Introduction
In United States, prostate cancer is a major threat
to the health of men (1), and, in
China, the incidence of prostate cancer has increased (2). Previous studies have revealed that
androgen serves an important role in the development of prostate
cancer. In the early stage of prostate cancer, androgen is
essential for the proliferation of the cancer cells, and is also
crucial in the recurrent or metastatic stage (3,4). Androgen
deprivation therapy (ADT) successfully inhibits this process in
patients (5,6). Methods of ADT include castration and
anti-androgen therapy, however, ADT presents numerous
disadvantages, including surgical trauma and complication, and
heavy economic burden, particularly in China (7). Therefore, it is important to identify
novel and more economical therapies for prostate cancer, and
inhibition of the histamine receptor has been suggested to have
potential (8,9).
Histamine receptor includes 4 subtypes, namely,
histamine h1 receptor (H1R), histamine h2 (H2R), h3 (H3R) and h4
(H4R). The histamine receptor is activated by histamine and
restrained by histamine receptor antagonists. In 1979, Armitage
(8) reported that a histamine
receptor antagonist (cimetidine) had an antitumor effect, however,
the mechanism was uncharacterized. Since, the majority of scholars
believe that histamine and histamine receptors serve a role in the
promotion in tumor growth and invasion, and histamine receptor
antagonists, the contrary (10–13). In
in vivo trials, histamine receptor antagonists have been
reported to increase the release of prolactin in male animals
(14). However, the proposed
mechanisms of the role of histamine in regulation of the release of
androgen are various, and Wang et al (9) suggest that histamine functions without
the histamine receptor. H3R has been demonstrated to be
overexpressed in multiple types of cancer (12,13), and
the present study demonstrated that histamine and H3R were
overexpressed in prostate cancer cells compared with normal
prostatic epithelial cells. Therefore, we hypothesized that H3R has
a potential role in the development of prostate cancer and aimed to
explore the underlying mechanism.
The present study can be divided into four parts: i)
Exploration of H3R expression in prostate cancer cell lines by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR), western blotting and immunohistochemistry of a prostate
cancer tissue array; ii) inhibition of the expression of H3R in
prostate cancer cell lines by antagonists or small-interfering RNA
(siRNA) to determine the role of H3R in cancer cells; iii)
investigate downstream signaling pathways following si-H3R
transfection, and iv) verify the effect of si-H3R on a nude mouse
xenograft model. Evidence indicates that H3R is involved in
multiple tumor processes, and the present study may broaden our
understanding of the underlying pathological mechanisms, as well as
aid in the discovery of novel treatment targets.
Materials and methods
Cell lines and prostate cancer tissue
chip
The prostate cancer cell lines, PC-3, LNCaP and
22RV1, were purchased from the Institute of Biochemistry and Cell
Biology (Chinese Academy of Sciences, Shanghai, China). C4-2 cells
were a gift from Dr Guo-wen Lin (Fudan University Shanghai Cancer
Center, Shanghai, China). The normal human prostate epithelial cell
line, RWPE-1, was purchased from the American Type Culture
Collection (Manassas, VA, USA). PC-3, LNCaP, C4-2 and 22RV1 cells
were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum (FBS) and 1% penicillin-streptomycin (all from Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). RWPE-1 cells
were cultured in keratinocyte serum-free medium (K-SFM; cat. no.
17005-042; Invitrogen; Thermo Fisher Scientific, Inc.) supplemented
with 0.05 mg/ml bovine pituitary extract (BPE), 5 ng/ml epidermal
growth factor (EGF; both from K-SFM medium kit; cat. no. 17005-042;
Invitrogen; Thermo Fisher Scientific, Inc), according to the
manufacturer's protocols, and 1% of penicillin-streptomycin
(Gibco). All media was changed every 2–3 days, and the cells were
passaged at 80–90% confluency. The prostate cancer tissue array was
purchased from Biomax USA (US Biomax, Inc., Derwood, MD, USA).
Cytotoxicity assays
Cell Counting kit-8 (CCK-8) assays were performed by
plating 5×103 LNCaP cells per well in a 96 well plate
(Thermo Fisher Scientific, Inc.). The following day, cells were
treated with 0, 10, 100 nM, and 1, 5, 10 µM of
R-(a)-methylhistamine (RAMH; cat. no. B5010; Apexbio, Houston, TX,
USA) and 0, 10, 100 nM and 1, 5, 10 µM ciproxifan (CPX; cat. no.
S2813; Selleck, Houston, TX, USA). The cells were exposed to the
drugs for 1–2 days prior to the addition of CCK-8 reagent (cat. no.
40203ES60; Yeasen; Shanghai Yi San Biotechnology Co., Ltd.,
Shanghai, China) for 2 h, according to the manufacturer's protocol,
and the absorbance of each well was detected using a microplate
reader (Thermo Fisher Scientific, Inc) at a wavelength of 450
nm.
H3R-knockdown studies
Transient knockdown was performed using siRNA
targeting H3R (Ruibobio, Guangzhou, Guangdong, China) and
non-targeting siRNA as a negative control. The siRNA sequences were
as follows: si-H3R, 5′-GCTATGCCGAGTTCTTCTACA-3′, and negative
control, 5′-TTCTCCGAACGTGTCACGT-3′. In brief, LNCaP cells
(5×104 in 24-well plate) were transfected with siRNA
(both 20 pmol in 500 µl/well) with Lipofectamine 2000 (1 µl in 500
µl/well, Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. The LNCaP cells used for transplant
were transfected with si-H3R-packaged lentivirus (MOI=20) and
selected using puromycin (2 µg/ml for 10 days) (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) according to the manufacturer's
protocol.
Migration and invasion assays
The upper chamber of a 24-well plate (Corning
Incorporated, Corning, NY, USA) was pre-coated with 50 µl Matrigel
(BD Biosciences, Franklin Lake, NJ, USA) for the invasion assay.
The following steps were the same for the two assays. A total of
1×105 LNCaP cells were suspended in 200 µl serum-free
RPMI-1640 medium and seeded into the upper chamber of a 24-well
plate. A total of 900 µl RPMI-1640 medium supplemented with 20% FBS
was added to lower chamber of each well. After 24 h, the cells were
fixed using 4% paraformaldehyde for 30 min at room temperature
(Mairel; Shanghai Miner Chemical Technology Co., Ltd., Shanghai,
China) and stained with 0.1% crystal violet (Mairel; Shanghai Miner
Chemical Technology Co., Ltd) for 30 min at room temperature. Cells
on the upper surface of the Transwell insert were removed with a
cotton swab and cells on the lower surface were counted under a
light microscope (Olympus Corporation, Tokyo, Japan) in three
fields of view (magnification, ×100).
