Introduction
Breast cancer (BC) is one of the top five malignant
cancer types in women worldwide in 2017 (1). According to the World Health
Organization, BC resulted in 521,000 mortalities in 2012 (2); additionally, ~1.7 million cases are
diagnosed every year (3,4), only 1% of which were diagnosed in males
(5). The most common primary symptom
of BC is a painless lump, and the diagnosis may be confirmed by
biopsy (5). BC is the most frequently
occurring cancer in women of reproductive age (15–49 years)
(6). In addition, genetic factors
account for 5–10% of all primary BC cases (7). Women with either a mother or sister
diagnosed with BC prior to 50 years of age have a 1.7% higher risk
compared with patients with no family history of BC (8). Prognoses of BC cases are different and
depend on a number of factors; therefore, outcomes vary between
different cases. Generally, the later the disease stage at point of
diagnosis, the poorer the prognosis (5). Therefore, it is important to understand
the process of BC pathogenesis.
MicroRNAs (miRNAs/miR) are important in activating
and modulating gene expression in a number of critical cellular
processes (7). They are small,
ubiquitous, endogenous single-chain molecules, 21–25 nucleotides in
length, and are non-protein coding. They may inhibit the
translation of the target mRNA, or reduce the activity of mRNA
through the complete or incomplete complementary binding to the
3′-untranslated region of the target gene and forming a complex
then inducing silence of the target gene (9). This may reflect a mechanistic potential,
enabling the bound complexes to avoid the mRNA clearing activity of
the ribosome (10). Several studies
have indicated that differential miRNAs expression in cancer
tissues is associated with human BC: For example, the downregulated
miRNAs miR-125b, miR-145, miR-21 and miR-155 were closely
associated with the biopathological features of different cancer
types (11). Overexpressed
has-miR-10b is also significantly associated with BC; it promotes
cancer cell migration and invasion (12).
Although a series of studies were conducted on the
miRNAs of BC, the results of the analysis of miRNAs expression
datasets of BC in previous studies remain inconsistent (11–13). These
inconsistencies may be due to a number of factors, including the
type of sequencing platform, sample selection, study design and
exposure assessment (13). By
analyzing the key miRNAs in the pathogenesis of BC, the aim of the
present study was to explore the association between miRNA and the
prevention and treatment of BC.
In the present study, a systematic review of the
miRNAs in BC identified in previous studies was conducted; the
function of the target genes regulated by the key miRNAs was also
analyzed to clarify the potential associations between the miRNAs
and the risk of BC. Specifically, results from the prediction of
the target genes of a number of BC-associated miRNAs were analyzed.
In addition, the target genes and corresponding transcription
factors (TF) associated with differentially expressed miRNAs were
identified, in order to gain an improved understanding of the
interaction of target genes and TF to the differentially expressed
miRNAs in BC. Additional studies are required to apply these key
miRNAs as biomarkers in BC diagnosis and site-specific
treatment.
Materials and methods
Identification of BC miRNA datasets
and screening of differentially expressed miRNAs
‘Breast Cancer’ and ‘miRNA’ were set as keywords to
screen previously published studies on Google Scholar (https://scholar.google.com/; date of assess, June 25th
2018). Following careful reading of original articles from the
records, a total of eight datasets that contained the expression of
miRNAs between BC and control tissues were determined. Then, the
information concerning all differentially expressed miRNAs in BC
was obtained. Subsequently, the details of up- and downregulated
miRNAs in BC, as compared with the control normal samples, were
also obtained from the reports of associated articles. The
frequencies of the up- and downregulated miRNAs in separate
datasets were calculated, and the miRNAs that appeared in at least
3 datasets were selected in the present study.
Indication of target genes of
differential expressed miRNA in BC
The sequence information of differentially expressed
miRNAs was obtained from the miRBase database (http://www.mirbase.org; version 21; June 1st 2017)
(14,15); then this sequence information was
transferred to a FASTA file (https://molbiol-tools.ca/Convert.htm). Using
TargetScan software (version 6.0; http://www.targetscan.org/mamm_31) (16,17), the
target genes associated with differentially expressed miRNA were
indicated. The parameter for maximum number of target genes for
each miRNA was set at 300, while the maximum non-target genes for
each miRNA were excluded.
Functional enrichment analysis of
target genes of differentially expressed miRNAs in BC
The predicted target genes of the differentially
expressed miRNAs were mapped into the Database for Annotation,
Visualization and Integrated Discovery (DAVID) (18) to perform the Gene Ontology (GO)
(19) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) pathway (20)
analyses. Then, the top 10 GO terms of the ‘biological processes’
from the GO annotation results were screened out, identified as
those with P<0.05.
TF analysis of target genes regulated
by up- and downregulated miRNA in BC
The up- and downregulated target genes of BC miRNAs
were uploaded to the TFactS database (21,22) at
(http://www.tfacts.org). The TFs were predicted
using the P-value, q-value, E-value and false discovery rate
(http://www.tfacts.org/); only TFs that were
<0.05 for all 4 indexes were considered to be reliable. The TFs
of target genes, which were mediated by up- and downregulated
miRNAs, were counted separately. Subsequently, the number of shared
TFs and different TFs of the target genes regulated by the up- and
downregulated miRNAs were calculated.
