Introduction
Colorectal cancer (CRC) is the third most common
cancer type in the world and is cancer with the second-highest
mortality rate, and its incidence rates continue to increase in a
number of countries, such as Brazil and Russia (1,2). Numerous
factors, including a high-fat diet, which may induce chronic
inflammation in the intestine, clearly increase the risk for CRC
(3–5).
Accumulating evidence suggests that cancer incidence in the colon
is increased compared with that of the small intestine (6,7). With
advances in metagenomics technology and use of germ-free mice, it
has been demonstrated that the intestinal microbiota serves a vital
role in CRC initiation and development (8,9). Indeed,
the microbiota and mucosal gene expression profiles in the gut of
patients with CRC differ from those in healthy subjects (10). Although a number of studies have
documented a critical association between the microbiota
microenvironment and development of CRC (11–13), the
roles of toll-like receptors (TLRs) and the underlying mechanisms
have not been investigated as thoroughly.
TLRs are germline-encoded type I transmembrane
receptors that serve as pathogen recognition receptors (PRRs) to
identify microbe-associated molecular patterns. TLR4 is one of the
characterized PRRs on a number of intestinal immune and non-immune
cell types, which recognizes lipopolysaccharide (LPS) from Gram
staining-negative bacteria in the gut. Generally, TLR4 is expressed
at low concentrations on intestinal mucosa under steady-state
conditions to maintain tolerance to commensal bacteria (14). Activation of TLR4 on colonic
epithelial cells induces a tumor-promoting microenvironment to
drive the tumorigenesis of colitis-associated cancer (15–17). In
addition, TLR4 innate immune signaling alters the colorectal cancer
chemotherapeutic response and radio-sensitivity through regulating
the autophagy pathway (18,19). The administration of a TLR4 inhibitor
or immune-modulator significantly regulates the TLR4 signal pathway
and decreases tumor burden (20,21). Thus,
gut microbiota are associated with carcinogenesis via modulating
TLR, particularly the TLR4 signal pathway in the colon epithelial
cells.
Generally, the metabolites from microbiota in the
gut also alter host intestinal cancer risk using different
mechanisms, including affecting host cell growth or turnover,
regulating the local immune microenvironment and metabolizing
ingested and host-derived products (22). Previous evidence revealed that
short-chain fatty acids (SCFA), primarily acetate, propionate and
butyrate, are major metabolites of microbiota with pleiotropic
effects on the epithelial and immune cells (23). SCFA are catabolic end products of the
microbial fermentation of dietary carbohydrates in the gut and
primarily resistant starches and dietary fiber (24). As a C4 fatty acid, butyrate functions
as the principle energy source for colonocytes. It also exerts
functions in the differentiation, maturation and apoptosis of a
number of different cell types, and may decrease the risk of
gastrointestinal inflammation, colon cancer and cardiovascular
disease (25). Numerous studies have
indicated that the different activities of butyrate and its
concomitant outcomes should be considered with the metabolism of
target cells (26–29). Indeed, in contrast to normal
colonocytes, cancerous colonocytes prefer to use aerobic glycolysis
for glucose metabolism, and butyrate acts as a histone deacetylase
(HDAC) inhibitor to suppress cell growth and to induce apoptosis by
altering gene expression profiles (30).
Therefore, as the coexisting elements of microbiota
in the gut, SCFAs and LPS have demonstrated the regulatory effect
on normal intestinal epithelial cells or colon cancer cells.
However, knowledge of the association between SCFAs and the TLR4
signaling pathway in the development of colon cancer remains
incomplete. The present study, by culturing human colon cancer
SW480 cells or mouse colon cancer CT26 cells with butyrate in
vitro, identified that butyrate suppressed cell growth and
proliferation, and increased the numbers of apoptotic cells.
Notably, the expression levels of TLR4 and cluster of
differentiation 14 (CD14) on these cells were clearly increased. In
addition, butyrate treatment induced the phosphorylation of
extracellular signal-regulated kinase (ERK), tumor protein 38
(p38), c-Jun NH2-terminal kinase (JNK) and nuclear factor-κB
(NF-κB) p65, and then promoted pro-inflammatory cytokine tumor
necrosis factor-α (TNF-α) secretion in SW480 and CT26 cells. These
results suggested that butyrate had the ability to regulate innate
immunity in colon cancer cells via promoting TLR4 expression and
the phosphorylation of mitogen activated protein kinases (MAPKs)
and NF-κB.
Materials and methods
Reagents and antibodies
RPMI-1640 medium and fetal calf serum was purchased
from PAN Biotech (PAN-Biotech GmbH, Aidenbach, Germany). The
CellTrace™ carboxyfluorescein succinimidyl ester (CFSE) Cell
Proliferation kit and the Pierce™ BCA protein assay kit were
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Butyrate, LPS, bovine serum albumin (BSA) and 2-mercaptoethanol
were purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany).
The 7-aminoactinomycin D (7-AAD), anti-human CD14-fluorescein
isothiocyanate (FITC; cat no. 11-0149-42), anti-mouse CD14-FITC
(cat no. 11-0141-82), anti-human TLR4-Alexa Fluor 488 (cat no.
53-9917-42), anti-mouse TLR4-Alexa Fluor 488 (cat no. 53-9917-42),
anti-ERK1/2 (cat no. LF-MA0178) and anti-phosphorylated
(phosphor)-ERK1/2 (cat no. PA5-36776) antibodies, interleukin
(IL)-6 and TNF-α ELISA kits (human and mouse) and Annexin V
Apoptosis Detection kit were purchased from the Thermo Fisher
Scientific, Inc. Radioimmunoprecipitation assay (RIPA) cell lysis
buffer and phenylmethane sulfonyl fluoride (PMSF) were purchased
from the Beyotime Institute of Biotechnology (Haimen, China). The
Cell Cycle Staining kit and goat anti-mouse IgG-horseradish
peroxidase (HRP) antibody were purchased from Hangzhou
MultiSciences (Lianke) Biotech Co., Ltd. (Hangzhou, China).
Antibodies against p38, phospho-p38, JNK, phospho-JNK, NF-κB p65,
NF-κB phospho-p65 and cleaved caspase-3 were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Goat anti-rabbit
IgG-HRP and HRP-conjugated-anti-actin antibodies were purchased
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Colon cancer cell lines
The human colon cancer SW480 cell line and mouse
colon cancer CT26 cell line were obtained from the Chinese Academy
of Sciences (Shanghai, China). Cells were grown in RPMI-1640 medium
supplemented with 10% heat-inactivated fetal calf serum, 100 U/ml
penicillin and 100 mg/ml streptomycin in a humidified incubator at
37°C and 5% CO2. In certain experiments, 5 mM butyrate and/or 1
µg/ml LPS were added into the culture wells.
Flow cytometric analysis
Cells were treated with trypsin for 2 min at room
temperature and then terminated with RPMI-1640 medium containing
10% fetal calf serum. Cells were dispersed with pipette and washed
with PBS twice and passed through 70 µm cell strainers (FalconTM,
Corning Incorporated, Corning, NY, USA). Then, the single cell
suspension was incubated with 2% rabbit serum (Stemcell
Technologies, Vancouver, BC, Canada) for 30 min at room temperature
to block Fc receptors on the cell membrane, followed by staining
with anti-TLR4-FITC or anti-CD14-FITC (both from Thermo Fisher
Scientific, Inc.) at 4°C for 30 min. Following washing with PBS 3
times, cells were re-suspended with PBS (1 mM EDTA) and analyzed
with a FACS Calibur flow cytometer (BD Bioscience, Franklin Lakes,
NJ, USA). All data were analyzed using CellQuest (version 6.1; BD
Biosciences) or FlowJo (version 8.7; Tree Star, Inc., Ashland, OR,
USA).
