Introduction
Lung cancer is one of the most malignant cancers,
with the highest morbidity and mortality among all cancers in the
world (1). Non-small cell lung cancer
(NSCLC) is closely associated with a long history of tobacco
smoking and accounts for ~80% of all lung cancer cases (2). The primary treatment for patients with
locally advanced or metastatic NSCLC includes systemic chemotherapy
(alone or in combination with radiation therapy) and targeted
biological therapy (3). However, lung
cancer cells frequently show significant resistance to
chemotherapeutic drugs (4).
Therefore, it is urgent to develop alternative therapeutic
strategies against lung cancer.
Traditional Chinese Medicine (TCM) has been
effectively used against diseases for thousands of years in China
(5,6).
Indeed, numerous Chinese medicinal herbs show promising effects in
the treatment of cancers (7).
Jinfukang (JFK), an Astragalus-based herbal formula consisting of
12 herbs (Table I), has been reported
to have anti-tumor efficacy against NSCLC (8–10). It has
been approved by the Chinese Food and Drug Administration as a drug
against NSCLC. Although a previous study reported that JFK could
inhibit proliferation and promote apoptosis of cancer cells in rats
(9), the underlying mechanisms remain
elusive.
 | Table I.Composition of JFK formula. |
Table I.
Composition of JFK formula.
| Botanical name | Herbal name | Common name | Chinese name | Ratio | Voucher no. |
|---|
| Astragalu
smembranaceus (Fish.) Bunge | Radix
astragali | Milkvetch root | Huang-qi | 8 |
SJTU-JFK140121-1 |
| Glehnia
littoralis F. Schmidt ex Miq. | Radix glehniae | Coastal glehnia
root | Bei-sha-shen | 8 |
SJTU-JFK140121-2 |
| Asparagus
cochinchinensis (Lour.) Merr | Radix asparagi | Cochinchinese
asparagus root | Tian-men-dong | 2 |
SJTU-JFK140121-3 |
| Ligustrum
lucidum W.T. Aiton | Fructus ligustri
lucidi | Glossy privet
fruit | Nv-zhen-zi | 2 |
SJTU-JFK140121-4 |
| Selaginella
doederleinii Hieron. | Herba
selaginellae | Spikemoss | Shi-Shang-Bai | 8 |
SJTU-JFK140121-5 |
| Paris
polyphylla Smith var. yunnanensis (Franch.) Hand Mazz | Rhizoma paridis
yunnanensis | Yunnan
manyleafparis rhizome | Chong-lou | 3 |
SJTU-JFK140121-6 |
| Epimedium
sagittatum (Siebold&Zucc.) Maxim | Folium
epimedii |
Shorthornedepimedium | Yin-yang-huo | 2 |
SJTU-JFK140121-7 |
| Gynostemma
pentaphyllum (Thunb.) Makino | Herbal
gynostemmatis | Five leaf
gynostemma herb | Jiao-gu-lan | 2 |
SJTU-JFK140121-8 |
| Cornus
officinalis Siebold & Zucc. | Fructus corni | Asiatic cornelian
cherry fruit | Shan-zhu-yu | 2 |
SJTU-JFK140121-9 |
| Salvia chinensis
Benth. | Herba salviae
chinensis | Chinese sage | Shi-jian-chuan | 8 |
SJTU-JFK140121-10 |
| Ophiopogon
japonicas (Thunb.) Ker Gawl | Radix
ophiopogon | Dwarf lilyturf | Mai-dong tuber | 2 |
SJTU-JFK140121-11 |
| Trigonella
foenum-graecum L. | Semen
trigonella | Common fenugreek
seed | Hu-lu-ba | 2 |
SJTU-JFK140121-12 |
As the majority of TCMs consist of various
components, it is necessary to utilize high-throughput strategies
to delineate the underlying mechanisms (11). Previous studies have shown that cDNA
profiling may be effectively used to characterize the antitumor
mechanism by measuring gene expression changes (12,13). With
the emergence of RNA-seq, this approach has provided a better
platform to investigate the mechanisms of various drugs due to its
performance in robustness and effectiveness (14,15).
A previous study indicated that JFK-induced
epigenetic alteration is involved in anti-lung cancer activity
(16). In present study, the
alteration in cellular growth and apoptosis upon JFK treatment was
examined in human lung cancer cell lines, and RNA-seq analysis was
performed to characterize the JFK-induced transcriptome. Among the
genes whose expression was modulated by JFK, the present study
further characterized those involved in the apoptosis pathway to
understand how JFK exerts its effects on lung cancer cells.
Materials and methods
Preparation of JFK
JFK contains 12 herbals described in Table I, and these raw herbs were obtained
from the pharmacy dispensary of Longhua Hospital, Shanghai
University of Traditional Chinese Medicine (Shanghai, China). The
names of herbs have been verified via www.theplantlist.org. Voucher specimens were deposited
at the herbarium in Shanghai Jiao Tong University. The raw
components of JFK (20 g) were minced and extracted with 200 ml 70%
ethanol at 80°C for 1 h according to the composition in Table I. Chromatographic and mass
spectrographic finger prints for JFK ethanol extract were examined
via LC-MS. The ethanol extract was filtered through a 0.45 µm
syringe filter and diluted to various concentrations used in the
present study.
Cell culture
The human lung cancer A549, NCI-H1975, NCI-H1650 and
NCI-H2228 cell lines were maintained with RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 100
U/ml penicillin and 100 µg/ml streptomycin (Thermo Fisher
Scientific, Inc.) at 37°C in a humidified atmosphere containing 5%
CO2.
Cell viability analysis
Human lung cancer cell lines were treated with
various concentrations of JFK extract (0, 0.041, 0.054, 0.081,
0.108, 0.162 and 0.216 mg/ml) for 48 h at 37°C, and then the cell
viability was evaluated using Cell Counting Kit 8 (CCK8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) by
spectrophotometric plate reader (Omega Bio-Tek, Inc., Norcross, GA,
USA). All cell viability results were tested by three independent
experiments.
Morphological changes and DAPI
staining
To evaluate the effects of JFK extract on cell
growth, 1×105 A549 cells were seeded in each well of
6-well plates, and exposed to JFK extract at a half maximal
inhibitory concentration (IC50; determined by CCK8 assay) for 48 h.
The morphological changes in A549 cells were observed with a
phase-contrast microscope (Model Ti-E; NIS4.0; Nikon, Tokyo,
Japan). A549 cells were fixed, stained with DAPI and observed using
fluorescence microscopy (Model Ti-E; NIS4.0; Nikon).