Results
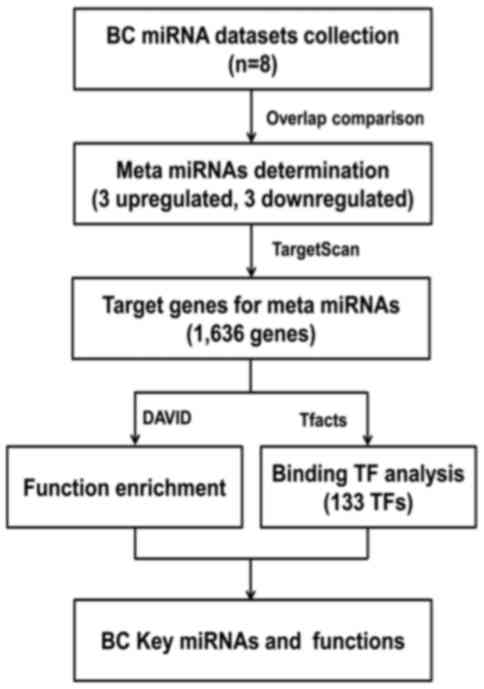
Method of screening
To analyze the key miRNAs and their target genes in
BC pathogenesis, and to explore the pathogenesis of BC, a
systematic review was performed to first identify the key up- and
downregulated miRNAs in BC. The target genes were predicted, and
functional and TF analyses of the target genes were subsequently
performed. The process is summarized in Fig. 1.
miRNAs identified through
screening
A total of 8 independent miRNA expression datasets
were identified from previous studies into miRNA expression
profiles in BC, which provided details of miRNA profiles in BC
tissues compared with in control tissues. In the present study, the
8 BC miRNA datasets were named based on the initials of the
authors, as follows: i) SY (23); ii)
IM (11); iii) VS (24); iv) HH (25); v) LA (26); vi) KL (27); vii) YL (28); and viii) SH (29). The primary features of the 8 datasets
used in the analysis are summarized in Table I; these 8 datasets differed according
to the sample type, number of samples, chip types and the number of
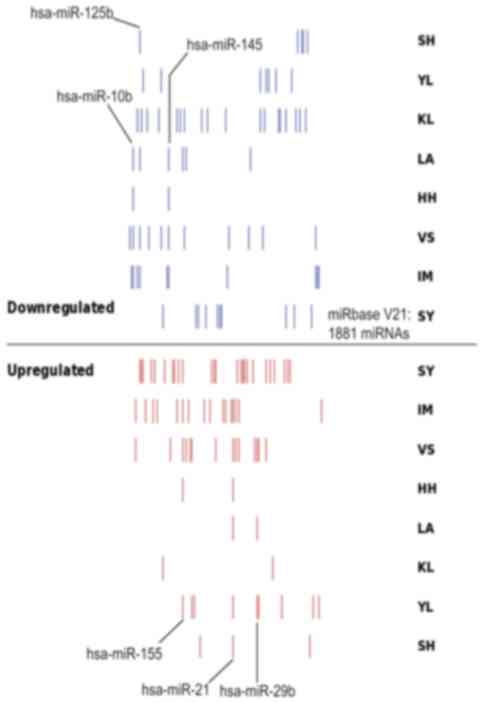
probes on the chip. By using SVG mapping, the distribution of the
differentially expressed miRNAs in the 8 datasets was analyzed, and
it was identified that there were notable differences among the up-
and downregulated miRNAs in each database (Fig. 2). As depicted in Fig. 2, the count and composition of
differentially expressed miRNAs exhibited notable differences
between different datasets, and only a limited number of overlapped
miRNAs were determined between datasets. There were also notable
differences among the differentially expressed miRNAs in the
distinct datasets. A total of 99 non-redundant differential miRNAs
were identified in the 8 BC miRNA datasets. Table II indicates the number of
differentially expressed miRNAs, and the results of the statistical
analysis of the differentially expressed miRNAs identified in the 8
BC miRNA datasets.
 | Table I.Basic characteristics of the breast
cancer datasets. |
Table I.
