Introduction
Colorectal cancer (CRC) is the third most common
type of cancer and the fourth most common cause of
cancer-associated mortality worldwide (1). In recent years, the overall effects of
the environment, lifestyle factors (2), and inherited and acquired
genetic/epigenetic alterations (3–5) on CRC,
and the interactions between them have been clarified. Dysplastic
adenomas are the most common form of premalignant precursor lesions
(6). Fearon and Vogelstein (7) first suggested the concept of the
adenoma-carcinoma sequence as a genetic model for colorectal
tumorigenesis; they proposed that colorectal carcinoma develops
from adenoma through a series of genetic events, including genetic
mutation and the loss of tumor suppressor genes. However, the
epigenetic mechanism of tumorigenesis in colorectal carcinoma
remains unclear.
Polycomb group proteins are global repressors of
gene expression that bring about transcriptional suppression
epigenetically through the formation of polycomb repressor
complexes (PRCs), including PRC1 and PRC2 (8,9). Enhancer
of zeste homolog 2 (EZH2) is a catalytic subunit of PRC2 (10–13). In
addition to EZH2, PRC2 contains the non-catalytic subunits
embryonic ectoderm development and suppressor of zeste 12. EZH2
serves as a histone lysine methyltransferase that mediates the
trimethylation of lysine 27 of histone H3 (H3K27) to silence
expression of PRC2-target genes involved in lineage differentiation
(14,15). EZH2 is overexpressed in several types
of cancer and its expression is associated with aggressiveness and
poor prognosis in breast, prostate and ovarian cancer, and
cholangiocarcinoma (16–21). Previously, EZH2 was reported to be
associated with accelerated cell proliferation and malignant
progression in pancreatic intraductal papillary mucinous neoplasm
and pancreatic cancer via the gene silencing of cyclin-dependent
kinase inhibitor (CDKN) 1B (p27) expression (22), and in melanoma and pancreatic cancer
via repression of CDKN1A (p21) (23,24).
However, the role of EZH2 in the progression of colorectal cancer
has not yet been investigated. Thus, the purpose of the present
study was to examine the role of alterations in EZH2 expression in
colorectal cancer progression.
Patients and methods
Patients
Between April 2009 and March 2013, 209 patients with
colorectal adenoma or carcinoma in adenoma (CIA) were treated
endoscopically in the Department of Gastroenterology and
Hepatology, Graduate School of Medical Sciences, Kumamoto
University (Kumamoto, Japan), and 33 patients with colorectal
adenoma and 18 patients with CIA underwent surgical resection at
the Department of Gastroenterological Surgery, Graduate School of
Medical Sciences, Kumamoto University. Between April 2013 and March
2014, 43 patients with early colorectal carcinoma underwent
surgical resection at the Department of Gastroenterological
Surgery, Graduate School of Medical Sciences, Kumamoto University.
Only the patients who provided written informed consent for the use
of their resected tissues for the current study were enrolled. A
total of 110 patients were enrolled in the current study, including
70 with adenoma lesions, 22 with CIA lesions, and 43 with carcinoma
lesions. Certain patients had more than two adenoma lesions. A
total of 42 lesions were resected endoscopically and 68 lesions
were resected surgically. The clinicopathological features of the
110 patients are summarized in Table
I. The average age was 67.7 years (range, 34–88), 62 patients
were male and 48 patients were female.
 | Table I.Clinicopathological characteristics
of patients. |
Table I.
Clinicopathological characteristics
of patients.
|
| Tissue type |
|
|---|
|
|
|
|
|---|
| Variable | Adenoma (n=47) | Carcinoma in
adenoma (n=20) | Carcinoma
(n=43) | P-value |
|---|
| Sex |
|
|
| 0.23 |
|
Male | 22 | 14 | 26 |
|
|
Female | 25 | 6 | 17 |
|
| Age,
yearsa | 69±9.57 | 66±18.4 | 71.5±13.7 | 0.44 |
| Location |
|
|
| 0.77 |
|
C/A/T/D/S/Rb | 8/12/4/5/8/10 | 2/4/2/1/5/6 | 3/6/6/5/9/14 |
|
| Tumor depth |
|
Tis/T1 | – | – | 23/20 |
|
| Tumor diameter,
mma | 27.5±22.5 | 29±23.1 | 46±25.9 | 0.51 |
The present study protocol was approved by the
institutional review board of Kumamoto University Hospital. The
study was conducted in compliance with the Declaration of Helsinki
and all patients provided written informed consent for the use of
their resected tissues for clinical study.
Immunohistochemical (IHC)
staining
IHC staining was performed on 4 µm sections obtained
from formalin-fixed (at 36°C, 10% formalin, 48 h) paraffin-embedded
blocks. Sections were pretreated through autoclaving (15 min, at
121°C) in Histofine antigen retrieval solution (pH 9.0) (Nichirei
Biosciences, Inc., Tokyo, Japan) for EZH2 and proliferation marker
protein Ki-67 (Ki-67) IHC, or through microwaving for 20 min in
Histofine antigen retrieval (pH 9.0) for CDKN2A (p16), p21, and
p27. Endogenous peroxidase activity was blocked for 5 min at room
temperature using 3% hydrogen peroxidase, and the sections were
incubated with diluted antibodies overnight at 4°C.
