Introduction
Ovarian cancer is a major malignant tumor type
affecting the female reproductive system, which has the highest
mortality rate of all gynecological tumors (1). Therapeutic drug resistance is a major
factor of the chemotherapy failure observed in the treatment of
ovarian cancer (2). Cisplatin (DDP)
is preferentially used for chemotherapy in ovarian cancer in
clinical practice; however, its efficacy is often restricted due to
its dose-limiting toxicities, including bone marrow toxicity,
nephrotoxicity and the development of drug resistance (3–5).
Identifying a method to limit DDP toxicity while maintaining its
efficacy is significantly important for successful chemotherapy in
ovarian cancer (6). Numerous studies
have evaluated that cancer cells, in comparison with normal cells,
are under increasing levels of oxidative stress associated with an
increased overall generation level of reactive oxygen species (ROS)
(7,8).
The moderately increased expression levels of ROS in cancer cells
may stimulate cellular proliferation and promote mutations and
genetic instability (9,10); however, excessive production of ROS
may inflict damage to various cellular components, including DNA,
protein and lipid membranes (11,12). This
increased intrinsic ROS stress in cancer cells provides a unique
opportunity for killing the malignant cells, due to their
vulnerability to additional ROS attack (13).
As a small molecular of nitroxide radicals, Tempol
(TPL) has been utilized as a biophysical tool for electron
paramagnetic resonance spectroscopy in numerous studies (14–16). TPL
has an unpaired electron and undergoes rapid reversible transfer
between 3 forms: Nitroxide, hydroxylamine and the oxoamonium cation
(17). Therefore, TPL is a potential
redox agent that may function as a reductive or oxidative agent
depending on the concentration in the cell (18). The clinical application of TPL at
<1 mM is usually as an antioxidative agent in the treatment of
inflammation (19,20), such as periodontitis in a rodent model
(21). And TPL also has a clinical
application in neurodegenerative diseases including including
Alzheimer's disease, Parkinson's disease and Huntington's disease
(22,23), or hypertension (24). In contrast to studies regarding the
antioxidative effects of TPL, another study has indicated that TPL,
at concentrations of >1 mM, may serve as a pro-oxidant by
producing ROS and oxidizing reduced transition metals (25). TPL is favorable for inhibiting the
growth of neoplastic cells by increasing cellular ROS production
(25,26). Based on these data, the present study
hypothesized that the pro-oxidative activity of TPL increased the
antitumor effects of DDP by increasing cellular ROS production and
inducing cell apoptosis. The present study investigated the
potentiating effect of TPL with an antitumor drug, DDP, on cellular
proliferation and apoptosis in ovarian cancer cells.
Materials and methods
Chemicals
Dulbecco's modified Eagle's medium (DMEM), trypsin
and fetal bovine serum (FBS) were purchased from Gibco (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). PBS was obtained from
Boster Biological Technology (Pleasanton, CA, USA). TPL was
purchased from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan).
Cisplatin and MTT were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). The Annexin V-fluorescein isothiocyanate
(FITC) Apoptosis Detection kit was obtained from BD Biosciences
(Franklin Lakes, NJ, USA). The primary antibodies used for western
blot analysis were purchased from multiple companies: Anti-Bcl-2
and anti-Bax were obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA) and anti-GAPDH was purchased from Sigma-Aldrich,
Merck KGaA. Dichlorofluorescin-diacetate (DCFH-DA) was obtained
from Sigma-Aldrich, Merck KGaA.
Cell culture
OVCAR3 and SKOV3 human ovarian cancer cell lines
were purchased from Southern Medical University Cancer Institute
(Guangzhou, China). Cells were cultured in DMEM supplemented with
10% FBS at 37°C in a humidified incubator at 5% CO2.
Cells were routinely subcultured using 0.05% trypsin-EDTA
solution.
Cell viability assay
Briefly, cells were seeded into 96-well plates at
the density of 5×103 cells/well. Following incubation at
37°C in humidified incubator for 24 h, the DMEM was removed and
replaced with a fresh medium containing increasing concentrations
of TPL (2, 4, 6, 8 or 10 mM) and cultured at 37°C for 48 h. In
order to estimate the effect of TPL on cell proliferation, growth
curve analyses of OVCAR3 and SKOV3 cells were performed. Cells were
seeded into 96-well plates at the density of 1×103
cells/well and cultured at 37°C with TPL (1.5 Mm in OVCAR3 cells
and 1 mM in SKOV3 cells) for 5 days. To investigate the effect of
combination treatment of TPL and DDP on OVCAR3 cells proliferation,
cells were cultured with 6 concentrations of DDP (1, 2, 4, 8, 16 or
32 µM) at 37°C for 48 h, and then 3 µM DDP combined with 1.5 and 2
mM TPL, respectively at 37°C for 48 h. Following treatment, the
medium was discarded, cells were washed with PBS once and 100 µl
MTT solution (0.5 mg/ml), which was diluted with 10% FBS and phenol
red-free DMEM (Thermo Fisher Scientific, Inc.), was added to each
well and incubated for 4 h at 37°C. Subsequently, the solution was
discarded, 50 µl dimethyl sulfoxide was added to each well and the
absorbance was evaluated at 490 nm. All experiments were performed
in triplicate.
Inverted phase contrast microscopy
observation
OVCAR3 cells were seeded in 6-well plates
(2×106 cells/well) and treated with 1.5 mM TPL, 3 µM DDP
or a combination of TPL and DDP (1.5 mM TPL and 3 µM DDP) at 37°C
for 48 h. Following a 48 h exposure to drug or control treatments,
the cells were observed at ×10 magnification in 22 fields of view
(18 mm) using an inverted phase contrast microscope. Images were
captured at magnification, ×200.