Basic characteristics of the breast
cancer datasets.
| Dataset | Sample type | Assay type | miRNA probes,
n | Method of
validation |
|---|
| SY (23) | Three human breast
tumors | Multiplex Real-time
PCR Assay (Applied Biosystems; Thermo Fisher Scientific, Inc.) | b- | Breast cancer cell
colony formation assay |
| IM (11) | 10 normal and 76
neoplastic breast tissues | miRNA microarray
chip (KCI version 1.0) and Perkin-Elmer ScanArray XL5K | 368 | Northern
blotting |
| HH (25) | 148 patients with
BC and 44 age-matched and disease-free controls | TaqMan (Applied
Biosystems; Thermo Fisher Scientific, Inc.) | – | RT-qPCR |
| LA (26) | – | – | – | – |
| KL (27) | MCF-7 and
MDA-MB-231 cell lines | RT2 miRNA PCR array
system (SABiosciences™ Corporation, Frederick, MD, USA) | 376 | RT-qPCR |
| YL (28) | 113 BC and 40
paired non-cancerous NATs | CapitalBio
(http://www.capitalbio.com) | 509 | RT-qPCR |
| SH (29) | 48 tissue and 100
serum samples of patients with primary BC and 20 control
samples | – | 10 | RT-qPCR |
| VS (40) | 63 primary tumors
and 177 normal tissues | Bead-based microRNA
profiling platform | 40-mer | Northern
blotting |
| Table II.GO annotation results of the target
genes of the up- and downregulated microRNAs in breast cancer
samples. |
Table II.
GO annotation results of the target
genes of the up- and downregulated microRNAs in breast cancer
samples.
| GO Term | Target gene
count | P-value | Fold
enrichment |
|---|
|
|---|
| A, Upregulated |
|---|
| GO:0045944:
Positive regulation of transcription from RNA polymerase II
promoter | 93 |
1.09×10−12 | 2.19 |
| GO:0006366:
Transcription from RNA polymerase II promoter | 59 |
1.79×10−11 | 2.66 |
| GO:0000122:
Negative regulation of transcription from RNA polymerase II
promoter | 67 |
5.70×10−9 | 2.15 |
| GO:0045893:
Positive regulation of transcription, DNA-templated | 50 |
1.85×10−7 | 2.25 |
| GO:0045669:
Positive regulation of osteoblast differentiation | 14 |
1.29×10−6 | 5.40 |
| GO:0006351:
Transcription, DNA-templated | 127 |
2.03×10−6 | 1.50 |
| GO:0001525:
Angiogenesis | 27 |
4.15×10−6 | 2.80 |
| GO:0008284:
Positive regulation of cell proliferation | 43 |
5.60×10−6 | 2.13 |
| GO:0021542: Dentate
gyrus development | 7 |
5.22×10−5 | 9.52 |
| GO:0006355:
Regulation of transcription, DNA-templated | 97 |
5.85×10−5 | 1.49 |
|
| B,
Downregulated |
|
| GO:0045944:
Positive regulation of transcription from RNA polymerase II
promoter | 81 |
5.22×10−8 | 1.88 |
| GO:0048013: Ephrin
receptor signaling pathway | 18 |
1.61×10−7 | 4.76 |
| GO:0006351:
Transcription, DNA-templated | 130 |
1.06×10−6 | 1.51 |
| GO:0007411: Axon
guidance | 21 |
2.19×10−5 | 3.01 |
| GO:0000122:
Negative regulation of transcription from RNA polymerase II
promoter | 55 |
8.54×10−5 | 1.74 |
| GO:0006366:
Transcription from RNA polymerase II promoter | 42 |
1.60×10−4 | 1.86 |
| GO:0045821:
Positive regulation of glycolytic process | 6 |
2.31×10−4 | 9.75 |
| GO:0045893:
Positive regulation of transcription, DNA-templated | 41 |
3.40×10−4 | 1.81 |
