Introduction
Hepatocellular carcinoma (HCC) is the most common
type of liver tumor, and the mortality rate for HCC is the third
highest of all types of cancer, following lung and stomach cancer
(1). Hepatitis B virus (HBV) chronic
infection is a risk factor for HCC (2), particularly in Asia and South Africa
(3). HBV X protein (HBx), a 17-kDa
multifunctional protein, serves an important role in HBV-associated
HCC (4–6).
During chronic HBV infection, HBV DNA can integrate
into the host genome (7). The HBx
gene can be maintained and transcribed in human HCC cells, although
complete HBV replication does not occur (7). Recent studies have demonstrated that HBx
influences the entire process of HBV-associated HCC, including
inflammation, cirrhosis and HCC development (8–10). HBx
regulates a variety of signaling pathways, including
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mechanistic
target of rapamycin, Wnt/β-catenin and P53, through modulating the
expression and activity of numerous genes (11–13). These
events also include epigenetic modification and mutation. However,
it is difficult to eliminate HBx once it has been integrated into
the host genome. Our previous study detected HBx, particularly
truncated HBx, in HBV-derived HCC cell lines (e.g., Hep3B and
SNU423) (14). Therefore, the
identification of an effective target is critical for inhibiting
the oncogenic function of HBx.
High mobility group box 1 (HMGB1) is a non-histone,
nuclear DNA-binding protein (15).
HMGB1 is also a multifunctional protein, and its function depends
on its location in the cell (15). In
the nucleus, as a DNA binding protein, HMGB1 has a role in a number
of crucial DNA events (16–20). Under certain conditions, including
starvation and reactive oxygen species (ROS), HMGB1 can translocate
to the cytoplasm and can transit to the extracellular environment
(21). Extra-nuclear HMGB1 regulates
several signaling pathways affecting cell functions, including
proliferation, autophagy and apoptosis (22–24).
Additionally, extracellular HMGB1 can affect the microenvironment
by inducing inflammation, fibrosis and angiogenesis (25–27). Our
previous study revealed that the deletion of HMGB1 worsened liver
ischemia/reperfusion injury (28).
Recently, increasing evidence has indicated that HMGB1 has a
significant association with HCC. For example, the HMGB1 serum
level is increased in patients with chronic HBV and this is
associated with the disease stage (29), and HMGB1 can also translocate outside
of the nucleus in response to HBV infection (15). These findings suggested that HBV may
regulate HMGB1.
The present study demonstrated that the HBx gene was
expressed in HBV-derived cell lines. The expression of HMGB1 was
associated with the HCC pathological grade and the overall survival
of patients. The non-HBV-derived cell line Huh7 was selected to
investigate the association between HBx and HMGB1. It was revealed
that HBx could regulate the expression of HMGB1 and knockdown of
HMGB1 could decrease the ability of HBx to promote cellular
proliferation. Next, several transcription factors that had been
reported to regulate HMGB1 were investigated using database
analysis (e.g., Genecards and Targetscan). Through analysis of
these transcription factors, the mechanism for HBx-mediated
regulation of HMGB1 was identified. By analyzing the genes
co-expressed with HMGB1 in patients with HBV, several key pathways
that could promote the formation of HCC were identified. These
findings suggested that HMGB1 is involved in the process of
HBV-induced HCC.
Materials and methods
Acquisition of human tissue
specimens
The three patients were aged 63, 48 and 46 years old
(mean was 52.3 years) and comprised of 2 females and 1 male. All
these three tissue were obtained in April 2015. Inclusion criteria
for the present study were: Each patient was infected by HBV and
HBx and HMGB1 had a higher expression in tumor tissue when compared
with normal tissue. Paired HCC and adjacent liver tissues were
obtained from patients who underwent hepatectomy at the Department
of Hepatopancreatobiliary Surgery of the Second Affiliated Hospital
of Harbin Medical University (Harbin, China). All human tissues
were acquired in accordance with the protocol approved by the
Ethics Committee of Harbin Medical University and written informed
consent was obtained from all the patients.
Cell lines
The non-HBV-derived HCC Huh7 cell line was obtained
from the Department of Surgery of the University of Pittsburgh
Medical Center. The cells were used immediately following receipt
and were cultured in Dulbecco's modified Eagle's medium with 10%
fetal bovine serum (Hyclone; GE Healthcare Life Sciences, Logan,
UT, USA) at 37°C with 5% CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. Next, 1 µg total RNA from
each sample was reverse transcribed to single-stranded cDNA with
RNA to cDNA EcoDry™ Premix (Clontech Laboratories, Inc.,
Mountainview, CA, USA). cDNA (1 µl) was diluted 50-fold with
nuclease-free water and was used as a template for RT-qPCR. All the
primers were purchased from Sangon Biotech Co., Ltd. (Shanghai,
China). HBx forward, 5′-TTCTTCGTCTGCCGTTCC-3′ and reverse,
5′-TCGGTCGTTGACATTGCT-3′; HMGB1 forward, 5′-TATGGCAAAAGCGGACAAGG-3′
and reverse, 5′-CTTCGCAACATCACCAATGGA-3′; GAPDH forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′; GATA3 forward,
5′-GCCCCTCATTAAGCCCAAG-3′ and reverse, 5′-TTGTGGTGGTCTGACAGTTCG-3′;
TBP forward, 5′-ACTCCACTGTATCCCTCCCC-3′ and reverse,
5′-TATATTCGGCGTTTCGGGCA-3′; HSF1 forward,
5′-CCATGAAGCATGAGAATGAGGC-3′ and reverse,
5′-CTTGTTGACGACTTTCTGTTGC-3′; ERBB3 forward,
5′-GGTGATGGGGAACCTTGAGAT-3′ and reverse,
5′-CTGTCACTTCTCGAATCCACTG-3′; KLF4 forward,
5′-CCCACATGAAGCGACTTCCC-3′ and reverse,
5′-CAGGTCCAGGAGATCGTTGAA-3′; NFKB1 forward,
5′-AACAGAGAGGATTTCGTTTCCG-3′ and reverse,
5′-TTTGACCTGAGGGTAAGACTTCT-3′. The thermo cycling conditions were
as follows: 94°C 20 sec, 60°C 20 sec, 72°C 2 min, 30 cycles.