Flow cytometry (FCM) analysis
Induction of apoptosis was determined using an
Annexin V-FITC Apoptosis Detection kit, according to the
manufacturer's protocol. OVCAR3 cells were seeded in culture dishes
at the density of 2×105 cells/well and incubated at 37°C
for 48 h with media containing 1.5 Mm TPL or 3 µM DDP or a
combination of TPL and DDP (1.5 mM TPL and 3 µM DDP). Annexin
V-FITC-positive staining is a hallmark of early apoptosis, whereas
late apoptosis and necrosis are indicated by additional positive
nuclear staining with propidium iodide (PI). The cells were
immediately detected with the probes by FCM analysis using FACS
Aria II (BD Biosciences) and analyzed by FlowJo Software (FlowJo
LLC, Ashland, OR, USA). Each sample was analyzed three times in
three independent experiments.
Western blot analysis
OVCAR3 cells were treated with TPL (1 mM), DDP (3
µM) or combination of 1 mM TPL and 3 µM DDP at 37°C for 48 h.
Following incubation, cells were digested with 0.05% trypsin-EDTA
solution at 37°C for 3 min, collected into a centrifuge tube and
washed twice with cold PBS. Protein lysis buffer (50 µl/dish) made
up of 1X protease inhibitors (Hangzhou Fude Biological Technology
Co., Ltd., Hangzhou, China) and 1 mM phenylmethanesulfonyl fluoride
(Fude Biological Technology Co., Ltd.) was added to centrifuge
tubes and cells were lyzed on ice for 10 min. The concentration of
the prepared protein was detected according to the manufacturer's
protocol of the Bicinchoninic Acid Protein Assay kit (Thermo Fisher
Scientific, Inc.). The whole cellular proteins (50 µg) were then
mixed with 5X SDS-PAGE sample loading buffer (Fude Biological
Technology Co., Ltd.) and boiled at 95°C for 10 min. The treated
samples were subjected to 12% SDS-PAGE and transferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). Nonspecific binding was blocked using 5% non-fat powdered
milk (dissolved in PBS) at room temperature for 1 h and the blots
were incubated overnight at 4°C with the following antibodies:
Bcl-2 (cat. no. 3498; 1:1,000), Bax (cat. no. 14796; 1:3,000) and
GAPDH (cat. no. G9545; 1:5,000). The following day, all the
membranes were washed three times with PBS supplemented with 0.1%
Tween-20 prior to incubation with the horseradish
peroxidase-conjugated goat anti-rabbit IgG secondary antibody (cat.
no. FD0128; 1:5,000; Fude Biological Technology Co., Ltd.) for 1 h
at room temperature. Finally, the membranes were washed with PBS
supplemented with 0.1% Tween-20 three times for 5 min each. The
bands were visualized by chemiluminescence reagents according to
the manufacturer's protocols (EMD Millipore) and visualized on the
ChemiDoc XRS+ Imaging System (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Image Lab Software (version 3.0; Bio-Rad Laboratories,