Apoptosis assay
Measurement of in vitro apoptosis was
performed by Annexin V-FITC/propidium iodide (PI) staining
Apoptosis Detection kit (Yeasen; Shanghai Yi San Biotechnology Co.,
Ltd.), according to the manufacturer's instructions, using a flow
cytometer (Beckman Coulter, Inc., Brea, CA, USA).
Reverse transcription-quantitative
polymerase chain reaction
Total RNA was extracted using a Takara RNeasy Mini
kit (cat. no. 9767; Takara Bio, Inc., Otsu, Japan), according to
manufacturer's instructions. RNA was transcribed into cDNA using a
PrimeScript RT reagent kit (cat. no. RR036A; Takara Bio, Inc.). A
Takara qPCR kit (cat. no. RR420Q; Takara Bio, Inc.) was used to
assess H3R gene expression using an ABI 7500 real-time system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions were as follows: 30 sec at 95°C, 5 sec at
95°C, 34 sec at 60°C and 40 cycles of 15 sec at 95°C, 60 sec at
60°C, and 15 sec at 95°C. Relative gene expression was normalized
to that of GAPDH. Gene expression was quantified using the
2−ΔΔCq method (15). The
primer sequences used were as follows: H3R forward,
5′-GCCACTGCTATGCCGAGTT-3′ and reverse, 5′-TGCGCCTCTGGATGTTCAG-3′;
GAPDH forward, 5′-TTTACCTTCCAGCAGCCCTA-3′ and reverse,
5′-GACAGAGTCCCAGATGAGCA-3′.
Western blotting
Total protein was extracted with
radioimmunoprecipitation assay (RIPA) buffer (Beyotime Institute of
Biotechnology, Haimen, China) adding 1 mM protease inhibitor
cocktail (Beyotime Institute of Biotechnology) at 4°C between 30
min and 1 h then concentrate under 12,000 rpm at 4°C for 15 min.
Total protein concentration was determined using a bicinchoninic
acid (BCA) kit (cat. no. P0010; Beyotime Institute of
Biotechnology, Haimen, China). The protein (30 µg per lane) samples
were resolved by 10% SDS-PAGE and transferred to polyvinylidene
fluoride (PVDF) membranes. The membranes were then blocked with 5%
non-fat milk for 1 h at room temperature. The membranes were probed
with the following primary antibodies overnight at 4°C: H3R
(dilution, 1:5,000; cat. no. ab124732; Abcam, Cambridge, UK), BAX
(BCL-2 associated X, apoptosis regulator; dilution, 1:1,000; cat.
no. D2E11), BCL2 (dilution, 1:1,000; cat. no. D17C4; Cell Signaling
Technology, Inc., Danvers, MA, USA), AR (dilution, 1:500; cat. no.
sc-7305; Santa Cruz Biotechnology, Dallas, TX, USA), GAPDH
(dilution, 1:5,000; cat. no. 70-Mab5465-040; Multi Sciences;
Hangzhou Lianke Biotechnology Co., Ltd., Hangzhou, China). The
membranes were then incubated with the appropriate horseradish
peroxidase-conjugated secondary antibody: Anti-rabbit IgG
(dilution, 1:1,000; cat. no. 7074) or anti-mouse IgG (dilution,
1:1,000; cat. no. 7076; both Cell Signaling Technology, Inc.) at
room temperature for 1 h. The two secondary antibodies were diluted
at 1:1,000 and were purchased from Cell Signaling Technology, Inc.
The blots were visualized using ECL-Plus reagent (cat. no.
WBKLS0010; Merck Millipore; Merck KGaA, Darmstadt, Germany). Images
were acquired with the ImageQuant LAS 4000mini (GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA) and analyzed with ImageJ 2.1.4.7
software (National Institutes of Health, Bethesda, MD, USA). The
results were expressed as the target protein/GAPDH ratio and
subsequently normalized to the values measured in the control
groups.
Immunohistochemistry
Prior to immunohistochemistry, the tissue array and
the xenograft tumor slides were placed in a washing solution of 3%
hydrogen peroxide (H2O2) and 60% methanol
phosphate buffered saline (PBS) (PH 7.4) for 30 min and
subsequently treated with 0.01 mol/l sodium citrate buffer for 95°C
in a microwave oven for 15 min for antigen retrieval. The tissue
array and the xenograft tumor slides were subsequently blocked in
5% normal goat serum and 5% bovine serum albumin (Yeasen; Shanghai
Yi San Biotechnology Co., Ltd) in PBS for 15 min at 37°C. Prior to
each step, the tissue array and the xenograft tumor slides were
rinsed three times in PBS buffer. The array was incubated with a
H3R primary antibody (dilution, 1:100; cat. no. ab124732; Abcam)
and the xenograft tumor slides with Ki67 (dilution, 1:400; cat. no.
9449; Cell Signaling Technology, Inc.) and TUNEL (dilution, 1:500;
cat. no. ab206386; Abcam) each for 12 h at 4°C. Subsequent to
rinsing with PBS, the sections were incubated with a secondary
horseradish peroxidase-conjugated antibody anti-rabbit IgG
(dilution, 1:200; cat. no. 7074; Cell Signaling Technology, Inc)
for 1 h at room temperature. 0.05% diaminobenzidine (Beyotime
Institute of Biotechnology) exposure was performed using chromogen
at room temperature about 5 min. Finally, the array and the
xenograft tumor slide were counterstained with 0.025% hematoxylin
(Beyotime Institute of Biotechnology) at room temperature for ~30
sec.
Xenograft tumor studies
All animal procedures were performed according to
the National Animal Experimentation guidelines and were approved by
the ethics committee of the Shanghai Jiao Tong University
(Shanghai, China). Male 4–8 week-old nude mice weighing 20–25 g
were used in the following experiments. A total of 8 mice were
divided equally and randomly into 2 groups. All mice were bred in
aseptic conditions at a constant humidity (~40–70%) between 20 and
26°C with standard 12 h light-dark cycles and free access to food
and water. For tumor xenograft studies, 5×106 LNCaP
cells were suspended in 0.1 ml serum-free media containing 50%
Matrigel, and injected subcutaneously into the oxter of the mice,
which were then observed every three days. Once the tumors reached
1 cm in diameter, the mice were sacrificed and the tumor volumes
were calculated using the following formula: 0.5 × length (mm) ×
width2 (mm2). The tumors were fixed in 10%
formalin at 4°C between 24 and 48 h, and embedded in paraffin for
incubation with the aforementioned primary antibodies, Ki67 and
TUNEL.