Cell proliferation and apoptosis
assays
For the cell proliferation assays, SW480 and CT26
cells were harvested with trypsin and the density of the cell
suspension was adjusted to 2×106/ml with PBS. An equal volume of 4
µM carboxyfluorescein succinimidyl ester (CFSE) labeling buffer
(Invitrogen; Thermo Fisher Scientific, Inc.) was added to the cell
suspension, and the cells were gently re-suspended. The cell
suspension was then incubated in the dark at 37°C for 20 min. Next,
RPMI-1640 complete medium was added to terminate CFSE labeling and
the cell suspension was then washed twice with PBS. Cells were
resuspended with fresh RPMI-1640 complete medium and cultured in an
incubator at 37°C and 5% CO2 for 24 h with or without 5 mM butyrate
treatment. Cells were digested with trypsin as previously described
and pelleted by centrifugation for 5 min in 150 × g at 4°C, and
then stained with 7-AAD (1:100) for 2 min at room temperature to
remove dead cells, and were then immediately analyzed with a flow
cytometer (BD FACSCalibur; BD Biosciences).
For cell apoptosis detection, cells were prepared
and harvested as aforementioned in the cell proliferation
experimental protocol, and then stained with the Annexin V
Apoptosis Detection kit according to the manufacturer's protocol.
Cells were additionally analyzed with a flow cytometer (BD
FACSCalibur; BD Biosciences), and the data were analyzed using
FlowJo (version 8.7; Tree Star, Inc., Ashland, OR, USA).
ELISA
The cell supernatant was collected following
centrifugation at 300 × g for 10 min at 4°C. The protein levels of
TNF-α and IL-6 in the supernatant were measured using ELISA kits
(Thermo Fisher Scientific, Inc.) following the manufacturer's
protocols.
Western blot analysis assay
Cells were prepared as aforementioned and washed
twice with cold PBS. Then, the cells were lysed with RIPA cell
lysis buffer and mixed with PMSF (100:1) and 1X phosphatase
inhibitor cocktail (Bimake, Houston, USA) for 20 min on ice. Lysed
cells were scraped into the 1.5 ml tube and centrifuged at 12,000 ×
g for 10 min at 4°C. The supernatants of the cell lysates were
harvested and the protein concentrations were quantified with the
Pierce™ BCA Protein Assay kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The lysate was mixed with
the 5X loading buffer, to which 5% 2-mercaptoethanol was added and
boiled at 100°C for 5 min. All lysate samples (30 µg per lane) were
subjected to SDS-PAGE for protein separation and then
electrophoretically transferred onto polyvinylidene difluoride
(PVDF) membranes (Merck KGaA). Depending on the molecular weight of
the target protein, 10% SDS-PAGE gels were used. Subsequent to
being blocked with TBS containing 5% BSA and 0.1% Tween-20 for 2 h
at room temperature, the PVDF membranes were incubated with
HRP-conjugated anti-actin (cat no., sc-47778; 1:10,000),
anti-ERK1/2 mAb (cat no., 14-9108-82; 1:1,000), anti-phospho-ERK1/2
mAb (cat no., 14-9109-82; MILAN8R; 1:1,000), anti-p38 MAPK mAb (cat
no., 2387; 1:1,000), anti-phospho-p38MAPKThr180/Tyr182 mAb (cat
no., 5175; 1:1,000), anti-SAPK/JNK mAb (cat no., 9252; 1:1,000),
anti-phospho-SAPK/JNKThr183/Tyr185 (cat no., 4668; 1:1,000),
anti-NF-κB P65 mAb (cat no., 8242; 1:1,000), anti-phospho-NF-kB
P65Ser536 (cat no., 3033; 1:1,000) and anti-cleaved caspase-3 (cat