Inc.) was used to quantify band densitometry. A total of three
independent experiments were performed.
Evaluation of cellular ROS by FCM
For the evaluation of the expression levels of ROS,
OVCAR3 cells that had been treated with 1.5 mM TPL at 37°C for 12 h
were harvested and washed twice with PBS. The cells were suspended
in 0.5 ml PBS and incubated with 20 µM DCFH-DA, a cell membrane
permeable fluorescence probe, for 30 min at 37°C. Subsequently, the
cells were washed and suspended in 0.5 ml PBS for analysis.
Fluorescence levels were detected by FCM analysis using FACS Aria
II (BD Biosciences) and analyzed by FlowJo Software. Each sample
was analyzed three times in three independent experiments.
Statistical analysis
Results represent data from triplicate experiments
for each treatment group and are presented as the mean ± standard
deviation. Student's t-test was performed for comparisons between 2
groups and one-way analysis of variance with the
Student-Newman-Keuls post-hoc test for comparisons between >2
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
TPL inhibits the proliferation of
OVCAR3 and SKOV3 cells
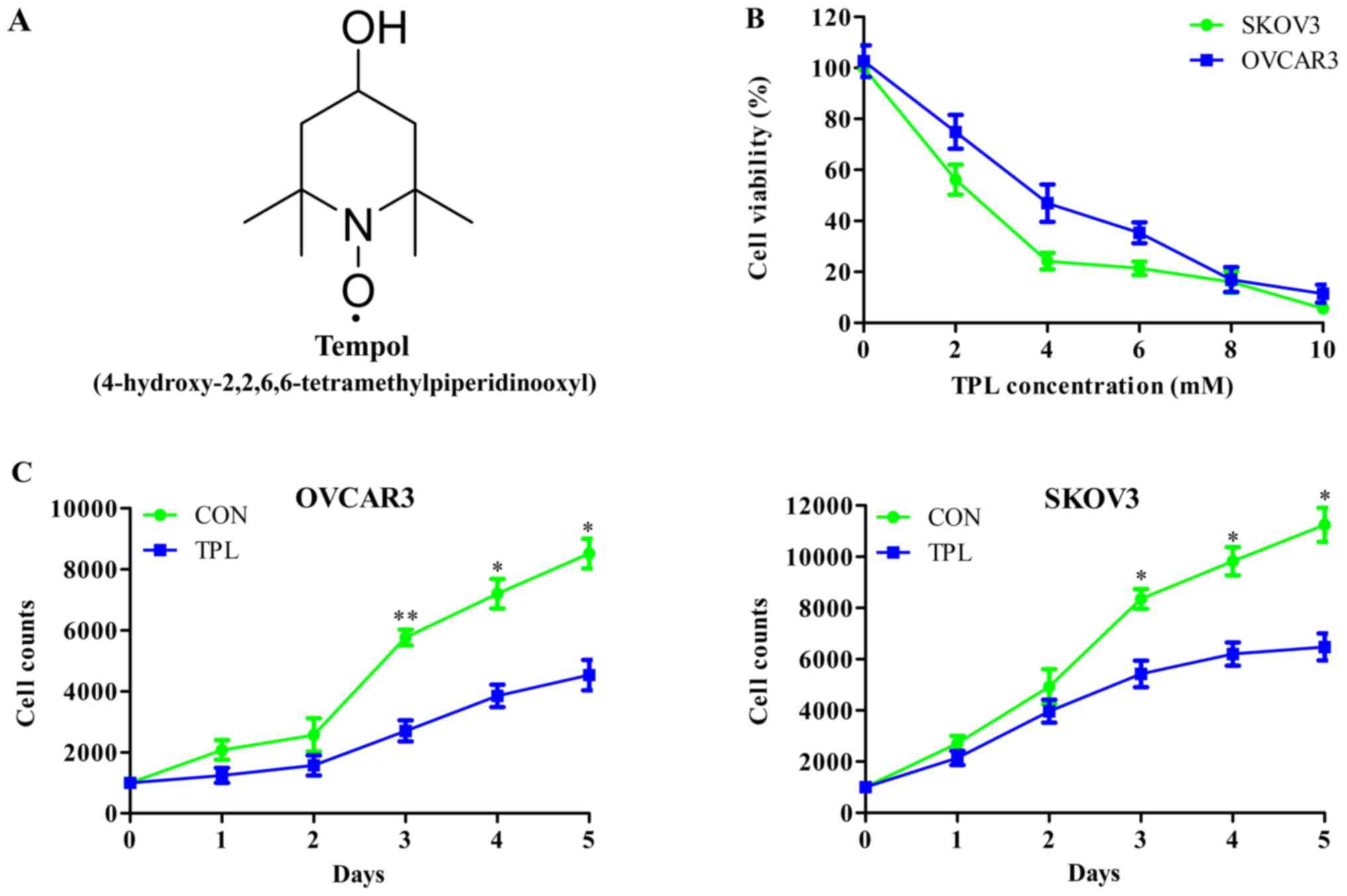
Fig. 1A depicted the
chemical structure of TPL. In order to investigate the effect of
TPL on cell proliferation, the present study performed an MTT assay
to evaluate cell viability following TPL treatment with
concentrations >1 mM. The cell viabilities of OVCAR3 and SKOV3
cells were suppressed by TPL in a dose-dependent manner. The
half-maximal inhibitory concentration (IC50) values of
TPL in OVCAR3 and SKOV3 cells were 3.72 and 2.32 mM, respectively,
after 48 h incubation (Fig. 1B).
Based on the IC50, a low concentration of drug without
cytotoxicity was used. Therefore, the present study selected the
concentration of 1.5 mM TPL in the OVCAR3 group and 1 mM TPL in the
SKOV3 group to analyze the effect of TPL on cell proliferation.
Cell growth curves indicated that the proliferation rate of OVACR3
cells was significantly inhibited by TPL, compared with the control
group, from the third day (third day, P=0.0097; fourth day,
P=0.0113; fifth day, P=0.0155); the proliferation rate of SKOV3
cells was also inhibited by TPL, compared with the control group,
from the third day (third day, P=0.0286; fourth day, P=0.0239;
fifth day, P=0.0174) (Fig. 1C). These
results suggested that TPL at concentrations of ≥1 mM were toxic
and suppressed cell proliferation in OVCAR3 and SKOV3 cells.
TPL increases DDP-induced
proliferation inhibition of OVCAR3 cells
It is universally recognized that DDP is the
chemotherapeutic drug for ovarian cancer in clinical practice
(28). MTT assays were used in the