Cell cycle analysis
Briefly, A549 cells were harvested by trypsin with
no EDTA (Thermo Fisher Scientific, Inc.), and washed twice with
PBS. The cells were fixed with cold 70% ethanol overnight, and then
stained with a solution consisting of 20 µg/ml propidium iodine
(PI) (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 0.1%
TritonX-100 (Sigma-Aldrich; Merck KGaA), 100 µg/ml RNase A
(Fermentas; Thermo Fisher Scientific, Inc.) for 15 min at 37°C in
the dark. Cell cycle progression was then analyzed by flow
cytometry (BD LSRFortessa; BD Biosciences, Franklin Lakes, NJ, USA)
and ModFit LT software (version 3.2; Verity Software House,
Topsham, ME, USA).
Cell apoptosis analysis
Determination of phosphatidyl serine (PS) and
membrane integrity was performed using the Annexin V-FITC/PI
Apoptosis kit (Beijing Zoman Biotechnology Co., Ltd., Beijing,
China). In brief, A549, NCI-H1975, NCI-H1650 and NCI-H2228 cells
were harvested by trypsin (no EDTA) and washed twice with PBS, then
stained with Annexin V-FITC/PI and analyzed by flow cytometry as
aforementioned. PI positive cells were designated as end stage
apoptotic cells, and FITC-positive cells were designated as early
stage apoptotic cells.
RNA-seq library construction
RNA-seq assays were performed in A549 cells without
and with JFK treatment for 48 h (0.054 mg/ml), as described in our
recent study (17). Briefly, total
RNA was extracted by TRIzol (Thermo Fisher Scientific, Inc.), and
further treated with DNase to remove genomic DNA contamination.
Isolation of mRNA was performed with Oligotex mRNA Mini kit
(Qiagen, Hilden, Germany) and then used for RNA-seq library
construction with NEBNext Ultra Directional RNA Library Prep kit
for Illumina (New England BioLabs, Inc., Ipswich, MA, USA), which
was subjected to Illumina sequencing. The raw sequencing data are
available in the EMBL database (www.ebi.ac.uk/arrayexpress) under accession number
E-MTAB-3592.
Analysis of differentially expressed
genes
The raw sequencing reads were mapped to the
reference genome (hg19) by TopHat. Cufflinks was used to
characterize the differential transcription pattern (18). The present study has taken biases in
library preparation into account. The gene expression level was
measured by reads per kilobase of transcript per million reads
mapped.
Functional annotation and pathway
analysis
The Database for Annotation, Visualization and
Integrated Discovery (DAVID) bioinformatics resource was used to
annotate gene functions and pathways (19,20).
Quantification of mRNA level
Transcription level of genes of interest was
evaluated using quantitative reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) with SYBR-Green PCR Master Mix
(Qiagen). RT-qPCR was performed according to our previous studies
(13,16,17). All
primer sequences used for RT-qPCR are presented in Table II.
| Table II.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
|
|
| Primer
sequence |
|---|
|
|
|
|
|---|
| Gene | GenBank | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| CDK2 | [NG_034014] |
TACACCCATGAGGTGACTCG |
GTCCCCAGAGTCCGAAAGAT |
| CDK4 | [NM_000075] |
CCCGAAGTTCTTCTGCAGTC |
CTGGTCGGCTTCAGAGTTTC |
| CCNA2 | [NM_001237] |
GGTACTGAAGTCCGGGAACC |
AAGATCCTTAAGGGGTGCAA |
| CCNB1 | [NM_172301] |
TGTGGATGCAGAAGATGGAG |
TTTGGTCTGACTGCTTGCTC |
| DR4 | [NM_003844] |
TGCAGGTCGTACCTAGCTCA |
GGACACAACTCTCCCAAAGG |
| Fas | [NG_009089] |
ATTGCTCAACAACCATGCTG |
CCAATCCCTTGGAGTTGATG |
| TP53BP2 | [NG_029950] |
TCCTTGGTCATTCAGGCTTC |
GGCCAACTGGATGGATTTTA |
| BNIP3L | [NM_004331] |
CAACAACAACTGCGAGGAAA |
TTGCTGCTGTTCATGGGTAG |
| GAPDH | [NG_007073] |
GCACCGTCAAGGCTGAGAAC |
ATGGTGGTGAAGACGCCAGT |
Western blot analysis
Cells were washed with PBS and collected in a 1.5 ml
tube. A549 cells were lysed using the procedures essentially the
same as described previously (13).
The total protein extracts were obtained following centrifugation
(18,000 × g for 15 min, at 4°C). The protein concentration was
evaluated by Pierce BCA protein assay kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Protein samples (~50 µg) were fractionated by SDS-PAGE (10%
polyacrylamide gels) and transferred to a polyvinylidene fluoride
membrane (EMD Millipore, Billerica, MA, USA). The protein-binding
membrane was incubated in PBS containing 0.1% Tween-20
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 60 min at 4°C.
5% skim milk was used to block non-specific binding for 3 h at 4°C.
This was followed by incubation overnight at 4°C with primary
antibody. The primary antibodies used in the present study
included; cyclin B1 (CCNB1; cat no. D160234-0100; dilution,
1:250; Sangon Biotech Co., Ltd., Shanghai, China), cyclin-dependent
kinase 4 (CDK4; cat no. 11026-1-AP; dilution, 1:100;
Proteintech Group, Inc., Wuhan, China), death receptor 4
(DR4; cat no. 24063-1-AP; dilution, 1:100; Proteintech
Group, Inc.), Fas (cat no. D220092-0100; dilution, 1:550;
Sangon Biotech Co., Ltd.), and GAPDH (cat no. 5174S; dilution,
1:2,000; Cell Signaling Technology, Inc., Danvers, MA, USA). The
membrane with protein antibody complexes were washed with PBS
containing 0.1% Tween-20, incubated for 2 h with a goat anti-rabbit
horseradish peroxidase-conjugated secondary antibody (cat no.
14708S; dilution, 1:15,000; Cell Signaling Technology, Inc.) at
4°C. Immunodetection was performed using ECL reagent (Santa Cruz
Biotechnology Inc., Dallas, TX, USA) and images were captured and
analyzed using the LI-COR Odyssey Infrared Imaging system (LI-COR
Biosciences, Lincoln, NE, USA). The protein amounts were
semi-quantified by analyzing blot intensity using GAPDH as loading
control.
siRNA and transfection
siRNA duplexes were designed to downregulate
DR4 and Fas. The DR4-targeting siRNA sequence
was 5′-CCUUCAAGUUUGUCGUCGUdTdT-3′ and the Fas-targeting
siRNA sequence was 5′-GUGCAGAUGUAAACCAAACdTdT-3′. siRNAs for in
vitro transfection were obtained from Shanghai Gene Pharma Co.,
Ltd. (Shanghai, China). Transfection was performed using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's instructions. Briefly, A549 and NCI-H2228 cells were
transfected with 50 nmol/l of siRNAs directed against Fas
and DR4 respectively, and incubated for 6 h at 37°C.