EnVision+ solution (Dako; Agilent Technologies, Inc.,
Santa Clara, CA, USA) was then applied for 30 min at room
temperature. The reaction products were visualized with a
diaminobenzidine solution followed by counterstaining with Mayer's
hematoxylin for 5 min at room temperature. The following primary
antibodies were used: Mouse monoclonal antibody (mAb) against EZH2
(1:50 dilution; cat. no. 612666; BD Biosciences, San Jose, CA,
USA); mouse mAb against Ki-67 (1:100 dilution; cat. no. M7240;
Dako; Agilent Technologies, Inc.), rabbit polyclonal Ab against p16
(1:50 dilution; cat. no. 4824; CST Biological Reagents Company
Limited, Shanghai, China), rabbit mAb against p21 (1:50 dilution;
cat. no. sc-397; Santa Cruz Biotechnology, Dallas, TX, USA); and
mouse mAb against p27 (1:150 dilution; cat. no. 610241; BD
Biosciences, San Jose, CA, USA). The following secondary antibodies
were used: anti-mouse IgG goat antibody (cat. no. K4001; Dako;
Agilent Technologies, Inc.) for EZH2, Ki-67 and p27 and anti-rabbit
IgG goat antibody (cat. no. K4003; Dako; Agilent Technologies,
Inc.) for p16 and p21, applied for 30 min at room temperature.
Chromogenic reaction was visualized with a diaminobenzidine
solution (cat. no. 349-00903; Wako Pure Chemical Industries, Ltd.,
Osaka, Japan). Positive controls for immunostaining were performed
using pancreatic intraductal papillary mucinous neoplasm tissue and
negative controls were prepared by omission of the primary
antibody. We evaluated IHC staining using OLYMPUS DP27 microscope
(confocal type laser scan) and we examined 5 fields (magnification,
×100) using DP2-BSW (ver. 2.1, Olympus Life Science). IHC analysis
was conducted using a dual scoring system of staining intensity and
staining extent. The staining intensity was defined as 0, 1 or 2,
denoting negative, weak, or strong staining, respectively. The
staining extent was defined as the percentage of cancer cells that
were positively stained. IHC score was calculated as the product of
the staining intensity and staining extent by 2 researchers, M.O.
and Y.S.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was successfully extracted from 32
selected lesions (4 normal lesions, 12 adenomas, 9 CIA lesions and
7 carcinomas). Formalin-fixed paraffin-embedded blocks were
sectioned into 3–5 serial 10-µm slices. Randomly selected lesions
were macrodissected by comparison with hematoxylin and eosin
staining (5 min at room temperature). Total RNA was extracted using
an RNeasy FFPE kit (Qiagen Sciences, Inc., Gaithersburg, MD, USA)
according to the manufacturer's protocol and converted to cDNA by
reverse transcription using Oligo(dT)12-18 primer (cat.
no. 18418-012), random primer (cat. no. 48190-011), 10 mM dNTP mix
(cat. no. 18427-088), SuperScriptTMIII Reverse Transcriptase (cat.
no. 18080-085) and RNaseOUTTM Recombinant Ribonuclease Inhibitor
(cat. no. 10777-019; all purchased from Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Random Hexamers Annealing was
performed for 5 min at 25°C, cDNA synthesis for 60 min at 50°C and
termination reaction for 15 min at 70°C. To determine the mRNA
expression levels of EZH2, p27, p21 and p16, a qPCR assay was
performed using a LightCycler 480 system (Roche Applied
Diagnostics, Basel, Switzerland) under the following reaction
conditions: Initial denaturation at 95°C for 10 min, 10 sec at
95°C, 30 sec at 60°C, 1 sec at 72°C and 10 sec at 40°C using
LightCycler 480 Probe Master (cat. no. 04-887-301-001, Roche Life
Science, Basel, Switzerland) and the 2-∆∆Cq method was used
(25). Probes and primers were
designed using the Roche probe library system (Table II). β-actin was used as a
reference.
| Table II.Primer sequences for polymerase chain
reaction analysis. |
Table II.
Primer sequences for polymerase chain
reaction analysis.
|
| Primer |
|---|
|
|
|
|---|
| Gene | Forward | Reverse |
|---|
| EZH2 |
GACTGGCGAAGAGCTGTTTT |
TVTTTVGATGCCGACATACTT |
| p21 |
GGCAGACCAGCATGACAGATT |
GCGGATTAGGGCTTCCTCTT |
| p16 |
TCGTGCTGATGCTACTGAGG |
ATCTATGCGGGCATGGTTAC |
| p27 |
GAGGTGGAGAGGGGCAGC |
TTCGGGGAACCGTCTGAAAC |
Statistical analysis
All statistical analysis was performed using
JMP® 10.0 software (SAS Institute Inc., Cary, NC, USA).
P<0.05 was considered to indicate a statistically significant
difference. Data are presented as the mean ± standard deviation and
analysis was performed 5 times per experiment. In comparison of
clinicopathological variables between tissue types, χ2
test and Fisher's exact test were used. In comparison of EZH2,
Ki-67, p21 expression score, unpaired Student's t-test was used. In
evaluation of association between EZH2, Ki-67 and p21, Pearson's
product-moment correlation coefficient was used. Following
univariate analysis, variables with a P<0.05 were selected for
multivariate analysis.