no., 9664; 1:1,000) for at least 12 h at 4°C. Then, the membranes
were incubated with HRP-conjugated secondary antibodies, including
mouse anti-rabbit IgG-HRP (cat no., sc-2357; 1:2,000) and goat
anti-mouse IgG-HRP (cat no., 70-GAM0072; 1:10,000) for 2 h at room
temperature. Following washing three times with TBS, the PVDF
membranes were visualized using the Immobilon™ Western
Chemiluminescent HRP Substrate (Merck KGaA) and quantified by
Fusion Solo 6 S Chemiluminescence (Vilber Lourmat Deutschland GmbH,
Eberhardzell, Germany).
Cell cycle assay
For cell cycle detection, cells were prepared and
harvested as aforementioned in the cell proliferation and apoptosis
experiments. Then, cells were labeled with Cell cycle staining kit
following the manufacture's protocols. The DNA content of cells was
detected using flow cytometry (BD FACSCalibur; BD Biosciences), and
the data were analyzed with Modfit LT 5.0 (American Verity Software
House, Topsham ME, USA).
Statistical analysis
Data were statistically analyzed using the
two-tailed unpaired Student's t-test or post-hoc test (Scheffe) for
two-way analysis of variance (ANOVA) on GraphPad Prism version 5.0
(GraphPad Software, Inc., La Jolla, CA, USA). All data are
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference. The
representative data presented are from more than 3 independent
experiments.
Results
Butyrate suppresses the proliferation
of colon cancer cells
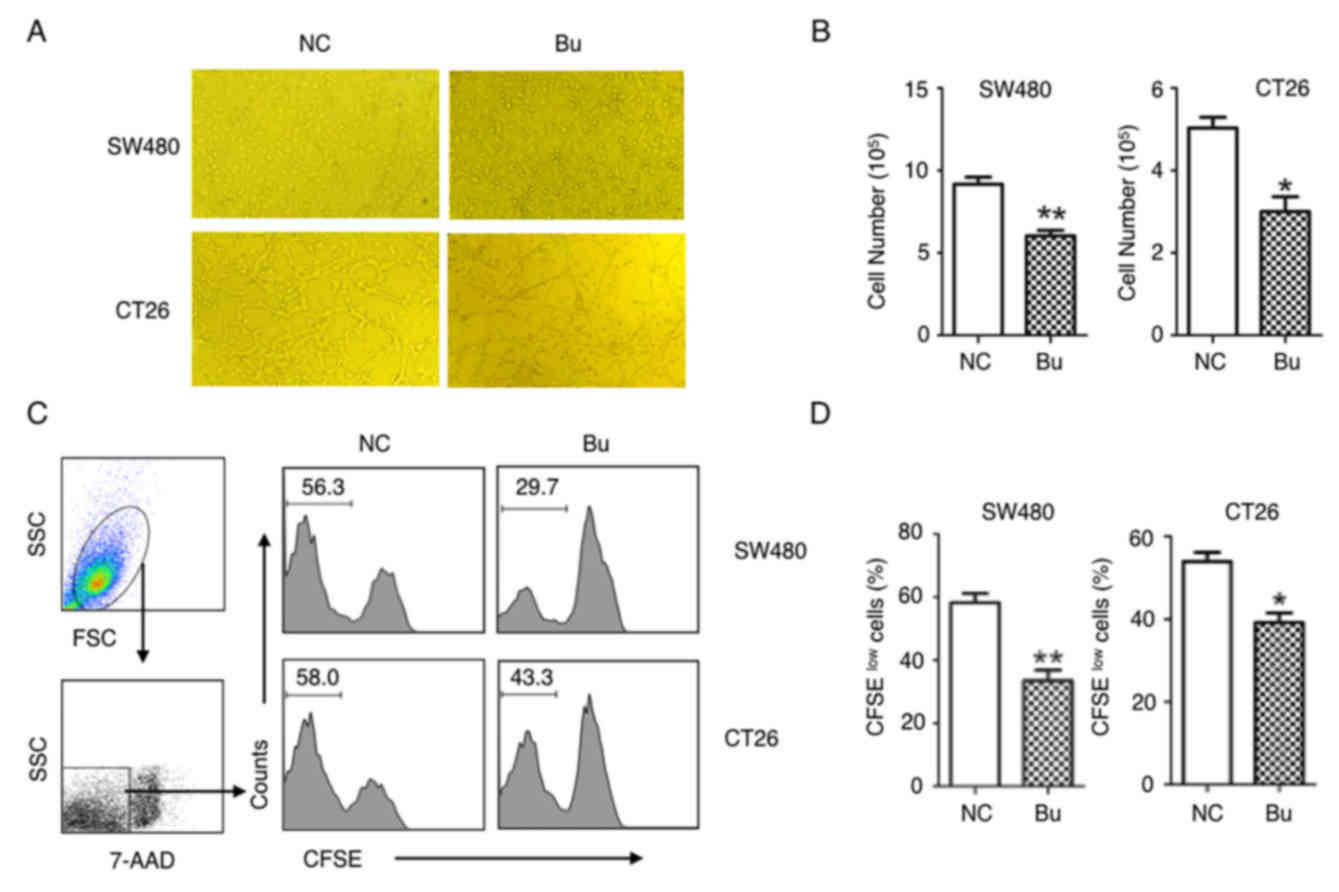
The effects of butyrate on the growth of colon
cancer cells were first examined. By treating human colon cancer
SW480 cells and mouse colon cancer CT26 cells with butyrate, it was
identified that the growth of colon cancer cells was evidently
inhibited and able to be directly observed under the microscope
(Fig. 1A). These results were
additionally confirmed by the data of the cell numbers of colon
cancer cells (Fig. 1B), and CFSE
tracing cell proliferation experiments (Fig. 1C and D). After 24 h culturing, the
proliferation of the SW480 and CT26 cells were significantly
suppressed by butyrate treatment (Fig. 1C
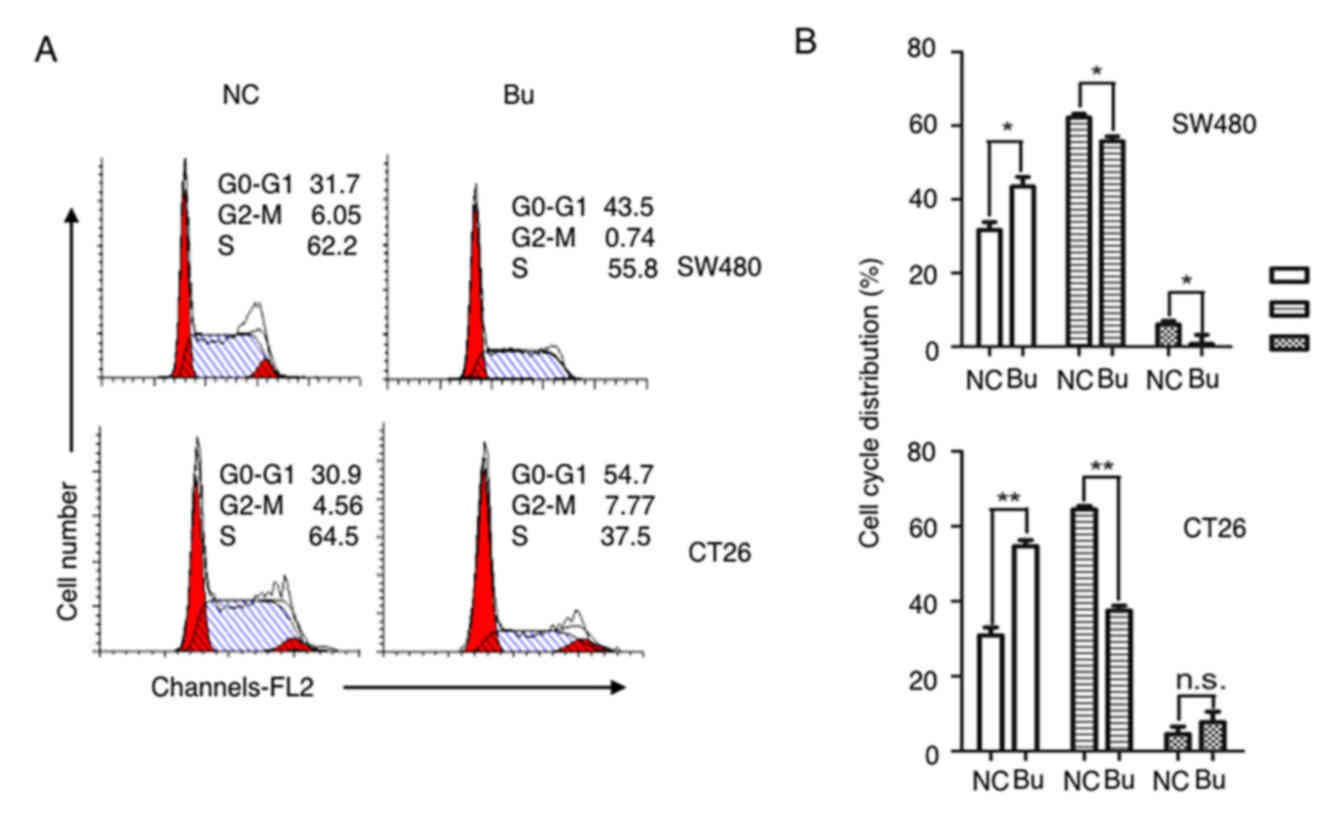
and D). By analyzing the cell cycle with a flow cytometer, an
increased number of cells in S phase were identified in the NC
group compared with the Bu group, and butyrate treatment increased
the numbers of cells in G1 and G2 phases compared with the NC group
(Fig. 2). Therefore, the growth and
the proliferation of colon cancer cells are significantly inhibited
by butyrate.
Butyrate promotes apoptosis in colon
cancer cells in vitro
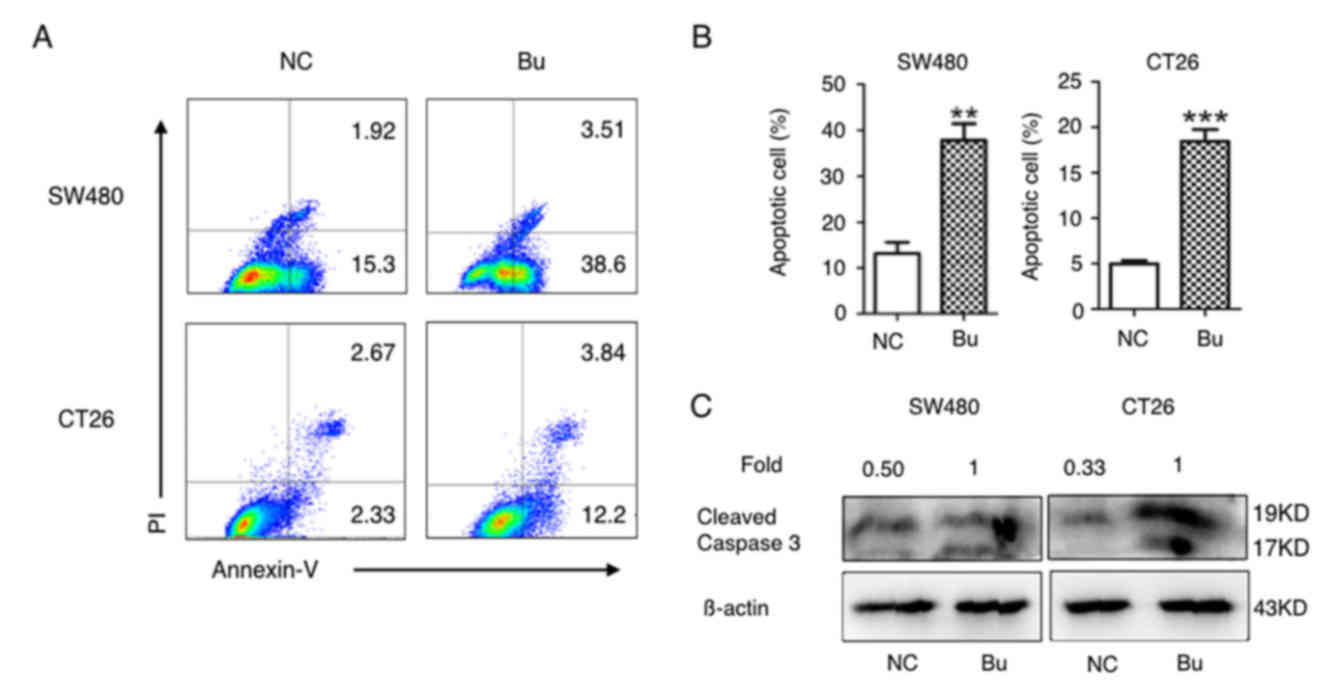
Next, the effects of butyrate on apoptosis induction
in colon cancer cells were investigated. As demonstrated in
Fig. 3A and B, the percentages of
Annexin V-positive apoptotic SW480 and CT26 cells were
significantly increased in butyrate treatment groups compared with
the NC groups. As caspases and their cleaved substrates contribute
to cell apoptosis (31), the levels
of cleaved caspase-3 protein (17 and 19 kD) in SW480 and CT26 cells
was additionally detected using western blot analysis. The data in
Fig. 3C indicates that butyrate
treatment evidently increased the protein levels of cleaved
caspase-3 in the SW480 and CT26 cells. Therefore, butyrate
treatment induces apoptosis in colon cancer cells in
vitro.