Statistical analysis
SPSS 22.0 (IBM Corp., Armonk, NY, USA) and Graph Pad
Prism (version 5; Graph Pad Software, Inc., La Jolla, CA, USA) were
used to analyze the data and all values are expressed as mean ±
standard deviation. One-way analysis of variance followed by the
LSD or Dunnett's T3 post-hoc tests was used for multiple
comparisons. The Mann-Whitney U test was used to compare
differences between H3R expression in prostate cancer and normal
tissues. Fisher's exact test was used to assess differences in H3R
expression in prostate cancer tissues with different Gleason
scores. P<0.05 was considered to indicate a statistically
significant differences.
Results
H3R is overexpressed in prostate
cancer tissues and cells
Previous studies have demonstrated that H3R is
overexpressed in various types of cancer, however, there are
limited reports of the expression of H3R in prostate cancer. In the
present study, the tissue array demonstrated positive
immunoreactivity of H3R in the majority of cancer tissues but
negative immunoreactivity in normal tissues, and the two tissue
groups were significantly different (P<0.01) (Fig. 1A-F; Table
I). It was demonstrated that the expression of H3R was
positively associated with Gleason score (P<0.01; Table II). RT-qPCR analysis demonstrated
that H3R mRNA is overexpressed in prostate cancer cells. In LNCaP,
22rv1 and C4-2 cells, the expression of H3R was >20-fold than
that of normal prostatic epithelial cells (P<0.01) (Fig. 1G). Western blotting demonstrated the
same result at the protein level. All cancer cell lines exhibited
higher expression of H3R than normal prostatic epithelial cells
(P<0.05). LNCaP cells exhibited the highest expression of H3R
(Fig. 1H-I), and were therefore,
selected for further experimentation. Thus, the tissue array,
RT-qPCR analysis and western blotting all suggested that expression
of H3R in prostate cancer is significantly higher than that in
normal prostatic epithelial tissues.
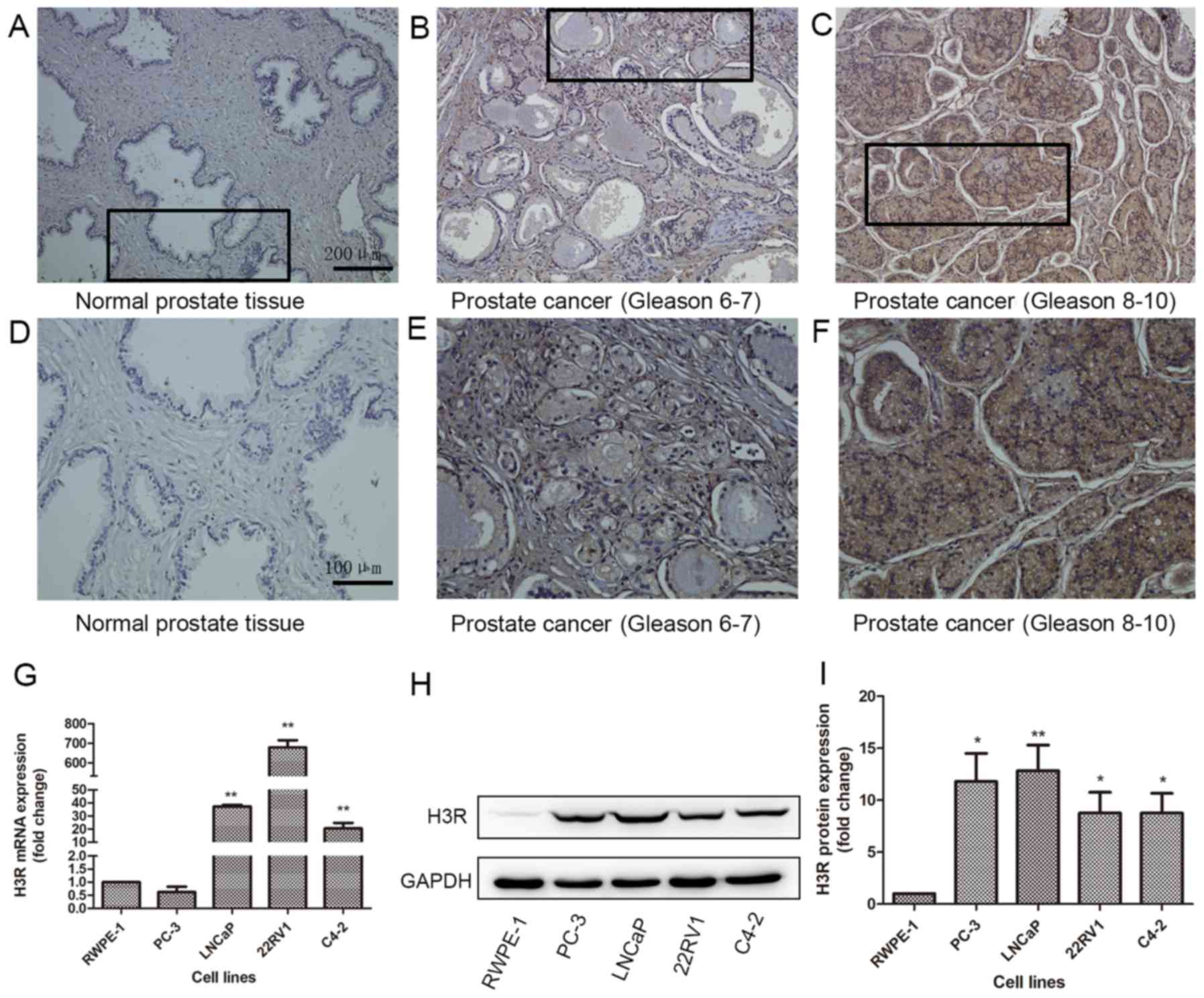 | Figure 1.Overexpression of H3R in prostate
cancer. Representative immunohistochemical images of H3R protein
expression in the prostate cancer tissue array. (A) Normal tissue,
(B) prostate cancer tissue (Gleason stage 6–7), (C) prostate cancer
tissue (Gleason stage 8–10), magnification, ×100. Enlarged images
of the outlined panels of (D) Normal tissue, (E) prostate cancer
tissue (Gleason 6–7), (F) prostate cancer tissue (Gleason stage
8–10), magnification, ×200. H3R expression measured by (G) reverse
transcription-quantitative polymerase chain reaction, and (H)
western blotting. (I) Quantification of western blotting results.