| GO:0006357:
Regulation of transcription from RNA polymerase II promoter | 36 |
5.37×10−4 | 1.86 |
| GO:0043524:
Negative regulation of neuron apoptotic process | 16 |
6.83×10−4 | 2.76 |
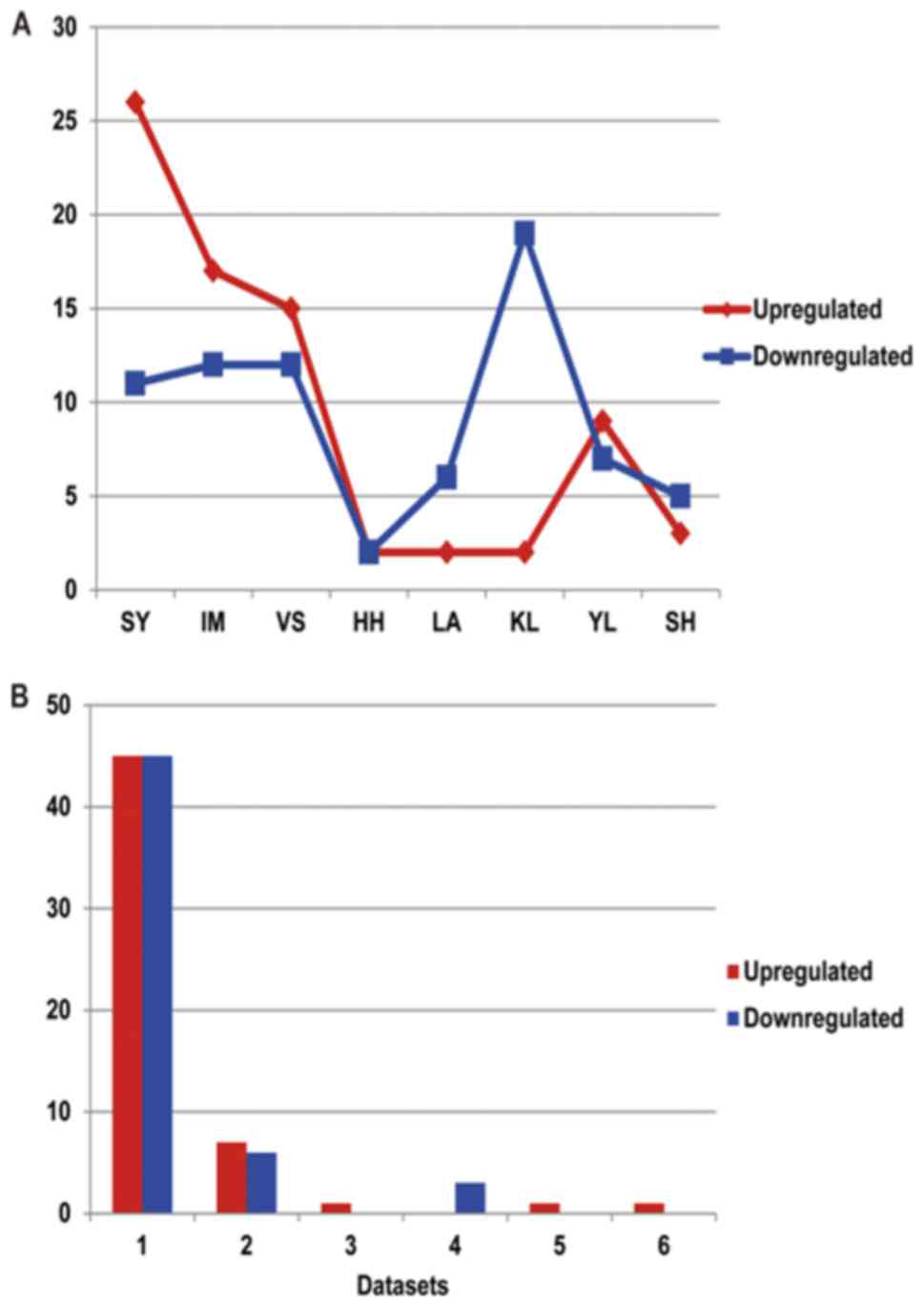
There were 4 datasets that had >20 differentially
expressed miRNAs: Dataset SY, 37; dataset IM, 29; dataset VS, 27;
and dataset KL, 21. Among the upregulated miRNAs, the largest
number for one dataset was 26 (SY), followed by dataset IM
(17); the HH, LA and KL datasets all
had the lowest numbers (all 2). Among the downregulated miRNAs, the
largest number for one dataset was 19 (KL), followed by the IM
(17) and VS (12) datasets; the HH dataset had the lowest
number (2).
Screening and identification of
differentially expressed miRNAs
Data from the differentially expressed miRNAs,
compared with the control samples, were collected from the previous
studies, as presented in Fig. 3. It
was identified that 99 non-redundant miRNAs were identified from
the 8 independent data sets, 55 of which were upregulated and 54 of
which were downregulated. This also indicates the requirement for a
systematic review of differential miRNA expression datasets in
BC.
The miRNAs expressed in at least 3 databases were
selected as the most likely to be correct miRNAs for subsequent
analysis. Consequently, a total of 6 meta miRNAs were identified in
the differential miRNA datasets of BC. The results indicated that a
total of 3 meta miRNAs were upregulated compared with the normal
tissues, which were hsa-miR-21b, hsa-miR-29b and hsa-miR-155, and a
total of 3 meta miRNAs were downregulated compared with the normal
tissues, which were hsa-miR-10b, hsa-miR-125b and hsa-miR-145.
These six were considered to be the most likely to be correct
differentially expressed miRNAs, and key factors affecting the
pathogenesis of BC.
In addition, information regarding miRNA location
was collected from the miRBase database, and the specific
differential miRNA expression profile data are summarized in
Table II. The chromosomal
distribution of these six meta miRNAs are relatively dispersed.
Specifically, two meta miRNAs (hsa-miR-155, hsa-miR-125b) were
located on chromosome 21, and the other four were located
separately on 4 different chromosomes: Chromosome 1 (hsa-miR-29b);
chromosome 2 (hsa-miR-10b); chromosome 5 (hsa-miR-145); and
chromosome 17 (hsa-miR-21). This sporadic distribution indicates
that the key miRNAs involved in the development of BC may have
large differences in biological function (30).
Prediction of target genes in
differentially expressed miRNAs
The target genes of the six meta-miRNAs (summarized
in Table II) were predicted using
TargetScan software. Although the default parameters were used, the
prediction results varied widely. In order to accurately identify
the target genes regulated by differentially expressed miRNAs, the
maximum number of target genes was set as 300. According to the
results, the number of target genes of all six differentially
expressed miRNAs reached 300.
