elmo1 was
identified, which was consistent with the results of previous
studies (15–20). Therefore, the present study further
assessed the influences of Dock1 and Elmo1 on cell proliferation.
The results revealed that the downregulation of Dock1 and Elmo1
significantly inhibited the proliferation of MDA-MB-231 cells.
Cell motility is important for the majority of
pathological and physiological processes, including early embryonic
development, wound repair, and tumor cell migration and invasion
(26). Previous studies have
demonstrated that Dock1 and Elmo1 serve crucial roles in the
migration and invasion of various types of tumor cells (23,27–29).
Therefore, the effects of Dock1 and Elmo1 on the migration and
invasion capacities of MDA-MB-231 cells were assessed. The results
indicated that the downregulation of Dock1 and Elmo1 could
significantly decrease the migration and invasion abilities of
MDA-MB-231 cells.
Furthermore, by using the Transwell chamber assay,
it was demonstrated that Dock1 and Elmo1 synergistically promote
the Rac-dependent cell migration process (24). Additionally, the present study also
evaluated the Rac1 activity of MDA-MB-231 cells transfected with
si-Dock1 and si-Elmo1. The results revealed that the downregulation
of Dock1 and Elmo1 significantly reduced the proliferation ability
and Rac1 activity of MDA-MB-231 cells. Furthermore, the impact of
Dock1 and Elmo1 silencing on the migration and invasion of
MDA-MB-231 cells was investigated in the present study. Based on
previous reports, the expression levels in MDA-MB-231 cells of
several migration-associated proteins, including FAK, Talin and
Vinculin, were assessed (30–32). The data revealed that the
downregulation of Dock1 and Elmo1 markedly reduced the expression
levels of FAK, Talin and Vinculin in MDA-MB-231 cells. Therefore,
it was confirmed that the downregulation of Dock1 and Elmo1 may
decrease the migration and invasion abilities of MDA-MB-231 cells
by regulating FAK, Talin and Vinculin expression.
Previous studies have demonstrated that the
RhoA/Rac1 pathway is involved in the migration and invasion of
tumor cells (33–36). In the migration and invasion of breast
cancer cells, the RhoA/Rac1 pathway also serves vital functions
(36–39). Nonetheless, in TNBC epithelial cells,
the role of this pivotal, migration-associated signaling pathway
remains unclear. Therefore, the present study measured the
expression levels of RhoA GTPase, total RhoA, Rac1 GTPase and total
Rac1 in MDA-MB-231 cells from each treatment group, and revealed
that the downregulation of Dock1 and Elmo1 markedly enhanced the
expression of RhoA GTPase, while reducing the expression level of
Rac1 GTPase. Additionally, there was no significant difference in
the total RhoA and Rac1 expression between MDA-MB-231 cells from
each treatment group. Therefore, these results confirmed that the
downregulation of Dock1 and Elmo1 affected the RhoA/Rac1 pathway in
MDA-MB-231 cells.
Taken together, the results of the present study
demonstrated that the downregulation of Dock1 and Elmo1 decreased
the migration and invasion abilities of MDA-MB-231 cells by
affecting the RhoA/Rac1 pathway. These results provided novel
insights into the pathogenesis of TMBC, giving rise to novel
possibilities for TNBC therapy.
In summary, the present study demonstrates that the
downregulation of Dock1 and Elmo1 decreased the migration and
invasion abilities of TNBC epithelial cells through the RhoA/Rac1
pathway. These results offer crucial insight into the mechanisms of
Dock1 and Elmo1 in TNBC epithelial cells. The observed effects of
Dock1 and Elmo1 on the migration and invasion of TNBC epithelial
cells suggested that Dock1 and Elmo1 may be promising targets for
TNBC therapies.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL was responsible for the design of the experiment.
SW and YZ were responsible for processing data. YL was a major
contributor in writing the manuscript. All authors confirmed the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Donepudi MS, Kondapalli K, Amos SJ and
Venkanteshan P: Breast cancer statistics and markers. J Cancer Res
Ther. 10:506–511. 2014.PubMed/NCBI
|
|
3
|
Weigelt B, Eberle C, Cowell CF, Ng CK and
Reis-Filho JS: Metaplastic breast carcinoma: More than a special
type. Nat Rev Cancer. 14:147–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Denkert C, Liedtke C, Tutt A and von
Minckwitz G: Molecular alterations in triple-negative breast
cancer-the road to new treatment strategies. Lancet. 389:2430–2442.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bianchini G, Balko JM, Mayer IA, Sanders
ME and Gianni L: Triple-negative breast cancer: Challenges and
opportunities of a heterogeneous disease. Nat Rev Clin Oncol.