present study determined the IC50 of DDP in the
experimental system. Fig. 2A
indicated that the IC50 of DDP was 4.71 µM following
incubation for 48 h. To investigate whether TPL increased the
effect of DDP on cell proliferation inhibition, OVCAR3 cells were
incubated with a combination of 3 µM DDP and 1.5 or 2.0 mM TPL.
Each concentration of TPL significantly increased DDP-induced cell
proliferation inhibition, compared with the signal DDP treatment
group (Fig. 2B), suggesting that TPL
improved the effect of DDP on inhibiting cell proliferation. When
incubated with TPL alone, the higher concentration of 2.0 mM
revealed a slightly higher toxicity compared with low concentration
of 1.5 mM TPL, but no significant statistical difference was
observed. Following treatment of TPL combined with DDP, the high
TPL concentration group exhibited significant proliferation
inhibition in OVCAR3 cells compared with the low TPL concentration
group (P<0.05), indicating that the proliferation inhibition
effect of TPL was more marked at higher concentrations.
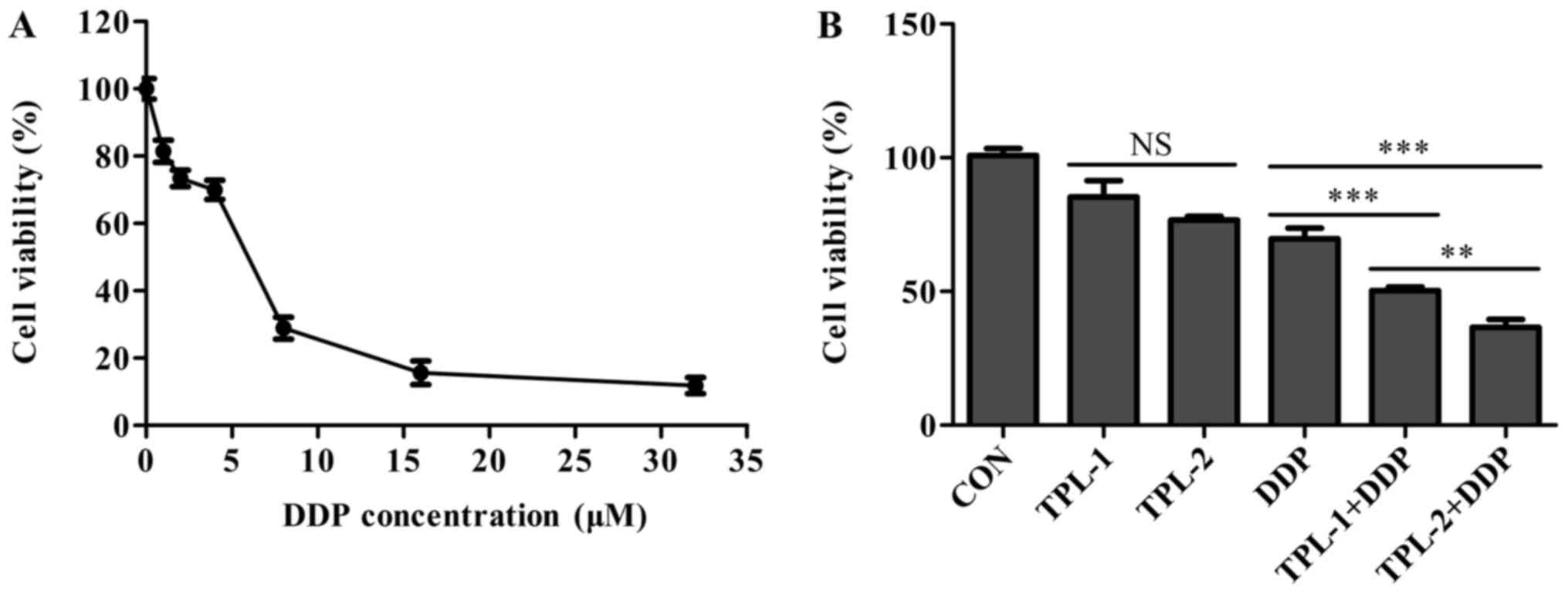 | Figure 2.Effect of TPL on DDP-induced
proliferation inhibition. OVCAR3 cells were incubated with
increasing concentrations of DDP (1, 2, 4, 6, 8, 16 or 32 µM) for
48 h. The proliferation of OVCAR3 cells with combination treatment
of TPL (1.5 and 2 mM) and DDP (3 µM) was increased compared with
the DDP-only treatment. (A) Changes in cell viability following DDP
treatment. (B) Changes in cell viability from the DDP and TPL alone
treatments compared with the combination treatment. **P<0.01 and
***P<0.001 vs. corresponding DDP group. NS, non-significant;
TPL, Tempol; DDP, cisplatin; CON, control' TPL-1, 1.5 mM TPL;
TPL-2, 2 mM TPL. |
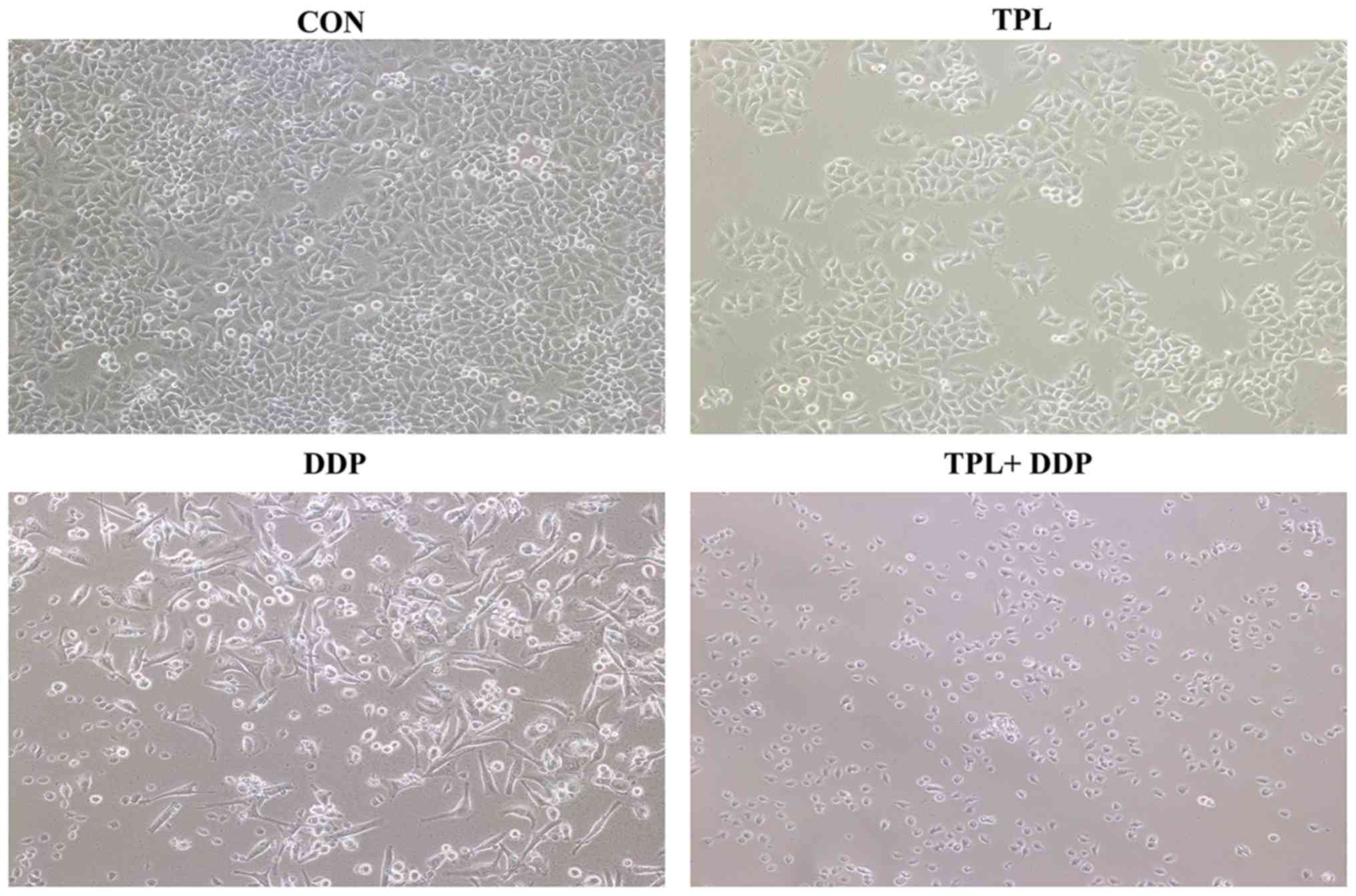
Morphological observation of OVCAR3
cells following combination treatment of TPL and DDP
Morphological observations of OVCAR3 cells following
treatment with TPL or DDP or the combination of two drugs are
demonstrated in Fig. 3. OVCAR3 cells,
cultured in the control group, were fibroblasts-like which had a
long and flat shape with irregular protrusions. Following TPL
treatment for 48 h, the cell shape was not altered significantly,
while the cell number was decreased compared with the control
group. In the DDP treatment group, the cell shape changed,
including some cells shriveling and others appearing round in
shape. In the combination treatment group, the cell number was
markedly decreased compared with the DDP-only treatment, and the
majority of the cells were round in shape and had not attached to
the culture dish. The combination treatment decreased cell
proliferation of OVCAR3 cells more effectively compared with
treatment with TPL or DDP alone.
