Introduction
Prostate cancer is the most common non-cutaneous
malignancy in males. It is also the second leading cause of
cancer-associated mortality in the western world amongst males
(1). Androgen-deprivation therapy, a
standard-of-care medicine for prostate cancer, is able to
proficiently control the development of androgen-dependent tumors
(2). Numerous patients with localized
disease have improved long-term survival and elevated cure rates
with standard therapies; however, the cancer eventually becomes
resistant to hormone deprivation and progresses to
castration-resistant prostate cancer (2,3). The
patients with locally advanced and metastatic disease have poor
prognoses, leading to high morbidity and mortality rates (2,3).
Therefore, understanding the mechanisms underlying cancer invasion
and subsequent metastasis are urgently required to develop
therapies for combating metastatic prostate cancer.
Thiazolidinediones (TZDs), including troglitazone
and ciglitazone, are synthetic ligands of peroxisome
proliferator-activated receptor γ (PPARγ) (4). They exhibit potential antitumor effects
on numerous types of cancers (4–10),
including prostate cancer (11–13). A
previous study performed on prostate cancer cell lines has
demonstrated that troglitazone (TGZ) decreases cellular
proliferation, which is associated with increased expression levels
of glutathione peroxidase 3 (GPx3) (11). TGZ has been investigated in clinical
trials against breast, colorectal and prostate cancer (14–16).
Treatment with TGZ in patients with advanced prostate cancer has
been associated with long periods of stable disease characterized
by the absence of new symptoms without new metastases (14), suggesting that TGZ may have clinical
value in suppressing cancer metastasis. However, the role of TGZ in
metastasis and the precise molecular mechanisms underlying its
action have not been fully elucidated.
During cancer progression, epithelial-mesenchymal
transition (EMT) is the main mechanism underlying the invasiveness
and metastasis of cancer cells. EMT is a process that converts
immotile epithelial cells to motile mesenchymal cells (17–19).
Downregulation of epithelial marker E-cadherin expression is the
hallmark of the EMT process. E-cadherin is a tumor suppressor that
serves crucial roles in cell-cell adhesion (19). Loss of E-cadherin expression or
function is associated with cancer cell invasion and metastasis
(20).
The present study aimed to investigate the
anti-invasive and anti-metastatic activities of TGZ and
hypothesized that TGZ may act on various stages of the metastatic
procedure to prevent cancer cells from metastasizing. The present
study therefore determined the effect of TGZ on the reduction of
cell invasive activity and also examined the effect of TGZ on the
expression levels of E-cadherin and GPx3 in PC-3 human prostate
cancer cells.
Materials and methods
Prostate cancer cell line
The human prostate cancer PC-3 cell line was
acquired from the American Type Culture Collection (Manassas, VA,
USA). These cells were cultured in RPMI medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and
1% penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.)
at 37°C in 95% air and 5% CO2. TGZ (Sigma-Aldrich; Merck
KGAa, Darmstadt, Germany) and GW9662 (Cayman Chemical Company, Ann
Arbor, MI, USA) were dissolved in 100% ethanol (Merck KGaA) to
obtain a concentration of 16 mM. The final ethanol concentration in
the solution was 0.25%. Ethanol alone at the same final
concentration of 0.25% was used as the control.
Cellular viability assay
The viability of PC-3 cells was analyzed using an
MTT (Sigma-Aldrich; Merck KGaA) assay based on the ability of live
cells to convert tetrazolium salt into purple formazan. Briefly,
PC-3 cells were seeded into 96-well cell culture plates at a
density of 8×103 cells/well in 200 µl media. Following
24 h incubation at 37°C, the medium was replaced with 1% FBS-RPMI
medium at 37°C for 24 h. Cells were treated with TGZ at
concentrations of 1, 10 and 40 µM or vehicle (ethanol) control and
cultured at 37°C for 48 h. The medium was then replaced with 100 µl
of MTT (diluted to 1 mg/ml in FBS-free medium, from a stock
solution of 10 mg/ml) and incubated at 37°C for 3 h. The
supernatant was removed and 100 µl of DMSO was supplemented to each
well to dissolve the formazan crystals. Plates were agitated at
room temperature for 5 min. The absorbance was read at 540 nm on an
Epoch BioTek microplate reader (BioTek Instruments, Inc., Winooski,
VT, USA). All treatments were performed in triplicate.
Cellular migration assay
PC-3 cell motility was analyzed using an in
vitro wound-healing assay. PC-3 cells were seeded into six-well
cell culture plates at a density of 8×104 cells/well and
grown until confluent at 37°C. Monolayers of confluent PC-3 cells
were then wounded using a pipette tip. Cell repair was monitored
using an inverted microscope (Olympus IX70, Tokyo, Japan) following
24 h exposure at 37°C to TGZ at concentrations of 1, 10 and 40 µM
or the vehicle (ethanol) control. The widths of the wounded areas
were measured by ImageJ software (version 1.48; National Institutes
of Health, Bethesda, MD, USA), and the relative wound closure
ratios of the distance to that at 0 h were calculated at 6, 9, 12
and 24 h. All treatments were performed in triplicate.