Statistical analysis
Data are presented as the mean ± standard deviation.
Comparisons between the groups were examined by standard one-way
analysis of variance ANOVA, SPSS for Windows 14.0 software package
(SPSS, Inc., Chicago, IL, USA), followed by Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
JFK induces cell growth arrest and DNA
damage in lung cancer cells
To examine the effects of JFK on lung cancer cell
growth, A549, NCI-H1975, NCI-H1650 and NCI-H2228 cells were treated
with JFK at various concentrations (0, 0.041, 0.054, 0.081, 0.108,
0.162 and 0.216 mg/ml) for 48 h and cell viability was assessed.
The cell viability of these human lung cancer cell lines decreases
significantly in a concentration-dependent manner following JFK
treatment (Fig. 1A). Additionally,
JFK was observed to exert an increased inhibition effect on A549
cells compared with other three human lung cancer cell lines, with
a IC50 of ~0.054 mg/ml. JFK treatment (0.054 mg/ml) for 48 h
reshapes a large number of A549 cells to be distinctly flat and
enlarged in size (Fig. 1B) and makes
the nucleus condensed and disintegrated (Fig. 1C), indicating multiple effects of JFK
on A549 cells. Based on these observations, A549 was next focused
on and the antitumor mechanisms of JFK were characterized.
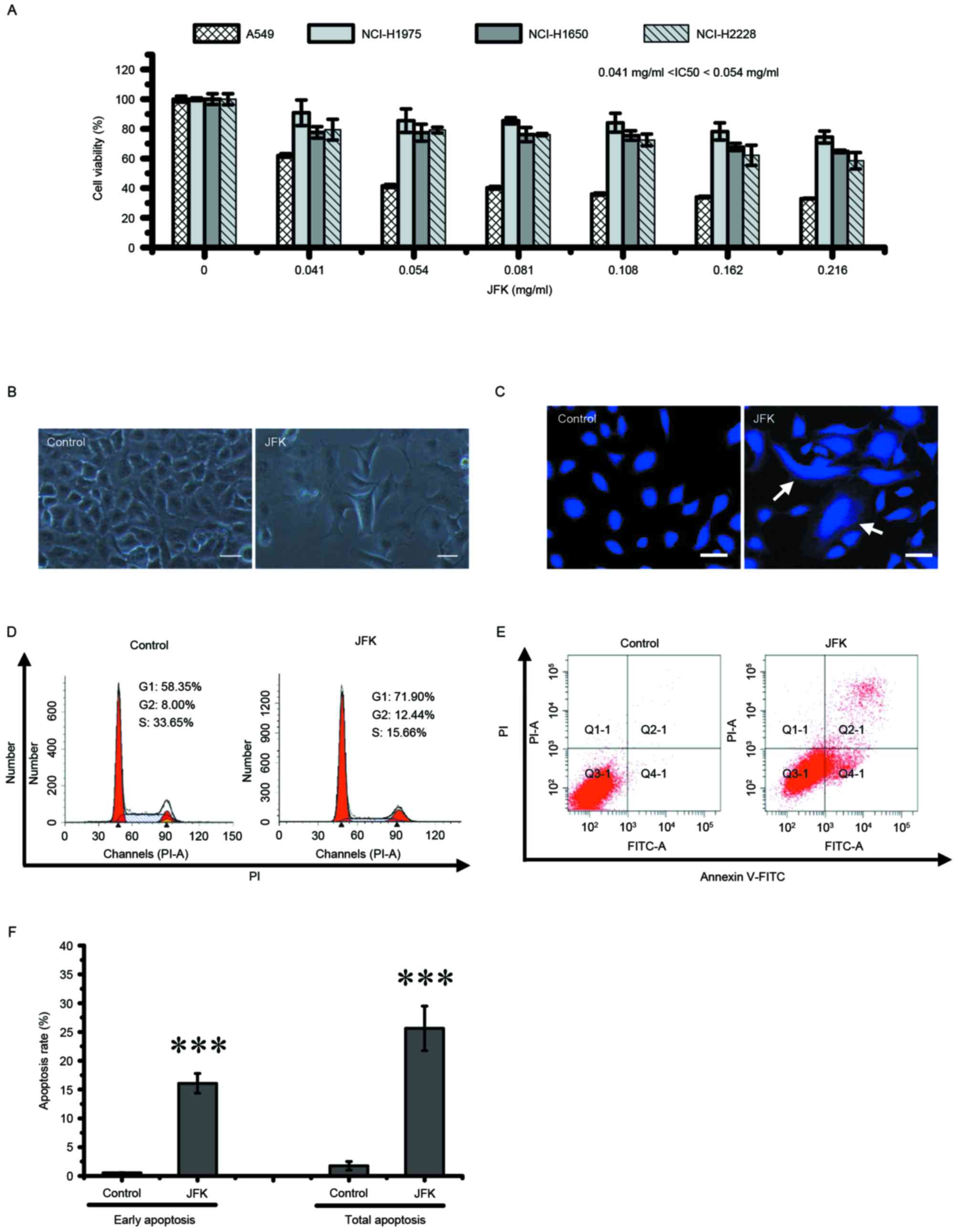 | Figure 1.Effect of JFK on cytotoxicity in lung
cancer cells. (A) A549, NCI-H1975, NCI-H1650 and NCI-H2228 cells
were treated with JFK (0, 0.041, 0.054, 0.081, 0.108, 0.162 and
0.216 mg/ml, respectively) for 48 h, and examined for viability.
Data are expressed as the mean ± SD (n=3). (B) A549 cells were
exposed to JFK (0.054 mg/ml) for 48 h. Morphological alteration was
examined by phase-contrast microscopy. Scale bar, 50 µm. (C) A549
cells were stained with DAPI for detecting nucleus changes using
fluorescence microscopy. Scale bar, 25 µm. (D) Cell cycle
distribution was evaluated by flow cytometry. (E) Apoptotic cells
were determined by flow cytometry after exposure to JFK for 48 h.
(F) Ratios of early apoptosis and total apoptosis were analyzed
basing on flow cytometric detection. Data are expressed as the mean
± SD (n=3). ***P<0.001. SD, standard deviation; JFK, Jinfukang;
PI, propidium iodide; FITC, fluorescein isothiocyanate; IC50, half
maximal inhibitory concentration. |
JFK leads to cell cycle arrest and
cell apoptosis in A549 cells
To further investigate whether JFK cytotoxicity
involves cell cycle arrest, a flow cytometry assay was performed to
characterize alterations of the cell cycle. As shown in Fig. 1D, cell cycle analysis indicates that a
significant portion of A549 cells accumulates in G1 and G2 phase
upon JFK (0.054 mg/ml) treatment, suggesting that JFK could induce
G1/S and G2/M transition arrest. To examine whether JFK triggers
apoptosis in A549 cells, the extent of apoptotic cells was
investigated by flow cytometry, and it was observed that numerous
A549 cells were induced to undergo apoptosis after exposure to JFK
for 48 h (Fig. 1E). This phenomenon
not only presents in early apoptosis, but also presents in total
apoptosis (Fig. 1F). Thus, the cell
apoptotic rates dramatically increase upon JFK treatment.
