Introduction
Renal cell carcinoma (RCC) is the most common
urological cancer, representing ~90% of adult human kidney cancer
cases (1). RCC accounts for just
under 3% of all adult neoplasms. The incidence of RCC has increased
progressively in the last few decades. Even though the mortality
rate of RCC is sustained at a high level, it is a remediable
disease when metastasis does not occur. Clinically, it has been
shown that RCC is commonly resistant to chemotherapy and radiation
therapy, and that tumor resection remains the only definitive
treatment for curative therapy (2).
Nearly 55% of patients with RCC survive following curative
nephrectomy. However, 20–40% of RCC patients develop postoperative
metastasis (3). Thus far, no early
diagnostic biomarkers for RCC have been identified, resulting in
the late detection of the disease and a poor therapeutic effect.
Therefore, the identification of RCC biomarkers that can improve
the early diagnosis and prognosis of RCC patients is an important
focus of cancer therapy.
miRNAs are ~22-nucleotide, single-stranded, small
non-coding RNAs with expression at the post-transcriptional level
in diverse biological processes (4).
The differences in the miRNA expression profiles between RCC and
normal tissues show that miRNAs participate in the cancer genesis
of kidney carcinoma, suggesting roles as cancer suppressors and
oncogenes (5–7). Thus, miRNAs may be able to function as
prognostic and diagnostic biomarkers in RCC. Previous studies have
researched the role of miRNAs in RCC using miRNA expression
profiling and revealed that the miRNAs provided a novel
post-transcriptional mechanism for controlling the expression of
specific pathways and genes associated with RCC (8–10).
Revealing the biological functions of miRNAs and their roles in
kidney tumorigenesis may improve the early detection and effective
treatment of RCC.
Recently, mechanism-based and integrative profiling
research has revealed the functions of miRNAs in RCC. To a certain
extent, this research is beneficial to improve our understanding of
the molecular mechanisms of kidney cancer genesis. Previous studies
have reported the miRNA expression profiling and functions in RCC,
such as for miR-133b (9), miR-135a
(9), miR-205 (10), miR-372 (11), miR-204 (12) and miR-1258 (13). However, due to the difference in
sequencing platforms, sample selection and filter conditions, the
miRNA expression profiling displays inconsistent results between
different studies (5,6,8).
Thus, to minimize the limitations found in these
studies, a meta-analysis was performed in the present study using
the robust rank aggregation method (14), followed by the target prediction of
identified meta-signature miRNAs and pathway enrichment analysis,
to identify miRNA dysregulation in RCC and the pathways that may be
controlled by these miRNAs. The transcription factors of
meta-signature miRNA target genes were then identified. The present
meta-analysis is important for developing deeper insight into
miRNAs in the regulation of RCC development.
Materials and methods
Selection of studies and datasets
RCC miRNA expression profiling studies were found
via PubMed (www.ncbi.nlm.nih.gov/pubmed) using the search terms
‘microRNAs,’ ‘miRNAs,’ ‘renal cell carcinoma’ and their
combinations. The last search was performed on March 2017. Article
titles and abstracts were screened. The full texts of articles were
evaluated. Articles that were original experimental studies, were
published in English and analyzed the miRNA expression profiling
between human RCC samples and control samples were included.
Studies were excluded if they met the following selection criteria:
i) Non-English studies; ii) studies using only cell lines; iii)
studies of individual preselected candidate miRNAs or genes; iv)
case reports and review articles; and v) studies that did not
include non-cancerous control samples. Lists of statistically
significantly upregulated and downregulated miRNAs were extracted
from the publications. A total of 7 publicly available and
independent miRNA datasets of RCC were included in this analysis.
The occurrence of upregulated and downregulated miRNAs was
recorded, respectively. Differentially expressed miRNAs were
required to be present in at least 4 databases as a filtering
condition. All miRNA names were standardized on the basis of
miRBase version 21 (http://www.mirbase.org/).
Meta-analysis
The ranked lists of gene sets for each analysis
obtained from the 7 datasets were integrated for analysis. Based on
statistical test P-values performed with independent Student's
t-tests using SPSS 19.0 (IBM Corp., Armonk, NY, USA), where
P<0.05 was considered significant, the lists of extracted miRNAs
were prioritized by fold-change values. The robust rank aggregation
method was used to ensure that the extracted miRNAs could be ranked
reliably (14).
Prediction of meta-signature miRNA
target genes
The nucleotide sequences of meta-signature miRNAs
were retrieved using miRBase version 21 (http://www.mirbase.org/) (15) and organized into FASTA files. The
genes targeted by these miRNAs were then predicted using TargetScan
(16). Other parameters were default
parameters.
Pathway enrichment analysis
To predict the potential functions of target genes,
Gene Ontology (GO) analysis (www.geneontology.org/) was performed (17). The Kyoto Encyclopedia of Genes and
Genomes (KEGG) was used to identify molecular functions (18). Using the Database for Annotation,
Visualization and Integrated Discovery (DAVID) online tool, GO and
KEGG pathway enrichment analyses were performed for the
differentially expressed miRNA target genes (19). The filtering criterion was
P<0.05.
Transcription factor analysis of
meta-signature miRNA target genes
The transcription factor analysis of upregulated and
downregulated miRNA target genes was performed in Tfacts
(http://www.tfacts.org/). The indices, including
P-value, q-value, E-value and false discovery rate were used to
select reliable transcription factors. The quantitative values of
all the indices should be <0.05.