*P<0.05 vs. RWPE-1; **P<0.01 vs. RWPE-1. H3R, histamine 3
receptor. |
 | Table I.The expression of H3R protein in
different prostate tissues in the array. |
Table I.
The expression of H3R protein in
different prostate tissues in the array.
|
| H3R
immu-noreactivity |
|
|---|
|
|
|
|
|---|
| Prostate
pathology | − | + | ++ | P-value |
|---|
| Prostate cancer
(n=40) | 0 | 17 | 23 | P<0.01 |
| Normal prostate
tissue (n=8) | 5 | 3 | 0 |
|
| Table II.The association between H3R expression
and gleason score in prostate cancer. |
Table II.
The association between H3R expression
and gleason score in prostate cancer.
|
| H3R
immu-noreactivity |
|
|---|
|
|
|
|
|---|
| Gleason score in
prostate cancer (n=36) | + | ++ | Difference in H3R
expression |
|---|
| 6–7 | 13 | 1 | P<0.01 |
| 8–10 | 4 | 18 |
|
H3R regulates the proliferation of
LNCaP cells
In order to reveal the role of H3R in LNCaP cells, a
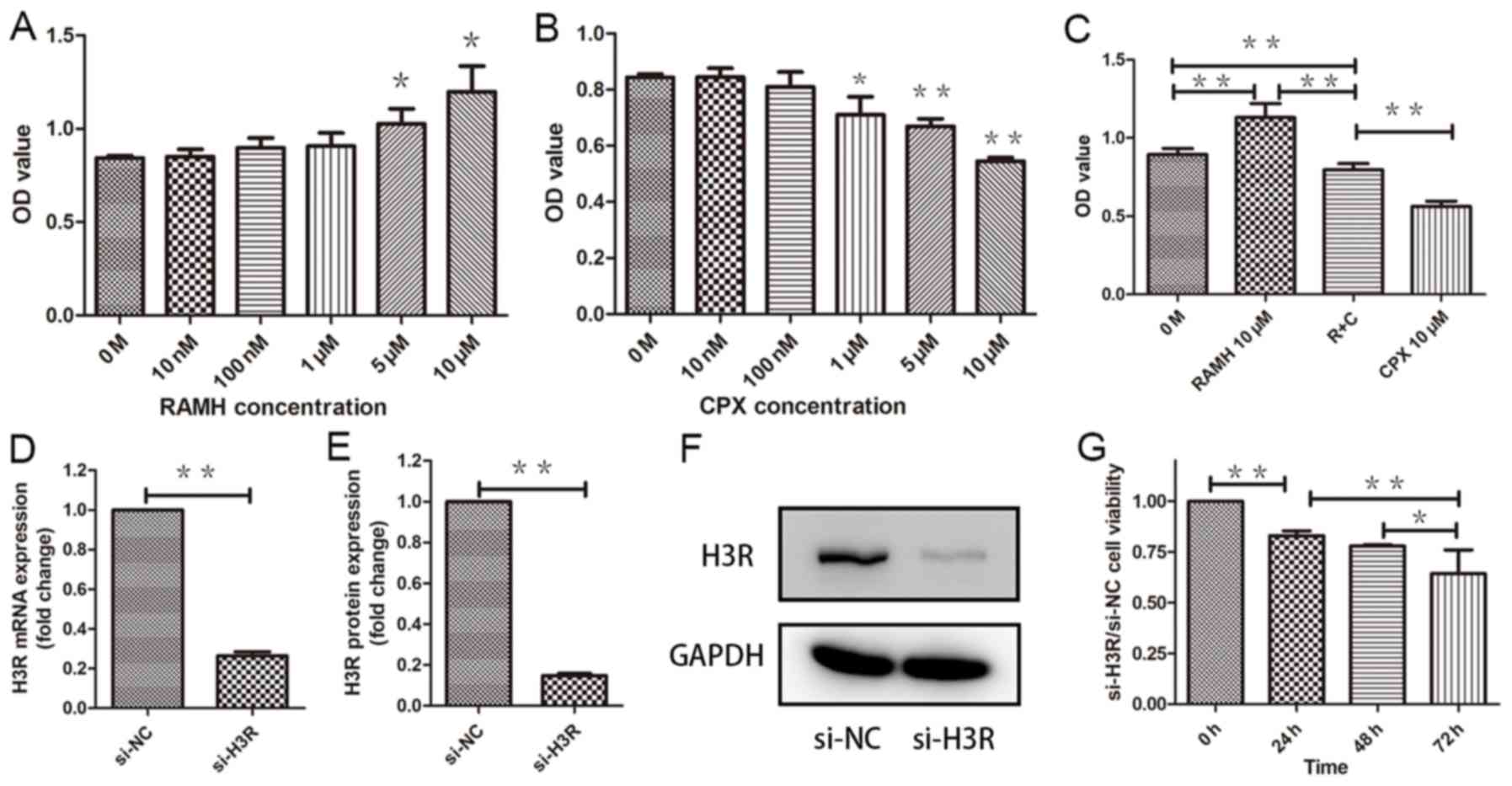
specific H3R agonist (RAMH) and antagonist (CPX) were used.
Pretreatment with RAMH promoted the proliferation of LNCaP cells in
a dose-dependent manner. In contrast, pretreatment with CPX
suppressed proliferation of LNCaP cells in a dose-dependent manner
(Fig. 2A-C). In previous studies by
our group, it was reported that histamine receptor antagonists may
function without a histamine receptor. Therefore, an siRNA was
designed to target H3R. Following si-H3R treatment of LNCaP cells
(Fig. 2D-F) their proliferation
declined by 20% more than that of si-NC LNCaP cells in one day
(P<0.01) (Fig. 2G).