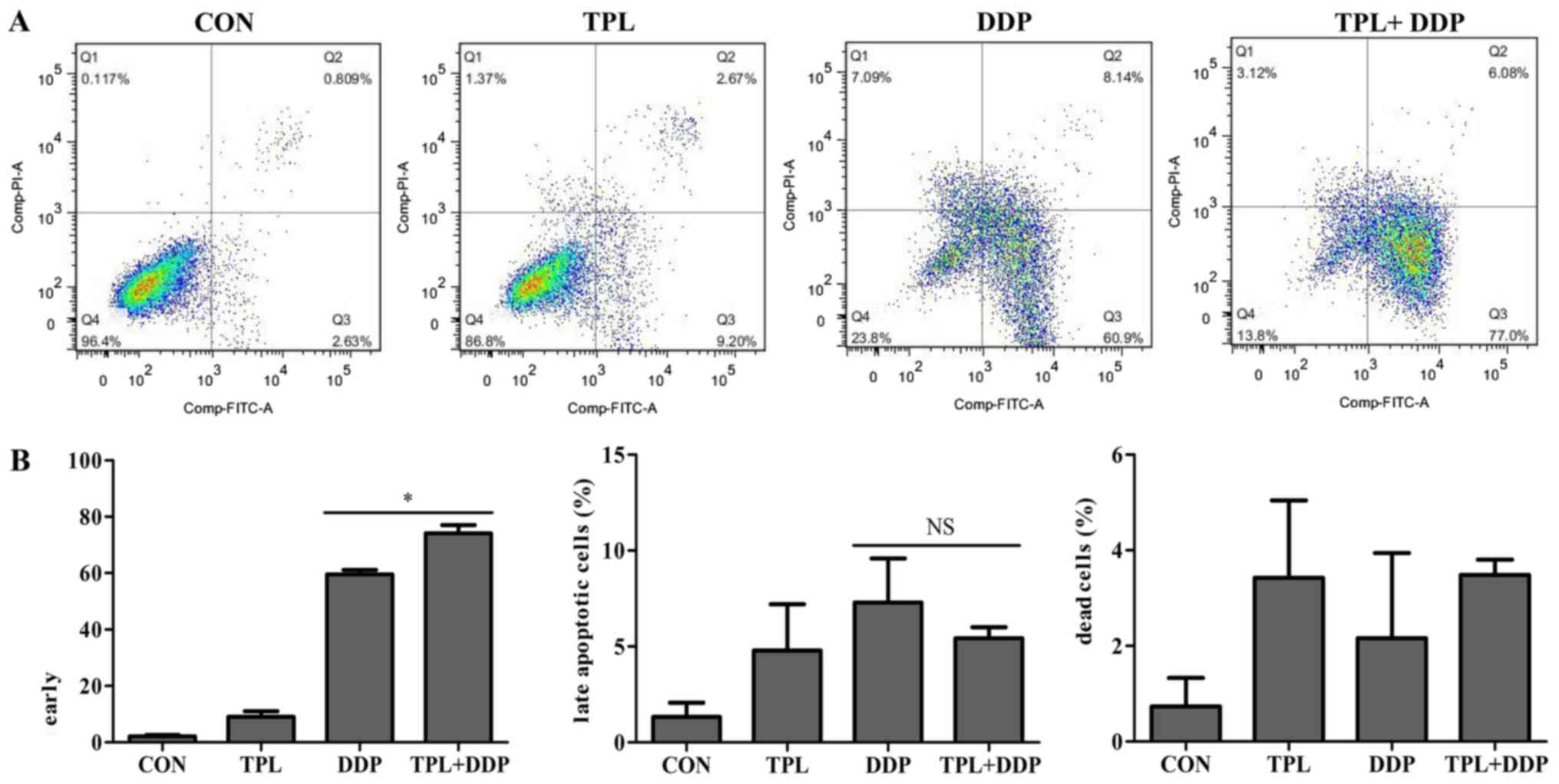
TPL improves DDP-induced apoptosis of
OVCAR3 cells
The effect of DDP in cancer therapy is dependent on
the induction of cell apoptosis. To evaluate the apoptotic stages
of the cells following stimulation with the combination treatment,
FCM analysis was used to distinguish the early apoptotic cells
stained with Annexin V-FITC and the late apoptotic cells stained
with PI. Representative cytograms for each treatment group are
presented in Fig. 4A. Stimulation of
OVCAR3 cells with the combination of 1.5 mM TPL and 3 µM DDP for 48
h produced a significant increase in the percentage of early
apoptotic cells (Annexin+/PI-; 74.1±2.9%) compared with the DDP
alone treatment (59.52±1.58%; Fig.
4B; P=0.0121). The percentage of late apoptotic cells in the
combination group (Annexin+/PI+; 5.43±0.58%) was similar to the DDP
group (7.29±2.3%; Fig. 4B). This
experiment indicated that TPL accelerated DDP-induced apoptosis,
primarily reflected in the increase in proportion of early
apoptotic cells.