Results
Study selection
According to the selection criteria, 7 publicly
available and independent RCC miRNA expression profiling datasets
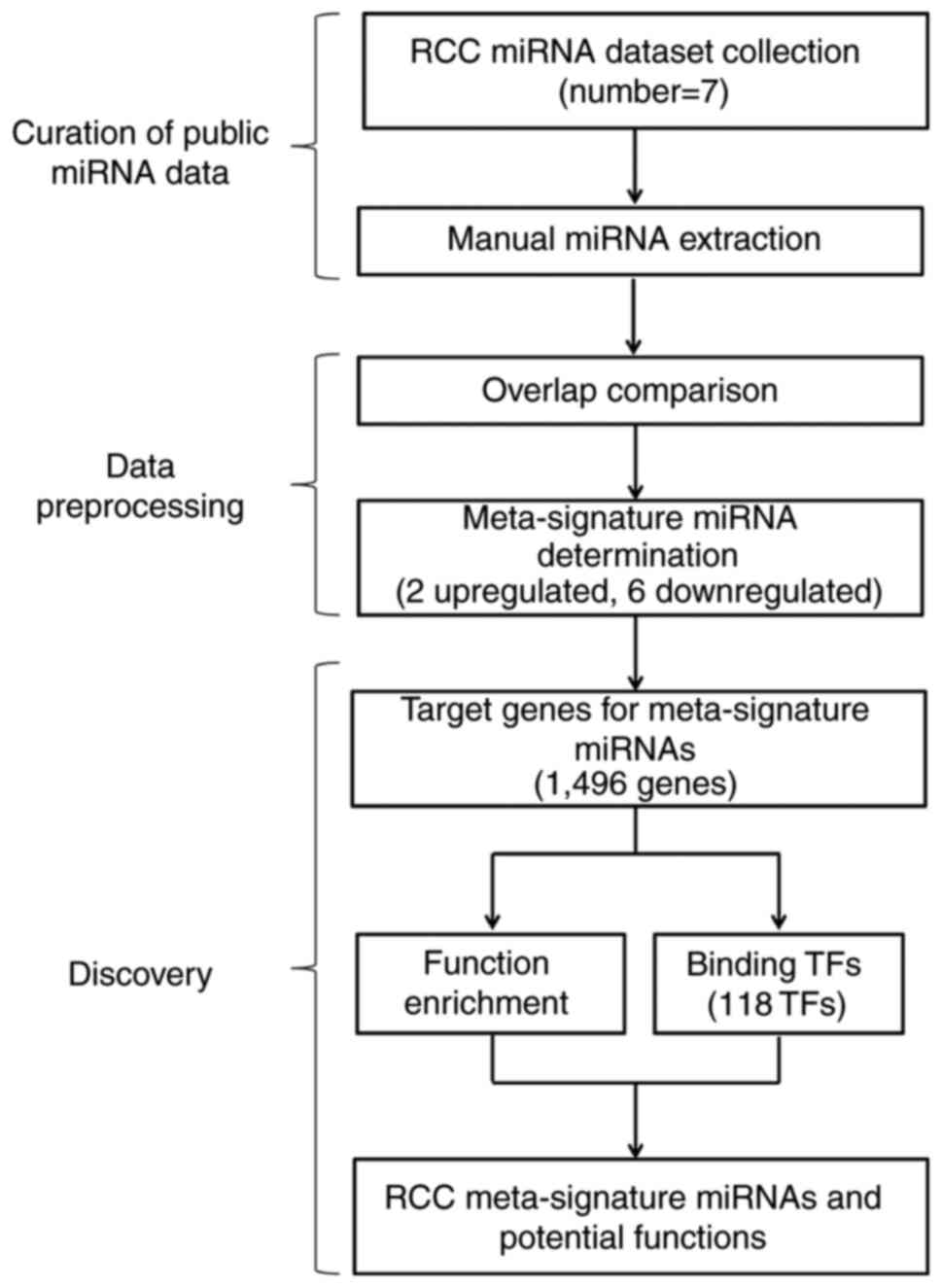
were collected in this analysis. The research strategy in this
study is shown in Fig. 1. These
datasets were performed with different populations and platforms.
The main characteristics of these studies and the acronyms by which
the studies are made reference to are presented in Table I. The 7 datasets were as follows: i)
MR (20); ii) XW (21); iii) SO (22); iv) CN (23); v) HH (13); vi) LW (24); and vii) FG (25). The 2 earliest studies were published
in 2007 and 2008, respectively, while the remaining 5 were
published between 2010 and 2012. Using the Scalable Vector Graphics
module of Perl (https://metacpan.org/pod/SVG) to analyze the
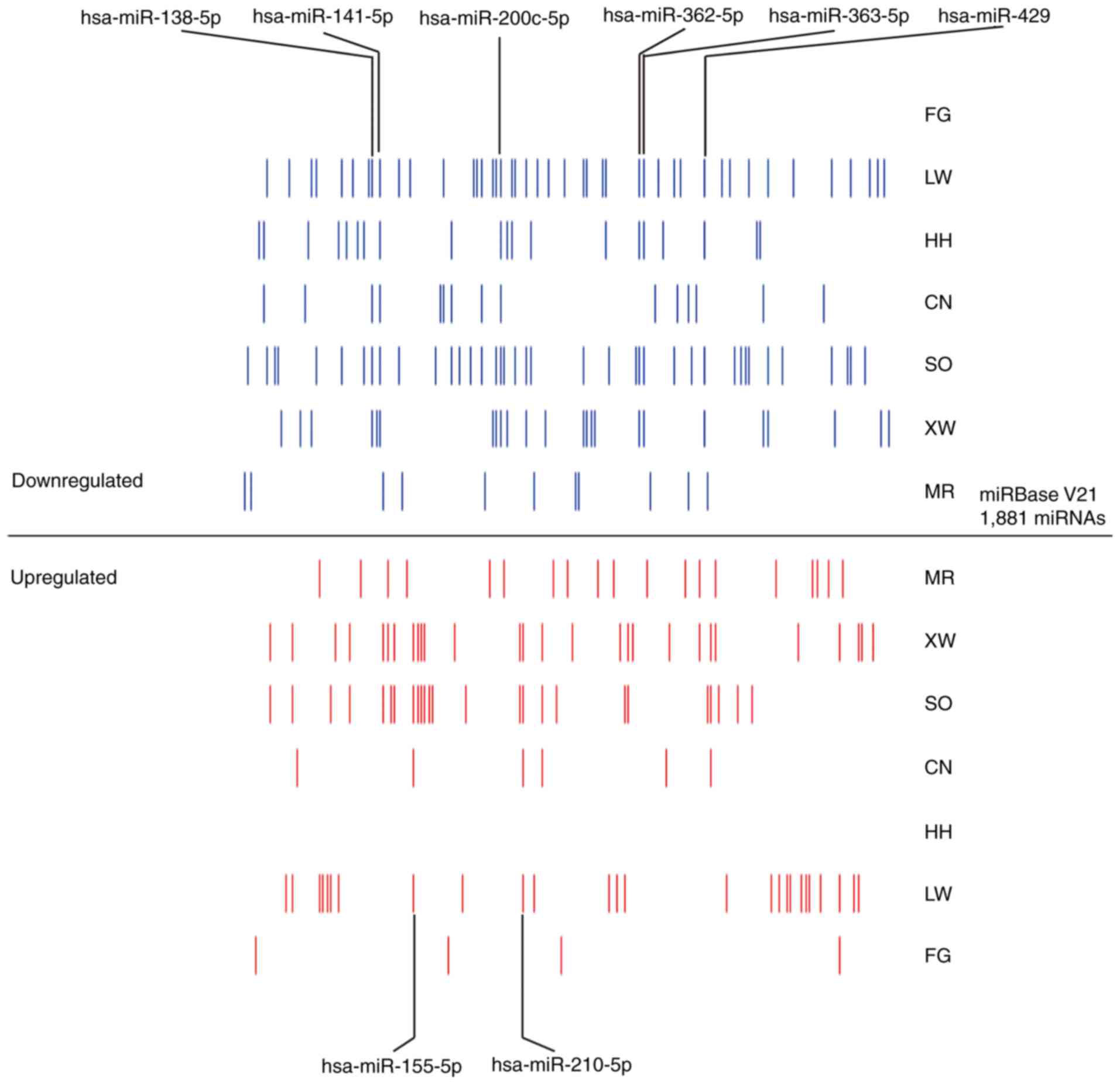
distribution of differentially expressed miRNAs, it was found that
the differentially expressed miRNAs were markedly different between
these 7 datasets (Fig. 2).