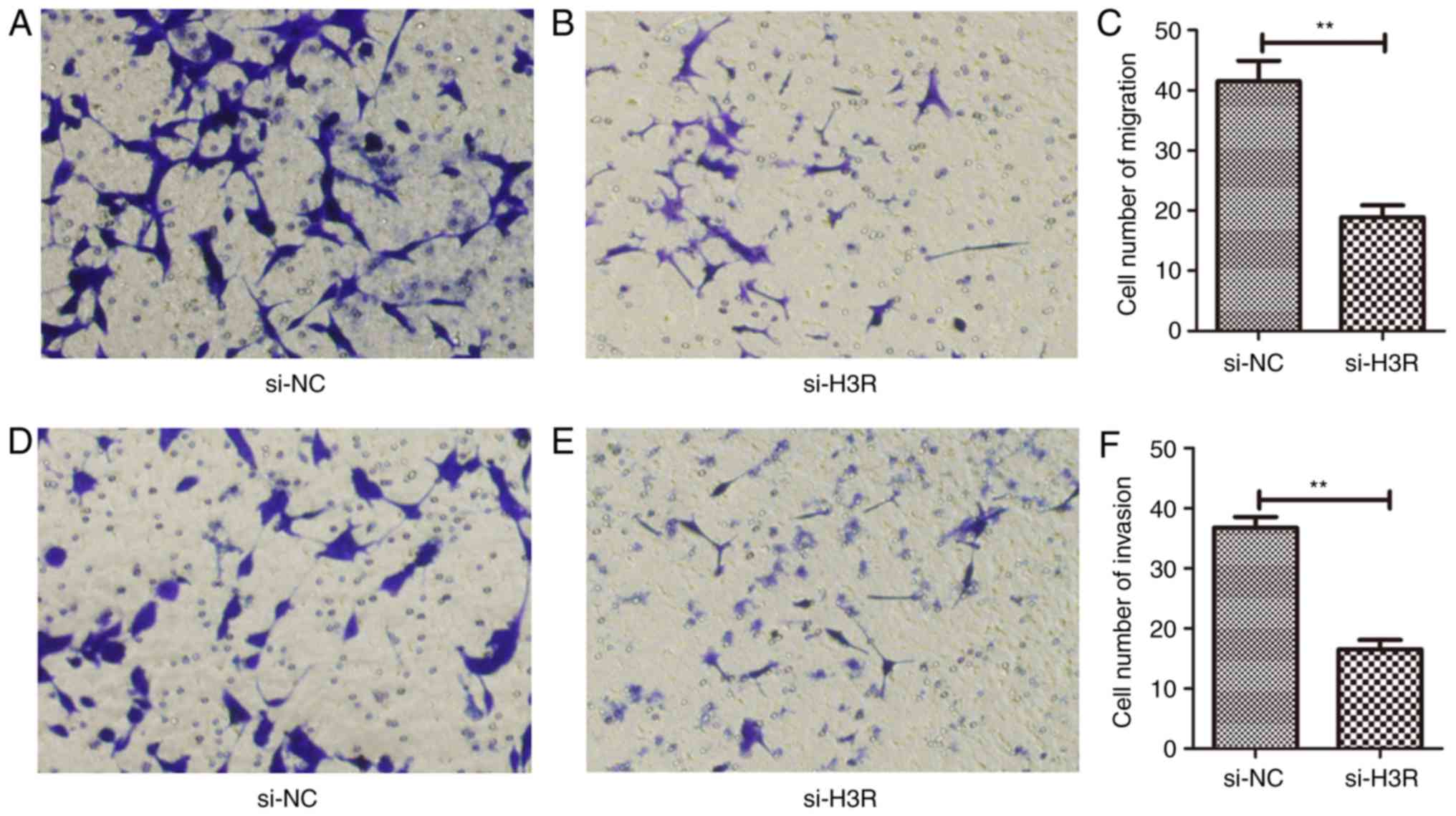
Inhibition of H3R expression
suppresses metastatic behaviour of LNCaP cells
Knockdown of H3R expression decreased the migration
(P<0.01) (Fig. 3A-C) and invasion
(P<0.01) (Fig. 3D-F) of the LNCaP
cells compared with si-NC-transfected cells.
Inhibition of H3R expression induces
apoptosis of LNCaP cells
Flow cytometric analysis of apoptosis revealed that
knockdown of H3R lead to increased apoptosis than control si-NC
cells (P<0.01) (Fig. 4E-F and H).
Treatment with 10 µM H3R agonist decreased apoptosis whereas
treatment with 10 µM H3R antagonist increased apoptosis. When used
in conjunction, the effects of both drugs were weakened (Fig. 4A-D and H). These results indicate that
knockdown of H3R lead to apoptosis of LNCaP cells.
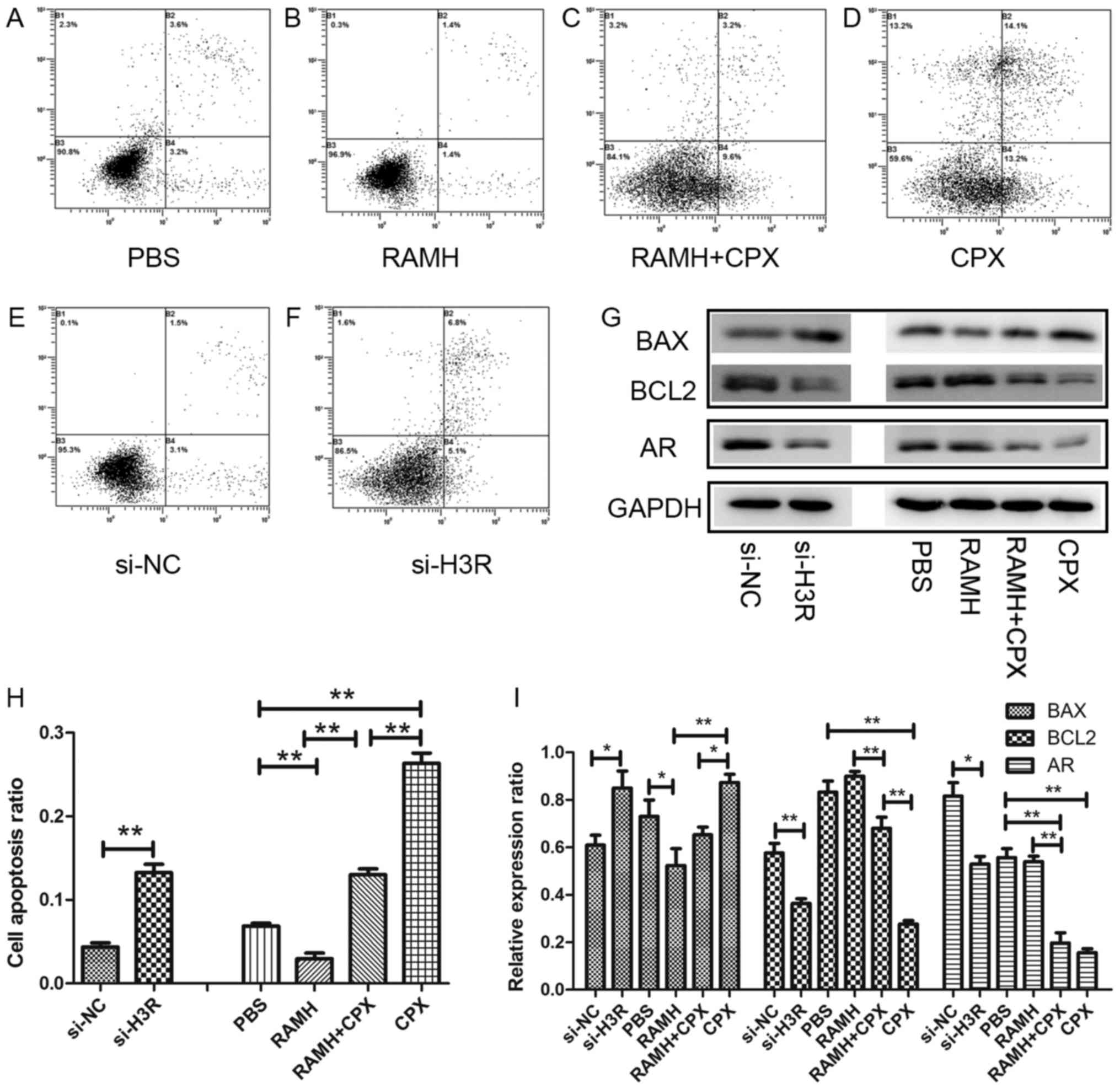 | Figure 4.Inhibition of H3R function by siRNA or
antagonist increased the apoptosis of LNCaP cells and their protein
expression profile. The effects of (A) PBS control, (B) RAMH, (C)
RAMH+CPX, (D) CPX, (E) si-NC and (F) si-H3R. (G) Western blotting
analysis of BAX, BCL-2 and AR protein expression. (H) Apoptosis of
LNCaP cells following treatment with siRNAs, RAMH and CPX. (I)
Quantification of protein expression. *P<0.05 and **P<0.01.