Butyrate upregulates the expression of
TLR4 and CD14 on colon cancer cells
Butyrate is produced in the colonic lumen, and a
high concentration of LPS is detected in the gut microenvironment
(32). Butyrate has been demonstrated
to exert immune modulatory effects on colonic epithelial cells
(32), and to possess a regulatory
function on colon cancer cells (33).
TLR4 and CD14 function as the receptor for LPS recognition
(34,35). Next, the effects of butyrate on the
expression of TLR4 and CD14 in the SW480 and CT26 cells were
detected. As demonstrated in Fig. 4,
there was a difference in the baseline expression levels of TLR4
and CD14 in the human colon cancer SW460 cells and mouse colon
cancer CT26 cells. Markedly low expression levels of TLR4 and CD14
were detected in the SW480 cells, but moderate expression of these
molecules in CT26 cells (Fig. 4).
Notably, regardless of what the baseline expression levels of TLR4
and CD14 on the cell membrane were, no marked effects on the
expression of these molecules were observed in the two types of
colon cancer cells following LPS stimulation. However, if SW480 and
CT26 cells were treated with butyrate itself or butyrate + LPS, the
expression levels of TLR4 and CD14 were significantly increased in
the two cell types (Fig. 4).
Therefore, these results suggested that butyrate upregulated the
expression of TLR4 and CD14 in SW480 and CT26 cells, and that the
effects of butyrate were species-independent.
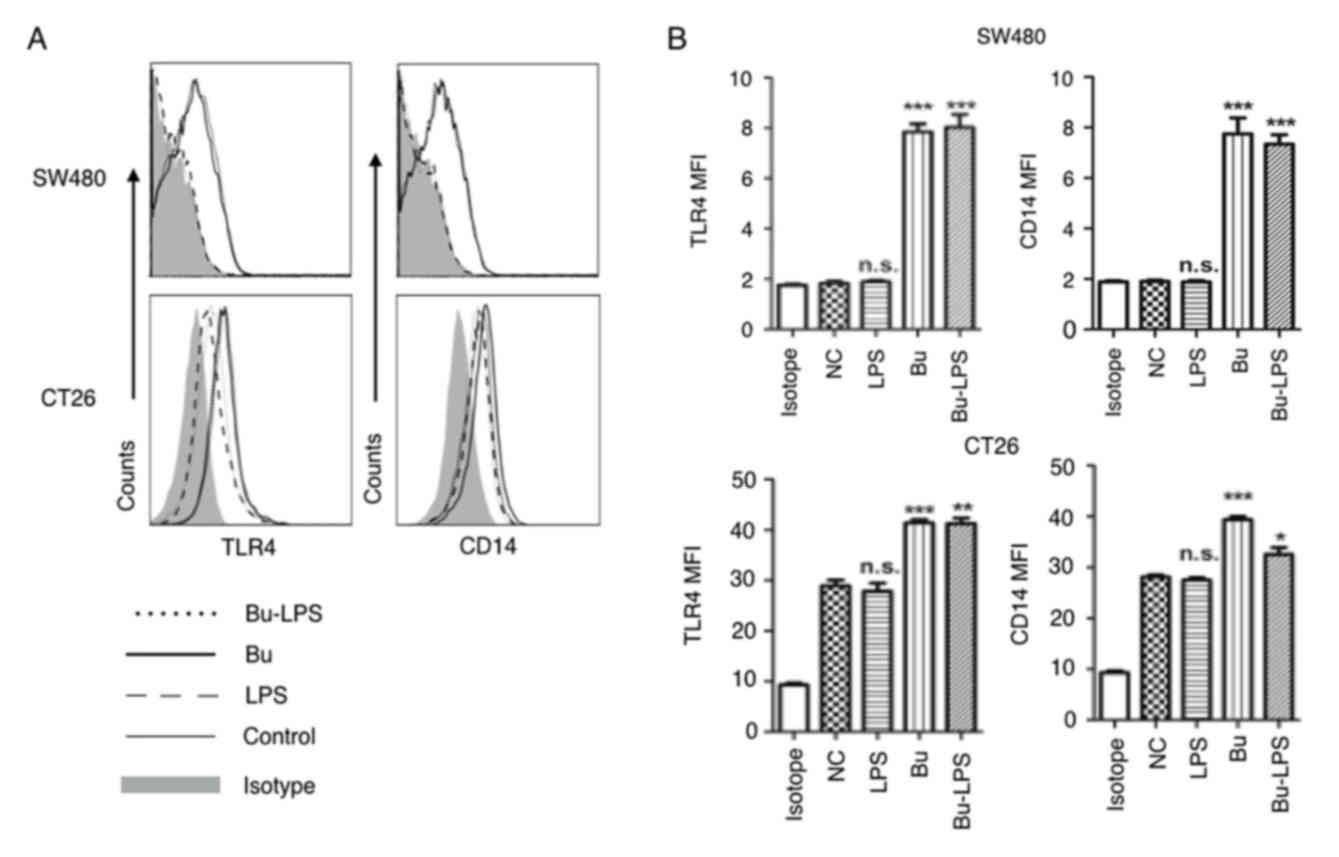 | Figure 4.Butyrate upregulates the levels of
TLR4 and CD14 on colon cancer cells. The expression levels of TLR4
and CD14 on the membrane of SW480 cells and CT26 cells treated by
butyrate and/or LPS were (A) analyzed using a flow cytometer, and
(B) the MFI values of TLR4 and CD14 were quantitative analyzed.
These experiments were repeated ≥3 times, and representative graphs
are presented. Compared with the NC group, *P<0.05, **P<0.01
and ***P<0.001. NS, not significant; MFI, mean fluorescence
index; Bu, butyrate; NC, negative control; LPS, lipopolysaccharide;
TLR4, toll-like receptor 4; CD14, cluster of differentiation
14. |
Butyrate induces the phosphorylation
of ERK, p38, JNK and NF-κB p65 in colon cancer cells
Stimulation of the TLR4 signaling pathway leads to
the activation of three distinct MAPKs, ERK, JNK, p38 and the
activation of NF-κB p65 (36). The
present study therefore elucidated the roles of the TLR4 signaling
pathway in the cellular response to butyrate and LPS in colon
cancer cells. The phosphorylation status of ERK, p38, JNK and NF-κB
p65 were analyzed using western blot analysis. In SW480 cells, as
indicated in Fig. 5, the amount of
p-ERK, p38, JNK and NF-κB p65 were not altered by LPS stimulation
alone but were evidently increased in the butyrate and butyrate +
LPS groups. Different from SW480, LPS-alone simulation increased
the phosphorylation of P-38 and JNK, but not the phosphorylation of
ERK and NF-κB p65 in CT26 cells. This difference may depend on the
different baseline expression levels of TLR4 and CD14 on SW480 and
CT26 cells (Fig. 4), and indicated
the species-specific LPS response in these colon cancer cells.
Notably, butyrate itself and butyrate +LPS stimulation increased
the phosphorylation of ERK, JNK, p38, NF-κB p65 protein in SW480
and CT26 cells (Fig. 5). These data
suggested that butyrate treatment resulted in the activation of
certain MAPKs (ERK, JNK and p38) and the NF-κB signaling pathway in
human and mouse colon cancer cells, and that the butyrate-mediated
activation of these signaling pathways in these cancer cells was
species-independent.