Functional analysis results of target
genes of differentially expressed miRNAs
The metabolic pathway enrichment results of each of
the target genes regulated by meta miRNAs were obtained, which are
presented in Fig. 4. According to
this figure, the KEGG Pathway functional enrichment results were
identified in all six differentially expressed miRNAs, but the
enrichment levels differed. Of these six miRNAs, the target genes
of hsa-miR-155 had the widest range of pathway enrichment results,
involving 26 KEGG terms; followed by hsa-miR-21, involving 24 KEGG
terms; then hsa-miR-145 (17 KEGG terms), hsa-miR-10b (11 KEGG
terms), and hsa-miR-125b (2 KEGG terms); and hsa-miR-29b (1
term).
The target genes identified to be associated with
the up- and downregulated miRNAs were uploaded to the DAVID website
for functional annotation analysis. Then, the GO enrichment results
of the most significant 10 biological processes (Biology Process)
corresponding to the target genes regulated by the up- and
downregulated miRNAs were collected, and the Pathway results with a
P<0.05 were identified. As summarized in Table III, upregulated miRNAs in BC were
predominantly enriched in the transcriptional regulation process.
Respectively, they depend on the RNA polymerase II promoter (GO:
0045944: Positive regulation of transcription from RNA polymerase
II promoter; GO: 000636: Transcription from RNA polymerase II
promoter; and GO: 0006366: Transcription from RNA polymerase II
promoter) and DNA template (GO: 0045893: Positive regulation of
transcription, DNA-templated). The target genes of downregulated
miRNAs in BC were enriched in the transcriptional regulation
process (GO: 0045944: Positive regulation of transcription from RNA
polymerase II promoter; GO: 0006351: Transcription, DNA-templated;
GO: 0000122: Negative regulation of transcription from RNA
polymerase II promoter; GO: 0045893: Positive regulation of
transcription, DNA-templated; and GO: 0006357: Regulation of
transcription from RNA polymerase II promoter). Additionally, they
were specifically enriched in the ephrin receptor signaling pathway
(GO: 0048013: ephrin receptor signaling pathway) and the biological
process of axon guidance (GO: 0007411: Axon guidance).
| Table III.List of key meta miRNAs in breast
cancer. |
Table III.
List of key meta miRNAs in breast
cancer.
| miRNA | Number of supported
datasets | Chr no. | Beg | End | Strand sense | Sequence |
|---|
|
|---|
| A, Upregulated |
|---|
| hsa-miR-21 | 6 | 17 | 59841273 | 59841294 | + |
UAGCUUAUCAGACUGAUGUUGA |
| hsa-miR-29b | 3 | 1 | 207802450 | 207802472 | – |
UAGCACCAUUUGAAAUCAGUGUU |
| hsa-miR-155 | 5 | 21 | 25573983 | 25574005 | + |
UUAAUGCUAAUCGUGAUAGGGGU |
|
| B,
Downregulated |
|
| hsa-miR-10b | 4 | 2 | 176150329 | 176150351 | + |
UACCCUGUAGAACCGAAUUUGUG |
| hsa-miR-125b | 4 | 21 | 16590290 | 16590311 | + |
UCACAAGUCAGGCUCUUGGGAC |
| hsa-miR-145 | 4 | 5 | 149430661 | 149430683 | + |
GUCCAGUUUUCCCAGGAAUCCCU |
The results of KEGG metabolic pathway enrichment
(Table IV) indicated that the target
genes of the upregulated miRNAs were specifically enriched in
cancer pathways: (hsa05200: Pathways in cancer) and several
signaling pathways [hsa04010: Mitogen-activated protein kinase
(MAPK) signaling pathway; hsa04014: Ras signaling pathway;
hsa04550: Signaling pathways regulating pluripotency of stem cells;
and hsa04151: Phosphoinositide 3-kinase/RAC-alpha serine/threonine
protein kinase signaling pathway]. Compared with this, the range
enrichment pathways of target genes in the downregulated miRNAs was
relatively varied, including: Proteoglycans in cancer (hsa05205);
axon guidance (hsa04360); circadian rhythm (hsa04710);
transcriptional misregulation in cancer (hsa05202); and MAPK
signaling pathway (hsa04010). This result further proved the
significance of axon guidance (hsa04360) in the pathogenesis of BC
and complexity of the pathogenesis of BC caused by multiple
metabolic pathways.
| Table IV.Kyoto Encyclopedia of Genes and
Genomes metabolic pathway enrichment results of the upregulated
miRNAs target genes in breast cancer samples. |
Table IV.