Results
EZH2 expression increases with
increased cell proliferative activity
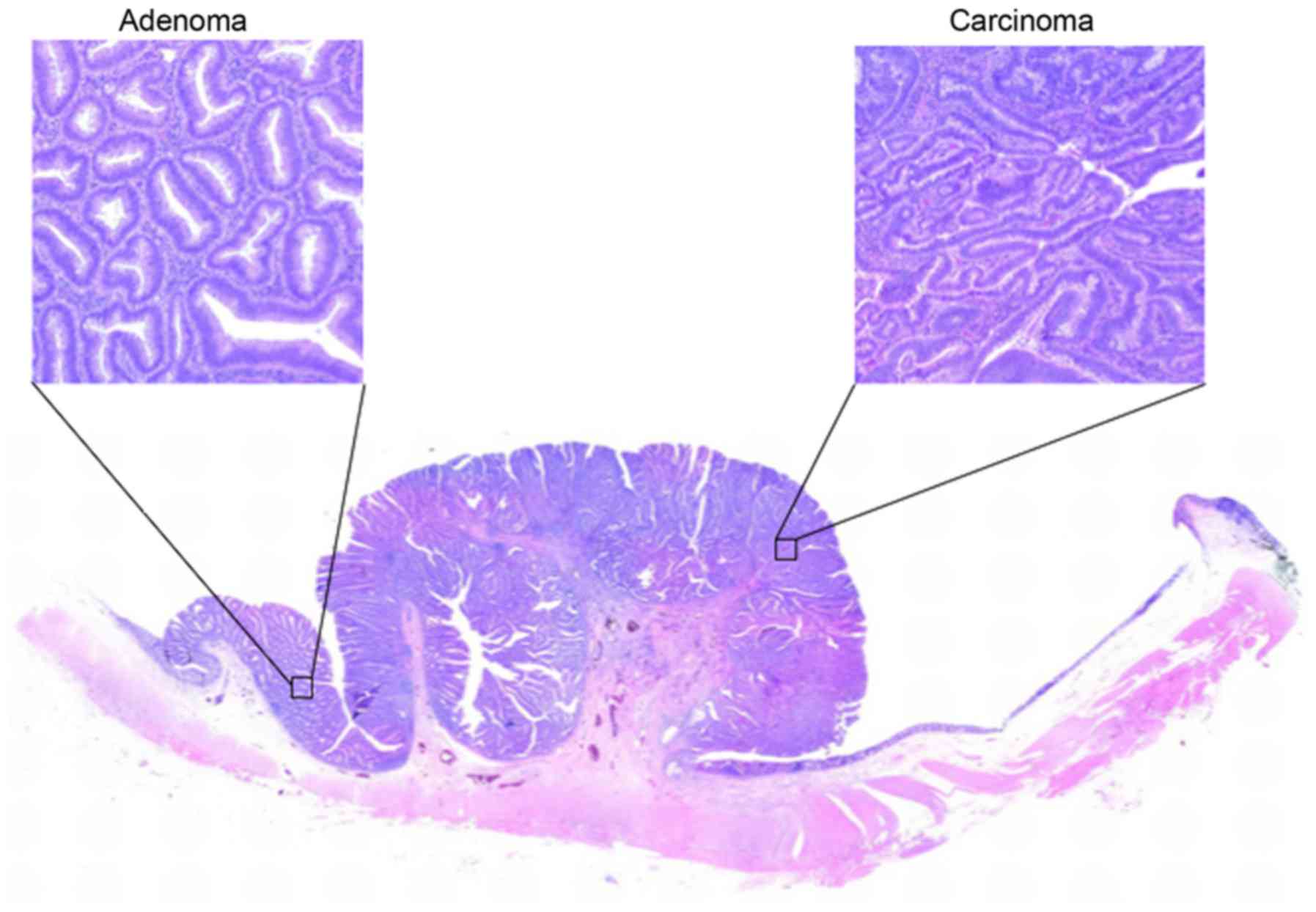
Carcinoma and adenoma cells are often present in the
same lesion (Fig. 1); this is
regarded as theoretical proof of the adenoma-carcinoma
sequence.