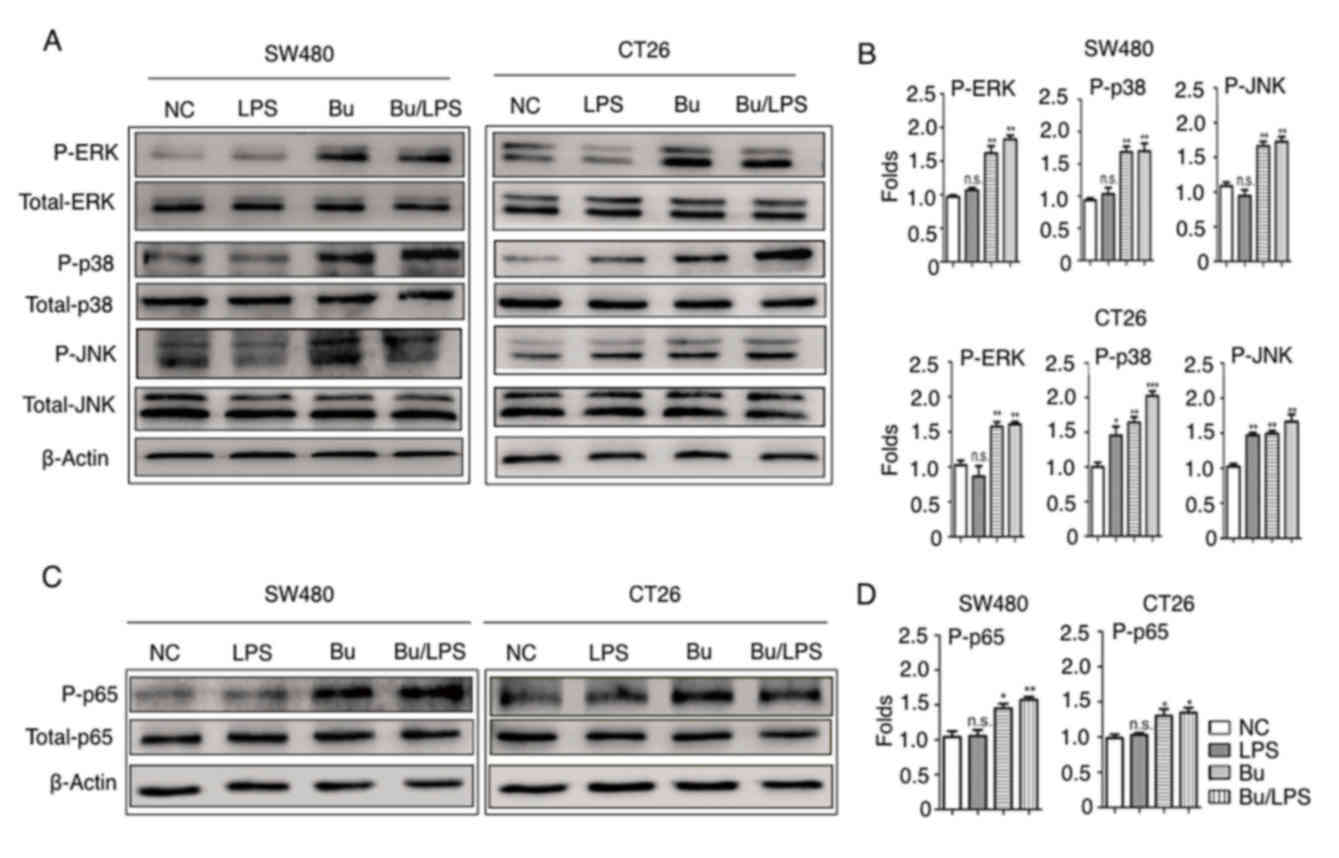 | Figure 5.Butyrate modifies the phosphorylation
of ERK, p38, JNK and NF-κB p65 in colon cancer cells. The
phosphorylation of ERK, p38, JNK and NF-κB p65 proteins in SW480
and CT26 cells treated with butyrate and/or LPS were detected using
a western blot analysis assay. (A) Representative western blot
analysis images of P-ERK/ERK, P-p38/p38 and P-JNK/JNK are
presented. (B) The ratios of P-ERK/ERK, P-p38/p38 and P-JNK/JNK
protein intensities were calculated. (C) Representative western
blot analysis images of P-NF-κB p65/NF-κB p65 protein expression in
cells are presented. (D) P-NF-κB p65/p65 protein intensities were
calculated. β-actin was used as the control. Experiments were
performed in triplicate and data are expressed as the mean ±
standard deviation. Compared with the NC group, *P<0.05,
**P<0.01 and ***P<0.001. These experiments were repeated ≥3
times, and representative images are presented. P, phosphorylated;
ERK, extracellular signal-regulated kinase; p38, tumor protein 38;
JNK, c-Jun NH2-terminal kinase; NS, not significant. |
Butyrate induces TNF-α secretion in
colon cancer cells
The inflammatory cytokine secretion of SW480 and
CT26 cells prior to and following butyrate treatment with or
without LPS was additionally analyzed. The data in Fig. 6A demonstrate that LPS alone was unable
to increase TNF-α production in SW40 cells, but that butyrate or
butyrate + LPS treatment significantly increased the secretion of
TNF-α (Fig. 6). In the case of CT26
cells, butyrate and LPS individually stimulated these cells to
produce increased levels of TNF-α (Fig.
6B). Notably, the production of IL-6 was not promoted by
butyrate and LPS in SW480 cells. Concurrently, in the CT26 cells,
LPS itself and butyrate + LPS induced high levels of IL-6, but
butyrate alone did not increase IL-6 production. These data
suggested that butyrate promoted TNF-α secretion, but not IL-6
production, in colon cancer cells.
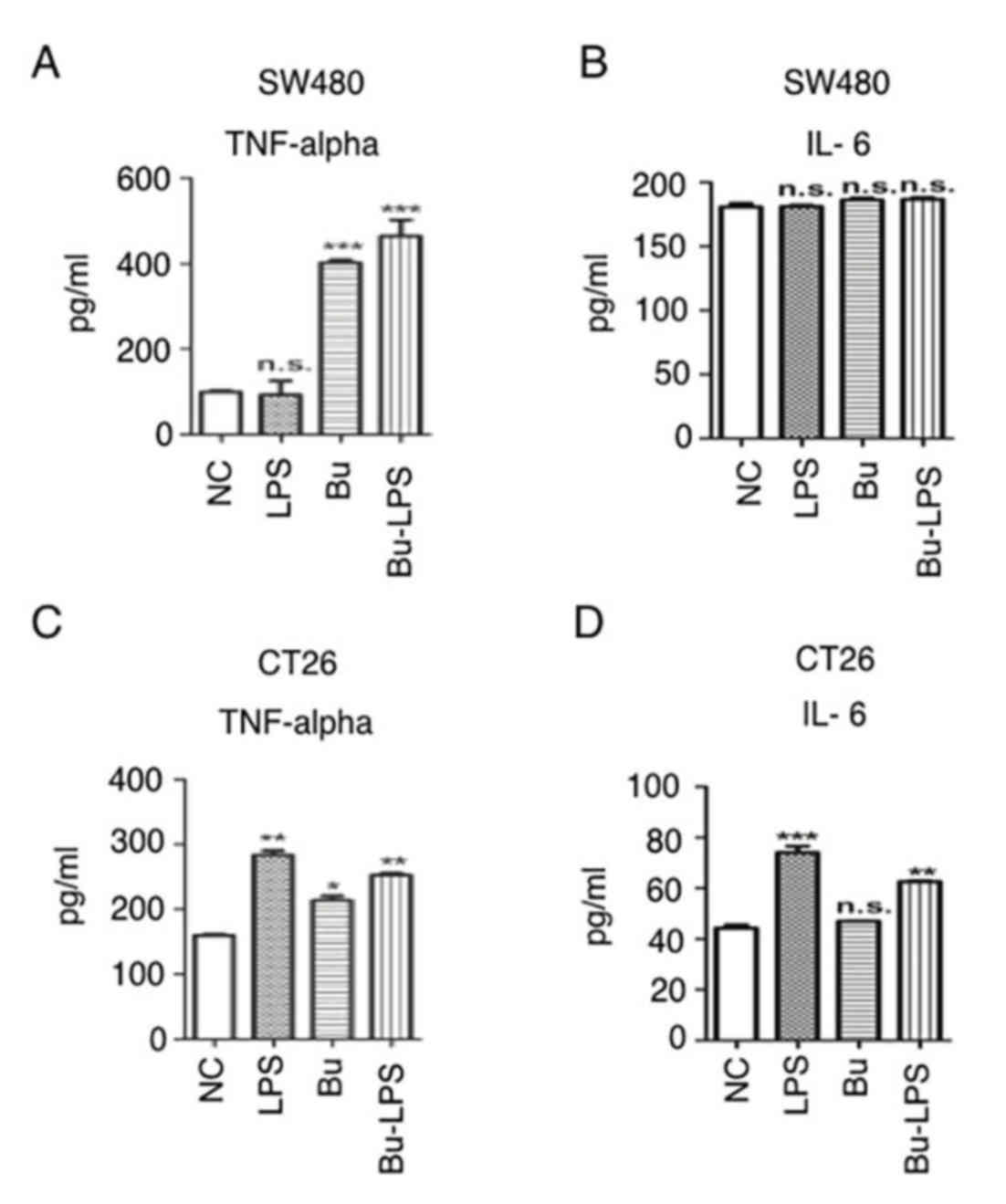 | Figure 6.Butyrate induces inflammatory
cytokine secretion in colon cancer cells. (A) SW480 cells were
cultured and treated with/without butyrate and/or LPS for 24 h, and
the supernatants were collected for inflammatory cytokine detection
using a TNF-α ELIA kit. (B) SW480 cells were cultured and treated
with/without butyrate and/or LPS for 24 h, and the supernatants
were collected for inflammatory cytokine detection using an IL-6
ELISA kit. (C) CT26 cells were cultured and treated with/without
butyrate and/or LPS for 24 h, and the supernatants were collected
for inflammatory cytokine detection using a TNF-α ELISA kit. (D)
CT26 cells were cultured and treated with or without butyrate
and/or LPS for 24 h, and the supernatants were collected for
inflammatory cytokine detection using an IL-6 ELISA kit. The
protein levels in the different groups were statistically analyzed.