Kyoto Encyclopedia of Genes and
Genomes metabolic pathway enrichment results of the upregulated
miRNAs target genes in breast cancer samples.
| A, Upregulated
miRNAs |
|---|
|
|---|
| Term | Target gene
count | P-value | Fold
enrichment |
|---|
| hsa05200: Pathways
in cancer | 39 |
1.93×10−6 | 2.29 |
| hsa04010: MAPK
signaling pathway | 27 |
3.66×10−5 | 2.44 |
| hsa04014: Ras
signaling pathway | 25 |
3.81×10−5 | 2.55 |
| hsa04550: Signaling
pathways regulating pluripotency of stem cells | 17 |
3.33×10−4 | 2.80 |
| hsa04151: PI3K-Akt
signaling pathway | 30 |
4.16×10−4 | 2.00 |
|
| B, Downregulated
miRNAs |
|
| hsa05205:
Proteoglycans in cancer | 24 |
1.17×10−5 | 2.81 |
| hsa04360: Axon
guidance | 17 |
8.68×10−5 | 3.14 |
| hsa04710: Circadian
rhythm | 8 |
2.56×10−4 | 6.04 |
| hsa05202:
Transcriptional misregulation in cancer | 18 |
7.52×10−4 | 2.51 |
| hsa04010: MAPK
signaling pathway | 23 |
1.19×10−3 | 2.11 |
Transcription factor analysis of
target genes regulated by differentially expressed miRNAs
In the present study, the TFs corresponding to the
target genes of up- and downregulated miRNAs (Table II) were also analyzed, and the
similarities and differences between the TFs of the two types of
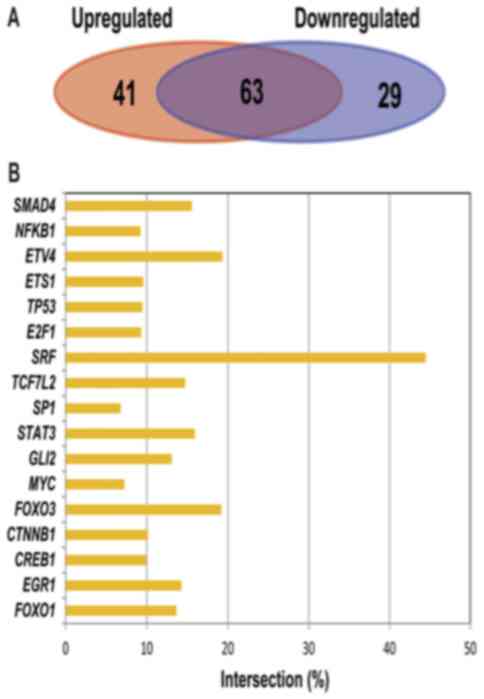
target genes identified. As indicated in Fig. 5A, 104 TFs interacted with 319 target
genes of the upregulated miRNAs, while 92 TFs interacted with 254
target genes of the downregulated miRNAs. According to the
similarities and differences of TFs between the two types of target
genes, demonstrated in Fig. 5, it was
identified that there were 63 TFs shared by the two types of target
genes, accounting for 47.3% of the total numbers of TFs. Fig. 5B indicated the crossover ratio, which
is the most significant TF. According to this figure, the E-value
was <0.05, and the highest crossover ratio value of
transcription factors (SRF) was 42%, which indicated that the
results of identified transcription factors were relatively
reliable.
Discussion
In the present systematic review, the results of
previous studies investigating the association between key miRNAs
and the progression of BC were analyzed. A total of six key miRNAs
were identified. The purpose of the present study was to analyze
the roles of miRNA in BC that may affect the prognosis of BC, then
to perform prognostic risk classification on patients, or identify
novel therapeutic methods for BC.
Upregulated hsa-miR-21 was identified as closely
associated with BC in the present study. In previous studies,
upregulated hsa-miR-21 has been demonstrated to be associated with
neuroblastoma (31) and lung cancer
(30). It was identified that the
consistent overexpression of hsa-miR-21 in tumors indicates
oncogenic activity, and may be classed as an oncomiR (31). These studies indicate that hsa-miR-21
may act as a universal indicator of cancer, instead of only for BC.
Patients with lung cancer and high hsa-miR-155 expression levels
exhibited poorer survival rates, compared with those with low
hsa-miR-155 expression levels (30).
It was also demonstrated that the role of hsa-miR-155 in BC
involved the promotion of tumor angiogenesis through targeting the
von Hippel-Lindau pathway, and was closely associated with poor
prognosis (32). Hsa-miR-29 is a
tumor promotor, which promotes metastasis in breast and colon
cancer (33). These data suggest that
miRNAs are important diagnostic biomarkers for different types of
cancer.
Of the three downregulated meta-miRNAs, hsa-miR-145
was identified to consistently exhibit decreased levels at certain
cancer stages of colorectal neoplasia (34). It was revealed previously that the
decreased expression of has-miR-10b may lead to tumor invasion and
metastasis through suppressing homeobox D10 and indirectly
activating the pro-metastatic gene Ras homolog family member C
(12). Hsa-miR-125b has been
demonstrated to function as a tumor suppressor in breast (35), bladder (36) and ovarian cancer (37). It may decrease expression of the
Mucin-1 oncoprotein, thereby promoting DNA damage-induced apoptosis
(38), suggesting that this miRNA is
also an important biomarker for cancer. All the six key miRNAs
identified in the present study have been demonstrated to be
closely associated with the progress of not only BC, but also a
range of cancer types. While five miRNAs indicated consistent roles
in different types of cancer, hsa-miR-29 exhibited various
functions (33). This may also be a
reason for the inconsistent outcomes of biomarker-associated miRNAs
in different miRNA studies (35).