Analysis of relative gene expression data was using real-time
quantitative PCR and the 2−ΔΔCq method (30). Gene expression of HBx and HMGB1 were
normalized to GAPDH. Gene expression of GATA3, TBP, HSF1, ERBB3,
KLF4 and NFKB1 were normalized to the control group. The reverse
transcription reactions were prepared using the TaqMan Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Each 15-µl multiplex reaction contained 10 ng total RNA as a
template.
Western blot analysis
Whole protein was extracted using cell lysis buffer
(Cell Signaling Technology, Inc., Danvers, MA, USA). The nuclear
protein was extracted as previously described (23). BCA was used as the protein
determination method. A total of 20 µg nuclear protein was
electrophoresed on 10% SDS-polyacrylamide gels and transferred onto
polyvinylidene difluoride membranes. Following blocking with 5%
skimmed milk at room temperature for 1 h, the membranes were
incubated with a 1:5,000 dilution of the primary antibodies with
bovine serum albumin (Thermo Fisher Scientific, Inc.) for HMGB1,
HBx and GAPDH (cat. nos. ab79823, ab39716 and ab181602,
respectively) at 4°C overnight. Next, the membranes were washed
with Tris-buffered saline and Tween-20 (TBST) three times,
incubated with a 1:10,000 dilution of the secondary antibody (goat
anti-rabbit; HRP; cat. no. ab191866) at room temperature for 1 h,
and developed onto X-ray film using a chemiluminescent reagent
(cat. no. c510043; Sangon Biotech, Co., Ltd.). All of the
antibodies were purchased from Abcam (Cambridge, UK).
Plasmid construct and cell
transfection
The PcDNA-3.1-HBx plasmid was purchased from Sangon
Biotech Co., Ltd. The control shRNA and HMGB1 shRNA were purchased
from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). The FuGene
transfection reagent (Promega Corporation, Madison, WI, USA) was
used at a reagent to plasmid ratio of 3:1 and the concentration was
1 µg/ml. Transfection was performed according to the manufacturer's
protocol, and cells were harvested after 48 h for the following
experiments.
Database analysis and statistical
analysis
The Cancer Genome Atlas (TCGA) data were obtained
from Synapse TCGA (https://www.synapse.org/). Gene expression data and
patient data from 377 patients were analyzed, however 6 patients
did not have HBV information. The gene expression association
analysis was completed using R2 (http://r2.amc.nl/) and Prism 5. The signaling pathway
analysis was performed using Toppgene (https://toppgene.cchmc.org/) and R2. The statistical
analysis was performed using Prism 5, and gene information was
collected from Genecards (http://www.genecards.org/). Data are presented as the
mean ± standard deviation. Two-way analysis of variance with
Tukey's multiple comparison test was used for raw data analysis and
P<0.05 was considered to indicate a statistically significant
difference.
Results
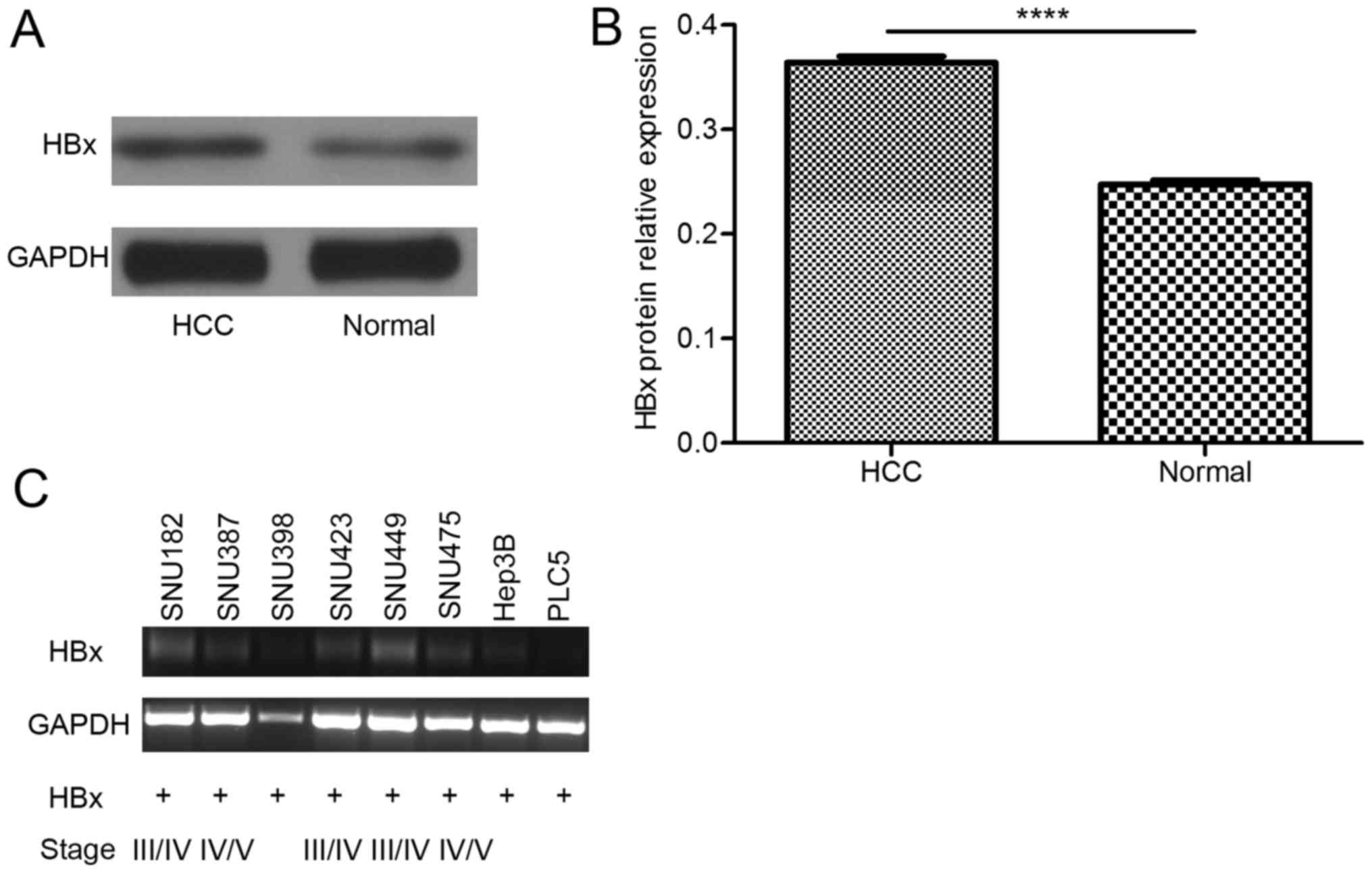
HBx expression is higher in tumor
tissue than in adjacent noncancerous tissue, and truncated HBx can
be detected in HBV-derived cell lines
The results of in vivo studies revealed that
HBx protein expression was higher in HCC tumor tissue than in the
adjacent non-cancerous tissue (Fig. 1A
and B). Once the HBx gene is integrated into the host genome,
it is difficult to eliminate. Accordingly, in in vitro
studies, truncated HBx was detected (Fig.