These experiments were repeated ≥3 times, and representative data
are presented as the mean ± standard deviation. *P<0.05,
**P<0.01 and ***P<0.001. TNF-α, tumor necrosis factor α;
IL-6, interleukin 6; Bu, butyrate; NC, negative control; LPS,
lipopolysaccharide; NS, not significant. |
Discussion
Microbiota and its induced inflammation in
colorectal mucosa are considered risk factors for the development
of colorectal carcinogenesis (8,37,38), but the potential molecular mechanisms
are yet to be elucidated. Previous evidence suggests that
microbiota-derived molecular mediators, including SCFAs, regulate
the metabolic and immune homeostasis in the colon (39). In the present study, by using culture
cells in vitro, it was identified that butyrate not only
induced apoptotic cancer cell production, but also promoted TNF-α
secretion and enhanced the TLR4 signaling pathway in SW480 and CT26
cells. So, in the pathological condition in colon cancer, butyrate
exhibits a function in upregulating the TLR4 sensors on cancer
cells, and then initiating bacterial mediated innate immunity to
maintain intestinal homeostasis.
Previous studies have suggested that butyrate exerts
different anti-proliferative properties in normal colonocytes and
colon cancer cells by altering cellular metabolism (40,41). For
normal colonic epithelia cells, butyrate is the primary energy
source and promotes the turnover of the colonic epithelium to
maintain normal homeostasis of the gut (42). However, cancerous colonocytes undergo
the Warburg effect and the preferred energy source for these cells
is glucose, not butyrate (42).
Therefore, accumulated butyrate functions as an HDAC inhibitor to
promote cell arrest and induce apoptosis (40,41). This
butyrate-mediated growth inhibition in colon cancer cells is
associated with the direct silencing of HDAC3 expression, the
repression of Cyclin B or the upregulation of cyclin-dependent
kinase inhibitor 1 (43,44). Furthermore, the present study
demonstrated that the activation of caspase-3 contributed to
butyrate-induced apoptosis in SW480 and CT26 colon cancer cells. In
accordance with the results in the present study, Zhang et
al (45) previously demonstrated
that butyrate effectively induces human colon cancer apoptosis
through JNK MAPK activation, characterized by an increased B-cell
lymphoma 2 (Bcl-2)-associated X protein (Bax)-to-Bcl-2 expression
ratio and caspase cascade activation. An additional study indicated
that microRNA transcription regulation or autophagy induction also
contributed to butyrate-mediated colon cancer cell arrest and
apoptosis (46). Therefore, the
inhibitory effects of butyrate on the growth and apoptosis of colon
cancer cells may depend on multiple molecular mechanisms.
In the gut, the expression, localization and
signaling of TLR4 on colonic epithelia are developmentally
regulated in a compartmentalized manner (47). Compared with the high expression on
postnatal day 1, decreased levels of TLR4 were detected in crypts
in the mature colon of humans and mice (48). These data indicate that the expression
of TLR4 on intestinal epithelial cells is closely associated with
the flora community in the intestine. Although data concerning the
expression levels of TLR4 on colon cells are contradictory, they
indicate an association between the TLR4 signaling pathway and the
development of colon cancer (49–51). The
present study identified that the expression levels of TLR4 on
human colon cancer SW480 cells and mouse colon cancer CT26 cells
were low (Fig. 3), but were
significantly increased following butyrate treatment. Collectively,
these data indicated that butyrate modified the immune
microenvironment in the intestinal tract by regulating the
expression of TLR4 and its signaling pathway in the colon
epithelial or colorectal cancer cells.
The myeloid differentiation primary response gene 88
(MyD88)-dependent and MyD88-independent pathways are initiated in
the TLR4 signaling pathway (52). LPS
treatment alone is not able to induce activation of the TLR4
signaling pathway, and no differences in the phosphorylation of
different MAPKs (ERK, p38 and JNK) and NF-κB p65 were identified
between the control group and LPS-treated SW480 cells (Fig. 4). However, the LPS treatment alone
significantly induced the phosphorylation of p38 and JNK in CT26
cells, although no difference was observed in ERK and p65 in the
control and LPS-treated groups. These different responses to
treatment with LPS alone may be caused by the different original
expression levels of TLR4 on SW480 and CT26 cells.
There are conflicting data about the association
between SCFAs and TLRs in the literature. Certain studies have
suggested that butyrate exhibits anti-inflammatory effects by
downregulating the TLR4-dependent signaling cascade and the
secretion of inflammatory cytokines, but others demonstrated its
function in promoting inflammatory cytokines (53,54). The
present study demonstrated that if butyrate or butyrate + LPS were
used, the expression levels of TLR4 on SW480 and CT26 cells were
significantly upregulated, and the protein levels of phosphorylated
ERK, p38, JNK and NF-κB p65 were increased. Similarly, Alva-Murillo
et al (55) identified that
butyrate induced p38 phosphorylation and improved antimicrobial
defense in bovine mammary epithelial cells. Butyrate also increased
TLR4 expression and the NF-κB response to TLR ligand stimulation in
human L-cells, HEK293 or HeLa epithelial cells (56,57). In
the present study, although butyrate upregulated TLR4 expression,
MAPKs and NF-κB phosphorylation in SW480 and CT26 cells, only an
increase in TNF-α production was induced and no change of IL-6
expression was detected (Fig. 5).
Similarly, this specific effect of butyrate on the expression of
different inflammatory cytokine genes was also identified in bovine
mammary epithelial cells (55). The
regulatory effects of butyrate on the production and functions of
TNF-α were previously explored in colonocytes (58,59).
Through interacting with TNF-α, butyrate induced higher NF-κB
activities in adenocarcinoma HT-29 and fetal FHC human colon cells
in vitro (59). Even in the
same types of cells, butyrate exhibited counter-regulatory effects
on TNF-α-induced complement and inflammatory protein production,
which were associated with the modulation of transcription factor
activation (58,59). Therefore, combined with the data in
the present study, it is suggested that the butyrate mediated
pro-inflammatory response may involve multiple mechanisms of
action.
In summary, butyrate inhibited the growth of colon
cancer cells, but also promoted the TLR4 expression, MAPKs and
NF-κB phosphorylation in these cells, and then induced microbe
mediated innate immunity to maintain microenvironment homeostasis
in the gut. Although, the potential mechanisms of butyrate in
intestinal tolerance and colon cancer development remain to be
elucidated, the findings of the present study indicated the
potential usage of butyrate in the treatment of colorectal
cancer.