Hsa-miR-29 and hsa-miR-125b were regarded as novel diagnostic and
prognostic biomarkers (33,38).
The KEGG pathway functional enrichment results were
calculated for all six differentially expressed miRNAs. The three
upregulated BC miRNAs were significantly enriched in the
transcriptional regulation process, in particular the progress of
RNA polymerase II promoter and DNA template. The target genes of
the downregulated BC miRNAs were also enriched in the
transcriptional regulation process. This may indicate that these
biological transcriptional regulation processes serve a vital role
in the pathogenesis of BC. Additionally, the three downregulated
miRNAs were specifically enriched in the ephrin receptor signaling
pathway (GO: 0048013) and the biological process of axon guidance
(GO: 0007411). It has been suggested previously that ephrin
receptors (Eph) and ephrins serve as tumor suppressors and
negatively regulate tumor growth (39). EphB6 is downregulated in invasive and
metastatic BC (39). The roles of
Ephs and ephrins are multi-faceted; the up- or downregulated ephrin
receptor signaling pathway also indicates a positive and negative
effect on the metastasis of BC (24).
Additional studies are required to explore the complex function of
this entire process. The axon guidance molecules, including ephrin,
are important for cell proliferation and the adhesion of normal
tissue (40). The downregulated
process of axon guidance in the present study indicated that it
serves as a tumor suppressor in BC (40). It has potential to serve as a
biomarker in BC diagnosis and targeted therapeutic strategies.
Early diagnoses are vital to decreasing the
mortality rate of cancer, and specific biomarkers are important for
diagnosis, disease prediction and prognosis. Although there is
evidence that miRNAs have critical effects on the pathology of BC,
these remain incompletely characterized.
In the present study, a systematic review identified
six miRNAs and the vital KEGG pathways, which suggests the
potential of selecting specific and typical biomarkers for BC, as
well as using the pathways and GO functions to further explore
effective therapeutics strategies.
In conclusion, miRNAs are important in the
development and progression of cancer. In the present study, 6 key
miRNAs involved in the pathological process of BC were identified,
including the regulated target genes and TFs. The systematic review
results indicated the potential for the prediction of BC prognosis
and metastasis. Additional exploration of the differential
expression of miRNAs and their associated pathways may provide
insight into additional useful biomarkers for the diagnosis and
prognosis of BC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FL, XZ and NT conceived and designed the study. MY,
JF, YP and JL performed the data analysis and interpretation of
data. FL and XZ wrote the paper. MY, JF and JL reviewed and edited
the manuscript. All authors read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare there are no competing
interests.
References
|
1
|
DeSantis CE, Ma J, Goding Sauer A, Newman
LA and Jemal A: Breast cancer statistics, 2017, racial disparity in
mortality by state. CA Cancer J Clin. 67:439–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nowbar AN, Howard JP, Finegold JA, Asaria
P and Francis DP: 2014 global geographic analysis of mortality from
ischaemic heart disease by country, age and income: Statistics from
world health organisation and united nations. Int J Cardiol.
174:293–298. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stewart BW and Wild CP: The global and
regional burden of cancerWorld Cancer Report 2014. World Health
Organization; Geneva: pp. 162014
|
|
4
|
Ferlay J, Soerjomataram I, Ervik M,
Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D and
Bray F: GLOBOCAN 2012 Cancer Incidence and Mortality Worldwide:
IARC CancerBase No. 11 [Internet]. v1.0. International Agency for
Research on Cancer; Lyon, France: 2013 Decembet 12–2016
|
|
5
|
Kosir MA: Breast Cancer. MSD manual
consumer version. https://www.msdmanuals.com/home/women-s-health-issues/breast-disorders/breast-cancer%C2%A0Decembet
12–2016
|
|
6
|
Reeder JG and Vogel VG: Breast cancer
prevention. Cancer Treatment and Res. 141:149–164. 2008. View Article : Google Scholar
|
|
7
|
Gage M, Wattendorf D and Henry LR:
Translational advances regarding hereditary breast cancer
syndromes. J Surg Oncol. 105:444–451. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Colditz GA, Kaphingst KA, Hankinson SE and
Rosner B: Family history and risk of breast cancer: Nurses' health
study. Breast Cancer Res Treat. 133:1097–1104. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dalmay T: Mechanism of miRNA-mediated
repression of mRNA translation. Essays Biochem. 54:29–38. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang C, Xie SH, Xu B, Lu S and Liu P:
Digitalis use and the risk of breast cancer: A systematic review
and meta-analysis. Drug Saf. 40:285–292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Griffiths-Jones S, Saini HK, van Dongen S
and Enright AJ: miRBase: Tools for microRNA genomics. Nucleic Acids
Res. 36:D154–D158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kozomara A and Griffiths-Jones S: miRBase:
Integrating microRNA annotation and deep-sequencing data. Nucleic
Acids Res. 39:152–157. 2011. View Article : Google Scholar
|
|
16
|
Lewis BP, Shih Ih, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dennis G, Sherman BT, Hosack DA, Yang J,
Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. Nat
Genet. 25:25–29. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanehisa M, Goto S, Kawashima S, Okuno Y
and Hattori M: The KEGG resource for deciphering the genome.