JFK-associated transcriptome analysis
in A549 cells
Given the observations of anti-proliferation and
pro-apoptosis effects induced by JFK, the present study
subsequently aimed to understand the underlying molecular
mechanisms by transcriptome-wide differential gene expression
analysis through RNA-seq. In total, 13,130,096 and 10,433,359 reads
were generated from JFK-treated and untreated A549 cells, 54 and
66% of which were uniquely mapped to the human genome,
corresponding to 13,720 and 17,519 expressed genes, respectively
(Table III). Compared with the
control, 5,281 differentially expressed genes (P<0.05) were
identified, with 2,777 (52.6%) downregulated and 2,504 (47.4%)
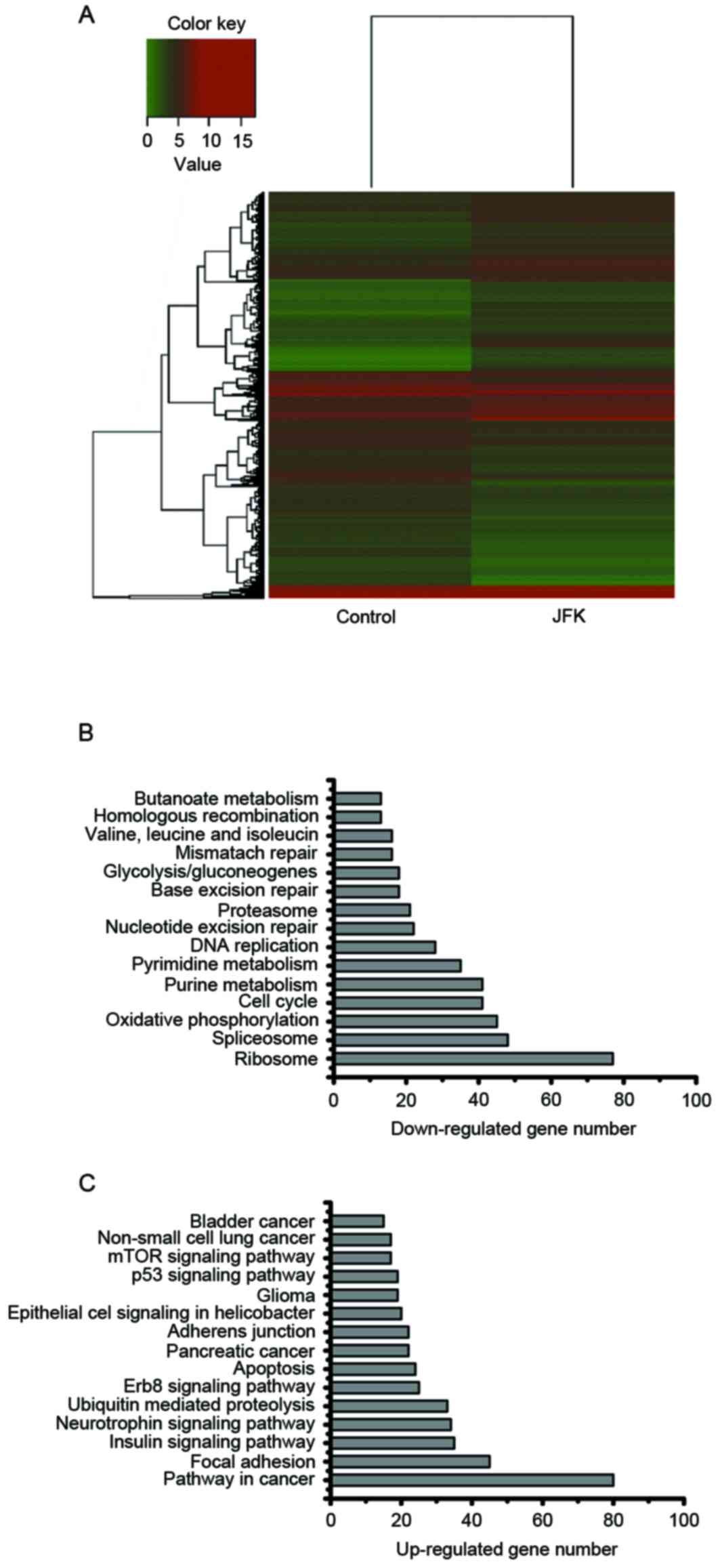
upregulated (Fig. 2A).
| Table III.The statistics of RNA-seq data
sets. |
Table III.
The statistics of RNA-seq data
sets.
| Group | Total reads | Unique mapping
reads | Mapping rate,
% | Expressed gene
number |
|---|
| Control | 10433359 | 6908569 | 66 | 17519 |
| JFK | 13130096 | 7035144 | 54 | 13720 |
Gene ontology analysis of
differentially expressed genes
Gene Ontology (GO) analysis of the genes with
JFK-induced differential expression was performed using DAVID
(19) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) pathways database (20). The top ten GO items were listed in
Table IV, in which
apoptosis-associated and cell cycle-associated biological processes
are statistically enriched in the upregulated and downregulated
genes, respectively. For the gene sets with differential
expression, KEGG pathway analysis was performed. Similarly, it was
observed that the cell cycle and apoptosis pathways are also
included in the top 10 enriched pathways (Fig. 2B and C). These results suggest that
JFK could induce growth inhibition potentially through repressing
pathways of cell cycle, nuclear division and translation pathways.
Simultaneously, it could induce apoptosis in A549 cells partially
through activating apoptosis, cell death and protein transport
pathways.
| Table IV.GO analysis of differentially
expressed genes. |
Table IV.