Cellular invasion assay
PC-3 cellular invasion was determined using a
Transwell insert. Briefly, 8 µm Transwell inserts (SPL Life
Sciences, Pocheon, Korea) were coated with Matrigel (Gibco; Thermo
Fisher Scientific, Inc.) and incubated at 37°C for 2 h to become
gelatinous. Cells were grown to subconfluence, detached by
trypsinization, washed twice with PBS and resuspended in serum free
RPMI medium. Subsequently, the cells added to the Transwell insert
(2×104 cells/insert) at a final concentration of
1×105 cells/ml together with TGZ at concentrations of 1,
10 and 40 µM or the vehicle (ethanol) control. Cell-free RPMI
medium was added to the lower chamber. Assays were incubated at
37°C for 30 h until cells had migrated to the bottom of the
membrane. Non-invasive cells in the upper surface of the membrane
were removed with a cotton swab. The remaining cells on the
membrane were fixed in methanol for 10 min at room temperature,
stained with hematoxylin for 10 min at room temperature and washed
with PBS. Invaded cells were counted in four fields per insert with
a light microscope at ×100 magnification. Cells were assessed for
their relative invasion ability as percentages of the vehicle
(ethanol) controls. All treatments were performed in
triplicate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from PC-3 cells using a
Hybrid-R RNA extraction kit (GeneAll Biotechnology, Seoul, South
Korea), and then cDNA was synthesized using a M-MLV cDNA Synthesis
kit (Enzynomics, Daejeon, South Korea) according to the
manufacturer's protocol. RT-qPCR was performed using TOPreal™ qPCR
2X PreMIX (SYBR Green with high ROX; Enzynomics) on a CFX Connect
Real-Time PCR Detection system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The reaction conditions were as follows:
Pre-incubation at 94°C for 10 min, followed by 40 cycles (94°C for
10 sec, 58°C for 20 sec, 72°C for 30 sec) ending with a melting
curve program with temperature rising from 60 to 95°C. The
specificity of the reactions was verified by melting curve
analysis. The primers used were: Human GPx3 sense,
5′-ACATGCCTACAGGTATGCGT-3 and antisense,
5′-GAGCAGAACAATTGGACCTA-3′; human E-cadherin sense,
5′-TTGCTACTGGAACAGGGACACT-3′ and antisense
5′-GGAGATGTATTGGGAGGAAGGTC-3′; and human β-actin sense,
5′-CATGTACGTTGCTATCCAGGC-3′ and antisense,
5′-CTCCTTAATGTCACGCACGAT-3′. The ratio of target gene fold-change
was normalized to human β-actin expression level using the
comparative 2−ΔΔCq method (21). All treatments were performed in
triplicate.
Western blot analysis
Cell lysates were subjected to SDS-PAGE (on 10%
acrylamide gel), transferred to nitrocellulose membranes and probed
with mouse monoclonal anti-E-cadherin primary antibody (sc-8426,
Santa Cruz Biotechnology, Inc., Dallas, TX, USA; 1:1,000 dilution),
mouse monoclonal anti-GPx3 primary antibody (ab27325, Abcam,
Cambridge, MA, USA; 1:500 dilution) and goat polyclonal anti-actin
primary antibody (sc-1616, Santa Cruz Biotechnology, Inc.; 1:1,000
dilution). Blots were incubated with horseradish
peroxidase-conjugated anti-mouse Ig secondary antibodies (61–0120,
Zymed; Thermo Fisher Scientific, Inc.; 1:2,000 dilution) and
anti-goat Ig (81–1620, Zymed; 1:2,000 dilution). Then, the antibody
reactive bands were developed using a chemiluminescent substrate
(DoGEN, Seoul, South Korea). Blots were quantified with ImageJ
software (version 1.48; National Institutes of Health), and the
relative ratio was calculated.
Statistical analysis
All data are presented as the mean ± standard error.
Statistical significance was further analyzed by Student's t-test
using Excel 2010 (Microsoft Corporation, Redmond, WA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
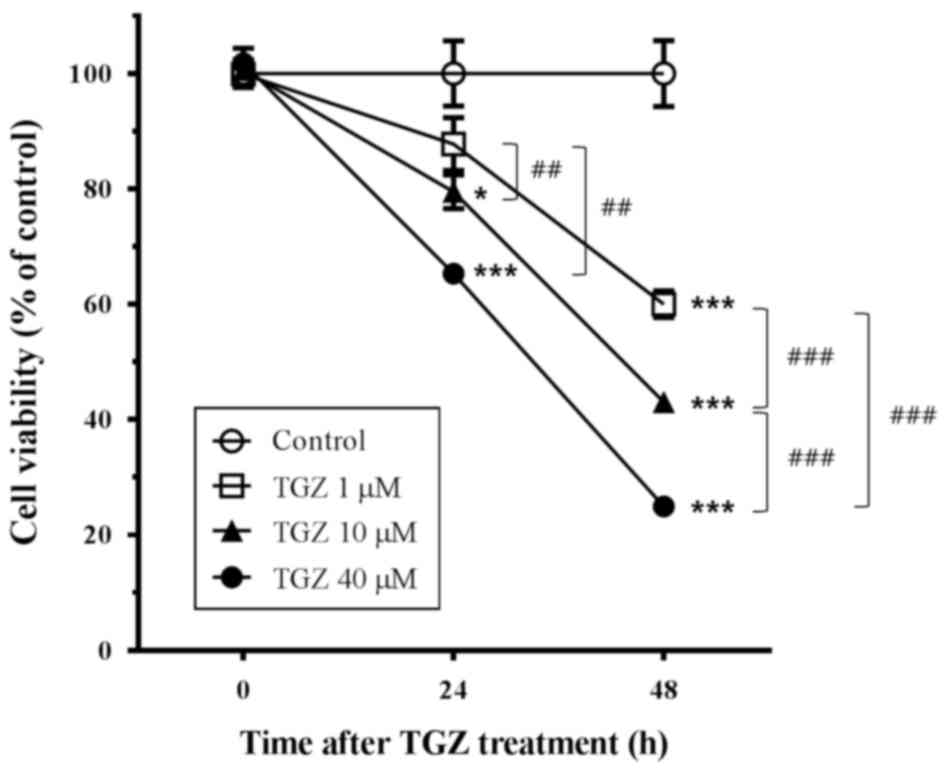
Treatment with TGZ decreased cellular
viability of PC-3 cells
To examine the effect of TGZ on the cellular
viability of PC-3 cells, the present study treated these cells with
1, 10 or 40 µM TGZ for 0, 24 or 48 h, and determined the rate of
cell survival using an MTT assay. Results are presented in Fig. 1. TGZ treatment decreased cellular
viability in a dose-dependent manner. Compared with the control
cells, cells treated with 10 and 40 µM TGZ for 24 h demonstrated
decreases in cell growth rates by 20.49 and 34.69%,
respectively.