 | Table I.Characteristics of analyzed
datasets. |
Table I.
Characteristics of analyzed
datasets.
| Dataset first
author | Acronym | Samples | Assay/sequencing
type | method | (Refs.) |
|---|
| Redova et
al | MR | Blood serum of 15
RCC patients and 12 matched healthy controls | TaqMan Low Density
Arrays | RT-qPCR | (20) |
| Wu et
al | XW | A set of benign
kidney specimens (n=10) and a 28-sample ccRCC training cohort,
including localized (pT1; n=13) and metastatic (M1; n=15) tumor
samples |
Agilenta
Human miRNA Microarray V2 | RT-PCR | (21) |
| Osanto et
al | SO | 11 fresh frozen
ccRCC and adjacent non-tumoral renal cortex pairs | Small RNA
sequencing | Stem-loop PCR | (22) |
| Nakada et
al | CN | 26 individuals,
representing 16 CCCs, 4 ChCCs and 6 normal kidneys |
Agilenta
G4470A Human miRNA Microarray | RT-qPCR | (23) |
| Hidaka et
al | HH | 10 cancer tissues
and 5 adjacent non-cancerous tissues | TaqMan LDA Human
MicroRNA Panel v2.0 | RT-qPCR | (13) |
| Weng et
al | LW | Paired frozen and
FFPE benign kidney (n=3 each) and ccRCC (n=3) specimens | Small RNA
sequencing and MicroRNA Human Version 2 Microarray | RT-PCR | (24) |
| Gottardo et
al | FG | 27 kidney specimens
(20 carcinomas, 4 benign renal tumors and 3 normal parenchyma) | MicroRNA
oligonucleotide microchips | – | (25) |
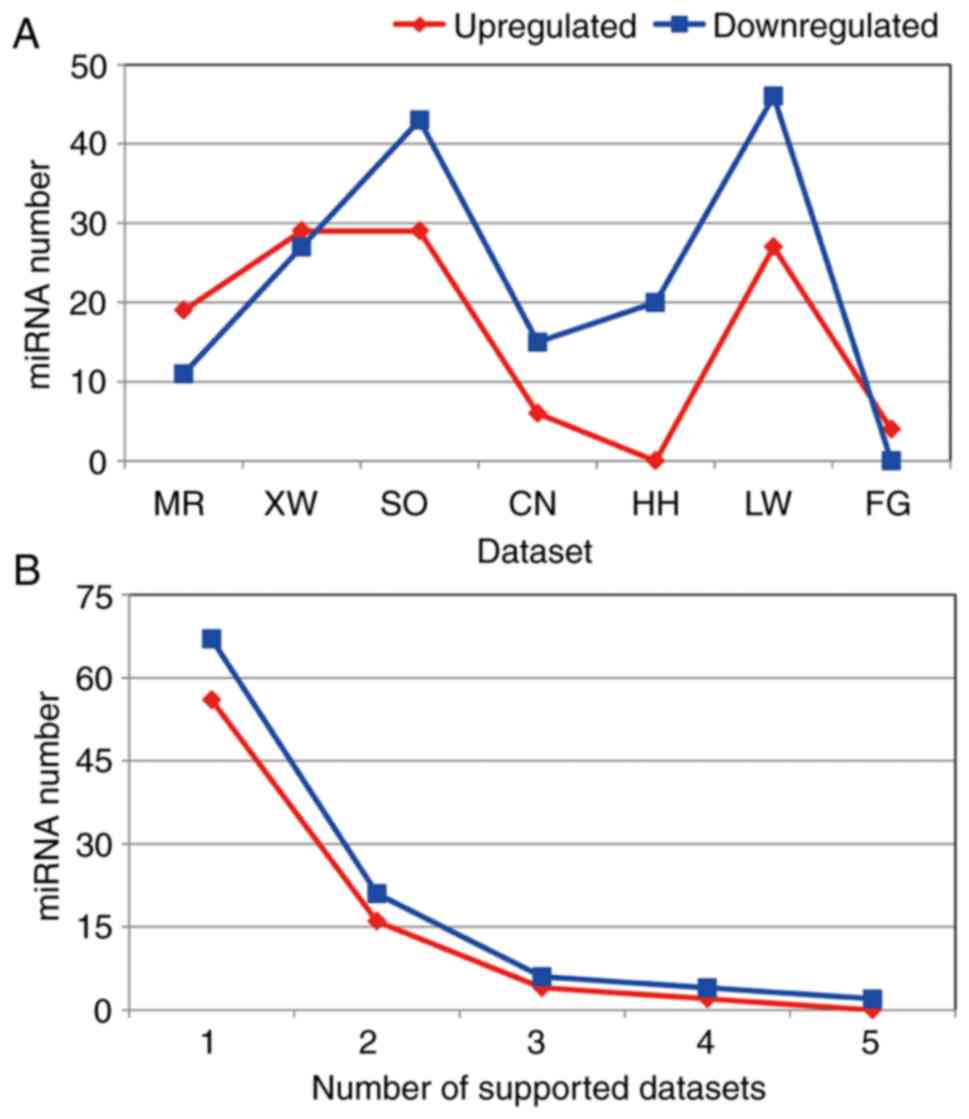
A total of 172 differentially expressed miRNAs were
revealed in the 7 miRNA expression profiling datasets. However, the
differentially expressed miRNA number for each dataset was clearly
different (Figs. 2 and 3). Although differences existed between each
miRNA dataset, the final lists of the deregulated miRNAs
corresponded. In total, 3 miRNA expression profiling datasets
included >50 differentially expressed miRNAs. The LW, SO and XW
datasets included 73, 72 and 56 differentially expressed miRNAs,
respectively. The XW and SO datasets contained the most upregulated
differentially expressed miRNAs, with 29 found. The LW dataset
contained 27 upregulated miRNAs, while the HH dataset did not
contain any. The LW and SO datasets included 46 and 43
downregulated miRNAs, respectively. However, the FG dataset did not
include any downregulated miRNAs.