1C), which is the form of HBx with a greater effect (14). These results suggested that HBx is
expressed in liver cells for a long time and contributes to the
process of HCC.
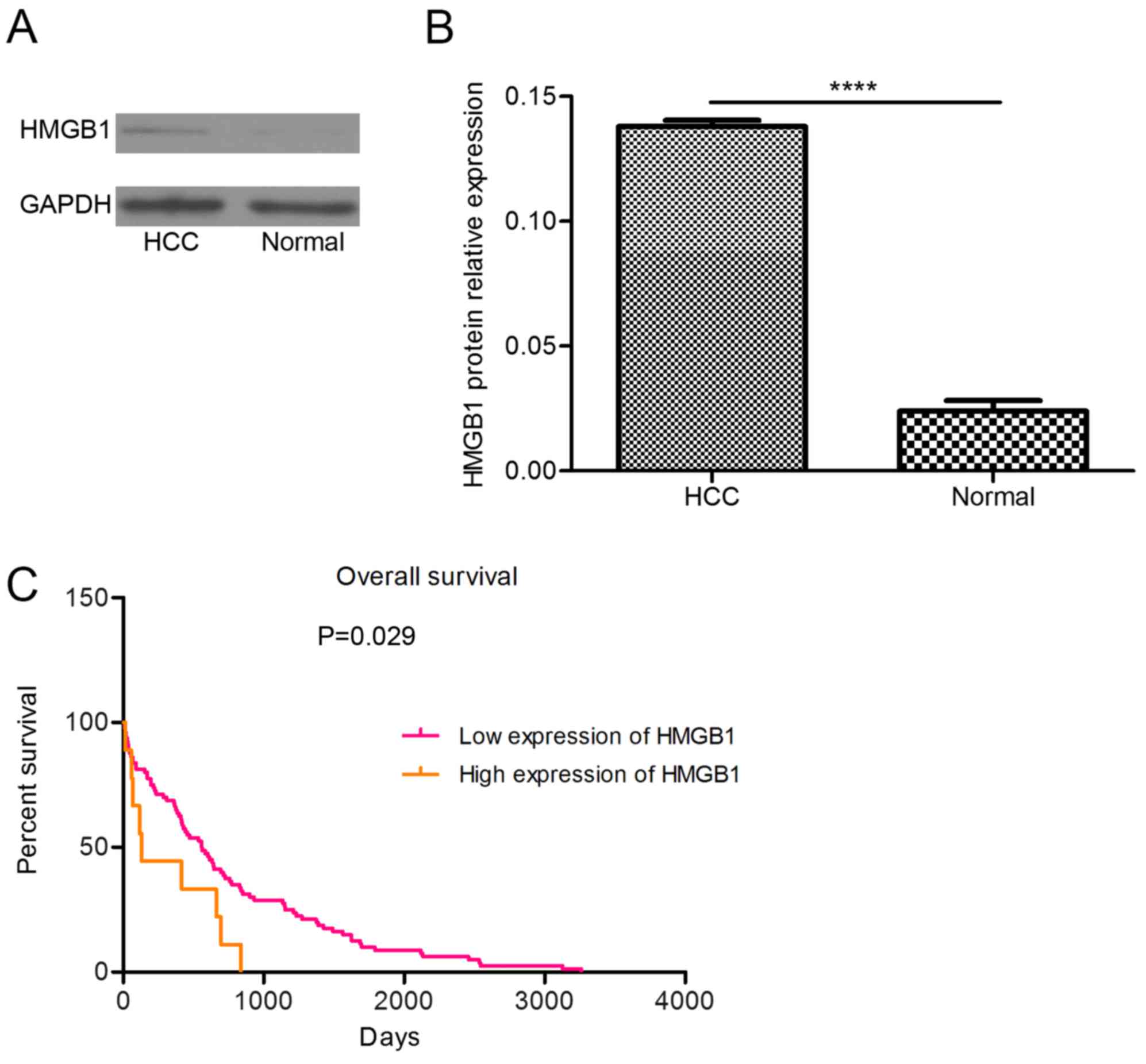
HMGB1 expression is higher in tumor
tissue than in adjacent non-cancerous tissue, and a high level of
HMGB1 is associated with the pathological grade of HCC and the
survival of patients
The protein level of HMGB1 was higher in the tumor
tissue than in the adjacent non-cancerous tissue (Fig. 2A and B). By analyzing TCGA data, the
present study revealed that the prognosis of patients with
high-level HMGB1 expression was poor (Fig. 2C). These findings suggested that
abnormal expression of HMGB1 can induce HCC.