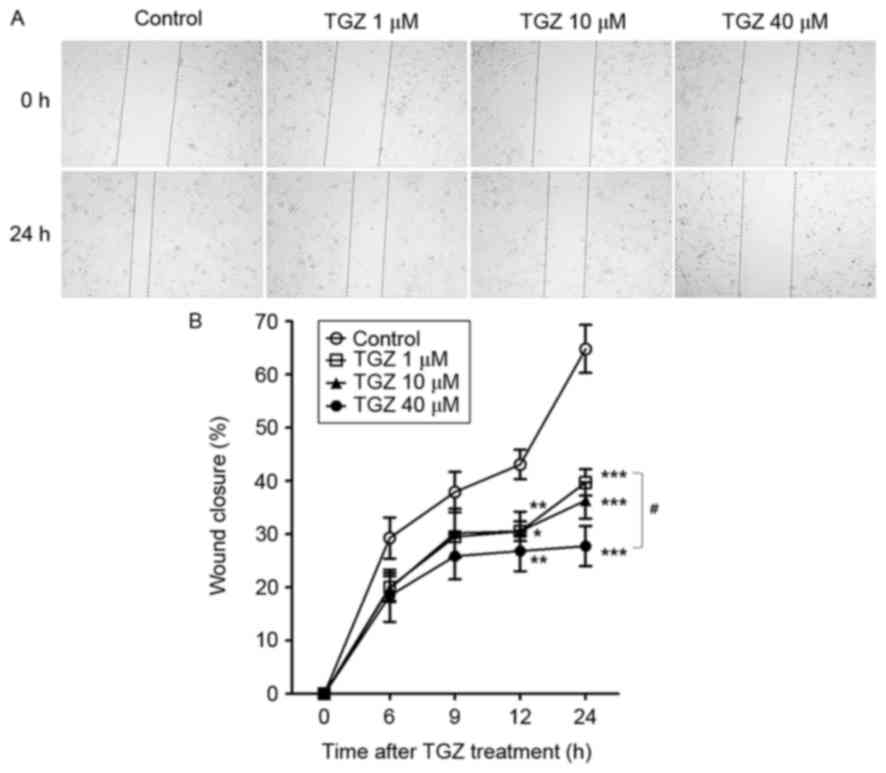
TGZ inhibits cell migration and
invasion of PC-3 cells
The present study evaluated cell migration and
invasion of PC-3 cells treated with TGZ by in vitro wound
healing and Matrigel assays. The results are presented in Figs. 2 and 3,
respectively. Following incubation with various concentrations of
TGZ for 24 h, the migration of PC-3 cells to the denuded area was
inhibited in a dose-dependent manner (Fig. 2), indicating that TGZ significantly
suppressed the motility of PC-3 cells.
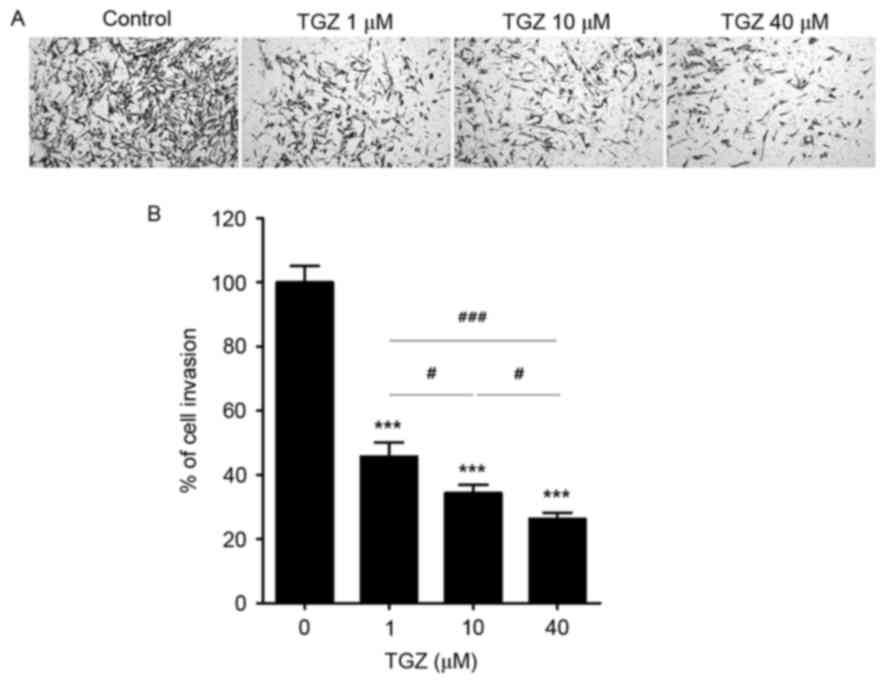
Results of the Matrigel assay revealed that TGZ
inhibited the invasion of PC-3 cells through the Matrigel-coated
filter pores in a dose-dependent manner (Fig. 3). Treatment with 1, 10 and 40 µM TGZ
inhibited 54.27, 65.63, and 73.70% of cell invasion, respectively.