RCC miRNA meta-signature
Using robust rank aggregation, a total of 8
significant meta-signature miRNAs were determined, including 2
upregulated and 6 downregulated miRNAs, from 7 RCC datasets
according to the permutation P-value (Fig. 2). All of the 8 meta-signature miRNAs
that attained statistical significance following Bonferroni's
correction were present in at least 4 datasets. The upregulated
miRNAs were hsa-miR-155-5p and hsa-miR-210-5p. The downregulated
miRNAs were hsa-miR-138-5p, hsa-miR-141-5p, hsa-miR-200c-5p,
hsa-miR-362-5p, hsa-miR-363-5p and hsa-miR-429.
The detailed location information of the 8
meta-signature miRNAs was extracted from the miRBase database
(Table II). The meta-signature miRNA
host genes were decentralized at different chromosomal locations,
with the exception of the hsa-miR-141-5p and hsa-miR-200c-5p genes,
and the hsa-miR-362-5p and hsa-miR-363-5p genes, which were located
on chr12 and chrX, respectively. hsa-miR-429, hsa-miR-210-5p,
hsa-miR-138-5p and hsa-miR-155-5p were included in chr1, chr11,
chr16 and chr21, respectively.
| Table II.Renal cell carcinoma meta-signature
miRNAs. |
Table II.
Renal cell carcinoma meta-signature
miRNAs.
| miRNA | Chromosome | Beginning | End | Strand | Support
datasets | Sequence |
|---|
| Upregulated |
|
|
|
|
|
|
|
hsa-miR-155-5p | chr21 | 25573983 | 25574005 | + | 4 |
UUAAUGCUAAUCGUGAUAGGGGU |
|
hsa-miR-210-5p | chr11 | 568150 | 568171 | – | 4 |
AGCCCCUGCCCACCGCACACUG |
| Downregulated |
|
|
|
|
|
|
|
hsa-miR-138-5p | chr16 | 56858527 | 56858549 | + | 4 |
AGCUGGUGUUGUGAAUCAGGCCG |
|
hsa-miR-141-5p | chr12 | 6964113 | 6964134 | + | 5 |
CAUCUUCCAGUACAGUGUUGGA |
|
hsa-miR-200c-5p | chr12 | 6963703 | 6963724 | + | 5 |
CGUCUUACCCAGCAGUGUUUGG |
|
hsa-miR-362-5p | chrX | 50008968 | 50008991 | + | 4 |
AAUCCUUGGAACCUAGGUGUGAGU |
|
hsa-miR-363-5p | chrX | 134169425 | 134169446 | – | 4 |
CGGGUGGAUCACGAUGCAAUUU |
|
hsa-miR-429 | chr1 | 1169055 | 1169076 | + | 4 |
UAAUACUGUCUGGUAAAACCGU |
Target prediction for meta-signature
miRNAs
TargetScan was used to gain predicted target genes
for the 8 meta-signature miRNAs. A total of 185 target genes were
obtained for hsa-miR-362-5p and >200 target genes for the other
7 meta-signature miRNAs.
Functional and pathway enrichment
analysis
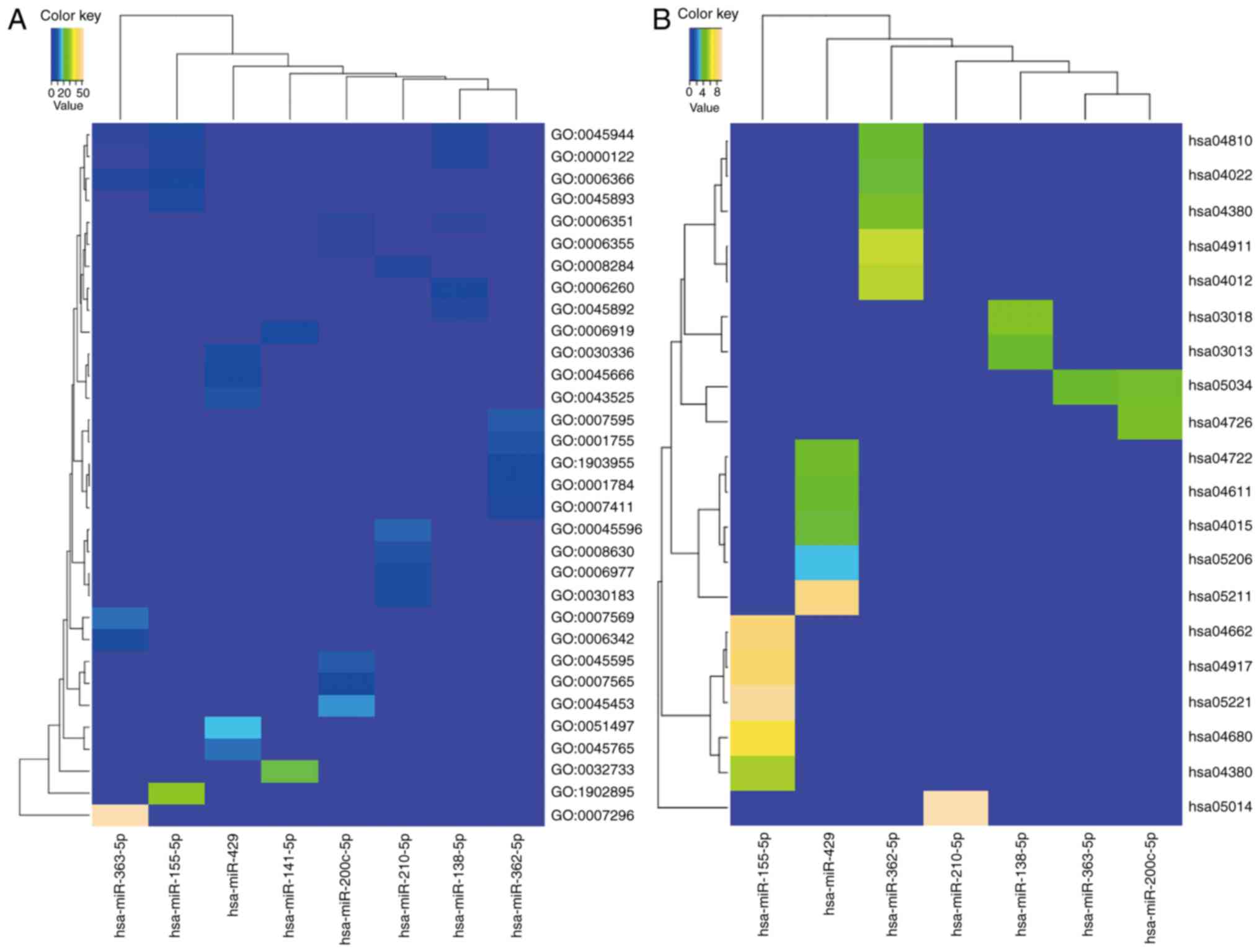
The functions of the target genes were analyzed
using the DAVID online tool, GO analysis and KEGG pathway
enrichment analyses. The enriched GO functions for the target genes
are presented in Fig. 4A and Table III. The predicted targets of
upregulated miRNAs were found to be significantly over-represented
in terms of the ‘transcriptional process’, ‘cell proliferation’ and
‘differentiation’. However, the downregulated miRNAs target genes
were mainly relevant to the ‘transcriptional process’, ‘cellular