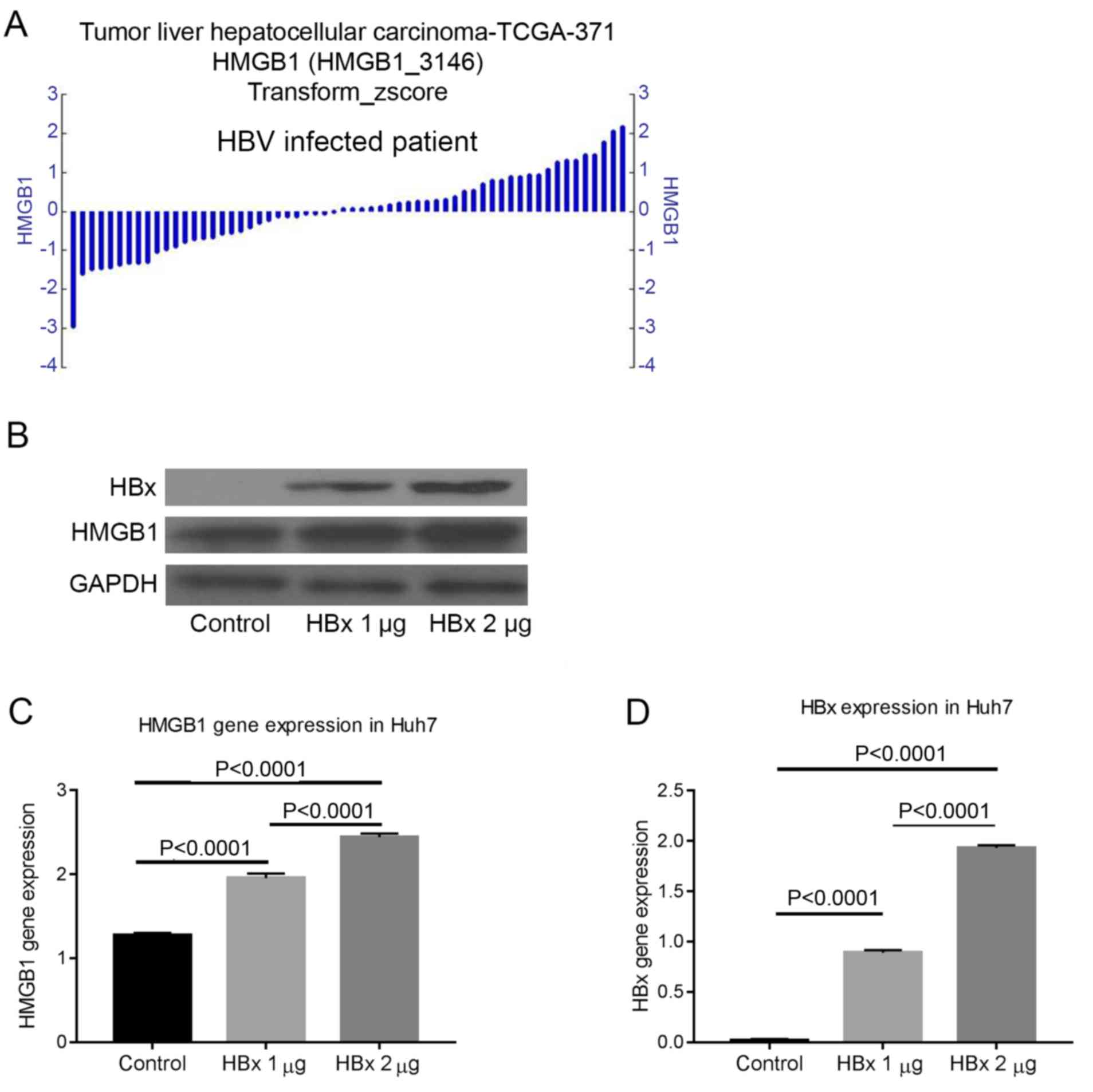
HBx can regulate the expression of
HMGB1
The gene expression of HMGB1 differed in patients
with HBV-induced HCC. This finding suggested that HBV may regulate
HMGB1 (Fig. 3A). To address this
hypothesis, the non-HBV-derived Huh7 cell line was transfected with
different quantities of pcDNA-3.1-HBx plasmid and the protein
expression was measured by western blot analysis after 72 h. The
results confirmed that HBx could regulate the expression of HMGB1
(Fig. 3B-D).
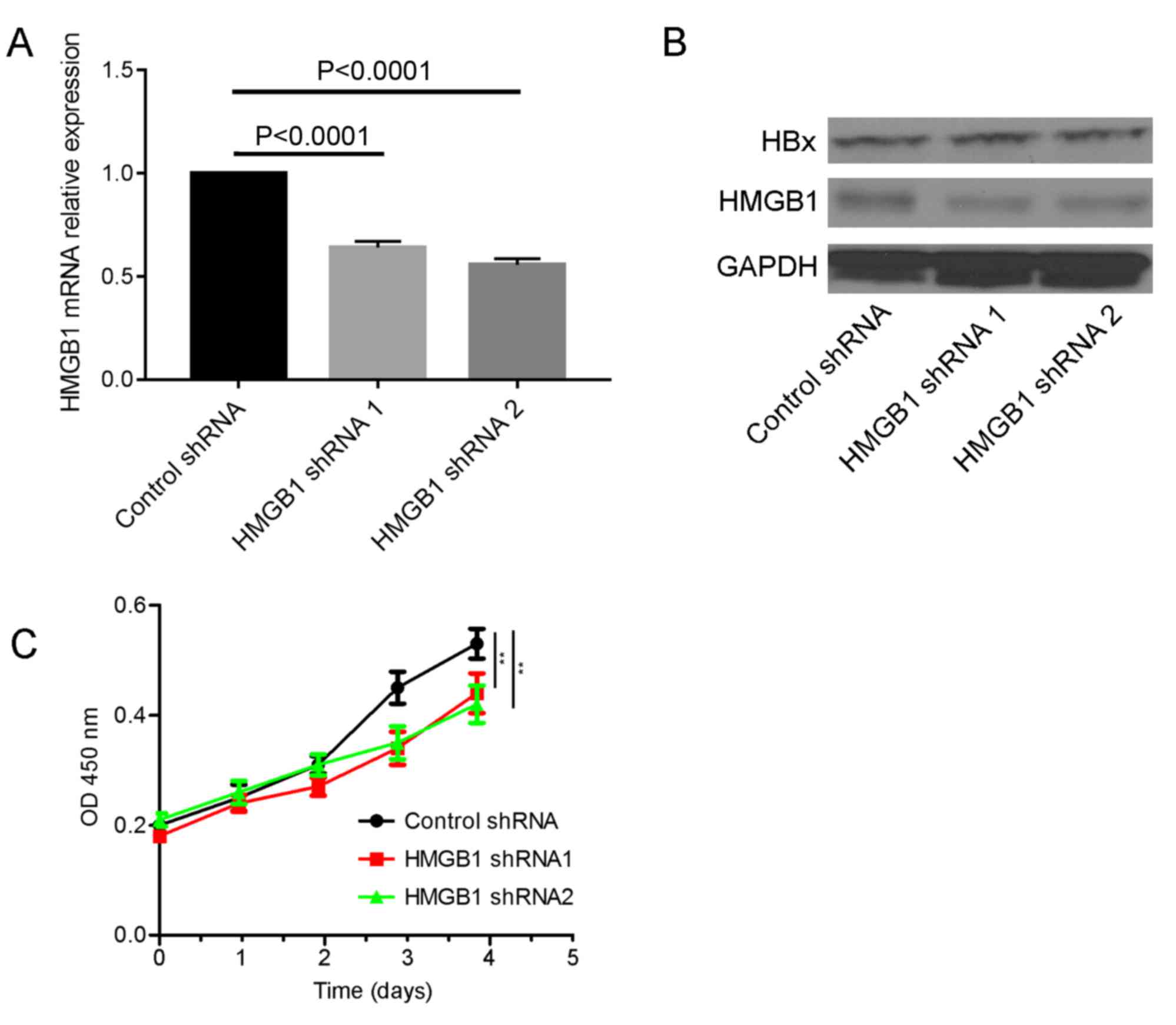
HMGB1 is required for HBx to promote
the cellular proliferation
To investigate the role of HMGB1 in the carcinogenic
process of HBx, a HBx stable expression cell line was created
(Fig. 4A and B). HMGB1 was knocked
down in the HBx stable expression cell line and it was revealed
that the ability of HBx to promote proliferation was decreased
(Fig. 4C).