These results demonstrated that TGZ significantly inhibited the
invasion of PC-3 cells. Taken together, these results revealed that
TGZ suppressed cellular migration and invasion of PC-3 cells.
TGZ increases the mRNA levels of
E-cadherin and GPx3 in PC-3 human prostate cancer cells
To investigate whether TGZ could enhance the
transcriptional levels of E-cadherin and GPx3 genes in PC-3 cells,
RT-qPCR was performed. The results are presented in Fig. 4. TGZ treatments of 1 and 10 µM
significantly increased the mRNA expression levels of E-cadherin in
a dose-dependent manner (Fig. 4A). A
TGZ treatment of 10 µM also significantly increased the mRNA
expression levels of GPx3 (Fig.
4B).
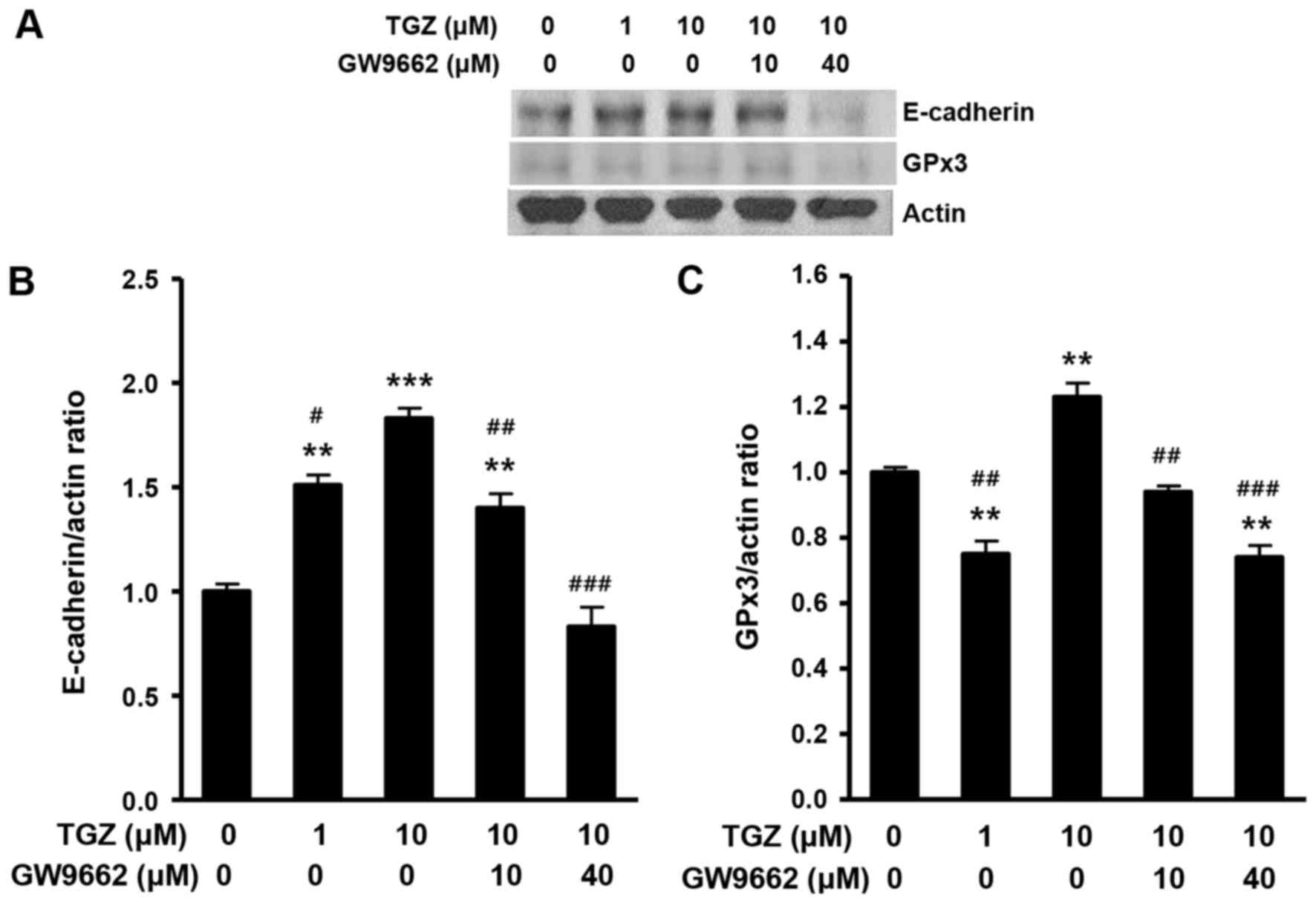
Although TGZ was able to affect the transcriptional
expression levels of E-cadherin and GPx3, the mechanisms underlying
these effects remain unclear. To prove whether these results were
dependent on the activation of PPARγ, GW9662 (a PPARγ antagonist)
was used to inhibit the function of PPARγ in PC-3 cells.
TGZ-induced upregulation of E-cadherin and GPx3 mRNAs in PC-3 cells
were decreased following treatment with GW9662 (Fig. 4). These results demonstrated that TGZ
increased the transcriptional expression levels of E-cadherin and
GPx3 in PC-3 cells in a PPARγ-dependent manner.