immunity’, ‘neuronal differentiation’ and apoptosis. The enriched
KEGG outputs for several miRNA target sets mainly showed
associations with ‘cellular immunity’ (T cell receptor signaling
pathway, B cell receptor signaling pathway and Rap1 signaling
pathway), ‘neuronal development’ (serotonergic synapse and Axon
guidance), ‘cell mobility’ (regulation of actin cytoskeleton) and
‘cancer’ (renal cell carcinoma and microRNAs in cancer). The target
gene of hsa-miR-141-5p was not shown since it lacked KEGG
enrichment outputs.
| Table III.KEGG terms for the GO/KEGG terms of
meta-signature miRNAs. |
Table III.
KEGG terms for the GO/KEGG terms of
meta-signature miRNAs.
| Meta miRNA | Category | Term | Count | % | P-value | List total | Pop hits | Pop total | Fold
enrichment | FDR |
|---|
| hsa-miR-138-5p | KEGG_PATHWAY | hsa03013:RNA
transport | 7 | 0.022856 |
7.15×10−3 | 70 | 172 | 6910 | 4.02 | 8.04 |
| hsa-miR-138-5p | KEGG_PATHWAY | hsa03018:RNA
degradation | 4 | 0.013061 |
4.13×10−2 | 70 | 77 | 6910 | 5.13 | 38.86 |
| hsa-miR-155-5p | KEGG_PATHWAY | hsa04660:T cell
receptor signaling pathway | 9 | 0.028164 |
2.90×10−5 | 84 | 103 | 6910 | 7.19 | 0.04 |
| hsa-miR-155-5p | KEGG_PATHWAY | hsa04380:Osteoclast
differentiation | 9 | 0.028164 |
1.61×10−4 | 84 | 131 | 6910 | 5.65 | 0.20 |
| hsa-miR-155-5p | KEGG_PATHWAY | hsa04662:B cell
receptor signaling pathway | 7 | 0.021905 |
1.64×10−4 | 84 | 69 | 6910 | 8.35 | 0.20 |
| hsa-miR-155-5p | KEGG_PATHWAY | hsa04917:Prolactin
signaling pathway | 7 | 0.021905 |
1.92×10−4 | 84 | 71 | 6910 | 8.11 | 0.24 |
| hsa-miR-155-5p | KEGG_PATHWAY | hsa05221:Acute
myeloid leukemia | 6 | 0.018776 |
5.23×10−4 | 84 | 56 | 6910 | 8.81 | 0.64 |
|
hsa-miR-200c-5p | KEGG_PATHWAY |
hsa05034:Alcoholism | 6 | 0.02374 |
7.99×10−3 | 50 | 177 | 6910 | 4.68 | 8.86 |
|
hsa-miR-200c-5p | KEGG_PATHWAY |
hsa04726:Serotonergic synapse | 4 | 0.015827 |
4.36×10−2 | 50 | 111 | 6910 | 4.98 | 40.28 |
| hsa-miR-210-5p | KEGG_PATHWAY |
hsa05014:Amyotrophic lateral sclerosis
(ALS) | 4 | 0.016675 |
8.32×10−3 | 59 | 50 | 6910 | 9.37 | 9.32 |
| hsa-miR-362-5p | KEGG_PATHWAY | hsa04810:Regulation
of actin cytoskeleton | 8 | 0.026455 |
3.78×10−3 | 67 | 211 | 6910 | 3.91 | 4.48 |
| hsa-miR-362-5p | KEGG_PATHWAY | hsa04360:Axon
guidance | 6 | 0.019841 |
7.08×10−3 | 67 | 127 | 6910 | 4.87 | 8.22 |
| hsa-miR-362-5p | KEGG_PATHWAY | hsa04911:Insulin
secretion | 5 | 0.016534 |
8.62×10−3 | 67 | 85 | 6910 | 6.07 | 9.92 |
| hsa-miR-362-5p | KEGG_PATHWAY | hsa04012:ErbB
signaling pathway | 5 | 0.016534 |
9.34×10−3 | 67 | 87 | 6910 | 5.93 | 10.71 |
| hsa-miR-362-5p | KEGG_PATHWAY | hsa04022:cGMP-PKG
signaling pathway | 6 | 0.019841 |
2.07×10−2 | 67 | 166 | 6910 | 3.73 | 22.35 |
| hsa-miR-363-5p | KEGG_PATHWAY |
hsa05034:Alcoholism | 6 | 0.024136 |
1.71×10−2 | 60 | 177 | 6910 | 3.90 | 17.62 |