H3R, histamine 3 receptor; si, small interfering; NC, negative
control; RAMH, R-(a)-methylhistamine; CPX, ciproxifan; BAX, BCL-2
associated X, apoptosis regulator; AR, androgen receptor. |
BCL-2 signaling pathways have been reported to serve
a key role in prostate cancer-cell apoptosis (16). The BCL-2 family contains BCL-2, BAX,
BAK and BCL-XL. It was demonstrated that knockdown of H3R
significantly decreased the level of BCL2 (anti-apoptosis gene)
expression (P<0.01), and increased the level of BAX
(apoptosis-associated gene) expression (P<0.05). It also
confirmed in specific H3R agonist and antagonist assay (Fig. 4).
H3R activates proliferation through
androgen receptor (AR) pathways
Numerous pathways have been demonstrated to be
involved in the development and progression of prostate cancer,
including AR, PI3K/AKT, MYC and PTEN (17–20). AR
has been demonstrated to accelerate the proliferation of prostate
cancer cells, therefore, in the present study, the expression of AR
was investigated following H3R-knockdown. Western blotting revealed
that the expression of AR was significantly reduced following
knockdown of H3R compared with si-NC-transfected cells (P<0.05).
This indicates that the expression of AR is associated with the
expression of H3R (Fig. 4G and I).
The H3R agonist and antagonist were used in parallel and CPX
reduced the expression of AR while RAMH increased the expression of
AR (Fig. 4).
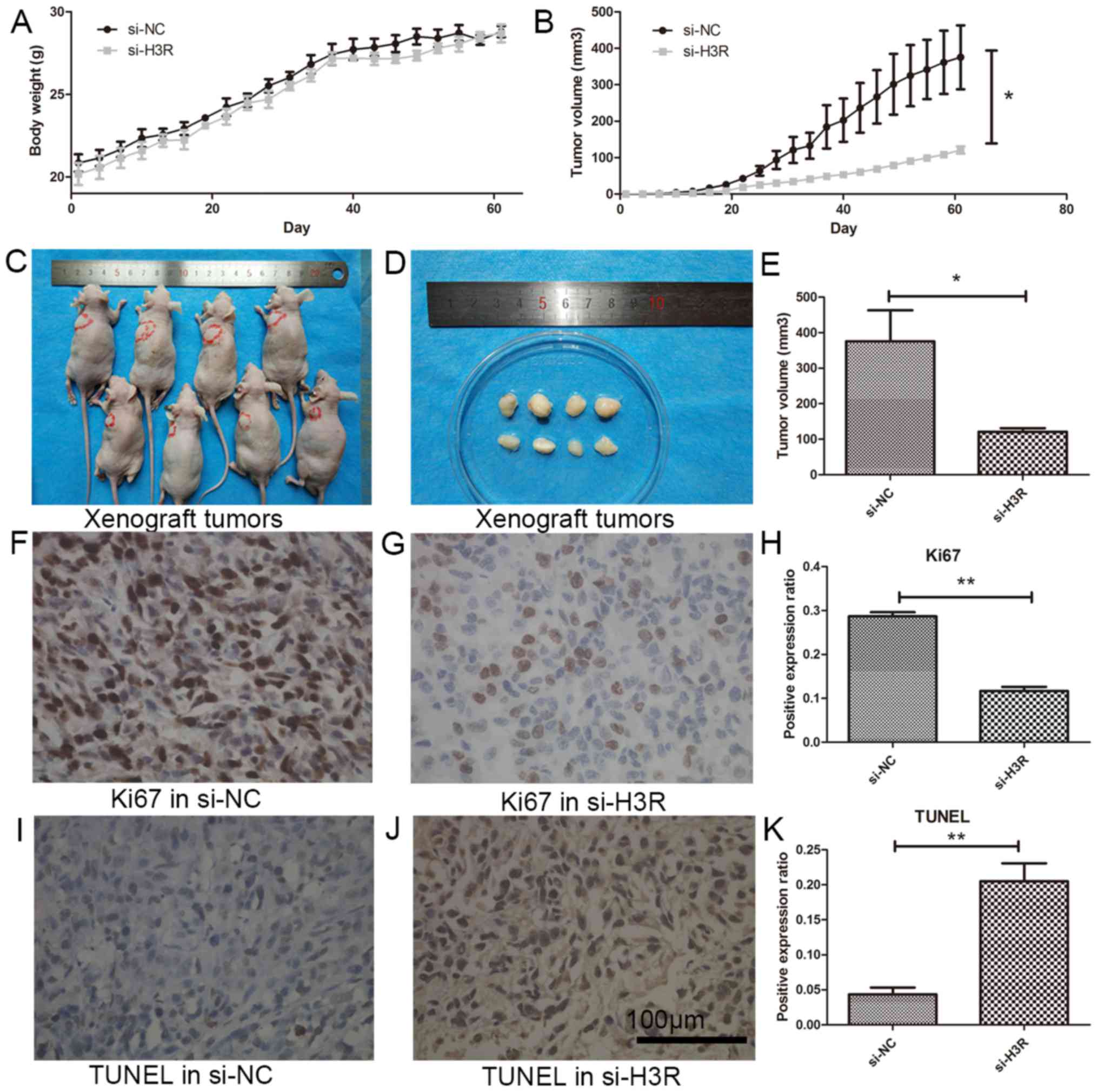
H3R knockdown suppresses prostate
cancer growth in vivo
To reveal the effect of H3R-knockdown in
vivo, a xenograft model was established in nude mouse using
si-H3R-transfected LNCaP cells (LNCaP siH3R) and negative control
(NC) transfected LNCaP cells (LNCaP si-NC). The length and width of
the tumor were measured twice a week. After 8 weeks, the tumors in
the LNCaP si-NC group grew to ~1 cm while tumors in the LNCaP
si-H3R group only grew to ~0.5 cm, and the difference was
significantly different (P<0.05) (Fig.