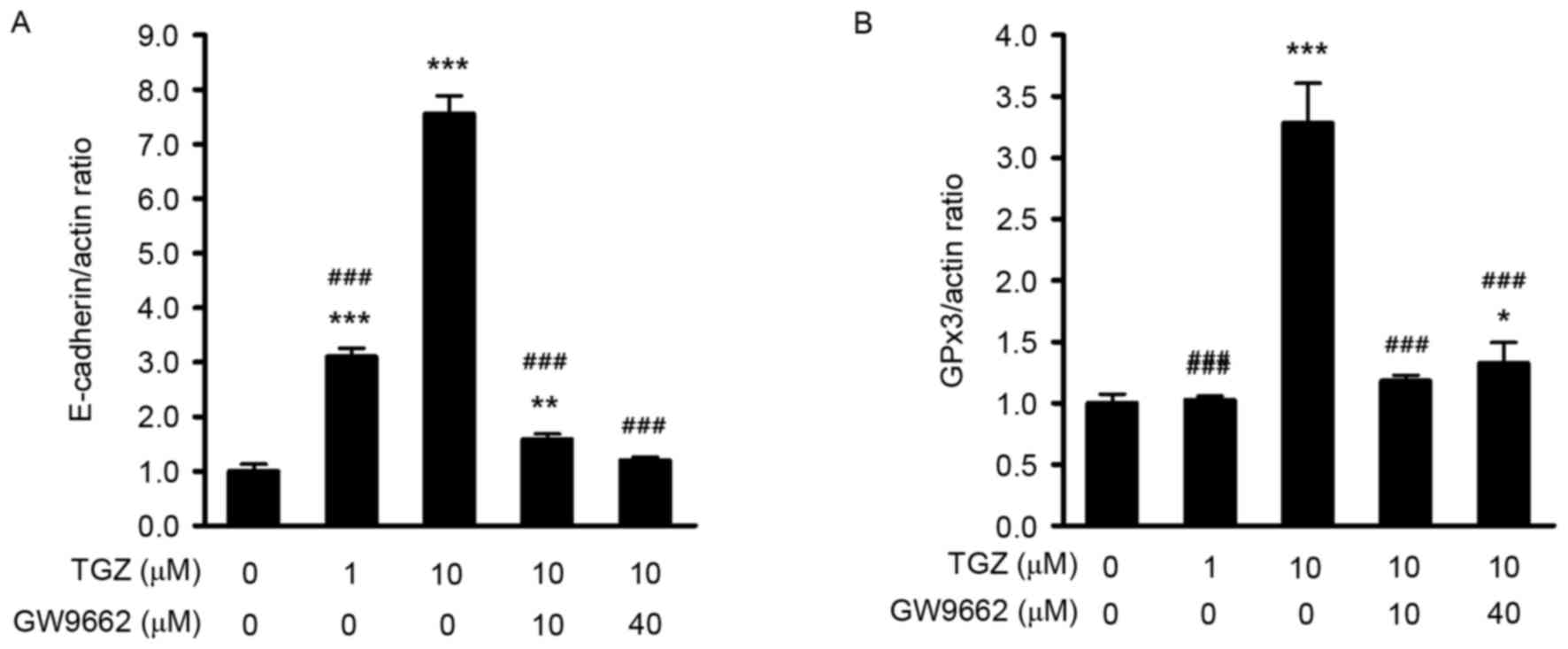
TGZ increases the protein levels of
E-cadherin and GPx3 in PC-3 human prostate cancer cells
The protein expression levels of E-cadherin and GPx3
in PC-3 cells were determined by western blot analysis following 48
h treatment with 0, 1 or 10 µM TGZ. Results are presented in
Fig. 5. TGZ at 1 and 10 µM
significantly increased the protein expression levels of E-cadherin
in a dose-dependent manner (Fig. 5A and
B). TGZ at 10 µM also significantly increased the protein
expression levels of GPx3 (Fig. 5A and
C). TGZ-induced upregulation of E-cadherin and GPx3 protein
expression levels in PC-3 cells were decreased following the
addition of GW9662 (Fig. 5). Thus,
TGZ upregulated E-cadherin and GPx3 protein expression levels in a
PPARγ-dependent manner, which was consistent with its upregulating
effect on E-cadherin and GPx3 mRNA expression levels.
Discussion
TZDs are a novel class of antidiabetics and are
specific ligands for PPARγ (22).
PPARγ is expressed at various levels depending on the type of
tissue and carcinoma (23). PPARγ has
been investigated as a therapeutic target for cancer treatment.
Certain previous studies have reported that PPARγ is able to induce
anti-proliferative, anti-angiogenic and pro-differentiation
signaling pathways in specific tissue types, thus serving a role in
the pathogenesis and progression of various types of cancer,
including prostate cancer (24,25). Using
TZDs as PPARγ ligands, previous studies have investigated the
effect of PPARγ on the metastatic potential and investigated its
underlying mechanisms (26–34). TZDs have been demonstrated to be able
to suppress cellular migration, invasion and metastasis of cancer
in the colon, liver, breast, lung, bladder and prostate gland
(26–34). For example, in colon cancer, TZD
inhibited the development and metastasis of HT-29 human colon
cancer cells via its differentiation-promoting effects both in
vivo and in vitro by involving the modulation of the
E-cadherin/β-catenin system (26). In
hepatocellular carcinoma, the suppression of cellular invasion and
migration mediated by PPARγ has been revealed to be mediated via
downregulation of matrix metalloproteinases, by and increased
expression levels of TIMP metallopeptidase inhibitor 3 and
E-cadherin (28,29).
However, the mechanisms underlying the inhibitory
action of TZDs on cell invasion in prostate cancer remain unclear.