Nucleic Acids Res. 32:D277–D280. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Essaghir A, Toffalini F, Knoops L, Kallin
A, van Helden J and Demoulin JB: Transcription factor regulation
can be accurately predicted from the presence of target gene
signatures in microarray gene expression data. Nucleic Acids Res.
38:e1202010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Essaghir A and Demoulin JB: A minimal
connected network of transcription factors regulated in human
tumors and its application to the quest for universal cancer
biomarkers. PLoS One. 7:e396662012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shimono Y, Zabala M, Cho RW, Lobo N,
Dalerba P, Qian D, Diehn M, Liu H, Panula SP, Chiao E, et al:
Downregulation of miRNA-200c links breast cancer stem cells with
normal stem cells. Cell. 138:592–603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:pp. 2257–2261.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heneghan HM, Miller N, Lowery AJ, Sweeney
KJ, Newell J and Kerin MJ: Circulating microRNAs as novel minimally
invasive biomarkers for breast cancer. Ann Surg. 251:499–505. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lowery AJ, Miller N, McNeill RE and Kerin
MJ: MicroRNAs as prognostic indicators and therapeutic targets:
Potential effect on breast cancer management. Clin Cancer Res.
14:360–365. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kastl L, Brown I and Schofield AC:
miRNA-34a is associated with docetaxel resistance in human breast
cancer cells. Breast Cancer Res Treat. 131:445–454. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan LX, Huang XF, Shao Q, Huang MY, Deng
L, Wu QL, Zeng YX and Shao JY: MicroRNA miR-21 overexpression in
human breast cancer is associated with advanced clinical stage,
lymph node metastasis and patient poor prognosis. RNA.
14:2348–2360. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Si H, Sun X, Chen Y, Cao Y, Chen S, Wang H
and Hu C: Circulating microRNA-92a and microRNA-21 as novel
minimally invasive biomarkers for primary breast cancer. J Cancer
Res Clin Oncol. 139:223–229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yanaihara N, Caplen N, Bowman E, Seike M,
Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, Tanaka T, et
al: Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Selcuklu SD, Donoghue MT and Spillane C:
miR-21 as a key regulator of oncogenic processes. Biochem Soc
Trans. 37:918–925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kong W, He L, Richards EJ, Challa S, Xu
CX, Permuth-Wey J, Lancaster JM, Coppola D, Sellers TA, Djeu JY and
Cheng JQ: Upregulation of miRNA-155 promotes tumour angiogenesis by
targeting VHL and is associated with poor prognosis and
triple-negative breast cancer. Oncogene. 33:679–689. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang H, Zhang G, Wu JH and Jiang CP:
Diverse roles of miR-29 in cancer (review). Oncol Rep.
31:1509–1516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Michael MZ, O'Connor SM, van Holst
Pellekaan NG, Young GP and James RJ: Reduced accumulation of
specific MicroRNAs in colorectal neoplasia. Mol Cancer Res.
1:882–891. 2003.PubMed/NCBI
|
|
35
|
Sun YM, Lin KY and Chen YQ: Diverse
functions of miR-125 family in different cell contexts. J Hematol
Oncol. 6:62013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang L, Luo J, Cai Q, Pan Q, Zeng H, Guo
Z, Dong W, Huang J and Lin T: MicroRNA-125b suppresses the
development of bladder cancer by targeting E2F3. Int J Cancer.
128:1758–1769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cowden Dahl KD, Dahl R, Kruichak JN and
Hudson LG: The epidermal growth factor receptor responsive miR-125a
represses mesenchymal morphology in ovarian cancer cells.
Neoplasia. 11:1208–1215. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rajabi H, Jin C, Ahmad R, McClary AC,
Joshi MD and Kufe D: Mucin 1 oncoprotein expression is suppressed
by the miR-125b oncomir. Genes Cancer. 1:62–68. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kaenel P, Mosimann M and Andres AC: The
multifaceted roles of Eph/ephrin signaling in breast cancer. Cell
Adh Migr. 6:138–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Harburg GC and Hinck L: Navigating breast
cancer: Axon guidance molecules as breast cancer tumor suppressors
and oncogenes. J Mammary Gland Biol Neoplasia. 16:257–270. 2011.
View Article : Google Scholar : PubMed/NCBI
|