GO analysis of differentially
expressed genes.
| A,
Downregulated |
|---|
|
| Gene ID | Function | Count | P-value |
|---|
| GO:0006414 | Translational
elongation | 85 |
6.79×10−57 |
| GO:0006412 | Translation | 140 |
6.93×10−39 |
| GO:0007049 | Cell cycle | 226 |
2.69×10−32 |
| GO:0022402 | Cell cycle
process | 178 |
7.65×10−30 |
| GO:0000278 | Mitotic cell
cycle | 132 |
7.16×10−28 |
| GO:0000279 | M phase | 120 |
2.39×10−26 |
| GO:0022403 | Cell cycle
phase | 134 |
1.00×10−23 |
| GO:0000087 | M phase of mitotic
cell cycle | 88 |
5.95×10−22 |
| GO:0000280 | Nuclear
division | 87 |
6.22×10−22 |
| GO:0007067 | Mitosis | 87 |
6.22×10−22 |
|
| B,
Upregulated |
|
| Gene ID |
Function | Count | P-value |
|
| GO:0046907 | Intracellular
transport | 150 |
5.33×10−15 |
| GO:0015031 | Protein
transport | 166 |
1.36×10−14 |
| GO:0045184 | Establishment of
protein localization | 167 |
1.51×10−14 |
| GO:0008104 | Protein
localization | 182 |
1.28×10−13 |
| GO:0010941 | Regulation of cell
death | 165 |
1.09×10−11 |
| GO:0042981 | Regulation of
apoptosis | 163 |
1.32×10−11 |
| GO:0043067 | Regulation of
programmed cell death | 164 |
1.55×10−11 |
| GO:0016265 | Death | 149 |
3.69×10−11 |
| GO:0008219 | Cell death | 148 |
4.29×10−11 |
| GO:0034613 | Cellular protein
localization | 95 |
4.45×10−10 |
JFK modulates the expression of genes
involved in cell cycle and apoptosis
For gene sets with a differential expression pattern
determined by RNA-seq analysis, cell cycle-associated genes,
including CDK2, CDK4, CCNB1 and cyclin A2 (CCNA2),
and apoptosis-associated genes, including Fas, DR4, tumor
protein P53 binding protein 2 (TP53BP2) and BCL2 interacting
protein 3 like (BNIP3L), for further validation (Fig. 3A). The transcriptional activities of
selected genes were measured by RT-qPCR, and the results were shown
in Fig. 3B and C. These results
confirm that JFK represses cell cycle-associated genes, and
activates apoptosis-associated genes as well. Among these selected
genes, the alterations in protein level of CDK4, CCNB1, Fas
and DR4 were examined by western blot analysis. As shown in
Fig. 3D and E, protein levels of
CDK4 and CCNB1 decrease and those of Fas and
DR4 increase after JFK (0.054 mg/ml) treatment.
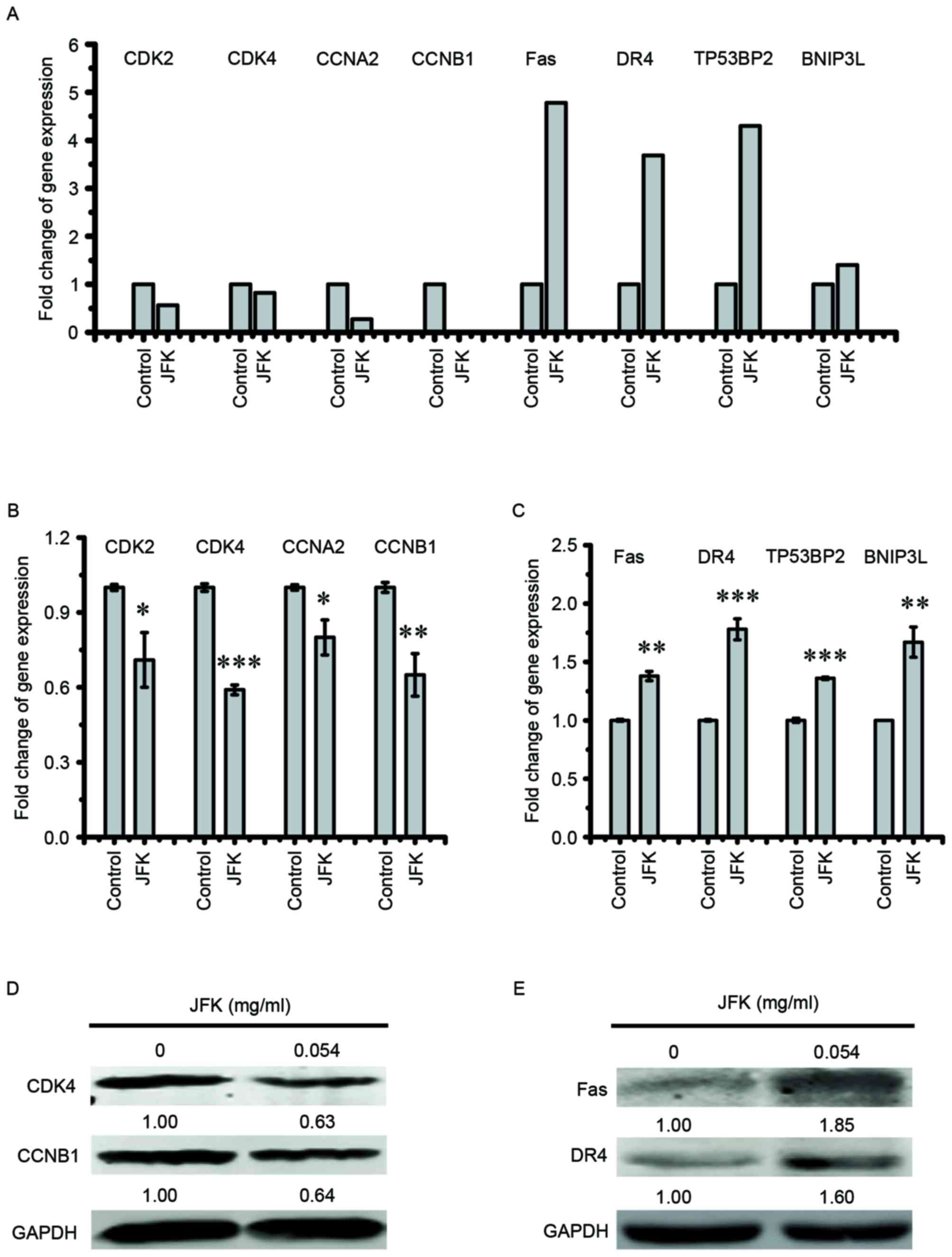 | Figure 3.Expression validation of genes
involved in the cell cycle and apoptosis upon JKF treatment in A549
cells. (A) Fold-change of cell cycle and cell apoptosis-associated
gene expression from RNA-seq results. (B and C) A549 cells were
treated with JFK (0 or 0.054 mg/ml) for 48 h, and then the mRNA
levels of cell cycle and apoptosis-associated genes were examined
by reverse transcription-quantitative polymerase chain reaction.
The data are expressed as the mean ± standard deviation (n=5).