Acknowledgements
Thanks for all members in Shao Lab and Zhou Lab of
School of Medicine, Jiangsu University (Zhenjiang, China) for
helpful discussion and technical assistance. Thanks to the
technician in the Clinical Lab of the affiliated hospital of
Jiangsu University for the flow cytometric analysis.
Funding
The present study was funded by grants from the
National Natural Science Foundation of China (grant nos., 31570879
and 31428006) and the Key Research and Development Program of
Jiangsu Province, China (grant no., BE2017696).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TX and SW performed the majority of experiments and
analyzed the data. CY and CZ were responsible for the culture of
cell lines. NY and JX performed the flow cytometric analysis for
the phenotype and the apoptosis of the cells. HJ, CL and YW
conducted the ELISA analysis and the western blotting. SX and QS
designed the experiments. SX analyzed the data and wrote the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Bu
|
butyrate
|
|
CRC
|
colorectal cancer
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
HDACs
|
histone deacetylases
|
|
IL
|
interleukin
|
|
JNK
|
c-Jun NH2-terminal kinase
|
|
LPS
|
lipopolysaccharide
|
|
MAPK
|
mitogen-activated protein kinase
|
|
NF-κB
|
nuclear factor-κB
|
|
SCFA
|
short-chain fatty acids
|
|
TLR
|
toll-like receptor
|
|
PRRs
|
pathogen recognition receptors
|
|
TNF
|
tumor necrosis factor
|
References
|
1
|
Center MM, Jemal A and Ward E:
International trends in colorectal cancer incidence rates. Cancer
Epidemiol Biomarkers Prev. 18:1688–1694. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Center MM, Jemal A, Smith RA and Ward E:
Worldwide variations in colorectal cancer. CA Cancer J Clin.
59:366–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eaden JA, Abrams KR and Mayberry JF: The
risk of colorectal cancer in ulcerative colitis: A meta-analysis.
Gut. 48:526–535. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hagemann T, Balkwill F and Lawrence T:
Inflammation and cancer: A double-edged sword. Cancer Cell.
12:300–301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei EK, Giovannucci E, Wu K, Rosner B,
Fuchs CS, Willett WC and Colditz GA: Comparison of risk factors for
colon and rectal cancer. Int J Cancer. 108:433–442. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tomkovich S, Yang Y, Winglee K, Gauthier
J, Mühlbauer M, Sun X, Mohamadzadeh M, Liu X, Martin P, Wang GP, et
al: Locoregional effects of microbiota in a preclinical model of
colon carcinogenesis. Cancer Res. 77:2620–2632. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tjalsma H, Boleij A, Marchesi JR and
Dutilh BE: A bacterial driver-passenger model for colorectal
cancer: Beyond the usual suspects. Nat Rev Microbiol. 10:575–582.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong SH, Zhao L, Zhang X, Nakatsu G, Han
J, Xu W, Xiao X, Kwong TNY, Tsoi H, Wu WKK, et al: Gavage of fecal
samples from patients with colorectal cancer promotes intestinal
carcinogenesis in germ-free and conventional mice.
Gastroenterology. 153(1621–1633): e62017.
|
|
9
|
Hold GL and Garrett WS: Gut microbiota.
Microbiota organization-a key to understanding CRC development. Nat
Rev Gastroenterol Hepatol. 12:128–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flemer B, Lynch DB, Brown JM, Jeffery IB,
Ryan FJ, Claesson MJ, O'Riordain M, Shanahan F and O'Toole PW:
Tumour-associated and non-tumour-associated microbiota in
colorectal cancer. Gut. 66:633–643. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zou S, Fang L and Lee MH: Dysbiosis of gut
microbiota in promoting the development of colorectal cancer.
Gastroenterol Rep (Oxf). 6:1–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ray D and Kidane D: Gut microbiota
imbalance and base excision repair dynamics in colon cancer. J
Cancer. 7:1421–1430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meng C, Bai C, Brown TD, Hood LE and Tian
Q: Human gut microbiota and gastrointestinal cancer. Genomics
Proteomics Bioinformatics. 16:33–49. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Frosali S, Pagliari D, Gambassi G,
Landolfi R, Pandolfi F and Cianci R: How the intricate interaction
among toll-like receptors, microbiota, and intestinal immunity can
influence gastrointestinal pathology. J Immunol Res.
2015:4898212015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fukata M, Hernandez Y, Conduah D, Cohen J,
Chen A, Breglio K, Goo T, Hsu D, Xu R and Abreu MT: Innate immune
signaling by toll-like receptor-4 (TLR4) shapes the inflammatory
microenvironment in colitis-associated tumors. Inflamm Bowel Dis.
15:997–1006. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fukata M, Shang L, Santaolalla R,
Sotolongo J, Pastorini C, España C, Ungaro R, Harpaz N, Cooper HS,
Elson G, et al: Constitutive activation of epithelial TLR4 augments
inflammatory responses to mucosal injury and drives
colitis-associated tumorigenesis. Inflamm Bowel Dis. 17:1464–1473.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pimentel-Nunes P, Gonçalves N,
Boal-Carvalho I, Afonso L, Lopes P, Roncon-Albuquerque R Jr, Soares
JB, Cardoso E, Henrique R, Moreira-Dias L, et al: Decreased
Toll-interacting protein and peroxisome proliferator-activated
receptor γ are associated with increased expression of Toll-like
receptors in colon carcinogenesis. J Clin Pathol. 65:302–308. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu T, Guo F, Yu Y, Sun T, Ma D, Han J,
Qian Y, Kryczek I, Sun D, Nagarsheth N, et al: Fusobacterium
nucleatum promotes chemoresistance to colorectal cancer by
modulating autophagy. Cell. 170(548–563): e162017.
|
|
19
|
Chung YH and Kim D: Enhanced TLR4
expression on colon cancer cells after chemotherapy promotes cell
survival and epithelial-mesenchymal transition through
phosphorylation of GSK3β. Anticancer Res. 36:3383–3394.
2016.PubMed/NCBI
|
|
20
|
Kuo WT, Lee TC and Yu LC: Eritoran
suppresses colon cancer by altering a functional balance in
toll-like receptors that bind lipopolysaccharide. Cancer Res.
76:4684–4695. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rafa H, Benkhelifa S, AitYounes S, Saoula
H, Belhadef S, Belkhelfa M, Boukercha A, Toumi R, Soufli I, Moralès
O, et al: All-trans retinoic acid modulates TLR4/NF-κB signaling
pathway targeting TNF-α and nitric oxide synthase 2 expression in
colonic mucosa during Ulcerative colitis and colitis associated
cancer. Mediators Inflamm. 2017:73532522017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Louis P, Hold GL and Flint HJ: The gut
microbiota, bacterial metabolites and colorectal cancer. Nat Rev
Microbiol. 12:661–672. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Koh A, De Vadder F, Kovatcheva-Datchary P
and Bäckhed F: From dietary fiber to host physiology: Short-chain
fatty acids as key bacterial metabolites. Cell. 165:1332–1345.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cummings JH, Pomare EW, Branch WJ, Naylor
CP and Macfarlane GT: Short chain fatty acids in human large
intestine, portal, hepatic and venous blood. Gut. 28:1221–1227.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wong JM, de Souza R, Kendall CW, Emam A
and Jenkins DJ: Colonic health: Fermentation and short chain fatty
acids. J Clin Gastroenterol. 40:235–243. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chisolm DA and Weinmann AS: Connections
between metabolism and epigenetics in programming cellular
differentiation. Annu Rev Immunol. 36:221–246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng L, Kelly CJ, Battista KD, Schaefer
R, Lanis JM, Alexeev EE, Wang RX, Onyiah JC, Kominsky DJ and Colgan
SP: Microbial-derived butyrate promotes epithelial barrier function
through IL-10 receptor-dependent repression of claudin-2. J
Immunol. 199:2976–2984. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mollica MP, Raso Mattace G, Cavaliere G,
Trinchese G, De Filippo C, Aceto S, Prisco M, Pirozzi C, Di Guida
F, Lama A, et al: Butyrate regulates liver mitochondrial function,
efficiency, and dynamics in insulin-resistant obese mice. Diabetes.