| hsa-miR-429 | KEGG_PATHWAY | hsa05211:Renal cell
carcinoma | 5 | 0.015331 |
2.64×10−3 | 63 | 65 | 6910 | 8.44 | 3.09 |
| hsa-miR-429 | KEGG_PATHWAY | hsa04015:Rap1
signaling pathway | 7 | 0.021463 |
1.09×10−2 | 63 | 210 | 6910 | 3.66 | 12.22 |
| hsa-miR-429 | KEGG_PATHWAY | hsa05206:MicroRNAs
in cancer | 8 | 0.024529 |
1.33×10−2 | 63 | 285 | 6910 | 3.08 | 14.67 |
| hsa-miR-429 | KEGG_PATHWAY |
hsa04722:Neurotrophin signaling
pathway | 5 | 0.015331 |
2.23×10−2 | 63 | 120 | 6910 | 4.57 | 23.41 |
| hsa-miR-429 | KEGG_PATHWAY | hsa04611:Platelet
activation | 5 | 0.015331 |
2.88×10−2 | 63 | 130 | 6910 | 4.22 | 29.28 |
Transcription factor analysis of
meta-signature miRNA target genes
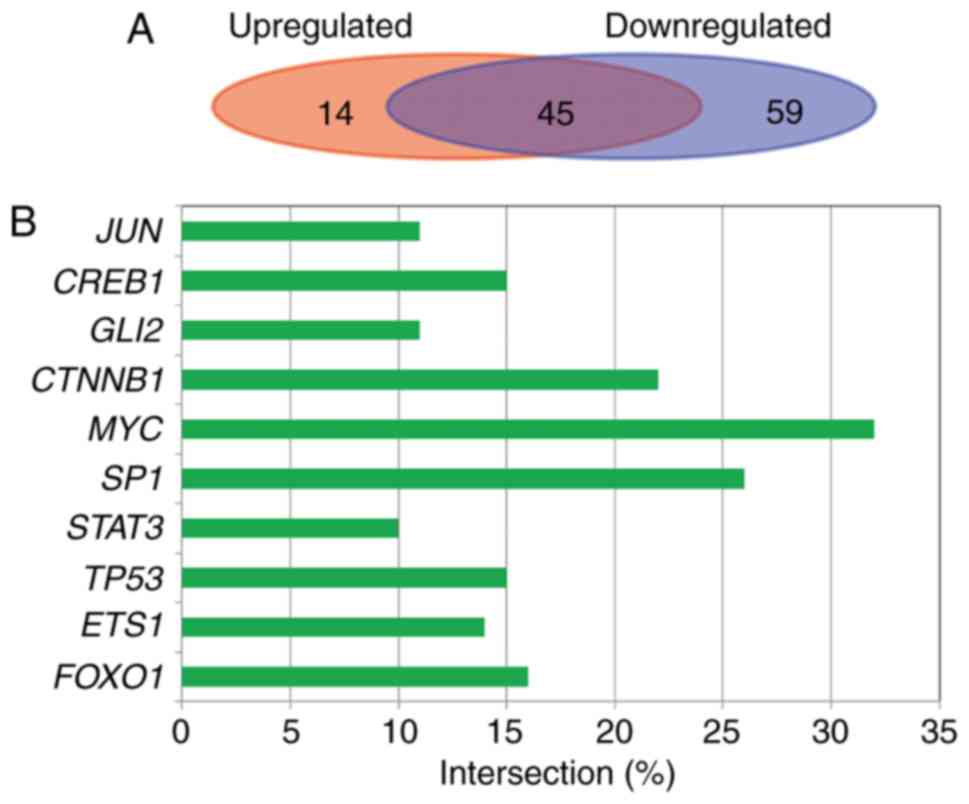
There were 106 interactions between 59 transcription
factors and 41 target genes for upregulated miRNAs, and 326
interactions between 104 transcription factors and 117 target genes
for downregulated miRNAs. In total, 45 transcription factors were
common to the upregulated and downregulated miRNAs target genes in
118 transcriptional factors (Fig.
5A). The anharmonic ratio of mainly transcription factors for
target genes is shown in Fig. 5B
(E-value, <0.05). Transcription factor MYC had the highest
anharmonic ratio and reached 32%.
Discussion
Numerous previous studies have reported associations
between miRNA expression and RCC. A number of these studies showed
that the differentially expressed miRNAs determined by the
different studies did not observe a consistent result in disease
samples compared with control samples. Differences among
technological platforms, sample size and etiological factors
attributed to the lack of uniformity (26). It is better to analyze individual
datasets and then analyze the resulting miRNA lists aggregately to
overcome these limitations. Therefore, such a comprehensive
evaluation of miRNA expression profiles was performed in kidney
cancer for the present study. An integrated analysis was performed
to identify differentially expressed miRNAs in 7 different
profiling datasets for RCC. Only the datasets that were generated
from miRNA microarrays or sequencing and further validated by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) or RT-PCR were selected in this study. Using the robust
rank aggregation method, 2 upregulated and 6 downregulated
meta-signature miRNAs were determined in at least 4 studies. The
location of the 8 meta-signature miRNAs on the chromosomes and
their nucleotide sequences are shown in Table II. The majority of them have
previously been determined as tumor suppressors and oncogenes via
oncogenic pathways and are known to be functionally important in
kidney carcinogenesis (27).
Therefore, the present results may highlight the potential of miRNA
to improve clinical prediction for RCC patients.