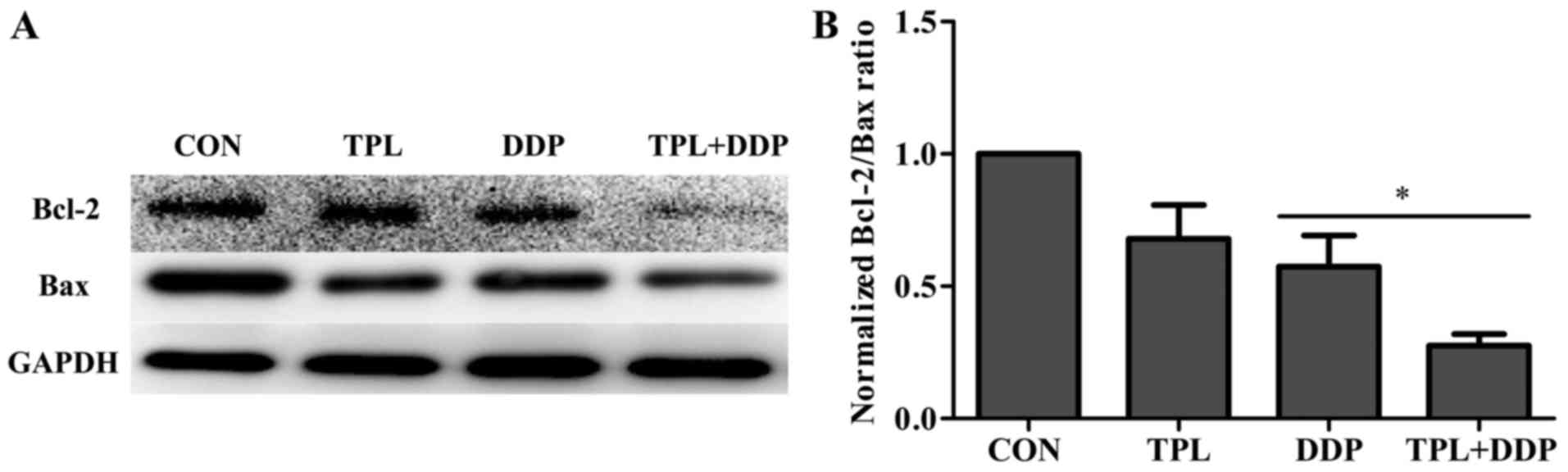
TPL decreases DDP-induced expression
ratio of Bcl-2:Bax in OVCAR3 cells
The expression ratio of Bcl-2:Bax serves an
important role in cellular apoptosis. To verify whether the
combination treatment was able to increase the apoptosis rate of
OVCAR3 cells, western blot analysis was used to detect the
expression of anti-apoptotic protein Bcl-2 and pro-apoptotic
protein Bax, and the expression ratio of Bcl-2:Bax. Fig. 5A depicted the protein expression after
TPL treatment, DDP treatment and combination treatment. The data in
Fig. 5B indicates that the
combination treatment decreased the protein level of Bcl-2 and the
expression ratio of Bcl-2:Bax induced by DDP (P=0.028). These
results suggest that the TPL treatment significantly promoted
DDP-induced apoptosis in OVCAR3 cells.
TPL increases the DDP-induced
apoptosis through ROS generation in OVCAR3 cells
To investigate the potential mechanism of
TPL-increased DDP-induced apoptosis in OVCAR3 cells, the cellular
ROS generation induced by TPL and DDP in OVCAR3 cells was examined
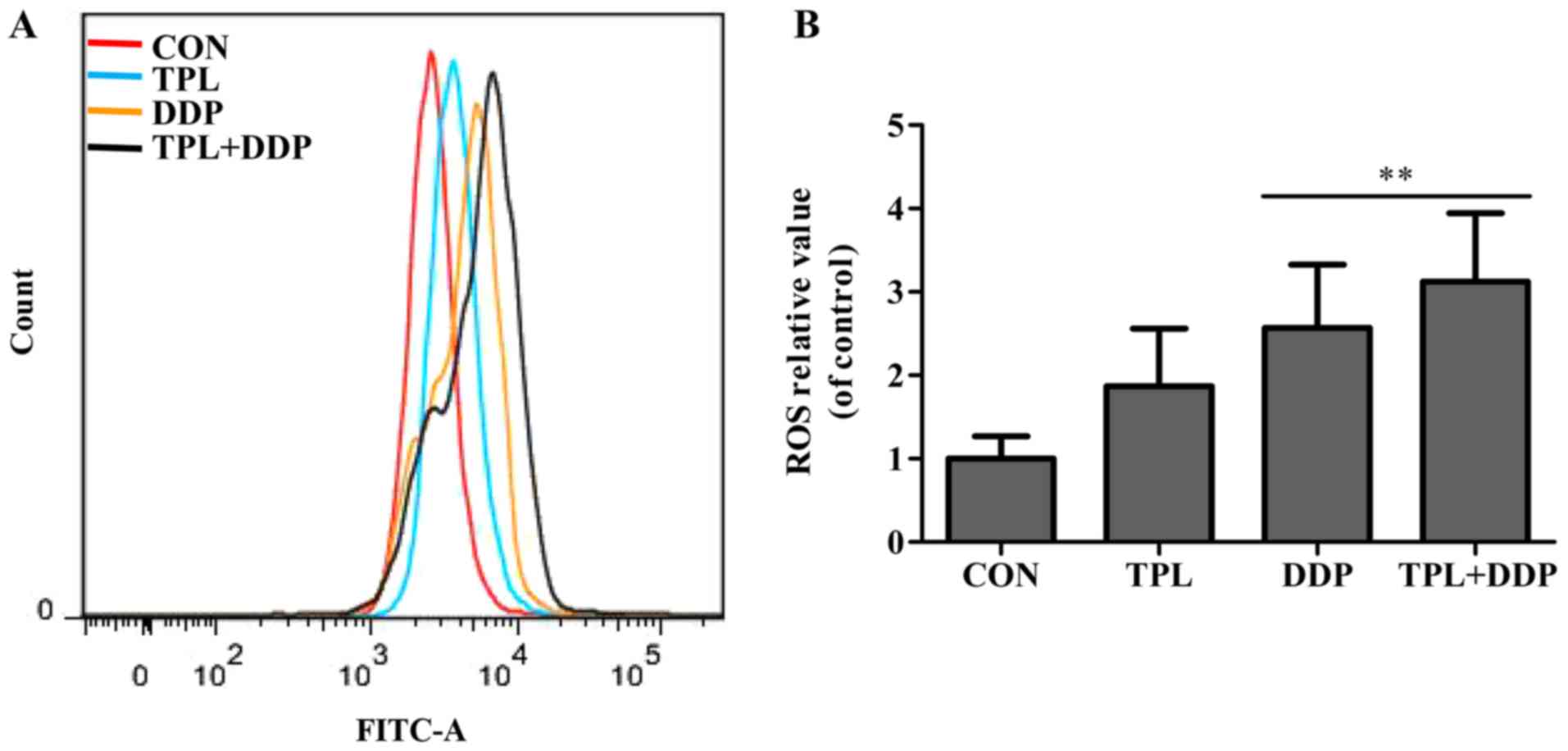
with DCFH-DA staining. As depicted in Fig. 6A, following treatment with TPL or DDP
for 12 h, TPL and DDP increased the fluorescence intensity of ROS
in OVCAR3 cells compared with the control group. In addition, the
fluorescence intensity of ROS in the combined treatment group was
significantly increased compared with the DDP alone group
(P=0.0035; Fig. 6B). Therefore, it
was concluded that TPL improved the cellular ROS production induced
by DDP, suggesting that TPL-increased DDP-induced apoptosis may be
associated with an increase in DDP-induced ROS generation.