5A-E). Furthermore, immunohistochemistry revealed that the
expression of Ki67 (proliferation marker) was significantly in the
LNCaP si-H3R group than the LNCaP si-NC group (P<0.01) (Fig. 5F-H). The expression of TUNEL
(apoptosis marker) was significantly increased in the LNCaP si-H3R
group compared with the LNCaP si-NC group (P<0.01) (Fig. 5I-K).
Discussion
The expression of histamine receptor in prostate
cancer has been insufficiently studied. It has been demonstrated
that histamine receptor serves a key role in the nervous system
(21). Lin et al revealed that
inhibition of the expression of H3R could suppress the growth and
invasion of glioblastoma tumor (13).
Previous studies have demonstrated that histamine receptor may
influence the secretion of sex hormones (22). Therefore, the present study aimed to
elucidate the role of H3R in prostate cancer.
Preliminarily, the expression of 4 histamine
receptors (H1R-H4R) were investigated in prostate cancer (data are
not shown). Among the 4 histamine receptors, the expression of H3R
was the highest, and H3R-knockdown lead to the inhibition of the
LNCaP cell proliferation, migration and invasion. si-H3R also
induced the apoptosis of prostate cancer cells and inhibited the
expression of androgen receptor.
Androgen is essential to maintain male sex
characteristics, and has also been demonstrated to promote the
occurrence of prostate cancer (23).
It has been indicated that androgen requires androgen receptor to
increase the growth of prostate cancer (24), but androgen has the ability to promote
production of androgen receptor (25). Androgen receptor is overexpressed in
prostate cancer can harbor mutations which result in spliceosomes
(AR-Vs) (26), these mutations allow
prostate cancer cells to grow in very low concentrations of
androgen (27,28). Therefore, targeting androgen and
androgen receptor has been clinically researched in prostate cancer
with good results (29,30). In the present study, it was
demonstrated that when H3R was inhibited, the expression of
androgen receptor decreased. This indicates that the influence of
H3R on LNCaP cells may occur via androgen receptor.
The role of H3R in tumor-cell proliferation has been
reported in various types of cancer (31–33),
however, reports of H3R expression in prostate cancer are limited.
In the present study, it was demonstrated that H3R is overexpressed
in prostate cancer, that it can influence the proliferation,
migration and invasion of prostate cancer cells, as well as the
downstream androgen receptor. However, the mechanism by which H3R
regulates the expression of androgen receptor remains undetermined.
Whether H3R has a similar role in castration-resistant prostate
cancer or whether it functions in the switch of hormone-responsive
prostate cancer to castration-resistant prostate cancer, also
remains to be determined. The results of the present study broaden
our understanding of the underlying pathological mechanisms and may
aid in the discovery of novel treatment targets in prostate cancer.
Future studies should consider the potential of combined treatment
of H3R-inhibition treatment with other standard therapies,
including chemotherapy or androgen deprivation therapy. The
biological functions of H3R determines provide a foundation for
further investigation.
Acknowledgements
The authors thank Dr Guo-wen Lin (Fudan University
Shanghai Cancer Center) for providing the C4-2 cell lines.
Funding
The present study was supported by the Med-Tech
Crossdisciplinary Foundation of Shanghai Jiao Tong University
(grant no. YG2012MS55).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Ethics approval and consent to
participate
The present study was approved by Shanghai Jiao Tong
University Ethics Committee. Written informed consent was obtained
from all participants.
Authors' contributions
JC and XH contributed to all experiments.
Patient consent for publication
The authors declare that the patients have provided
written informed consent for the publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A6, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Graff RE, Meisner A, Ahearn TU, Fiorentino
M, Loda M, Giovannucci EL, Mucci LA and Pettersson A:
Pre-diagnostic circulating sex hormone levels and risk of prostate
cancer by ERG tumour protein expression. Br J Cancer. 114:939–944.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cornford P, Bellmunt J, Bolla M, Briers E,
De Santis M, Gross T, Henry AM, Joniau S, Lam TB, Mason MD, et al:
EAU-ESTRO-SIOG guidelines on prostate cancer. Part II: Treatment of
relapsing, metastatic and castration-resistant prostate cancer. Eur
Urol. 71:630–642. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
James ND, de Bono JS, Spears MR, Clarke
NW, Mason MD, Dearnaley DP, Ritchie AWS, Amos CL, Gilson C, Jones
RJ, et al: Abiraterone for prostate cancer not previously treated
with hormone therapy. N Engl J Med. 377:338–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sweeney CJ, Chen YH, Carducci M, Liu G,
Jarrard DF, Eisenberger M, Wong YN, Hahn N, Kohli M, Cooney MM, et
al: Chemohormonal therapy in metastatic hormone-sensitive prostate
cancer. N Engl J Med. 373:737–746. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sammon JD, Abdollah F, Reznor G, Pucheril
D, Choueiri TK, Hu JC, Kim SP, Schmid M, Sood A, Sun M, et al:
Patterns of declining use and the adverse effect of primary
androgen deprivation on all-cause mortality in elderly men with
prostate cancer. Eur Urol. 68:32–39. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Armitage JO and Sidner RD: Antitumour
effect of cimetidine. Lancet. 1:882–883. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang WT, Chen YH, Hsu JL, Leu WJ, Yu CC,
Chan SH, Ho YF, Hsu LC and Guh JH: Terfenadine induces
anti-proliferative and apoptotic activities in human
hormone-refractory prostate cancer through histamine
receptor-independent Mcl-1 cleavage and Bak up-regulation. Naunyn
Schmiedebergs Arch Pharmacol. 387:33–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blaya B, Nicolau-Galmés F, Jangi SM,
Ortega-Martínez I, Alonso-Tejerina E, Burgos-Bretones J,
Pérez-Yarza G, Asumendi A and Boyano MD: Histamine and histamine
receptor antagonists in cancer biology. Inflamm Allergy Drug
Targets. 9:146–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng Y, Xu M, Li X, Jia J, Fan K and Lai
G: Cimetidine suppresses lung tumor growth in mice through
proapoptosis of myeloid-derived suppressor cells. Mol Immunol.