66:1405–1418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Astakhova L, Ngara M, Babich O, Prosekov
A, Asyakina L, Dyshlyuk L, Midtvedt T, Zhou X, Ernberg I and
Matskova L: Short chain fatty acids (SCFA) reprogram gene
expression in human malignant epithelial and lymphoid cells. PLoS
One. 11:e01541022016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fellows R, Denizot J, Stellato C, Cuomo A,
Jain P, Stoyanova E, Balázsi S, Hajnády Z, Liebert A, Kazakevych J,
et al: Microbiota derived short chain fatty acids promote histone
crotonylation in the colon through histone deacetylases. Nat
Commun. 9:1052018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Julien O and Wells JA: Caspases and their
substrates. Cell Death Differ. 24:1380–1389. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Inan MS, Rasoulpour RJ, Yin L, Hubbard AK,
Rosenberg DW and Giardina C: The luminal short-chain fatty acid
butyrate modulates NF-kappaB activity in a human colonic epithelial
cell line. Gastroenterology. 118:724–734. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Katzenmaier EM, André S, Kopitz J and
Gabius HJ: Impact of sodium butyrate on the network of
adhesion/growth-regulatory galectins in human colon cancer in
vitro. Anticancer Res. 34:5429–5438. 2014.PubMed/NCBI
|
|
34
|
Brubaker SW, Bonham KS, Zanoni I and Kagan
JC: Innate immune pattern recognition: A cell biological
perspective. Annu Rev Immunol. 33:257–290. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kitchens RL: Role of CD14 in cellular
recognition of bacterial lipopolysaccharides. Chem Immunol.
74:61–82. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang H, Young DW, Gusovsky F and Chow JC:
Cellular events mediated by lipopolysaccharide-stimulated toll-like
receptor 4. MD-2 is required for activation of mitogen-activated
protein kinases and Elk-1. J Biol Chem. 275:20861–20866. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mariani F, Sena P and Roncucci L:
Inflammatory pathways in the early steps of colorectal cancer
development. World J Gastroenterol. 20:9716–9731. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pesic M and Greten FR: Inflammation and
cancer: Tissue regeneration gone awry. Curr Opin Cell Biol.
43:55–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sivaprakasam S, Bhutia YD, Ramachandran S
and Ganapathy V: Cell-surface and nuclear receptors in the colon as
targets for bacterial metabolites and its relevance to colon
health. Nutrients. 9:pii: E856. 2017.PubMed/NCBI
|
|
40
|
Blouin JM, Penot G, Collinet M, Nacfer M,
Forest C, Laurent-Puig P, Coumoul X, Barouki R, Benelli C and
Bortoli S: Butyrate elicits a metabolic switch in human colon
cancer cells by targeting the pyruvate dehydrogenase complex. Int J
Cancer. 128:2591–2601. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bultman SJ: Molecular pathways:
Gene-environment interactions regulating dietary fiber induction of
proliferation and apoptosis via butyrate for cancer prevention.
Clin Cancer Res. 20:799–803. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hague A, Butt AJ and Paraskeva C: The role
of butyrate in human colonic epithelial cells: An energy source or
inducer of differentiation and apoptosis? Proc Nutr Soc.
55:937–943. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Archer SY, Johnson J, Kim HJ, Ma Q, Mou H,
Daesety V, Meng S and Hodin RA: The histone deacetylase inhibitor
butyrate downregulates cyclin B1 gene expression via a
p21/WAF-1-dependent mechanism in human colon cancer cells. Am J
Physiol Gastrointest Liver Physiol. 289:G696–G703. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wilson AJ, Byun DS, Popova N, Murray LB,
L'Italien K, Sowa Y, Arango D, Velcich A, Augenlicht LH and
Mariadason JM: Histone deacetylase 3 (HDAC3) and other class I
HDACs regulate colon cell maturation and p21 expression and are
deregulated in human colon cancer. J Biol Chem. 281:13548–13558.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang Y, Zhou L, Bao YL, Wu Y, Yu CL,
Huang YX, Sun Y, Zheng LH and Li YX: Butyrate induces cell
apoptosis through activation of JNK MAP kinase pathway in human
colon cancer RKO cells. Chem Biol Interact. 185:174–181. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hu S, Liu L, Chang EB, Wang JY and Raufman
JP: Butyrate inhibits pro-proliferative miR-92a by diminishing
c-Myc-induced miR-17-92a cluster transcription in human colon
cancer cells. Mol Cancer. 14:1802015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ortega-Cava CF, Ishihara S, Rumi MA,
Kawashima K, Ishimura N, Kazumori H, Udagawa J, Kadowaki Y and
Kinoshita Y: Strategic compartmentalization of Toll-like receptor 4
in the mouse gut. J Immunol. 170:3977–3985. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Meng D, Zhu W, Shi HN, Lu L, Wijendran V,
Xu W and Walker WA: Toll-like receptor-4 in human and mouse colonic
epithelium is developmentally regulated: A possible role in
necrotizing enterocolitis. Pediatr Res. 77:416–424. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang EL, Qian ZR, Nakasono M, Tanahashi T,
Yoshimoto K, Bando Y, Kudo E, Shimada M and Sano T: High expression
of Toll-like receptor 4/myeloid differentiation factor 88 signals
correlates with poor prognosis in colorectal cancer. Br J Cancer.
102:908–915. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lu CC, Kuo HC, Wang FS, Jou MH, Lee KC and
Chuang JH: Upregulation of TLRs and IL-6 as a marker in human
colorectal cancer. Int J Mol Sci. 16:159–177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Paarnio K, Väyrynen S, Klintrup K, Ohtonen
P, Mäkinen MJ, Mäkelä J and Karttunen TJ: Divergent expression of
bacterial wall sensing Toll-like receptors 2 and 4 in colorectal
cancer. World J Gastroenterol. 23:4831–4838. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rosadini CV and Kagan JC: Early innate
immune responses to bacterial LPS. Curr Opin Immunol. 44:14–19.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wu JL, Zou JY, Hu ED, Chen DZ, Chen L, Lu
FB, Xu LM, Zheng MH, Li H, Huang Y, et al: Sodium butyrate
ameliorates S100/FCA-induced autoimmune hepatitis through
regulation of intestinal tight junction and toll-like receptor 4
signaling pathway. Immunol Lett. 190:169–176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Iraporda C, Errea A, Romanin DE, Cayet D,
Pereyra E, Pignataro O, Sirard JC, Garrote GL, Abraham AG and Rumbo
M: Lactate and short chain fatty acids produced by microbial
fermentation downregulate proinflammatory responses in intestinal
epithelial cells and myeloid cells. Immunobiology. 220:1161–1169.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Alva-Murillo N, Medina-Estrada I,
Báez-Magaña M, Ochoa-Zarzosa A and López-Meza JE: The activation of
the TLR2/p38 pathway by sodium butyrate in bovine mammary
epithelial cells is involved in the reduction of Staphylococcus
aureus internalization. Mol Immunol. 68:(2 pt B). 445–455. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lin MY, de Zoete MR, van Putten JP and
Strijbis K: Redirection of epithelial immune responses by
short-chain fatty acids through inhibition of histone deacetylases.
Front Immunol. 6:5542015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Larraufie P, Doré J, Lapaque N and
Blottière HM: TLR ligands and butyrate increase Pyy expression
through two distinct but inter-regulated pathways. Cell Microbiol.
19:2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Andoh A, Fujiyama Y, Hata K, Araki Y,
Takaya H, Shimada M and Bamba T: Counter-regulatory effect of
sodium butyrate on tumour necrosis factor-alpha (TNF-alpha)-induced
complement C3 and factor B biosynthesis in human intestinal
epithelial cells. Clin Exp Immunol. 118:23–29. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hýzd'alová M, Hofmanová J, Pacherník J,
Vaculová A and Kozubík A: The interaction of butyrate with
TNF-alpha during differentiation and apoptosis of colon epithelial
cells: Role of NF-kappaB activation. Cytokine. 44:33–43. 2008.
View Article : Google Scholar : PubMed/NCBI
|