HBx increases the expression of HMGB1
by regulating transcription factors associated with HMGB1
To clarify the molecular mechanism of HBx-mediated
regulation of HMGB1, several transcription factors associated with
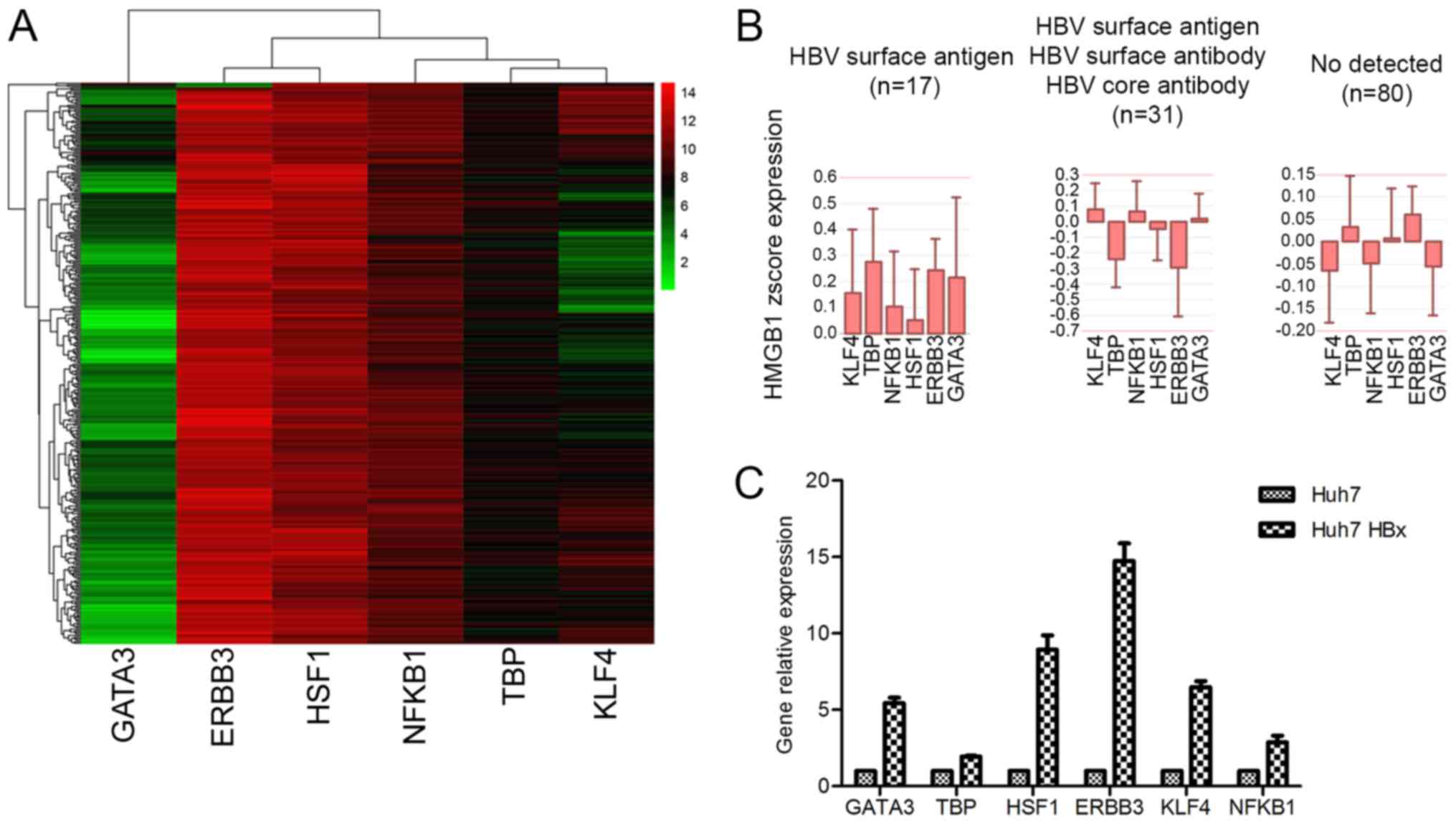
HMGB1 that could be expressed in HCC were investigated. Eventually,
six possible transcription factors: GATA3, ERBB3, HSF1, NFKB1, TBP
and KLF4 were identified (Fig. 5A).
Several of these transcription factors were highly expressed in
patients with HBV (Fig. 5B).