13:674–690. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wojciak-Stothard B and Ridley AJ: Rho
GTPases and the regulation of endothelial permeability. Vascul
Pharmacol. 39:187–199. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kamai T, Yamanishi T, Shirataki H, Takagi
K, Asami H, Ito Y and Yoshida K: Overexpression of RhoA, Rac1, and
Cdc42 GTPases is associated with progression in testicular cancer.
Clin Cancer Res. 10:4799–4805. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rihet S, Vielh P, Camonis J, Goud B,
Chevillard S and de Gunzburg J: Mutation status of genes encoding
RhoA, Rac1, and Cdc42 GTPases in a panel of invasive human
colorectal and breast tumors. J Cancer Res Clin Oncol. 127:733–738.
2001.PubMed/NCBI
|
|
9
|
Pan Y, Bi F, Liu N, Xue Y, Yao X, Zheng Y
and Fan D: Expression of seven main Rho family members in gastric
carcinoma. Biochem Biophys Res Commun. 315:686–691. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simpson KJ, Dugan AS and Mercurio AM:
Functional analysis of the contribution of RhoA and RhoC GTPases to
invasive breast carcinoma. Cancer Res. 64:8694–8701. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cote JF and Vuori K: GEF what? Dock180 and
related proteins help Rac to polarize cells in new ways. Trends
Cell Biol. 17:383–393. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng H, Hu B, Liu KW, Li Y, Lu X, Cheng T,
Yiin JJ, Lu S, Keezer S, Fenton T, et al: Activation of Rac1 by
Src-dependent phosphorylation of Dock180 (Y1811) mediates
PDGFRα-stimulated glioma tumorigenesis in mice and humans. J Clin
Invest. 121:4670–4684. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gumienny TL, Brugnera E, Tosello-Trampont
AC, Kinchen JM, Haney LB, Nishiwaki K, Walk SF, Nemergut ME, Macara
IG, Francis R, et al: CED-12/ELMO, a novel member of the
CrkII/Dock180/Rac pathway, is required for phagocytosis and cell
migration. Cell. 107:27–41. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim K, Lee J, Lee SA, Moon H, Park B, Kim
D, Joo YE and Park D: Intermolecular steric inhibition of Ephexin4
is relieved by Elmo1. Sci Rep. 7:44042017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hamoud N, Tran V, Croteau LP, Kania A and
Cote JF: G-protein coupled receptor BAI3 promotes myoblast fusion
in vertebrates. Proc Natl Acad Sci USA. 111:3745–3750. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hochreiter-Hufford AE, Lee CS, Kinchen JM,
Sokolowski JD, Arandjelovic S, Call JA, Klibanov AL, Yan Z, Mandell
JW and Ravichandran KS: Phosphatidylserine receptor BAI1 and
apoptotic cells as new promoters of myoblast fusion. Nature.
497:263–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Katoh H and Negishi M: RhoG activates Rac1
by direct interaction with the Dock180-binding protein Elmo.
Nature. 424:461–464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park D and Ravichandran KS: Emerging roles
of brain-specific angiogenesis inhibitor 1. Adv Exp Med Biol.
706:167–178. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park D, Tosello-Trampont AC, Elliott MR,
Lu M, Haney LB, Ma Z, Klibanov AL, Mandell JW and Ravichandran KS:
BAI1 is an engulfment receptor for apoptotic cells upstream of the
ELMO/Dock180/Rac module. Nature. 450:430–434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiao Y, Peng Y, Wan J, Tang G, Chen Y,
Tang J, Ye WC, Ip NY and Shi L: The atypical guanine nucleotide
exchange factor Dock4 regulates neurite differentiation through
modulation of Rac1 GTPase and actin dynamics. J Biol Chem.
288:20034–20045. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Laurin M, Huber J, Pelletier A, Houalla T,
Park M, Fukui Y, Haibe-Kains B, Muller WJ and Côté JF: Rac-specific
guanine nucleotide exchange factor DOCK1 is a critical regulator of
HER2-mediated breast cancer metastasis. Proc Natl Acad Sci USA.
110:7434–7439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jarzynka MJ, Hu B, Hui KM, Bar-Joseph I,
Gu W, Hirose T, Haney LB, Ravichandran KS, Nishikawa R and Cheng
SY: ELMO1 and Dock180, a bipartite Rac1 guanine nucleotide exchange
factor, promote human glioma cell invasion. Cancer Res.