*P<0.05, **P<0.01 and ***P<0.001. (D and E) A549 cells
were treated with or without JFK (0.054 mg/ml) for 48 h, and then
the protein levels of CDK4, CCNB1, Fas and DR4 were
examined by western blot analysis. CDK4, cyclin-dependent kinase 4;
CDK2, cyclin-dependent kinase 2; CCNB1, cyclin B1; CCNA2, cyclin
A2; TP53BP2, tumor protein P53 binding protein 2; BNIP3L, BCL2
interacting protein 3 like; JFK, Jinfukang. |
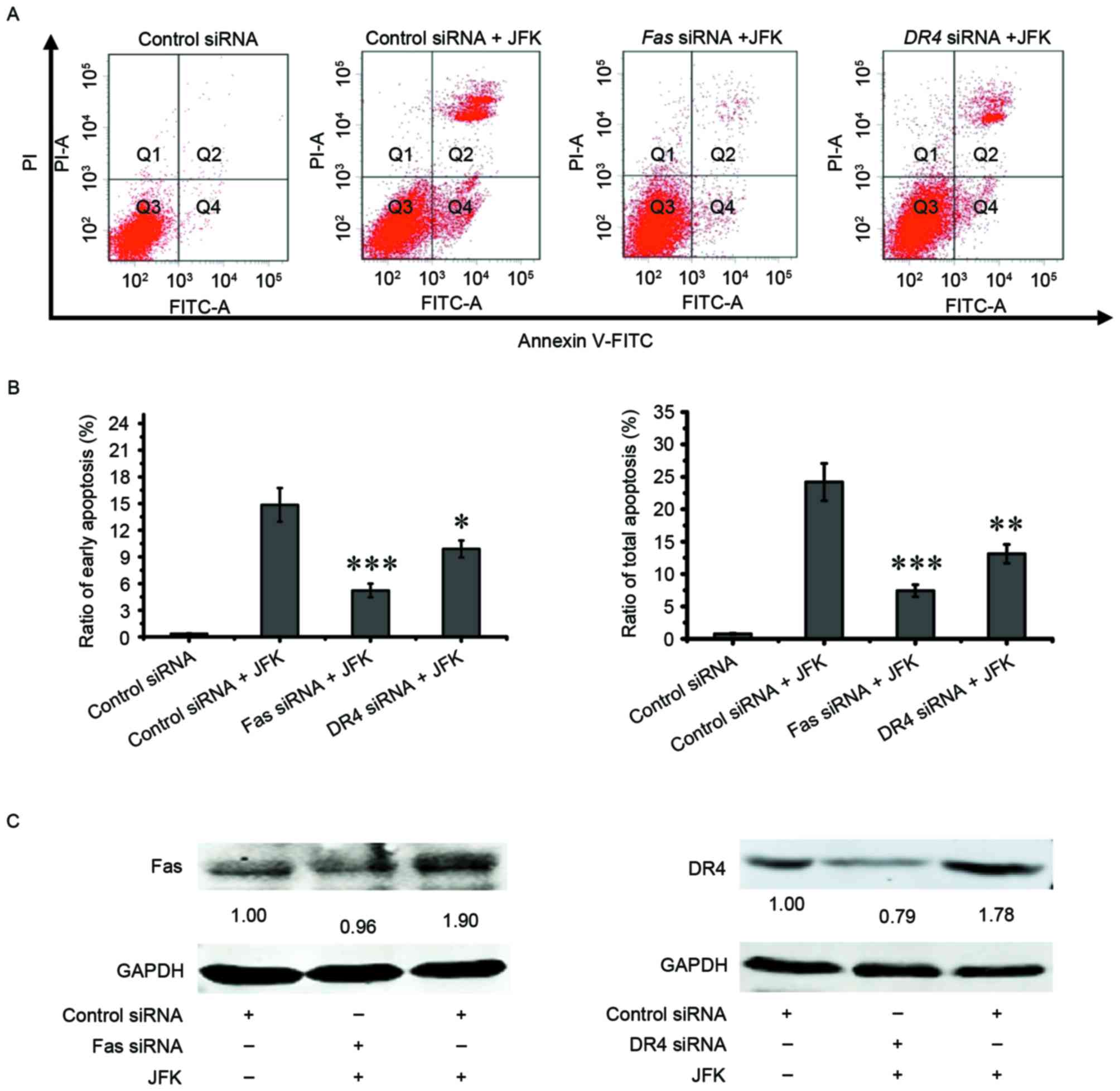
Activation of Fas and DR4 contributes
to JFK-induced pro-apoptotic effect in A549 cells
To explore whether Fas and DR4 mediate
the JFK-induced apoptosis in A549 cells, lastly siRNA assays were
performed to suppress the expression of Fas and DR4
in JFK-treated A549 cells. As shown in Fig. 4A and B, the apoptotic rates (including
early apoptosis rate and total apoptosis rate) significantly
decrease when Fas and DR4 were knocked down. The
protein expression levels of Fas and DR4 also drop
when Fas and DR4 were knocked down (Fig. 4C). These results indicate that
activation of Fas and DR4 is partially involved in
JFK-induced apoptosis.
Discussion
Herbal intervention has been widely used for the
treatment and prevention of various diseases for thousands of years
in China (21). The majority of known
anticancer drugs are derived from herbal plants (22). JFK, a type of Astragalus-based herbal
formula, has been reported to be clinically effective in NSCLC
patients (8,10,23). In
the present study, the anticancer mechanisms of JFK were explored.
The cellular behavior of lung cancer cell lines treated with JFK
were examined, and it was found that JFK could induce cellular
growth arrest and the cell cycle is regulated by a complex cascade
of events and mediated by cell cycle regulatory proteins, including
cyclins and CDKs (24). CDK2 and CDK4
are two key members of the cyclin-dependent kinase family, whose
activities are restricted to the G1-S phase, and are essential for
the G1-Stransition (25,26). CCNA2 is a critical regulatory factor
that can interact with CDK kinases for regulating G1-S transition
and G2-M transition (27). CCNB1
usually has an important role in mitosis, and indicates a poor
prognosis when overexpressed in NSCLC (28). The present results show that A549
cells could be induced to a state of G1 and G2 phase enrichment
upon JFK treatment (Fig. 1D),
accompanying downregulation of cell cycle-associated genes,
including CDK2, CDK4, CCNB1 and CCNA2 (Figs. 3A and 4B). These results suggest that JFK leads to
A549 cell cycle arrest through the suppression of the cell cycle
pathway.
Induction of apoptosis in tumor cells is one of the
most common anticancer mechanisms in cancer therapies (29). The death receptor apoptotic pathway
has previously been proposed as an anti-cancer drug target in human
lung cancer (30). Fas and
DR4, two members of the death receptor family, have a vital
role during the process of apoptosis. TP53BP2 induces
apoptosis through the mitochondrial death pathway (31). BNIP3L has been reported to
promote cell death via apoptosis (32). In the present study, it was found the
expression activities of Fas and DR4 in A549 cells
are upregulated in both mRNA and protein levels after JFK
treatment. TP53BP2 and BNIP3L are also activated by
JFK (Fig. 3A and C). Particularly, it
was observed that downregulation of Fas and DR4 could
attenuate JFK-induced apoptosis in A549 cells (Fig. 4). However, this phenomenon was not
observed to occur in the human lung cancer NCI-H1650 and NCI-H228
cell lines, although JFK also exerts an inhibitory viability effect
on these two cell lines (Fig. 5A-C).