Numerous previous studies have suggested that a
number of miRNAs serve noticeable roles in kidney cancer
development. For example, Chen et al (10) showed that miRNA-205 may be a candidate
RCC suppressor through targeting zinc finger E-box-binding homeobox
2 (ZEB2). hsa-miR-155-5p and hsa-miR-210-5p were reported to be
upregulated in the RCC samples (22),
which was also observed in the present study. By contrast,
hsa-miR-138-5p and hsa-miR-141-5p were identified for the first
time in the present study. miR-205 was markedly upregulated in the
non-tumor tissues compared with that in the RCC samples and cell
lines. Overexpression of miR-205 suppressed ACHN cell
proliferation, migration and invasion, and induced cell apoptosis
(10). A study conducted by Zhou
et al (9) reported that
miRNA-133b and miRNA-135a suppressed cell proliferation and induced
apoptosis through the tyrosine-protein kinase JAK2/signal
transducer and activator of transcription 3 signaling pathway in
RCC. Huang et al (11)
demonstrated that miRNA-372 was upregulated in control normal
samples compared with that in RCC tissues and cell lines. miRNA-372
overexpression inhibited proliferation and invasion via targeting
insulin-like growth factor 2 in RCC cell lines (11). It is noteworthy that the majority of
RCC meta-signature miRNAs in the present study have also been
demonstrated to serve functionally important roles in kidney cancer
genesis. In addition, the targeted gene enrichment analysis
suggested effects on several pathways that participate in
cancerogenesis, including the rap1 signaling pathway, renal cell
carcinoma and microRNAs in cancer (Fig.
4). The majority of the 8 meta-signature miRNAs in the present
study were identified to be associated with RCC development.
Therefore, these miRNAs may be potential candidates for the
development of early detection and diagnosis methods for RCC. The
meta-analysis showed that high levels of miR-155-5p and miR-210-5p
were associated with RCC. Gao et al (28) reported that miR-155 overexpression
decreased transcription factor E2F2 (E2F2) expression, while
inhibition elevated E2F2 expression in RCC cell lines, and that
miR-155 may function as a cancer-promoting miRNA through targeting
E2F2. Nakada et al (29)
suggested that the upregulation of miR-210 was associated with
hypoxia-inducible factor 1α accumulation in response to hypoxic
conditions in renal cancer (30).
miR-210 was also associated with a number of biological processes,
including mitochondrial metabolism, angiogenesis, cell cycle
regulation and apoptosis (30).
Furthermore, miR-141-5p, miR-200c-5p and miR-429 are members of the
miR-200 family (31). Previous
studies have reported that ZEB2, which is a transcriptional
repressor for CDH1/E-cadherin, is a hypothetical target of miR-200c
and miR-141 in a variety of cancer types (31). miR-141 and miR-200c overexpression
decreased ZEB2 expression and increased E-cadherin expression in
lung cancer cells and renal carcinoma cell lines (32,33). A
study conducted by Liang et al (34) showed that miR-138 was able to induce
SN-12 cell senescence by increasing the expression of tumor protein
p16 and targeting histone-lysine N-methyltransferase EZH2 (34). In addition, miR-429 was demonstrated
to inhibit cell proliferation, migration and invasion of RCC
through targeting transcription factor Sp1 (35). miR-363-3p transfection caused a
decrease in cyclic AMP-responsive element-binding protein 1 (CREB1)
expression and suppressed cell proliferation, migration and
apoptosis reduction in human RCC through targeting CREB1 (36).
To the best of our knowledge, only a few
meta-analyses of miRNA profiling investigations have been performed
specifically for RCC. Tang and Xu (37) identified a statistically significant
miRNA meta-signature of two upregulated (hsa-miR-21 and
hsa-miR-210) and three downregulated (hsa-miR-141, hsa-miR-200c and
hsa-miR-429) miRNAs in patients with RCC. The serum levels of
hsa-miR-193a-3p, hsa-miR-362 and hsa-miR-572 in patients with RCC
were significantly increased, whereas the levels of hsa-miR-28-5p
and hsa-miR-378 were markedly decreased (8). In addition, White et al (6) reported that hsa-miR-122, hsa-miR-155 and
hsa-miR-210 had the highest overexpression, and hsa-miR-200c,
hsa-miR-335 and hsa-miR-218 had the most downregulation in patients
with RCC. In present study, a meta-analysis approach was used to
integrate multitudinous and independent miRNA expression profiling
datasets for RCC. A meta-signature, composed of 8 differentially
expressed miRNAs, including 2 upregulated miRNAs, hsa-miR-155-5p
and hsa-miR-210-5p, and 6 downregulated miRNAs, hsa-miR-138-5p,
hsa-miR-141-5p, hsa-miR-200c-5p, hsa-miR-362-5p, hsa-miR-363-5p and
hsa-miR-429, from 7 different and independent datasets was
determined. The subsequent target prediction of meta-signature
miRNAs and pathway enrichment analysis identified genes and
pathways that may be regulated by these meta-signature miRNAs.
Furthermore, the transcription factors of the meta-signature miRNA
target genes were identified. One potential limitation of the
current study is that the miRNA dataset from blood serum samples
was not included, which may have different miRNA expression
compared with that of the tumor tissues. The present analysis may
be useful in the research into predictive markers for the early
detection of RCC and assist in obtaining a promising treatment in
anticancer therapy. However, further in vivo validation
studies will be required.
Acknowledgements
Not applicable.
Funding
This study was supported by a grant from the Project
of Clinical Research Funds of Zhejiang Medical Association (no.
2011ZYC-A62).
Availability of data and materials
The datasets used an/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GY, RW and FW conceived and designed this study. GY,
RW, MX, XF, QEH and CZ performed the experiments. GY, RW and FW
wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Banumathy G and Cairns P: Signaling
pathways in renal cell carcinoma. Cancer Biol Ther. 10:658–664.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pantuck AJ, Zisman A and Belldegrun AS:
The changing natural history of renal cell carcinoma. J Urol.