67:7203–7211. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Grimsley CM, Kinchen JM, Tosello-Trampont
AC, Brugnera E, Haney LB, Lu M, Chen Q, Klingele D, Hengartner MO
and Ravichandran KS: Dock180 and ELMO1 proteins cooperate to
promote evolutionarily conserved Rac-dependent cell migration. J
Biol Chem. 279:6087–6097. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, Dai JM, Che YL, Gao YM, Peng HJ,
Liu B, Wang H and Linghu H: Elmo1 helps dock180 to regulate Rac1
activity and cell migration of ovarian cancer. Int J Gynecol
Cancer. 24:844–850. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Balassa T, Berta G, Jakab L, Bohonyi N and
Szekeres-Bartho J: The effect of the Progesterone-Induced Blocking
Factor (PIBF) on E-cadherin expression, cell motility and invasion
of primary tumour cell lines. J Reprod Immunol. 125:8–15. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sanders MA, Ampasala D and Basson MD:
DOCK5 and DOCK1 regulate Caco-2 intestinal epithelial cell
spreading and migration on collagen IV. J Biol Chem. 284:27–35.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi L, Zhang B, Sun X, Zhang X, Lv S, Li
H, Wang X, Zhao C, Zhang H, Xie X, et al: CC chemokine ligand 18
(CCL18) promotes migration and invasion of lung cancer cells by
binding to Nir1 through Nir1-ELMO1/DOC180 signaling pathway. Mol
Carcinog. 55:2051–2062. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tajiri H, Uruno T, Shirai T, Takaya D,
Matsunaga S, Setoyama D, Watanabe M, Kukimoto-Niino M, Oisaki K,
Ushijima M, et al: Targeting ras-driven cancer cell survival and
invasion through selective inhibition of DOCK1. Cell Rep.
19:969–980. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gu S, Papadopoulou N, Nasir O, Föller M,
Alevizopoulos K, Lang F and Stournaras C: Activation of membrane
androgen receptors in colon cancer inhibits the prosurvival signals
Akt/bad in vitro and in vivo and blocks migration via
vinculin/actin signaling. Mol Med. 17:48–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang C, Rajfur Z, Yousefi N, Chen Z,
Jacobson K and Ginsberg MH: Talin phosphorylation by Cdk5 regulates
Smurf1-mediated talin head ubiquitylation and cell migration. Nat
Cell Biol. 11:624–630. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sieg DJ, Hauck CR, Ilic D, Klingbeil CK,
Schaefer E, Damsky CH and Schlaepfer DD: FAK integrates
growth-factor and integrin signals to promote cell migration. Nat
Cell Biol. 2:249–256. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dreissigacker U, Mueller MS, Unger M,
Siegert P, Genze F, Gierschik P and Giehl K: Oncogenic K-Ras
down-regulates Rac1 and RhoA activity and enhances migration and
invasion of pancreatic carcinoma cells through activation of p38.
Cell Signal. 18:1156–1168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gulhati P, Bowen KA, Liu J, Stevens PD,
Rychahou PG, Chen M, Lee EY, Weiss HL, O'Connor KL, Gao T and Evers
BM: mTORC1 and mTORC2 regulate EMT, motility, and metastasis of
colorectal cancer via RhoA and Rac1 signaling pathways. Cancer Res.
71:3246–3256. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Parri M and Chiarugi P: Rac and Rho
GTPases in cancer cell motility control. Cell Commun Signal.
8:232010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao Y, Li J, Xing Y, Wang J, Lu C, Xin X
and Geng M: N-acetylglucosaminyltransferase V mediates cell
migration and invasion of mouse mammary tumor cells 4TO7 via RhoA
and Rac1 signaling pathway. Mol Cell Biochem. 309:199–208. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baugher PJ, Krishnamoorthy L, Price JE and
Dharmawardhane SF: Rac1 and Rac3 isoform activation is involved in
the invasive and metastatic phenotype of human breast cancer cells.
Breast Cancer Res. 7:R965–R974. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bourguignon LY, Zhu H, Shao L and Chen YW:
CD44 interaction with tiam1 promotes Rac1 signaling and hyaluronic
acid-mediated breast tumor cell migration. J Biol Chem.
275:1829–1838. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Burbelo P, Wellstein A and Pestell RG:
Altered Rho GTPase signaling pathways in breast cancer cells.
Breast Cancer Res Treat. 84:43–48. 2004. View Article : Google Scholar : PubMed/NCBI
|