JFK enhances the transcription of DR4 and Fas in
NCI-H2228 cells, but induces marginal apoptosis in siRNA knockdown
assays (Fig. 5D-F), suggesting
JFK-induced apoptosis is cellular context-dependent.
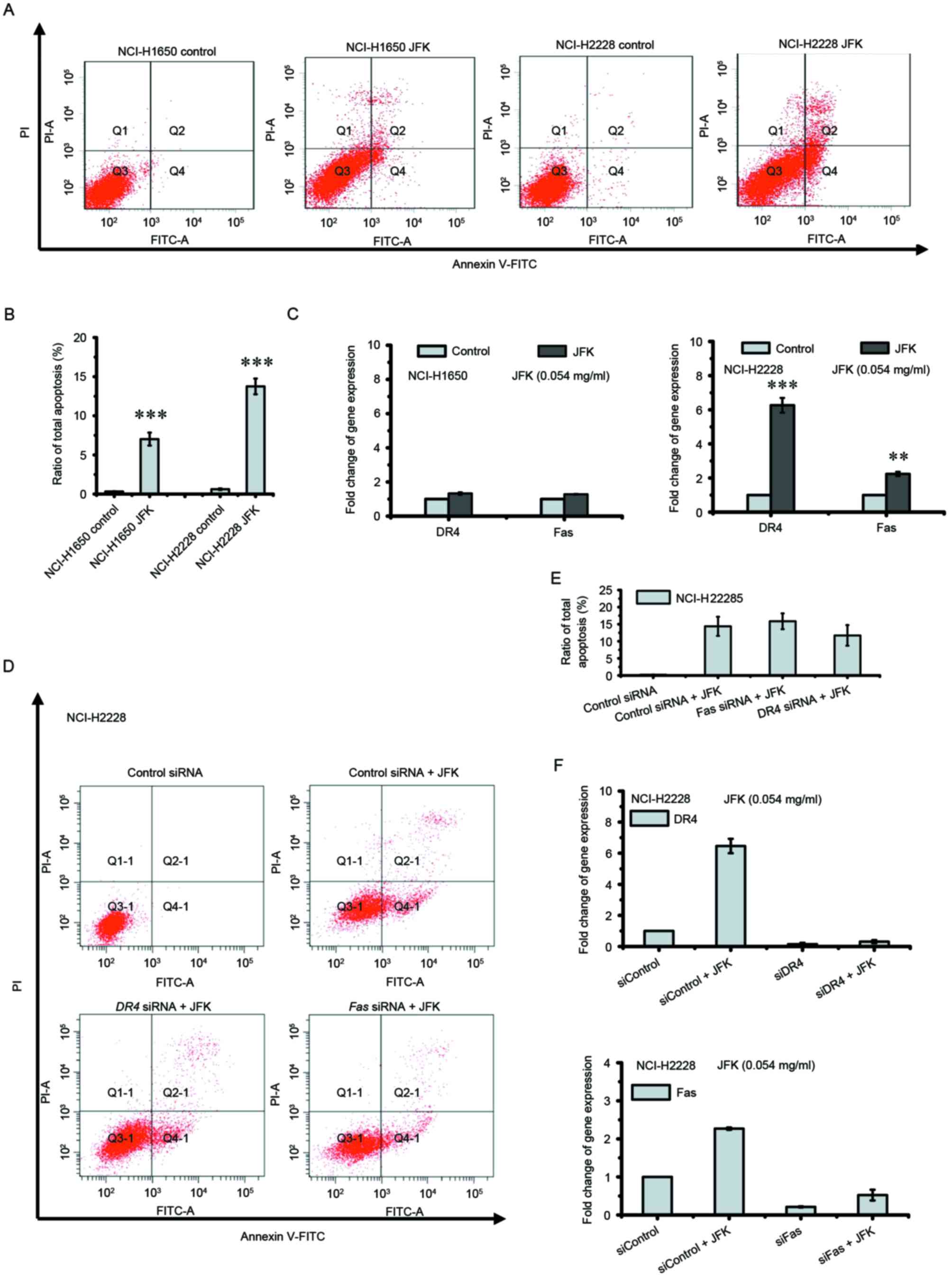 | Figure 5.DR4 and Fas are not
involved in JFK-induced apoptosis in NCI-H1650 and NCI-H2228 cells.
(A) The apoptotic cells were detected by flow cytometry, after JFK
(0.054 mg/ml) treatment for 48 h in NCI-H1650 and NCI-H2228 cells.
(B) Apoptosis rate was analyzed based on flow cytometry. Data were
expressed as the mean ± SD (n=3), ***P<0.001. (C) mRNA levels
were examined by reverse transcription-quantitative polymerase
chain reaction. Data were expressed as the mean ± SD (n=3),
**P<0.01 and ***P<0.001. (D and E) DR4 and
Fas-knockdown NCI-H2228 cells were treated with JFK (0.054
mg/ml) for 48 h, and then the percentage of apoptosis was
determined by flow cytometry. Data are expressed as the mean ± SD
(n=3). (F) DR4 and Fas knockdown NCI-H2228 cells were
treated with JFK (0.054 mg/ml) for 48 h, and then the mRNA levels
were examined by reverse transcription-quantitative polymerase
chain reaction. Data are expressed as the mean ± SD (n=3). SD,
standard deviation; DR4, death receptor 4; JFK, Jinfukang; siRNA,
small interfering RNA; PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
Overall, the present study reported that JFK induces
cellular growth arrest and apoptosis in lung cancer cells. The
transcriptome profiling analysis identified 5,281 genes whose
expression was modulated by JFK and it was demonstrated that JFK
contributes to A549 cellular apoptosis partially through activation
of DR4 and Fas. The present results provide new
insight for understanding how this TCM formula exerts its effects
against lung cancer.
Acknowledgements
The authors would like to thank Professor Daniel M.
Czajkowsky (School of Biomedical Engineering and Bio-ID Center,
Shanghai Jiao Tong University, Shanghai, China) for helpful
discussion and careful proofreading. The present study was
supported by Longhua Medical Project of State Clinical Research
Center of TCM in Longhua Hospital (grant nos. LYTD-21 and
JDZX2012123), National Basic Research Program of China (grant no.
2013CB967402) and National Natural Science Foundation of China
(grant nos. 81373623 and 91229123).