First, IHC staining and RT-qPCR were
performed for EZH2
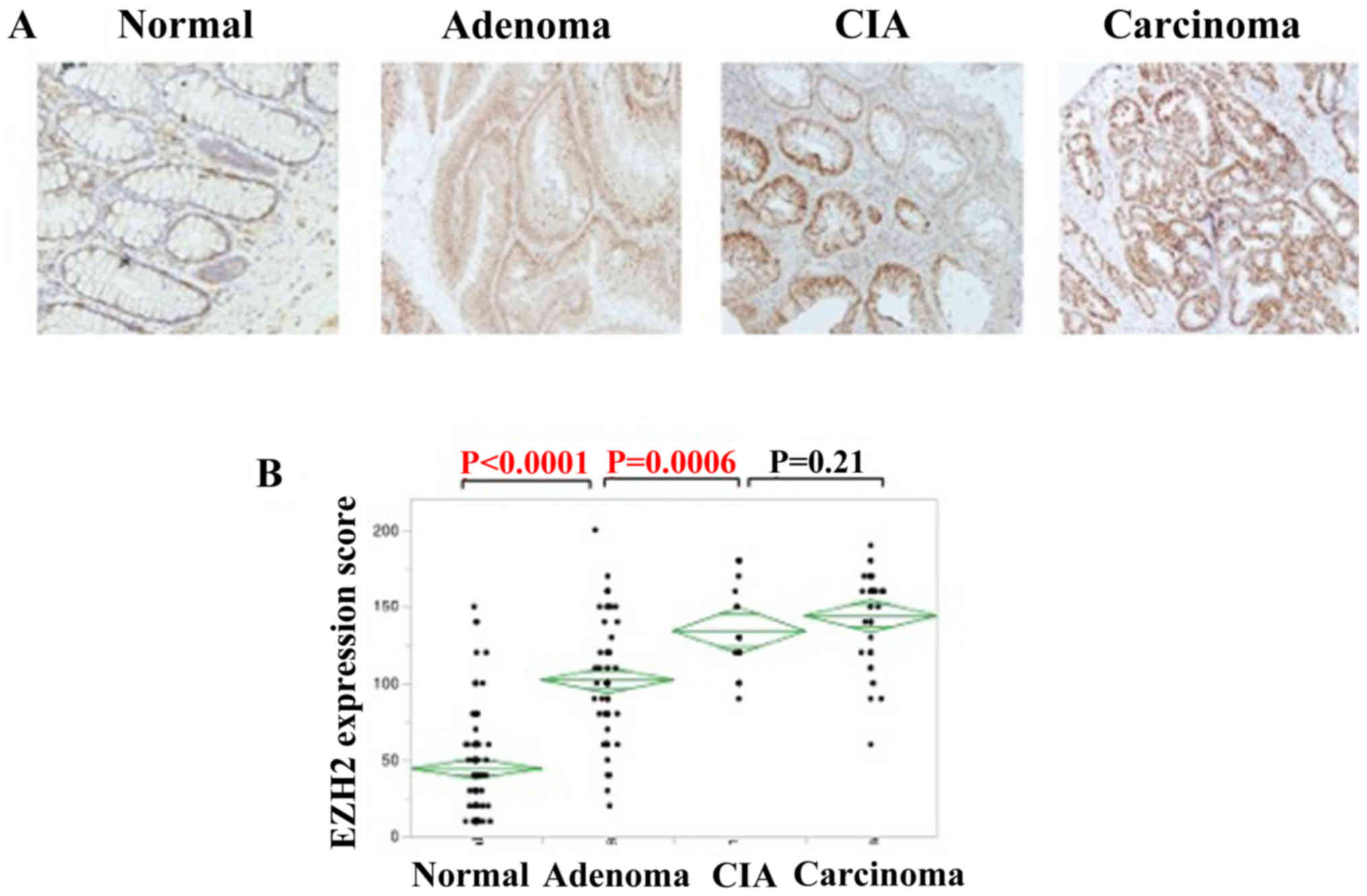
EZH2 expression was detected in the nuclei of
colorectal adenomas, CIA lesions and colorectal adenocarcinomas
(Fig. 2A). IHC staining was used to
analyze EZH2 expression. The IHC score increased parallel to
pathological changes among normal tissue, adenoma and carcinoma.
The expression of EZH2 was significantly higher in colorectal
adenomas compared with that of normal lesions (P<0.0001), and
also higher in CIA lesions compared with colorectal adenomas
(P=0.0006); however, no significant association was identified in
EZH2 expression between CIA and colorectal carcinomas (Fig. 2B).
Ki-67 expression associates with EZH2
expression
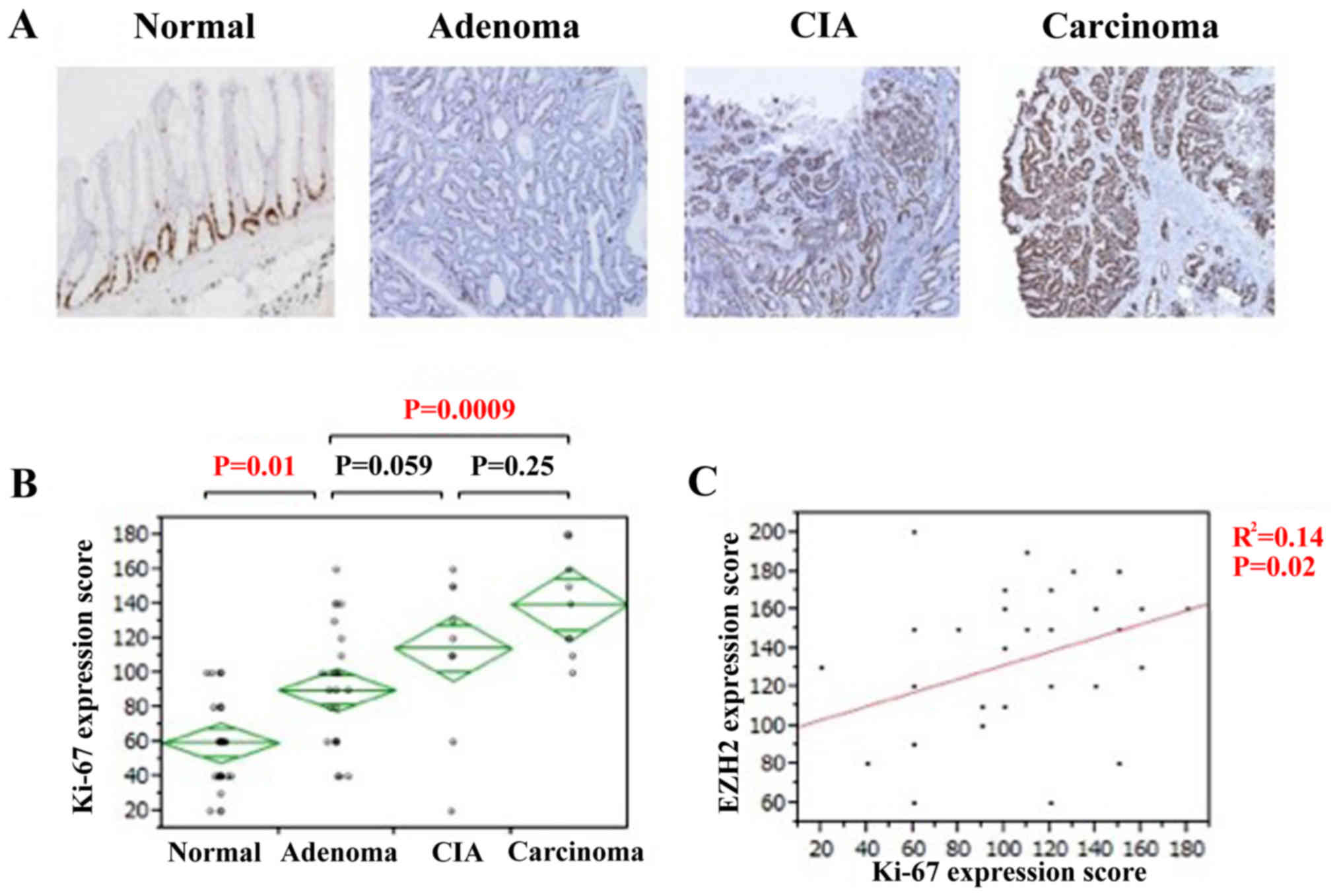
Proliferative activity was assessed using Ki-67
immunostaining. Ki-67 expression was also observed in the nuclei of
all tissue types (Fig. 3A). The IHC
score of Ki-67 gradually increased along with the pathological
change from normal tissue to adenoma and carcinoma. The expression
of Ki-67 was significantly higher in colorectal adenomas compared
with that of normal lesions (P=0.01; Fig.
3B). Although no significant association was identified in
Ki-67 expression between colorectal adenomas and CIA lesions,
expression was significantly higher in colorectal carcinomas
compared with adenoma lesions (P=0.0009; Fig. 3B). There was a significant positive
association identified between EZH2 and Ki-67 expression (P=0.02;
Fig. 3C). Therefore, the
proliferative activity of the cell increased during the disease
progression of colorectal adenocarcinomas, particularly during the
change from normal to adenoma, and it is likely that EZH2 serves a
notable role in the adenoma-carcinoma sequence of colorectal
adenocarcinoma.
Tumor suppressor p21 is downregulated
with the progression of colorectal carcinomas
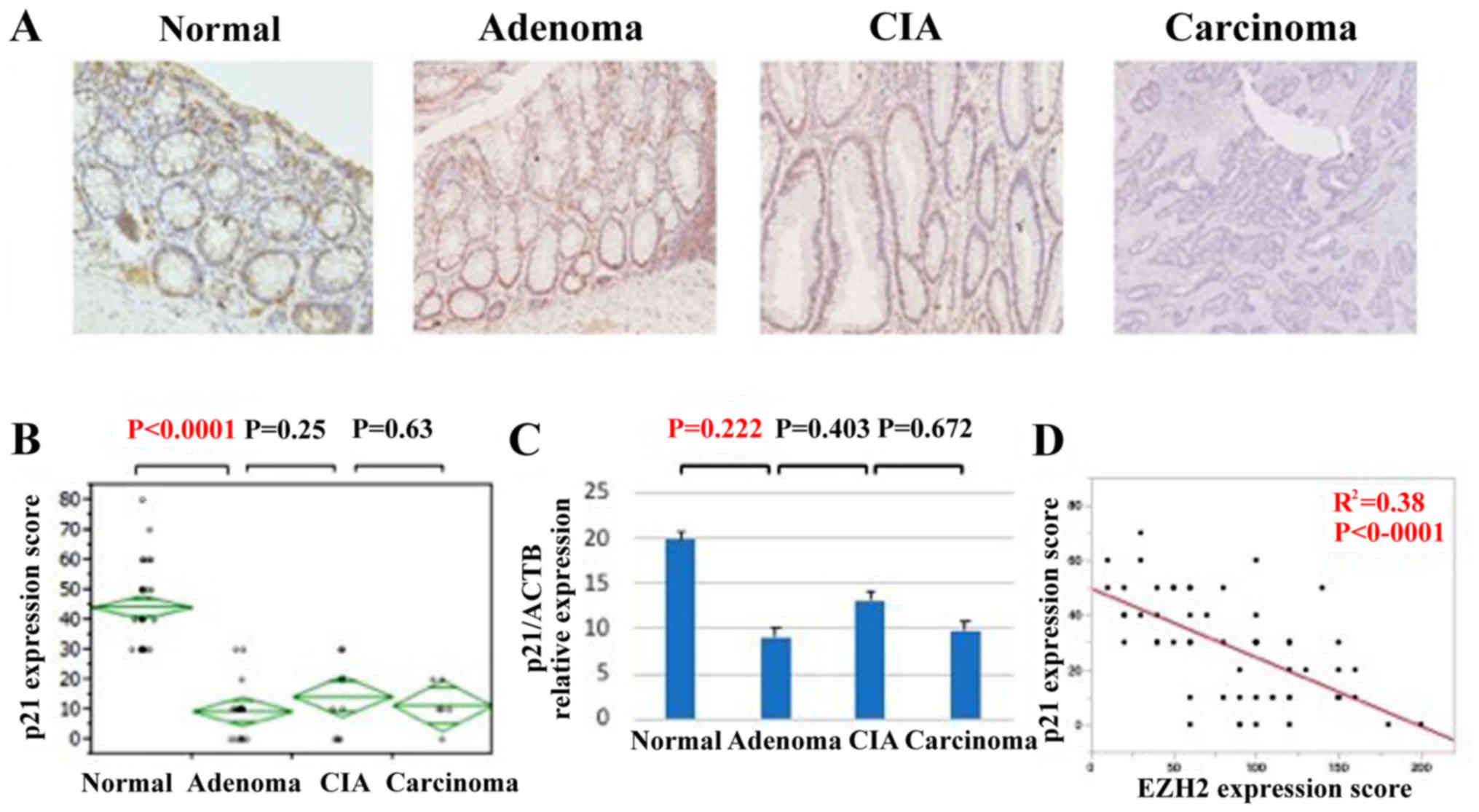
EZH2 has been reported to promote cellular
proliferation via downregulation of tumor suppressor proteins,