HBx affects a variety of cellular
functions through regulation of HMGB1
A number of genes that are co-expressed with HMGB1
were abnormally expressed in HBV-induced HCC. Therefore, several
genes that exhibited a significant associated with HMGB1 were
further examined. Next, the top 19 genes were selected to analyze
the functions associated with HMGB1 (Table I). By analyzing these genes using
Toppgene, the top 16 signaling pathways enriched for genes that are
co-expressed with HMGB1 were obtained (Table II).
 | Table I.Top 19 genes co-expressed with HMGB1
in patients with HBV. |
Table I.
Top 19 genes co-expressed with HMGB1
in patients with HBV.
| Gene | Function | R2 | P-value |
|---|
| CDKN1B | Cell cycle,
development | 0.303 |
2.5×10−9 |
| PCNA | DNA repair,
signaling transduction | 0.289 |
1.5×10−8 |
| CDK2 | Cell cycle,
kinase | 0.204 |
7.6×10−5 |
| CREB1 | TF, drug target,
signal transduction, transcription regulator activator | 0.199 |
1.2×10−4 |
| CCNE2 | Cell cycle | 0.193 |
1.9×10−4 |
| BIRC5 | Apoptosis, cell
cycle | 0.188 |
2.7×10−4 |
| NFKBIA | Apoptosis, signal
transduction | 0.186 |
3.1×10−4 |
| CCNA2 | Cell cycle | 0.171 |
9.8×10−4 |
| YWHAQ | Signal
transduction | 0.169 |
1.1×10−3 |
| E2F1 | TF, apoptosis, cell
cycle, development, transcription regulator activator | 0.168 |
1.2×10−3 |
| CREB3L4 | TF, transcription
regulator activator | 0.144 |
5.5×10−3 |
| CDK4 | Cell cycle,
kinase | 0.144 |
5.5×10−3 |
| RB1 | TF, cell cycle,
drug target, signal transduction, transcription regulator
activator | 0.133 |
1.0×10−2 |
| TGFBR1 | Kinase, membrane,
signal transduction | 0.123 |
2.0×10−2 |
| E2F2 | TF, cell cycle,
transcription regulator activator | 0.117 |
2.0×10−2 |
| SMAD4 | TF, development,
signal transduction, transcription regulator activator | 0.113 |
3.0×10−2 |
| STAT4 | TF, signal
transduction, transcription regulator activator | 0.109 |
4.0×10−2 |
| APAF1 | Apoptosis,
development, drug target | 0.105 |
4.0×10−2 |
| BAX | Apoptosis, cell
cycle, development, differentiation, membrane | 0.102 |
5.0×10−2 |
| Table II.Top 16 signaling pathways enriched
for genes that were co-expressed with high mobility group box
1. |
Table II.
Top 16 signaling pathways enriched
for genes that were co-expressed with high mobility group box
1.
| Name | Source | P-value | FDR B&H | FDR B&Y | Bonferroni | Gene input |
|---|
| Cell cycle | BioSystems:
WikiPathways |
6.16×10−17 |
1.57×10−14 |
1.09×10−13 |
3.6×10−14 | 11 |
| p53 signaling
pathway | MSigDB C2 BIOCARTA
(v5.1) |
1.07×10−15 |
1.04×10−13 |
7.23×10−13 |
6.25×10−13 | 7 |
| DNA damage
response | BioSystems:
WikiPathways |
8.61×10−15 |
6.28×10−13 |
4.37×10−12 |
5.03×10−12 |
|
| Viral
carcinogenesis | BioSystems:
KEGG |
1.67×10−13 |
7.49×10−12 |
5.20×10−11 |
9.74×10−11 | 11 |
| Pathways in
cancer | BioSystems:
KEGG |
8.38×10−13 |
3.49×10−11 |
2.43×10−10 |
4.89×10−10 | 12 |
| PI3K-Akt signaling
pathway | BioSystems:
KEGG |
1.64×10−10 |
6.26×10−11 |
4.35×10−10 |
9.55×10−10 | 12 |
| E2F transcription
factor network | BioSystems: Pathway
Interaction Database |
1.82×10−12 |
6.26×10−11 |
4.35×10−10 |
1.06×10−9 | 8 |
| DNA
replication | BioSystems:
REACTOME |
2.01×10−9 |
4.71×10−8 |
3.27×10−7 |
1.18×10−6 | 7 |
| Regulation of IFNA
signaling | BioSystems:
REACTOME |
2.10×10−9 |
4.72×10−8 |
3.28×10−7 |
1.23×10−6 | 5 |
| TRAF6-mediated IRF7
activation | BioSystems:
REACTOME |
6.38×10−9 |
1.1×10−7 |
7.61×10−7 |
3.73×10−6 | 5 |
| Regulation of
autophagy | BioSystems:
KEGG |
8.79×10−9 |
1.39×10−7 |
9.64×10−7 |
5.13×10−6 | 5 |
| Integrated cancer
pathway | BioSystems:
WikiPathways |
1.02×10−8 |
1.57×10−7 |
1.09×10−6 |
5.98×10−6 | 5 |
| Regulation of p27
phosphorylation during cell cycle progression | MSigDB C2 BIOCARTA
(v5.1) |
1.27×10−8 |
1.85×10−7 |
1.29×10−6 |
7.41×10−6 | 4 |
| RIG-I/MDA5-mediated
induction of IFN-α/β pathways | BioSystems:
REACTOME |
1.55×10−8 |
2.21×10−7 |
1.54×10−6 |
9.06×10−6 | 6 |
| IL-7 signaling
pathway | BioSystems:
WikiPathways |
3.38×10−8 |
4.45×10−7 |
3.09×10−6 |
1.97×10−5 | 5 |
| Regulation of TLR
signaling pathway | BioSystems:
WikiPathways |
6.74×10−8 |
8.2×10−7 |
5.7×10−6 |
3.94×10−5 | 6 |
Discussion
HBx serves an important role in the process of
HBV-induced HCC (Fig. 1A and B). The
carcinogenic mechanisms of HBx have been reported in numerous
previous studies (7–13). The HBx gene integrates into the host
cell genome during HBV infection (Fig.
1C) (14) and is very difficult
to completely remove. Therefore, the best method for inhibiting the
function of HBx is to identify an effective target to prevent
integration.