To better comprehend the molecular mechanisms involved in
TGZ-induced inhibition of cell invasion, the present study analyzed
the effect of TGZ on the expression levels of E-cadherin and GPx3.
E-cadherin and GPx3 may be involved in cancer cell invasion. The
present study demonstrated that TGZ reduced cell migration and
invasion of PC-3 cells. Consistently, TGZ positively regulated the
mRNA and protein expression levels of E-cadherin and GPx3 in a
PPARγ-dependent manner. The results from the present study may
provide preliminary evidence for further studies investigating the
mechanisms underlying the suppression of metastasis by TZDs.
A number of human solid tumors are carcinomas
originating from various epithelial cell types (18,19). Tumor
cells change from a highly differentiated epithelial morphology to
a migratory and invasive phenotype during the conversion from a
benign tumor to a malignant invasive tumor (18). This process of EMT can induce the loss
of cell-cell contacts and an increase in cell motility (18,35). It
also induces the dissemination of single tumor cells from primary
epithelial tumors to distant organs (19). These alterations are necessary for
invasion. Throughout the EMT process, numerous genes involved in
cellular adhesion, migration and invasion and mesenchymal
differentiation are transcriptionally regulated (19). Amongst these genes, loss of adhesive
function of E-cadherin in epithelial cells has been suggested to be
a hallmark of EMT and metastatic carcinoma (17,19,20,35).
E-cadherin formation is preserved in the majority of
differentiated tumors, including carcinomas of the skin, head and
neck, breast, lung, liver, colon and prostate (17–20).
However, E-cadherin expression levels demonstrated an inverse
correlation with cancer grade or patient survival (17). In a number of human and mouse types of
cancer, carcinoma progression and poor prognosis have been revealed
to be associated with partial loss of E-cadherin expression
(19). Increasing E-cadherin function
in cancer cells and mouse cancer models suppresses the ability to
invade and metastasize (19).
Conversely, disrupted expression of E-cadherin in noninvasive tumor
cells induces invasive and metastatic activities (18,19). In
the present study, an increase in E-cadherin expression level
induced increased cell-cell interactions and decreased motility of
PC-3 cells following TGZ treatment. Therefore, changing the
functional protein expression levels of E-cadherin may be an
alternative approach for cancer therapy.
GPx3 belongs to the family of glutathione
peroxidases (36). It is well known
that glutathione peroxidases are among the most important reactive
oxygen species scavengers that protect cells from oxidative damage
(36). Downregulation of GPx3 by
hypermethylation has been revealed in numerous types of cancer
(37–45), including prostate cancer (11,46,47). In
prostate cancer, a negative correlation between GPx3 expression
levels and poor clinical outcomes has been demonstrated (47,48).
Forced expression of GPx3 in prostate cancer cell lines is able to
suppress colony formation and cellular proliferation (11,47,49).
Xenografts with higher GPx3 expression levels have resulted in
reduced tumor volume, metastasis and animal mortality (47). Our previous study revealed that GPx3
is downregulated in prostates of TRAMP mice and that disruption of
GPx3 expression in TRAMP mice increased prostate cancer development
and metastasis (50). It has been
demonstrated that silencing GPx3 expression promoted cancer
metastasis in human thyroid and gastric cancer cells (37,38). In
the present study, it was notable that GPx3 expression level was
upregulated following treatment with TGZ in human prostate cancer
cells. This induction was closely associated with the anti-invasion
effect of TGZ. All these results suggested that GPx3 acted as a
negative regulator of prostate tumor development and metastasis,
producing further evidence for its role as a tumor suppressor.
In conclusion, the present study revealed that TGZ
effectively abrogated the migration and invasion of PC-3 cells
in vitro by increasing the expression levels of E-cadherin
and GPx3. Upregulated expression levels of E-cadherin and GPx3 is a
possible mechanism underlying the anti-migration and anti-invasion
effects of TGZ. Altered expression levels induced by TGZ may
account for the mechanisms underlying the invasion inhibition
ability of TZDs and its mode of action.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Basic Science
Research Program of the National Research Foundation of Korea
funded by the Ministry of Education, Republic of Korea (grant no.,
NRF-2012R1A1A2009579) and the Research Institute for Veterinary
Science and the BK21 PLUS Program for Creative Veterinary Science
Research, College of Veterinary Medicine, Seoul National
University, Korea.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SC performed all experiments, analyzed the data and
wrote the manuscript. JL contributed to the cell culture and
RT-qPCR. HO, UK, and BR participated in the MTT assay, wound
healing assay and data collection. JP conceived the idea,
interpreted the data and helped to draft the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chi KN, Bjartell A, Dearnaley D, Saad F,
Schröder FH, Sternberg C, Tombal B and Visakorpi T:
Castration-resistant prostate cancer: From new pathophysiology to
new treatment targets. Eur Urol. 56:594–605. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sun Y, Wang BE, Leong KG, Yue P, Li L,
Jhunjhunwala S, Chen D, Seo K, Modrusan Z, Gao WQ, et al: Androgen
deprivation causes epithelial-mesenchymal transition in the
prostate: Implications for androgen-deprivation therapy. Cancer
Res. 72:527–536. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grommes C, Landreth GE and Heneka MT:
Antineoplastic effects of peroxisome proliferator-activated
receptor gamma agonists. Lancet Oncol. 5:419–429. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weng JR, Chen CY, Pinzone JJ, Ringel MD
and Chen CS: Beyond peroxisome proliferator-activated receptor
gamma signaling: The multi-facets of the antitumor effect of
thiazolidinediones. Endocr Relat Cancer. 13:401–413. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wei S, Yang J, Lee SL, Kulp SK and Chen
CS: PPARgamma-independent antitumor effects of thiazolidinediones.