54:74–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tanaka S, Sakaguchi M, Yoneyama H, Usami Y
and Harusawa S: Histamine H3 receptor antagonist OUP-186 attenuates
the proliferation of cultured human breast cancer cell lines.
Biochem Biophys Res Commun. 480:479–485. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin JJ, Zhao TZ, Cai WK, Yang YX, Sun C,
Zhang Z, Xu YQ, Chang T and Li ZY: Inhibition of histamine receptor
3 suppresses glioblastoma tumor growth, invasion, and
epithelial-to-mesenchymal transition. Oncotarget. 6:17107–17120.
2015.PubMed/NCBI
|
|
14
|
Donoso AO and Banzan AM: H1- and
H2-histamine receptor antagonists and induced release of prolactin
in male rats. Neuroendocrinology. 30:11–14. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin Y, Fukuchi J, Hiipakka RA, Kokontis JM
and Xiang J: Up-regulation of Bcl-2 is required for the progression
of prostate cancer cells from an androgen-dependent to an
androgen-independent growth stage. Cell Res. 17:531–536. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu C, Lou W, Armstrong C, Zhu Y, Evans CP
and Gao AC: Niclosamide suppresses cell migration and invasion in
enzalutamide resistant prostate cancer cells via Stat3-AR axis
inhibition. Prostate. 75:1341–1353. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barber AG, Castillo-Martin M, Bonal DM,
Jia AJ, Rybicki BA, Christiano AM and Cordon-Cardo C: PI3K/AKT
pathway regulates E-cadherin and Desmoglein 2 in aggressive
prostate cancer. Cancer Med. 4:1258–1271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Koh CM, Bieberich CJ, Dang CV, Nelson WG,
Yegnasubramanian S and De Marzo AM: MYC and prostate cancer. Genes
Cancer. 1:617–628. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gray IC, Stewart LM, Phillips SM, Hamilton
JA, Gray NE, Watson GJ, Spurr NK and Snary D: Mutation and
expression analysis of the putative prostate tumour-suppressor gene
PTEN. Br J Cancer. 78:1296–1300. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Green AJ, Gelfand JM, Cree BA, Bevan C,
Boscardin WJ, Mei F, Inman J, Arnow S, Devereux M, Abounasr A, et
al: Clemastine fumarate as a remyelinating therapy for multiple
sclerosis (ReBUILD): A randomised, controlled, double-blind,
crossover trial. Lancet. 390:2481–2489. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rossing MA, Scholes D, Cushing-Haugen KL
and Voigt LF: Cimetidine use and risk of prostate and breast
cancer. Cancer Epidemiol Biomarkers Prev. 9:319–323.
2000.PubMed/NCBI
|
|
23
|
Ragnarsson O, Johannsson G, Geterud K,
Lodding P and Dahlqvist P: Inadequate testosterone suppression
after medical and subsequent surgical castration in a patient with
prostate cancer. BMJ Case Rep. 2013:pii: bcr20130103952013.
View Article : Google Scholar
|
|
24
|
Wang Q, Li W, Zhang Y, Yuan X, Xu K, Yu J,
Chen Z, Beroukhim R, Wang H, Lupien M, et al: Androgen receptor
regulates a distinct transcription program in androgen-independent
prostate cancer. Cell. 138:245–256. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yeap BB, Krueger RG and Leedman PJ:
Differential posttranscriptional regulation of androgen receptor
gene expression by androgen in prostate and breast cancer cells.
Endocrinology. 140:3282–3291. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu X, Ledet E, Li D, Dotiwala A,
Steinberger A, Feibus A, Li J, Qi Y, Silberstein J, Lee B, et al: A
whole blood assay for AR-V7 and ARv567es in patients with prostate
cancer. J Urol. 196:1758–1763. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu Z, Chen S, Sowalsky AG, Voznesensky OS,
Mostaghel EA, Nelson PS, Cai C and Balk SP: Rapid induction of
androgen receptor splice variants by androgen deprivation in
prostate cancer. Clin Cancer Res. 20:1590–1600. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sharma NL, Massie CE, Ramos-Montoya A,
Zecchini V, Scott HE, Lamb AD, MacArthur S, Stark R, Warren AY,
Mills IG and Neal DE: The androgen receptor induces a distinct
transcriptional program in castration-resistant prostate cancer in
man. Cancer Cell. 23:35–47. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mills IG: Maintaining and reprogramming
genomic androgen receptor activity in prostate cancer. Nat Rev
Cancer. 14:187–198. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Z, Alyamani M, Li J, Rogacki K, Abazeed
M, Upadhyay SK, Balk SP, Taplin ME, Auchus RJ and Sharifi N:
Redirecting abiraterone metabolism to fine-tune prostate cancer
anti-androgen therapy. Nature. 533:547–551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pfanzagl B, Mechtcheriakova D,
Meshcheryakova A, Aberle SW, Pfragner R and Jensen-Jarolim E:
Activation of the ileal neuroendocrine tumor cell line P-STS by
acetylcholine is amplified by histamine: Role of H3R and H4R. Sci
Rep. 7:13132017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Davenas E, Rouleau A, Morisset S and
Arrang JM: Autoregulation of McA-RH7777 hepatoma cell proliferation
by histamine H3 receptors. J Pharmacol Exp Ther. 326:406–413. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Medina V, Croci M, Crescenti E, Mohamad N,
Sanchez-Jiménez F, Massari N, Nuñez M, Cricco G, Martin G, Bergoc R
and Rivera E: The role of histamine in human mammary
carcinogenesis: H3 and H4 receptors as potential therapeutic
targets for breast cancer treatment. Cancer Biol Ther. 7:28–35.
2008. View Article : Google Scholar : PubMed/NCBI
|