Discussion
DDP chemotherapy is one of the primary treatments
for ovarian cancer and has been applied for a number of years,
despite the fact that DDP treatment alone does not demonstrate
satisfactory effects in current clinical practice. Drug resistance
has emerged as the major impediment to effective ovarian cancer
therapy (3–5). Therefore, novel strategies that are able
to improve the effects of DDP urgently required. TPL, a cyclic
nitroxide, protects normal organs from oxidative damage and notably
inhibits the growth of neoplastic cells, compared with the growth
of normal cells (29), which suggests
that TPL may be a promising agent for cancer treatment. The present
study investigated whether TPL was able to potentiate the effect of
DDP on chemotherapy-induced apoptosis in ovarian cancer cells.
Firstly, the combination treatment of TPL and DDP significantly
inhibited the proliferation of OVCAR3 cells, as indicated by MTT
assay. Subsequently, FCM analysis indicated that the effect of DDP
in inducing apoptosis was markedly increased following TPL and DDP
combination therapy in OVCAR3 cells. This effect may provide the
possibility of a decrease in the effective dose of DDP in clinical
therapy, and consequent decrease in the dose-associated toxicity of
DDP.
ROS generation, a hallmark of apoptosis induction,
has been demonstrated to be associated with an inhibition of the
mitochondrial respiration chain, leading to rupture of and
alterations to the membrane potential of mitochondria (30). It has also been demonstrated that ROS
generation consequently leads to the release of cytochrome C from
the mitochondria to the cytosol, and activation of the
mitochondrial apoptotic pathway (31). Bcl-2 and Bax serve important roles in
regulating caspase-dependent and caspase-independent apoptosis,
mediated by the mitochondrial pathway. These types of apoptosis are
accompanied by an upregulation of Bax and downregulation of Bcl-2,
or downregulation of the Bcl-2:Bax expression ratio, and promotion
of the formation of apoptosome (32).
Western blot analysis demonstrated that co-treatment markedly
decreased the protein level of Bcl-2 and the Bcl-2:Bax expression
ratio compared with the DDP-only treatment. Previous studies
demonstrated that TPL has an unpaired electron and interferes with
the electron transport carriers in the mitochondria, particularly
Complex I, leading to the formation of ROS (25,27). The
DCFH-DA staining assay in the present study indicated that TPL
increased the cellular ROS generation induced by DDP. These results
indicated that the combination treatment of TPL and DDP induced
apoptosis through increasing the cellular ROS level in OVCAR3
cells. However, there are potential harmful interactions between
TPL with enzyme complexes that may alter essential cell functions,
including inhibition of glucose transport and impairing of
mitochondrial adenosine 5′-triposphate production (33,34).
Therefore, the potential side effects of TPL may result in damage
to the function of some normal cells.
In conclusion, the present study identified that TPL
increases the rate of DDP-induced apoptosis in OVCAR3 cells,
highlighting the significant therapeutic implications of TPL
combined with DDP in OC. Whether this combination will affect other
types of human cancer cells similarly remains to be determined.
Acknowledgements
The authors thank Guangzhou Key Laboratory of Tumor
Immunology Research, Southern Medical University, Guangzhou, China,
for providing experimental platforms.
Funding
The present study was financially supported by the
National Natural Science Foundation of China (grant no. 81472834),
Guangzhou Science and Technology Research project (grant no.
201400000001-1), National Major Basic Research Program of China
(grant no. 2010CB529401), National Natural Science Foundation of
China (grant no. 61427807) and the Introduced Major Research and
Development Project Funded by Fujian Province (grant no.
2012I2004).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MW participated in the design of the study,
performed all experiments and wrote the manuscript. KL assisted
performing cells cultures and western blotting. ZZ assisted
performing flow cytometry analysis. LL assisted performing inverted
phase contrast microscopy observation. QW, LZ and WG helped with
the statistical analysis. YW, WH, RL and KY participated in the
design of the study. QL conceived of the study and assisted editing
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Salehi F, Dunfield L, Phillips KP, Krewski
D and Vanderhyden BC: Risk factors for ovarian cancer: An overview
with emphas is on hormonal factors. J Toxicol Environ Health B Crit
Rev. 11:301–321. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Markman M: Pharmaceutical management of
ovarian cancer: Current status. Drugs. 68:771–789. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mckeage MJ: Comparative adverse effect
profiles of platinum drugs. Drug Saf. 13:228–244. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aisner J, Jacobs M, Sinabaldi V, Gray W
and Eisenberger M: Chemoradiotherapy for the treatment of
regionally advanced head and neck cancers. Semin Oncol. 21 5 Suppl
12:S35–S44. 1994.
|
|
5
|
Ali BH and Al Moundhri MS: Agents
ameliorating or augmenting the nephrotoxicity of cisplatin and
other platinum compounds: A review of some recent research. Food
Chem Toxicol. 44:1173–1183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Santos NA, Catao CS, Martins NM, Curti C,
Bianchi ML and Santos AC: Cisplatin-induced nephrotoxicity is
associated with oxidative stress, redox state unbalance, impairment
of energetic metabolism and apoptosis in rat kidney mitochondria.