including p27, p21, and p16 (22–25). To
assess the mechanism of this accelerated cell proliferation during
the adenoma-carcinoma sequence of colorectal cancer, IHC staining
and qPCR analyses of p27, p21, and p16 expression were performed
(Fig. 4). Whereas p21 exhibited
strong nuclear localization in normal tissue, its expression was
downregulated in adenomas, CIA lesions and carcinomas (Fig. 4A). The IHC score was significantly
lower in adenomas compared with that of normal tissue (Fig. 4B). Similarly, p21 expression at the
mRNA level tended to be significantly lower in adenomas, CIA
lesions, and carcinomas compared with in normal lesions (Fig. 4C). EZH2 and p21 expression was
identified to be significantly inversely associated (Fig. 4D). However, the expression of p27 and
p16 did not change significantly during the progression of
colorectal carcinoma (data not shown). Thus, the downregulation of
p21 appears to be an important factor in accelerated cell
proliferation during colorectal cancer progression.
Next, as p21 is a well-known transcriptional target
of p53 (26), p53 expression was
assessed via IHC staining. Although the IHC score of p53
significantly increased with the pathological change from normal to
adenoma and carcinoma, no significant association was identified
between p21, and p53 expression (data not shown).
Discussion
The present study demonstrated that increased EZH2
expression was associated with the progression of colorectal
cancer; concomitantly, expression of the tumor suppressor protein
p21 was decreased.
Tumorigenesis is a multi-step process associated
with genetic and epigenetic alterations. In 1990, Fearon et
al and Vogelstein et al reviewed the genetic model for
colorectal tumorigenesis (7,27). Allelic losses of chromosome 5q, which
is associated with the gene for familial adenomatous polyposis,
have been observed in ~30% of colorectal adenomas in patients
without polyposis (28,29), suggesting that this allelic loss is an