Cancer Lett. 276:119–124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu HN, Lee YR, Noh EM, Lee KS, Kim JS,
Song EK, Han MK, Lee YC, Kwon KB, Lee SJ, et al: Induction of G1
phase arrest and apoptosis in MDA-MB-231 breast cancer cells by
troglitazone, a synthetic peroxisome proliferator-activated
receptor gamma (PPARgamma) ligand. Cell Biol Int. 32:906–912. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Colin-Cassin C, Yao X, Cerella C, Chbicheb
S, Kuntz S, Mazerbourg S, Boisbrun M, Chapleur Y, Diederich M,
Flament S and Grillier-Vuissoz I: PPARγ-inactive Δ2-troglitazone
independently triggers ER stress and apoptosis in breast cancer
cells. Mol Carcinog. 54:393–404. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ye J, Yin L, Xie P, Wu J, Huang J, Zhou G,
Xu H, Lu E and He X: Antiproliferative effects and molecular
mechanisms of troglitazone in human cervical cancer in vitro. Onco
Targets Ther. 8:1211–1218. 2015.PubMed/NCBI
|
|
10
|
Li MY, Deng H, Zhao JM, Dai D and Tan XY:
PPARgamma pathway activation results in apoptosis and COX-2
inhibition in HepG2 cells. World J Gastroenterol. 9:1220–1226.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chang SN, Han J, Abdelkader TS, Kim TH,
Lee JM, Song J, Kim KS and Park JH and Park JH: High animal fat
intake enhances prostate cancer progression and reduces glutathione
peroxidase 3 expression in early stages of TRAMP mice. Prostate.
74:1266–1277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shiau CW, Yang CC, Kulp SK, Chen KF and
Chen CS, Huang JW and Chen CS: Thiazolidenediones mediate apoptosis
in prostate cancer cells in part through inhibition of Bcl-xL/Bcl-2
functions independently of PPARgamma. Cancer Res. 65:1561–1569.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Santha S, Viswakarma N, Das S, Rana A and
Rana B: Tumor necrosis factor-related apoptosis-inducing ligand
(TRAIL)-troglitazone-induced apoptosis in prostate cancer cells
involve AMP-activated protein kinase. J Biol Chem. 290:21865–21875.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mueller E, Smith M, Sarraf P, Kroll T,
Aiyer A, Kaufman DS, Oh W, Demetri G, Figg WD, Zhou XP, et al:
Effects of ligand activation of peroxisome proliferator-activated
receptor gamma in human prostate cancer. Proc Natl Acad Sci USA.
97:10990–10995. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Burstein HJ, Demetri GD, Mueller E, Sarraf
P, Spiegelman BM and Winer EP: Use of the peroxisome
proliferator-activated receptor (PPAR) gamma ligand troglitazone as
treatment for refractory breast cancer: A phase II study. Breast
Cancer Res Treat. 79:391–397. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kulke MH, Demetri GD, Sharpless NE, Ryan
DP, Shivdasani R, Clark JS, Spiegelman BM, Kim H, Mayer RJ and
Fuchs CS: A phase II study of troglitazone, an activator of the
PPARgamma receptor, in patients with chemotherapy-resistant
metastatic colorectal cancer. Cancer J. 8:395–399. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Christofori G: New signals from the
invasive front. Nature. 441:444–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Frixen UH, Behrens J, Sachs M, Eberle G,
Voss B, Warda A, Löchner D and Birchmeier W: E-cadherin-mediated
cell-cell adhesion prevents invasiveness of human carcinoma cells.
J Cell Biol. 113:173–185. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Day C: Thiazolidinediones: A new class of
antidiabetic drugs. Diabet Med. 16:179–192. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ikezoe T, Miller CW, Kawano S, Kawano S,
Heaney A, Williamson EA, Hisatake J, Green E, Hofmann W, Taguchi H
and Koeffler HP: Mutational analysis of the peroxisome
proliferator-activated receptor gamma gene in human malignancies.
Cancer Res. 61:5307–5310. 2001.PubMed/NCBI
|
|
24
|
Matsuyama M and Yoshimura R: Peroxisome
proliferator-activated receptor-gamma is a potent target for
prevention and treatment in human prostate and testicular cancer.
PPAR Res. 2008:2498492008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sikka S, Chen L, Sethi G and Kumar AP:
Targeting PPARgamma signaling cascade for the prevention and
treatment of prostate cancer. PPAR Res. 2012:9680402012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoshizumi T, Ohta T, Ninomiya I, Terada I,
Fushida S, Fujimura T, Nishimura G, Shimizu K, Yi S and Miwa K:
Thiazolidinedione, a peroxisome proliferator-activated
receptor-gamma ligand, inhibits growth and metastasis of HT-29
human colon cancer cells through differentiation-promoting effects.
Int J Oncol. 25:631–639. 2004.PubMed/NCBI
|
|
27
|
Takano S, Kubota T, Nishibori H, Hasegawa
H, Ishii Y, Nitori N, Ochiai H, Okabayashi K, Kitagawa Y, Watanabe
M and Kitajima M: Pioglitazone, a ligand for peroxisome
proliferator-activated receptor-gamma acts as an inhibitor of colon
cancer liver metastasis. Anticancer Res. 28:3593–3599.