Glossary
Abbreviations
Abbreviations:
|
JFK
|
Jinfukang formula
|
|
NSCLC
|
non-small cell lung cancer
|
|
TCM
|
Traditional Chinese Medicine
|
|
RNA-Seq
|
RNA sequencing
|
|
CCK8
|
Cell Counting Kit 8
|
|
PI
|
propidium iodine
|
|
GO
|
gene ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
References
|
1
|
Li C and Hong W: Research status and
funding trends of lung cancer biomarkers. J Thorac Dis. 5:698–705.
2013.PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ettinger DS, Wood DE, Akerley W, Bazhenova
LA, Borghaei H, Camidge DR, Cheney RT, Chirieac LR, D'Amico TA,
Demmy TL, et al: Non-small cell lung cancer, version 1.2015. J Natl
Compr Canc Netw. 12:1738–1761. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shanker M, Willcutts D, Roth JA and Ramesh
R: Drug resistance in lung cancer. Lung Cancer (Auckl). 1:23–36.
2010.PubMed/NCBI
|
|
5
|
Efferth T, Li PC, Konkimalla VS and Kaina
B: From traditional Chinese medicine to rational cancer therapy.
Trends Mol Med. 13:353–361. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu J, Xu J and Shi Q: Effect of ethanol
extract of HPRS, a Traditional Chinese Medicine formula, on HCT116
cell line. J Tradit Chin Med. 36:760–767. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han R: Highlight on the studies of
anticancer drugs derived from plants in China. Stem Cells.
12:53–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiao L, Wang Y, Xu L and You M: Lung
cancer prevention and therapy using the JinFuKang herbal mixture.
Curr Pharmacol Rep. 1:346–353. 2015. View Article : Google Scholar
|
|
9
|
Sun JL and Liu JX: Effect of ‘Jinfukang
Oral Solution’ on expression of apoptosis related genes of human
lung adenocarcinoma cells transplanted in nude mice. Shanghai J
Tradit Chin Med. 10:2007.
|
|
10
|
Liu JX, Shi ZM, Li HG, Xu ZY, Zhu YW, Zhao
LH, Gao H, Liu LS, Zhu HR and Zhang H: Clinical observation on 271
cases of non-small cell lung cancer treated with Yifei Kangliu Yin.
Chin J Integr Tradit West Med. 7:247–250. 2001.
|
|
11
|
Xue R, Fang Z, Zhang M, Yi Z, Wen C and
Shi T: TCMID: Traditional Chinese medicine integrative database for
herb molecular mechanism analysis. Nucleic Acids Res. 41(Database
issue): D1089–D1095. 2013.PubMed/NCBI
|
|
12
|
Bonham M, Arnold H, Montgomery B and
Nelson PS: Molecular effects of the herbal compound PC-SPES:
Identification of activity pathways in prostate carcinoma. Cancer
Res. 62:3920–3924. 2002.PubMed/NCBI
|
|
13
|
Lu J, Chen J, Xu N, Wu J, Kang Y, Shen T,
Kong H, Ma C, Cheng M, Shao Z, et al: : Activation of AIFM2
enhances apoptosis of human lung cancer cells undergoing
toxicological stress. Toxicol Lett. 258:227–236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pickrell JK, Marioni JC, Pai AA, Degner
JF, Engelhardt BE, Nkadori E, Veyrieras JB, Stephens M, Gilad Y and
Pritchard JK: Understanding mechanisms underlying human gene
expression variation with RNA sequencing. Nature. 464:768–772.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Z, Gerstein M and Snyder M: RNA-Seq:
A revolutionary tool for transcriptomics. Nat Rev Genet. 10:57–63.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu J, Zhang X, Shen T, Ma C, Wu J, Kong H,
Tian J, Shao Z, Zhao X and Xu L: Epigenetic profiling of H3K4Me3
reveals herbal medicine Jinfukang-induced epigenetic alteration is
involved in anti-lung cancer activity. Evid Based Complement
Alternat Med. 2016:132016. View Article : Google Scholar
|
|
17
|
Qi Y, Zhang X, Kang Y, Wu J, Chen J, Li H,
Guo Y, Liu B, Shao Z and Zhao X: Genome-wide transcriptional
profiling analysis reveals annexin A6 as a novel EZH2 target gene
involving gastric cellular proliferation. Mol Biosyst.
11:1980–1986. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Trapnell C, Roberts A, Goff L, Pertea G,
Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL and Pachter L:
Differential gene and transcript expression analysis of RNA-seq
experiments with TopHat and Cufflinks. Nat Protoc. 7:562–578. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hesketh T and Zhu WX: Health in China.
Traditional Chinese medicine: One country, two systems. BMJ.
315:115–117. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cassileth BR, Rizvi N, Deng G, Yeung KS,
Vickers A, Guillen S, Woo D, Coleton M and Kris MG: Safety and
pharmacokinetic trial of docetaxel plus an Astragalus-based herbal
formula for non-small cell lung cancer patients. Cancer Chemother
Pharmacol. 65:67–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nigg EA: Cyclin-dependent protein kinases:
Key regulators of the eukaryotic cell cycle. Bioessays. 17:471–480.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chohan TA, Qian H, Pan Y and Chen JZ:
Cyclin-dependent kinase-2 as a target for cancer therapy: Progress
in the development of CDK2 inhibitors as anti-cancer agents. Curr
Med Chem. 22:237–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Volm M, Koomägi R, Mattern J and Stammler
G: Cyclin A is associated with an unfavourable outcome in patients
with non-small-cell lung carcinomas. Br J Cancer. 75:1774–1778.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Soria JC, Jang SJ, Khuri FR, Hassan K, Liu
D, Hong WK and Mao L: Overexpression of cyclin B1 in early-stage
non-small cell lung cancer and its clinical implication. Cancer
Res. 60:4000–4004. 2000.PubMed/NCBI
|
|
29
|
Ouyang L, Shi Z, Zhao S, Wang FT, Zhou TT,
Liu B and Bao JK: Programmed cell death pathways in cancer: A
review of apoptosis, autophagy and programmed necrosis. Cell
Prolif. 45:487–498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oh SB, Hwang CJ, Song SY, Jung YY, Yun HM,
Sok CH, Sung HC, Yi JM, Park DH, Ham YW, et al: Anti-cancer effect
of tectochrysin in NSCLC cells through overexpression of death
receptor and inactivation of STAT3. Cancer Lett. 353:95–103. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi S, Kajino S, Takahashi N,
Kanazawa S, Imai K, Hibi Y, Ohara H, Itoh M and Okamoto T: 53BP2
induces apoptosis through the mitochondrial death pathway. Genes
Cells. 10:253–260. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fei P, Wang W, Kim Sh, Wang S, Burns TF,
Sax JK, Buzzai M, Dicker DT, McKenna WG, Bernhard EJ and El-Deiry
WS: Bnip3L is induced by p53 under hypoxia, and its knockdown
promotes tumor growth. Cancer Cell. 6:597–609. 2004. View Article : Google Scholar : PubMed/NCBI
|