early event in colorectal tumor progression. In addition,
Vogelstein et al (27)
revealed that mutations in RAS were observed in intermediate and
late adenoma, whereas allelic loss of chromosome 17p, which was
associated with the TP53 gene, was observed in late adenoma
carcinoma. These allelic losses induce other alterations, including
tumor suppressor gene hypomethylation (7), which has also been identified to occur
early in colorectal tumorigenesis; in one study, ~1/3 of the DNA
regions had lost methyl groups, even in extremely small adenomas
(27). Thus, genetic and epigenetic
alterations occur in the early stage of colorectal tumorigenesis.
In the present study, EZH2 expression was increased in adenomas,
suggesting that EZH2 expression is an early event in colorectal
tumorigenesis. This finding suggests that EZH2 overexpression may
be involved in epigenetic alterations that occur early in the
progression from normal epithelial cells to adenoma.
In the present study, of the tumor suppressor
proteins p27, p21 and p16, only p21 was expressed at higher levels
in normal tissue than in cancerous tissue; its expression was
inversely associated with EZH2 expression and no significant
association was identified between p21 expression and p53
expression. Moreover, expression of EZH2 correlated with the
expression of Ki-67. p21 is a mediator of p53 tumor suppressor
activity and inhibits cell-cycle progression by inhibiting the
activity of cyclin-dependent kinase-cyclin complexes and
proliferating cell nuclear antigen (30). It has been reported that p21 is
stimulated by multiple pathways that are independent of p53
(30). The findings of the present
study suggest that EZH2-mediated acceleration of cell proliferation
via downregulation of p21 may serve an essential role in the
progression of normal cells to adenoma, and that p21 is regulated
not by p53 but by EZH2 in the progression of colorectal cancer. A
reduction in the expression of p21 has been associated with the
progression of colorectal cancer (31,32).