Arch Toxicol. 81:495–504. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawanishi S, Hiraku Y, Pinlaor S and Ma N:
Oxidative and nitrative DNA damage in animals and patients with
inflammatory diseases in relation to inflammation-related
carcinogenesis. Biol Chem. 387:365–372. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Janssen-Heininger YM, Mossman BT, Heintz
NH, Forman HJ, Kalyanaraman B, Finkel T, Stamler JS, Rhee SG and
van der Vliet A: Redox-based regulation of signal transduction:
principles, pitfalls, and promises. Free Radic Biol Med. 45:1–17.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ames BN, Shigenaga MK and Hagen TM:
Oxidants, antioxidants, and the degenerative diseases of aging.
Proc Natl Acad Sci USA. 90:7915–7922. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schafer ZT, Grassian AR, Song L, Jiang Z,
Gerhart-Hines Z, Irie HY, Gao S, Puigserver P and Brugge JS:
Antioxidant and oncogene rescue of metabolic defects caused by loss
of matrix attachment. Nature. 461:109–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Perry G, Raina AK, Nunomura A, Wataya T,
Sayre LM and Smith MA: How important is oxidative damage? Lessons
from Alzheimer's disease. Free Radic Biol Med. 28:831–834. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moloney JN and Cotter TG: ROS signalling
in the biology of cancer. Semin Cell Dev Biol. 80:50–64. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brasch RC, London DA, Wesbey GE, Tozer TN,
Nitecki DE, Williams RD, Doemeny J, Tuck LD and Lallemand DP: Work
in progress: Nuclear magnetic resonance study of a paramagnetic
nitroxide contrast agent for enhancement of renal structures in
experimental animals. Radiology. 147:773–779. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tabaczar S, Talar M and Gwoździński K:
Nitroxides as antioxidants-possibilities of their application in
chemoprevention and radioprotection. Postepy Hig Med Dosw (Online).
65:46–54. 2011.(In Polish). View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Saito K, Takeshita K, Ueda J and Ozawa T:
Two reaction sites of a spin label, TEMPOL
(4-hydroxy-2,2,6,6-tetramethylpiperidine-N-oxyl), with hydroxyl
radical. J Pharm Sci. 92:275–280. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krishna MC, Grahame DA, Samuni A, Mitchell
JB and Russo A: Oxoammonium cation intermediate in the
nitroxide-catalyzed dismutation of superoxide. Proc Natl Acad Sci
USA. 89:5537–5541. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wilcox CS and Pearlman A: Chemistry and
antihypertensive effects of tempol and other nitroxides. Pharmacol
Rev. 60:418–469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Volk T, Hensel M, Schuster H and Kox WJ:
Secretion of MCP-1 and IL-6 by cytokine stimulated production of
reactive oxygen species in endothelial cells. Mol Cell Biochem.
206:105–112. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cuzzocrea S, McDonald MC, Mota-Filipe H,
Mazzon E, Costantino G, Britti D, Mazzullo G, Caputi AP and
Thiemermann C: Beneficial effects of tempol, a membrane-permeable
radical scavenger, in a rodent model of collagen-induced arthritis.
Arthritis Rheum. 43:320–328. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Di Paola R, Mazzon E, Zito D, Maiere D,
Britti D, Genovese T and Cuzzocrea S: Effects of Tempol, a
membrane-permeable radical scavenger, in a rodent model
periodontitis. J Clin Periodontol. 32:1062–1068. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moosmann B and Behl C: Antioxidants as
treatment for neurodegenerative disorders. Expert Opin Investig
Drugs. 11:1407–1435. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grasbon-Frodl EM, Kösel S, Riess O, Müller
U, Mehraein P and Graeber MB: Analysis of mitochondrial targeting
sequence and coding region polymorphisms of the manganese
superoxide dismutase gene in German Parkinson disease patients.
Biochem Biophys Res Commun. 255:749–752. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schnackenberg CG and Wilcox CS: Two-week
administration of tempol attenuates both hypertension and renal
excretion of 8-Iso prostaglandin f2alpha. Hypertension. 33:424–428.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Monti E, Supino R, Colleoni M, Costa B,
Ravizza R and Gariboldi MB: Nitroxide TEMPOL impairs mitochondrial
function and induces apoptosis in HL60 cells. J Cell Biochem.
82:271–276. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Offer T, Russo A and Samuni A: The
pro-oxidative activity of SOD and nitroxide SOD mimics. FASEB J.
14:1215–1223. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gariboldi MB, Rimoldi V, Supino R, Favini
E and Monti E: The nitroxide tempol induced oxidative stress,
p21WAF/CIP1, and cell death in HL60 cells. Free Radic Biol Med.
29:633–641. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Agarwal R and Kaye SB: Ovarian cancer:
Strategies for overcoming resistance to chemotherapy. Nat Rev
Cancer. 3:502–516. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gariboldi MB, Lucchi S, Caserini C, Supino
R, Oliva C and Monti E: Antiproliferative effect of the piperidine
nitroxide TEMPOL on neoplastic and nonneoplastic mammalian cell
lines. Free Radic Biol Med. 24:913–923. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Perrone GG, Tan SX and Dawes IW: Reactive
oxygen species and yeast apoptosis. Biochim Biophys Acta.
1783:1354–1368. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Todorova VK, Harms SA, Kaufmann Y, Luo S,
Luo KQ, Babb K and Klimberg VS: Effect of dietary glutamine on
tumor glutathione levels and apoptosis-related proteins in
DMBA-induced breast cancer of rats. Breast Cancer Res Treat.
88:247–256. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alpert E, Gruzman A, Totary H, Kaiser N,
Reich R and Sasson S: A natural protective mechanism against
hyperglycemia in vascular endothelial and smooth-muscle cells: Role
of glucose and 12-hydroxyeicosatetraenoic acid. Biochem J.
362:413–422. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Behrooz A and Ismail-Beigi F: Stimulation
of glucose transport by hypoxia: Signals and mechanisms. News
Physiol Sci. 14:105–110. 1999.PubMed/NCBI
|