HMGB1 is a multifunctional protein, and its function
depends on its location. Abnormal expression of HMGB1 in different
organs can induce different diseases (15). High expression of HMGB1 can influence
several cell functions and the microenvironment of the liver
(15–27); accordingly, HMGB1 has been associated
with the development and progression of HCC (Fig. 2) and has also been associated with HBV
infection (29). In the present
study, it was confirmed that HBx could increase the expression of
HMGB1 through the regulation of transcription factors (Figs. 3 and 5).
Therefore, HMGB1 may be an ideal target for inhibiting the
carcinogenic function of HBx.
To study the molecular mechanisms of HBx-induced HCC
through the regulation of HMGB1, genes significantly associated
with HMGB1 in patients with HBV were investigated (Table I). Next, these genes were sorted into
signaling pathways, as presented in Table II. Abnormalities in some of these
pathways have been reported to be associated with HBx and HMGB1.
For example, the P53 signaling pathway is a classical tumor
suppressor pathway, and HBx and HMGB1 can regulate cell
proliferation and apoptosis by suppressing P53 (2,31).
PI3K/Akt is an important signaling pathway through which HBx can
induce autophagy (32), and HMGB1 is
involved in the same pathway when responding to oxidative stress
(33). Furthermore, the Toll-like
receptor (TLR) signaling pathway also contributes to HBx-induced
carcinogenesis (34), and a previous
study demonstrated that HMGB1 could increase the expression of
mitochondria in HCC by binding to TLRs (35).
In conclusion, HBx and HMGB1 have a synergistic
effect in promoting HCC. The present study confirmed that HBx could
regulate HMGB1. Additionally, several signaling pathways associated
with HBx and HMGB1 that may serve as the mechanism whereby HBx
promotes HCC through the regulation of HMGB1 were identified.
Nevertheless, further investigation is required to identify the
precise mechanism through which HBx induces HCC through HMBG1
regulation.
Acknowledgements
Not applicable.
Funding
This study was supported by Heilongjiang Natural
Science Foundation (grant no. H2018025).
Availability of data and materials
HCC TCGA data were obtained from http://www.synapse.org/. All other data analyzed
during the current study are included in the published article.
Authors' contributions
DW and HL contributed to experiment design, drafting
the manuscript and revising. HW, CD, HY and JZ conducted the
western blotting and PCR experiments, and data collection. YP, BS,
ZS, JW and TC contributed to data analysis. ST contributed to
experiment design and drafting the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Harbin Medical University and written informed consent
was obtained from all the patients.
Consent for publication
All patients provided consent for the publication of
the paper and any associated images.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ERBB3
|
Erb-B2 receptor tyrosine kinase 3
|
|
GATA3
|
GATA binding protein 3
|
|
HBx
|
hepatitis B virus-X protein
|
|
HMGB1
|
high mobility group box 1
|
|
HSF1
|
heat shock transcription factor 1
|
|
KLF4
|
Kruppel-like factor 4
|
|
NFKB1
|
nuclear factor κB subunit 1
|
|
TBP
|
TATA-box binding protein
|
References
|
1
|
Hernandez-Gea V, Toffanin S, Friedman SL
and Llovet JM: Role of the microenvironment in the pathogenesis and
treatment of hepatocellular carcinoma. Gastroenterology.
144:512–527. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lu JW, Yang WY, Tsai SM, Lin YM, Chang PH,
Chen JR, Wang HD, Wu JL, Jin SL and Yuh CH: Liver-specific
expressions of HBx and src in the p53 mutant trigger
hepatocarcinogenesis in zebrafish. PLoS One. 8:e769512013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen JY, Chen YJ, Yen CJ, Chen WS and
Huang WC: HBx sensitizes hepatocellular carcinoma cells to
lapatinib by up-regulating ErbB3. Oncotarget. 7:473–489.
2016.PubMed/NCBI
|
|
4
|
Liu XY, Tang SH, Wu SL, Luo YH, Cao MR,
Zhou HK, Jiang XW, Shu JC, Bie CQ, Huang SM, et al: Epigenetic
modulation of insulin-like growth factor-II overexpression by
hepatitis B virus X protein in hepatocellular carcinoma. Am J
Cancer Res. 5:956–978. 2015.PubMed/NCBI
|
|
5
|
Zhao J, Wu J, Cai H, Wang D, Yu L and
Zhang WH: E3 Ubiquitin ligase Siah-1 is Down-regulated and fails to
target natural HBx truncates for degradation in hepatocellular
carcinoma. J Cancer. 7:418–426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu JJ, Song W, Zhang SD, Shen XH, Qiu XM,
Wu HZ, Gong PH, Lu S, Zhao ZJ, He ML and Fan H: HBx-upregulated
lncRNA UCA1 promotes cell growth and tumorigenesis by recruiting
EZH2 and repressing p27Kip1/CDK2 signaling. Sci Rep. 6:235212016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Amaddeo G, Cao Q, Ladeiro Y, Imbeaud S,
Nault JC, Jaoui D, Gaston Mathe Y, Laurent C, Laurent A,
Bioulac-Sage P, et al: Integration of tumour and viral genomic
characterizations in HBV-related hepatocellular carcinomas. Gut.
64:820–829. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Levrero M and Zucman-Rossi J: Mechanisms
of HBV-induced hepatocellular carcinoma. J Hepatol. 64 Suppl
1:S84–S101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Geng M, Xin X, Bi LQ, Zhou LT and Liu XH:
Molecular mechanism of hepatitis B virus X protein function in
hepatocarcinogenesis. World J Gastroenterol. 21:10732–10738. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tarocchi M, Polvani S, Marroncini G and
Galli A: Molecular mechanism of hepatitis B virus-induced
hepatocarcinogenesis. World J Gastroenterol. 20:11630–11640. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu M, Guo J, Li W, Xia H, Lu Y, Dong X,
Chen Y, Xie X, Fu S and Li M: HBx induced AFP receptor expressed to
activate PI3K/AKT signal to promote expression of Src in liver
cells and hepatoma cells. BMC Cancer. 15:3622015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ding SL, Yang ZW, Wang J, Zhang XL, Chen
XM and Lu FM: Integrative analysis of aberrant Wnt signaling in
hepatitis B virus-related hepatocellular carcinoma. World J
Gastroenterol. 21:6317–6328. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chan C, Wang Y, Chow PK, Chung AY, Ooi LL
and Lee CG: Altered binding site selection of p53 transcription
cassettes by hepatitis B virus X protein. Mol Cell Biol.