Bukholm and Mesland (31) reported an
association between reduced/absent p21 expression, and the
development of metastases and mortality owing to cancer disease.
Furthermore, Pasz-Walczak et al (32) reported an inverse correlation between
p21 expression and clinical stage. In addition, Hubaux et al
(33) revealed that inhibiting EZH2
using short hairpin RNA increased p21 protein levels in small cell
lung cancer (33) and pancreatic
ductal adenocarcinoma (34). It was
proposed that the mechanism underlying the regulation of p21
expression by EZH2 may involve downregulation of runt-related
transcription factor 3 (RUNX3), a transactivator of p21, by EZH2
via trimethylation of H3K27 in the promoter region (35). Chi et al (36) revealed that RUNX3 suppresses cell
growth by inducing p21 expression in gastric cancer. However,
Kodach et al (37) reported
that knockdown of EZH2 did not result in RUNX3 re-expression in
colorectal cancer cell lines. The authors demonstrated that the
downregulation of RUNX3 was associated with RUNX3 DNA methylation
and that knockdown of EZH2 prevented the re-silencing of RUNX3
following the removal of demethylating agents (37). Thus, further studies are required to
clarify the association between EZH2 and p21 expression.
A notable limitation of the present study is the
small number of samples due to the single-institutional study
design. In particular, CIA samples were relatively rare compared
with adenomas or carcinomas. Further studies including a larger
number of samples are therefore required. In conclusion, the
results of the present study suggest that EZH2 upregulation serves
an important role in the progression of colorectal cancer via p21
downregulation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YSak, YM and HB designed and directed the project.
MO performed the experiments and collected all the
clinicopathological data. YSak, RT, YK, KN, DI, KK, KH, JK, MI, YB
and NY supported the experiments. TS, HN and YSas assisted with the
collection of clinicopathological data. MO wrote the paper and
YSak, YM and HB revised the manuscript and agreed that this study
was appropriately investigated and to be accountable for all
aspects of the study. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study protocol was approved by the
institutional review board of Kumamoto University Hospital. All
patients provided written informed consent for the use of their
resected tissues for clinical study.
Patient consent for publication
Patients provided their consent for the publication
of this data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khandekar MJ, Cohen P and Bruce M:
Molecular mechanisms of cancer development in obesity. Nat Rev
Cancer. 11:886–895. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saif MW and Chu E: Biology of colorectal
cancer. Cancer J. 16:196–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Issa JP: Colon cancer: It's CIN or CIMP.
Clin Cancer Res. 14:5939–5940. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jass JR: Classification of colorectal
cancer based on correlation of clinical, morphological and
molecular features. Histopathology. 50:113–130. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fearon ER and Vogelstein B: A genetic
model for colorectal tumorigenesis. Cell. 61:759–767. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alkema MJ, Bronk M, Verhoeven E, Otte A,
van't Veer LJ, Berns A and van Lohuizen M: Identification of
Bmi1-interacting proteins as constituents of a multimeric mammalian
polycomb complex. Gene Dev. 11:226–240. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kuzmichev A, Nishioka K,
Erdjument-Bronmage, Tempst P and Reinberg D: Histone
methyltransferase activity associated with a human multiprotein
complex containing the Enhancer of Zeste protein. Gene Dev.
16:2893–2905. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ketel CS, Andersen EF, Vargas ML, Suh J,
Strome S and Simon JA: Subunit contributions to histone
methyltransferase activities of fly and worm polycomb group
complexes. Mol Cell Biol. 25:6857–6868. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schwartz YB, Kahn TG, Nix DA, Li XY,
Bourgon R, Biggin M and Pirrotta V: Genome-wide analysis of
Polycomb targets in Drosophila melanogaster. Nat Genet. 38:700–705.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schuettengruber B, Ganapathi M, Leblanc B,
Portoso M, Jaschek R, Tolhuis B, van Lohuizen M, Tanay A and
Cavalli G: Functional anatomy of polycomb and trithorax chromatin
landscapes in Drosophila embryos. PLoS Biol. 7:e132009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Boyer LA, Piath K, Zeitlinger J, Brambrink
T, Medeiros LA, Lee TI, Levine SS, Wernig M, Tajonar A, Ray MK, et
al: Polycomb complexes repress developmental regulators in murine
embryonic stem cells. Nature. 441:349–353. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cao R and Zhang Y: The functions of
E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr
Opin Genet Dev. 14:155–164. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simon JA and Lange CA: Roles of the EZH2
histone methyltransferase in cancer epigenetics. Mutat Res.
647:21–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kleer CG, Cao Q, Varambally S, Shen R, Ota
I, Tomlins SA, Ghosh D, Sewalt RG, Otte AP, Hayes DF, et al: EZH2
is a marker of aggressive breast cancer and promotes neoplastic
transformation of breast epithelial cells. Proc Natl Acad Sci USA.
100:11606–11611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bachmann IM, Halvorsen OJ, Collett K,
Stefansson IM, Straume O, Haukaas SA, Salvesen HB, Otte AP and
Akslen LA: EZH2 expression is associated with high proliferation
rate and aggressive tumor subgroups in cutaneous melanoma and
cancers of the endometrium, prostate, and breast. J Clin Oncol.