2008.PubMed/NCBI
|
|
28
|
Shen B, Chu ES, Zhao G, Man K, Wu CW,
Cheng JT, Li G, Nie Y, Lo CM, Teoh N, et al: PPARgamma inhibits
hepatocellular carcinoma metastases in vitro and in mice. Br J
Cancer. 106:1486–1494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee HJ, Su Y, Yin PH, Lee HC and Chi CW:
PPAR(gamma)/PGC-1(alpha) pathway in E-cadherin expression and
motility of HepG2 cells. Anticancer Res. 29:5057–5063.
2009.PubMed/NCBI
|
|
30
|
Magenta G, Borenstein X, Rolando R and
Jasnis MA: Rosiglitazone inhibits metastasis development of a
murine mammary tumor cell line LMM3. BMC Cancer. 8:472008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu H, Zang C, Fenner MH, Possinger K and
Elstner E: PPARgamma ligands and ATRA inhibit the invasion of human
breast cancer cells in vitro. Breast Cancer Res Treat. 79:63–74.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoo JY, Yang SH, Lee JE, Cho DG, Kim HK,
Kim SH, Kim IS, Hong JT, Sung JH, Son BC and Lee SW: E-cadherin as
a predictive marker of brain metastasis in non-small-cell lung
cancer, and its regulation by pioglitazone in a preclinical model.
J Neurooncol. 109:219–227. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang DR, Lin SJ, Ding XF, Miyamoto H,
Messing E, Li LQ, Wang N and Chang C: Higher expression of
peroxisome proliferator-activated receptor γ or its activation by
agonist thiazolidinedione-rosiglitazone promotes bladder cancer
cell migration and invasion. Urology. 81:1109.e1–e6. 2013.
View Article : Google Scholar
|
|
34
|
Qin L, Gong C, Chen AM, Guo FJ, Xu F, Ren
Y and Liao H: Peroxisome proliferatoractivated receptor γ agonist
rosiglitazone inhibits migration and invasion of prostate cancer
cells through inhibition of the CXCR4/CXCL12 axis. Mol Med Rep.
10:695–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: New insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brigelius-Flohé R and Maiorino M:
Glutathione peroxidases. Biochim Biophys Acta. 1830:3289–3303.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao H, Li J, Li X, Han C, Zhang Y, Zheng
L and Guo M: Silencing GPX3 expression promotes tumor metastasis in
human thyroid cancer. Curr Protein Pept Sci. 16:316–321. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Peng DF, Hu TL, Schneider BG, Chen Z, Xu
ZK and El-Rifai W: Silencing of glutathione peroxidase 3 through
DNA hypermethylation is associated with lymph node metastasis in
gastric carcinomas. PLoS One. 7:e462142012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Murawaki Y, Tsuchiya H, Kanbe T, Harada K,
Yashima K, Nozaka K, Tanida O, Kohno M, Mukoyama T, Nishimuki E, et
al: Aberrant expression of selenoproteins in the progression of
colorectal cancer. Cancer Lett. 259:218–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Agnani D, Camacho-Vanegas O, Camacho C,
Lele S, Odunsi K, Cohen S, Dottino P and Martignetti JA: Decreased
levels of serum glutathione peroxidase 3 are associated with
papillary serous ovarian cancer and disease progression. J Ovarian
Res. 4:182011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brigelius-Flohé R and Kipp A: Glutathione
peroxidases in different stages of carcinogenesis. Biochim Biophys
Acta. 1790:1555–1568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen B, Rao X, House MG, Nephew KP, Cullen
KJ and Guo Z: GPx3 promoter hypermethylation is a frequent event in
human cancer and is associated with tumorigenesis and chemotherapy
response. Cancer Lett. 309:37–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
He Y, Wang Y, Li P, Zhu S, Wang J and
Zhang S: Identification of GPX3 epigenetically silenced by CpG
methylation in human esophageal squamous cell carcinoma. Dig Dis
Sci. 56:681–688. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mohamed MM, Sabet S, Peng DF, Nouh MA,
El-Shinawi M and El-Rifai W: Promoter hypermethylation and
suppression of glutathione peroxidase 3 are associated with
inflammatory breast carcinogenesis. Oxid Med Cell Longev.
2014:7871952014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee OJ, Schneider-Stock R, McChesney PA,
Kuester D, Roessner A, Vieth M, Moskaluk CA and El-Rifai W:
Hypermethylation and loss of expression of glutathione peroxidase-3
in Barrett's tumorigenesis. Neoplasia. 7:854–861. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lodygin D, Epanchintsev A, Menssen A,
Diebold J and Hermeking H: Functional epigenomics identifies genes
frequently silenced in prostate cancer. Cancer Res. 65:4218–4227.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu YP, Yu G, Tseng G, Cieply K, Nelson J,
Defrances M, Zarnegar R, Michalopoulos G and Luo JH: Glutathione
peroxidase 3, deleted or methylated in prostate cancer, suppresses
prostate cancer growth and metastasis. Cancer Res. 67:8043–8050.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sekine Y, Osei-Hwedieh D, Matsuda K,
Raghavachari N, Liu D, Furuya Y, Koike H, Suzuki K and Remaley AT:
High fat diet reduces the expression of glutathione peroxidase 3 in
mouse prostate. Prostate. 71:1499–1509. 2011.PubMed/NCBI
|
|
49
|
Wang H, Luo K, Tan LZ, Ren BG, Gu LQ,
Michalopoulos G, Luo JH and Yu YP: p53-induced gene 3 mediates cell
death induced by glutathione peroxidase 3. J Biol Chem.
287:16890–16902. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chang SN, Lee JM, Oh H and Park JH:
Glutathione peroxidase 3 inhibits prostate tumorigenesis in TRAMP
mice. Prostate. 76:1387–1398. 2016. View Article : Google Scholar : PubMed/NCBI
|