33:485–497. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ng KY, Chai S, Tong M, Guan XY, Lin CH,
Ching YP, Xie D, Cheng AS and Ma S: C-terminal truncated hepatitis
B virus X protein promotes hepatocellular carcinogenesis through
induction of cancer and stem cell-like properties. Oncotarget.
7:24005–24017. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kang R, Chen R, Zhang Q, Hou W, Wu S, Cao
L, Huang J, Yu Y, Fan XG, Yan Z, et al: HMGB1 in health and
disease. Mol Aspects Med. 40:1–116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cato L, Stott K, Watson M and Thomas JO:
The interaction of HMGB1 and linker histones occurs through their
acidic and basic tails. J Mol Biol. 384:1262–1272. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
El Mezayen R, El Gazzar M, Seeds MC,
McCall CE, Dreskin SC and Nicolls MR: Endogenous signals released
from necrotic cells augment inflammatory responses to bacterial
endotoxin. Immunol Lett. 111:36–44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang C, Peng L and Su J: Two HMGB1 genes
from grass carp Ctenopharyngodon idella mediate immune responses to
viral/bacterial PAMPs and GCRV challenge. Dev Comp Immunol.
39:133–146. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Zhu Y, Lai Y, Wu X, Feng Z, Yu Y,
Bast RC Jr, Wan X, Xi X and Feng Y: Follicle-stimulating hormone
inhibits apoptosis in ovarian cancer cells by regulating the OCT4
stem cell signaling pathway. Int J Oncol. 43:1194–1204. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Furuita K, Murata S, Jee JG, Ichikawa S,
Matsuda A and Kojima C: Structural feature of bent DNA recognized
by HMGB1. J Am Chem Soc. 133:5788–5790. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang D, Kang R, Livesey KM, Cheh CW,
Farkas A, Loughran P, Hoppe G, Bianchi ME, Tracey KJ, Zeh HJ III
and Lotze MT: Endogenous HMGB1 regulates autophagy. J Cell Biol.
190:881–892. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sundberg E, Fasth AE, Palmblad K, Harris
HE and Andersson U: High mobility group box chromosomal protein 1
acts as a proliferation signal for activated T lymphocytes.
Immunobiology. 214:303–309. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu H, Huang L, Zhu S, Li X, Li Z, Yu C
and Yu X: Regulation of autophagy by systemic admission of
microRNA-141 to target HMGB1 in l-arginine-induced acute
pancreatitis in vivo. Pancreatology. 16:337–346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gdynia G, Keith M, Kopitz J, Bergmann M,
Fassl A, Weber AN, George J, Kees T, Zentgraf HW, Wiestler OD, et
al: Danger signaling protein HMGB1 induces a distinct form of cell
death accompanied by formation of giant mitochondria. Cancer Res.
70:8558–8568. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Q, Yang XP, Fang D, Ren X, Zhou H,
Fang J, Liu X, Zhou S, Wen F, Yao X, et al: High-mobility group
box-1 mediates toll-like receptor 4-dependent angiogenesis.
Arterioscler Thromb Vasc Biol. 31:1024–1032. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jia L, Clear A, Liu FT, Matthews J, Uddin
N, McCarthy A, Hoxha E, Durance C, Iqbal S and Gribben JG:
Extracellular HMGB1 promotes differentiation of nurse-like cells in
chronic lymphocytic leukemia. Blood. 123:1709–1719. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Z, Lin C, Peng L, Ouyang Y, Cao Y,
Wang J, Friedman SL and Guo J: High mobility group box 1 activates
Toll like receptor 4 signaling in hepatic stellate cells. Life Sci.
91:207–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang H, Nace GW, McDonald KA, Tai S,
Klune JR, Rosborough BR, Ding Q, Loughran P, Zhu X, Beer-Stolz D,
et al: Hepatocyte-specific high-mobility group box 1 deletion
worsens the injury in liver ischemia/reperfusion: A role for
intracellular high-mobility group box 1 in cellular protection.
Hepatology. 59:1984–1997. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng BQ, Jia CQ, Liu CT, Lu XF, Zhong N,
Zhang ZL, Fan W and Li YQ: Serum high mobility group box
chromosomal protein 1 is associated with clinicopathologic features
in patients with hepatocellular carcinoma. Dig Liver Dis.
40:446–452. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Q, Li J, Wen T, Zeng W, Peng C, Yan S,
Tan J, Yang K, Liu S, Guo A, et al: Overexpression of HMGB1 in
melanoma predicts patient survival and suppression of HMGB1 induces
cell cycle arrest and senescence in association with p21
(Waf1/Cip1) up-regulation via a p53-independent, Sp1-dependent
pathway. Oncotarget. 5:6387–6403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu M, Guo J, Li W, Xia H, Lu Y, Dong X,
Chen Y, Xie X, Fu S and Li M: HBx induced AFP receptor expressed to
activate PI3K/AKT signal to promote expression of Src in liver
cells and hepatoma cells. BMC Cancer. 15:3622015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang FP, Li L, Li J, Wang JY, Wang LY and
Jiang W: High mobility group box-1 promotes the proliferation and
migration of hepatic stellate cells via TLR4-dependent signal
pathways of PI3K/Akt and JNK. PLoS One. 8:e643732013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Cai J, Zeng X, Chen Y, Yan W,
Ouyang Y, Xiao D, Zeng Z, Huang L and Liu A: Downregulation of
toll-like receptor 4 induces suppressive effects on hepatitis B
virus-related hepatocellular carcinoma via ERK1/2 signaling. BMC
Cancer. 15:8212015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tohme S, Yazdani HO, Liu Y, Loughran P,
van der Windt DJ, Huang H, Simmons RL, Shiva S, Tai S and Tsung A:
Hypoxia mediates mitochondrial biogenesis in hepatocellular
carcinoma to promote tumor growth through HMGB1 and TLR9
interaction. Hepatology. 66:182–197. 2017. View Article : Google Scholar : PubMed/NCBI
|