24:268–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Varambally S, Dhanasekaran SM, Ahou M,
Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt
RG, Otte AP, et al: The polycomb group protein EZH2 is involved in
progression of prostate cancer. Nature. 419:624–629. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saramäki OR, Tammela TL, Martilkainen PM,
Vessella RL and Visakorpi T: The gene for polycomb group protein
enhancer of zeste homolog 2 (EZH2) is amplified in late-stage
prostate cancer. Gene Chromosome Canc. 45:639–645. 2006. View Article : Google Scholar
|
|
20
|
Li H, Cai Q, Godwin AK and Zhang R:
Enhancer of zeste homolog 2 promotes the proliferation and invasion
of epithelial ovarian cancer cells. Mol Cancer Res. 8:1610–1618.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakagawa S, Okabe H, Sakamoto Y, Hayashi
H, Hashimoto D, Yokoyama N, Sakamoto K, Kuroki H, Mima K, Nitta H,
et al: Enhancer of zeste homolog 2 (EZH2) promotes progression of
cholangiocarcinoma cells by regulating cell cycle and apoptosis.
Ann Surg Oncol. 20 Suppl 3:S667–S675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuroki H, Hayashi H, Okabe H, Hashimoto D,
Takamori H, Nakahara O, Nakagawa S, Fukushima Y, Chikamoto A, Beppu
T, et al: EZH2 is associated with malignant behavior in pancreatic
IPMN via p27Kip1 downregulation. PLoS One. 9:e1009042014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan T, Jiang S, Chung N, Alikhan A, Ni C,
Lee CC and Hornyak TJ: EZH2-dependent suppression of a cellular
senescence phenotype in melanoma cells by inhibition of p21/CDKN1A
expression. Mol Cancer Res. 9:418–29. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ougolkov AV, Billim V and Billadeau DD:
Regulation of pancreatic tumor cell proliferation and
chemoresistance by the histone methyltransferase enhancer of zeste
homologue 2. Clin Cancer Res. 14:6790–6796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Abbas T and Dutta A: p21 in cancer,
intricate networks and multiple activities. Nat Rev Cancer.
9:400–414. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vogelstein B, Fearon ER, Hamilton SR, Kern
SE, Preisinger AC, Leppert M, Nakamura Y, White R, Smits AM and Bos
JL: Genetic alterations during colorectal-tumor development. N Eng
J Med. 319:525–532. 1988. View Article : Google Scholar
|
|
28
|
Sasaki M, Okamoto M, Sato C, Sugio K,
Soejima J, Iwama T, Ikeuchi T, Tonomura A, Miyaki M and Sasazuki T:
Loss of constitutional heterozygosity in colorectal tumors from
patients with familial polyposis coli and those with nonpolyposis
colorectal carcinoma. Cancer Res. 49:4402–4406. 1989.PubMed/NCBI
|
|
29
|
Goelz SE, Vogelstein B, Jamilton SR and
Feinberg AP: Hypomethylation of DNA from benign and malignant human
colon neoplasms. Science. 228:187–190. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Abbas T and Dutta A: p21 in cancer:
Intricate networks and multiple activities. Nat Rev Cancer.
9:400–414. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bukholm IK and Mesland JM: Protein
expression of p53, p21 (WAF1/CIP1), bcl-2, Bax, cyclin D1 and pRb
in human colon carcinomas. Virchows Arch. 436:224–228. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pasz-Walczak G, Kordek R and Faflik M: P21
(WAF1) expression in colorectal cancer: Correlation sith P53 and
cyclin D1 expression, clinicopathological parameters and prognosis.
Pathol Res Pract. 197:683–689. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hubaux R, Thu KL, Coe BP, MacAulay C, Lam
S and Lam WL: EZH2 promotes E2F-driven SCLC tumorigenesis through
modulation of apoptosis and cell-cycle regulation. J Thorac Oncol.
8:1102–1106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Batchu RB, Qazi AM, Gruzdyn OV, Semaan A,
Seward SM, Chamala S, Dhulipala VB, Bouwman DL, Weaver DW and
Gruber SA: EZH2-shRNA-mediated upregulation of p21waf1/cip1 and its
transcriptional enhancers with concomitant downmodulation of mutant
p53 in pancreatic ductal adenocaricinoma. Surgery. 154:739–747.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yoo KH and Hennighausen L: EZH2
methyltransferase and H3K27 methylation in breast cancer. Int J
Biol Sci. 8:59–65. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chi XZ, Yang JO, Lee KY, Ito K, Sakakura
C, Li QL, Kim HR, Cha EJ, Lee YH, Kaneda A, et al: RUNX3 suppresses
gastric epithelial cell growth by inducing p21(WAF1/Cip1)
expression in cooperation with transforming growth factor
{beta}-activated SMAD. Mol Cell Biol. 25:8097–8107. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kodach LL, Jacobe RJ, Heijmans J, van
Noesel CJ, Langers AM, Verspaget HW, Hommes DW, Offerhaus GJ, van
den Brink GR and Hardwick JC: The role of EZH2 and DNA methylation
in the silencing of the tumour suppressor RUNX3 in colorectal
cancer. Carcinogenesis. 31:1567–1575. 2010. View Article : Google Scholar : PubMed/NCBI
|