166:1611–1623. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel D: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yi Z, Fu Y, Zhao S, Zhang X and Ma C:
Differential expression of miRNA patterns in renal cell carcinoma
and nontumorous tissues. J Cancer Res Clin Oncol. 136:855–862.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
White NM, Bao TT, Grigull J, Youssef YM,
Girgis A, Diamandis M, Fatoohi E, Metias M, Honey RJ, Stewart R, et
al: miRNA profiling for clear cell renal cell carcinoma: Biomarker
discovery and identification of potential controls and consequences
of miRNA dysregulation. J Urol. 186:1077–1083. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meng X, Chen X, Chen D, Bing Y and Huang
Z: FoxO1: A novel insight into its molecular mechanisms in the
regulation of skeletal muscle differentiation and fiber type
specification. Oncotarget. 8:10662–10674. 2017.PubMed/NCBI
|
|
8
|
Wang C, Hu J, Lu M, Gu H, Zhou X, Chen X,
Zen K, Zhang CY, Zhang T, Ge J, et al: A panel of five serum miRNAs
as a potential diagnostic tool for early-stage renal cell
carcinoma. Sci Rep. 5:76102015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou W, Bi X, Gao G and Sun L: miRNA-133b
and miRNA-135a induce apoptosis via the JAK2/STAT3 signaling
pathway in human renal carcinoma cells. Biomed Pharmacother.
84:722–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Z, Tang ZY, He Y, Liu LF, Li DJ and
Chen X: miRNA-205 is a candidate tumor suppressor that targets ZEB2
in renal cell carcinoma. Oncol Res Treat. 37:6582014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang X, Huang M, Kong L and Li Y: miR-372
suppresses tumour proliferation and invasion by targeting IGF2BP1
in renal cell carcinoma. Cell Prolif. 48:593–599. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiong F, Liu K, Zhang F, Sha K, Wang X,
Guo X and Huang N: miR-204 inhibits the proliferation and invasion
of renal cell carcinoma by inhibiting RAB22A expression. Oncol Rep.
35:3000–3008. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hidaka H, Seki N, Yoshino H, Yamasaki T,
Yamada Y, Nohata N, Fuse M, Nakagawa M and Enokida H: Tumor
suppressive microRNA-1285 regulates novel molecular targets:
Aberrant expression and functional significance in renal cell
carcinoma. Oncotarget. 3:44–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kolde R, Laur S, Adler P and Vilo J:
Robust rank aggregation for gene list integration and
meta-analysis. Bioinformatics. 28:573–580. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Griffiths-Jones S, Saini HK, van Dongen S
and Enright AJ: miRBase: Tools for microRNA genomics. Nucleic Acids
Res. 36:D154–D158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are MicroRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tweedie S, Ashburner M, Falls K, Leyland
P, McQuilton P, Marygold S, Millburn G, Osumi-Sutherland D,
Schroeder A, Seal R and Zhang H: FlyBase Consortium: FlyBase:
Enhancing Drosophila gene ontology annotations. Nucleic Acids Res.
37:D555–D559. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rédei GP: Kyoto Encyclopedia of Genes and
Genomes. John Wiley and Sons, Inc.; Hoboken, NJ: 2012
|
|
19
|
Huang DW, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Redova M, Poprach A, Nekvindova J, Iliev
R, Radova L, Lakomy R, Svoboda M, Vyzula R and Slaby O: Circulating
miR-378 and miR-451 in serum are potential biomarkers for renal
cell carcinoma. J Transl Med. 10:552012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu X, Weng L, Li X, Guo C, Pal SK, Jin JM,
Li Y, Nelson RA, Mu B, Onami SH, et al: Identification of a
4-microRNA signature for clear cell renal cell carcinoma metastasis
and prognosis. PLoS One. 7:e356612012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osanto S, Qin Y, Buermans HP, Berkers J,
Lerut E, Goeman JJ and van Poppel H: Genome-wide microRNA
expression analysis of clear cell renal cell carcinoma by next
generation deep sequencing. PLoS One. 7:e382982012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakada C, Matsuura K, Tsukamoto Y,
Tanigawa M, Yoshimoto T, Narimatsu T, Nguyen LT, Hijiya N, Uchida
T, Sato F, et al: Genome-wide microRNA expression profiling in
renal cell carcinoma: significant down-regulation of miR-141 and
miR-200c. J Pathol. 216:418–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weng L, Wu X, Gao H, Mu B, Li X, Wang JH,
Guo C, Jin JM, Chen Z, Covarrubias M, et al: MicroRNA profiling of
clear cell renal cell carcinoma by whole-genome small RNA deep
sequencing of paired frozen and formalin-fixed, paraffin-embedded
tissue specimens. J Pathol. 222:41–51. 2010.PubMed/NCBI
|
|
25
|
Gottardo F, Liu CG, Ferracin M, Calin GA,
Fassan M, Bassi P, Sevignani C, Byrne D, Negrini M, Pagano F, et
al: Micro-RNA profiling in kidney and bladder cancers. Urol Oncol.
25:387–392. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Diaz G, Melis M, Tice A, Kleiner DE,
Mishra L, Zamboni F and Farci P: Identification of microRNAs
specifically expressed in hepatitis C virus-associated
hepatocellular carcinoma. Int J Cancer. 133:816–824. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ortholan C, Puissegur MP, Ilie M, Barbry
P, Mari B and Hofman P: MicroRNAs and lung cancer: New oncogenes
and tumor suppressors, new prognostic factors and potential
therapeutic targets. Curr Med Chem. 16:1047–1061. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao Y, Ma X, Yao Y, Li H, Fan Y, Zhang Y,
Zhao C, Wang L, Ma M, Lei Z and Zhang X: miR-155 regulates the
proliferation and invasion of clear cell renal cell carcinoma cells
by targeting E2F2. Oncotarget. 7:20324–20337. 2016.PubMed/NCBI
|
|
29
|
Nakada C, Tsukamoto Y, Matsuura K, Nguyen
TL, Hijiya N, Uchida T, Sato F, Mimata H, Seto M and Moriyama M:
Overexpression of miR-210, a downstream target of HIF1α, causes
centrosome amplification in renal carcinoma cells. J Pathol.
224:280–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu TY, Zhang H, Du SM, Li J and Wen XH:
Expression of microRNA-210 in tissue and serum of renal carcinoma
patients and its effect on renal carcinoma cell proliferation,
apoptosis, and invasion. Genet Mol Res. 15:150177462016.PubMed/NCBI
|
|
31
|
Song F, Yang D, Liu B, Guo Y, Zheng H, Li
L, Wang T, Yu J, Zhao Y, Niu R, et al: Integrated microRNA network
analyses identify a poor-prognosis subtype of gastric cancer
characterized by the miR-200 family. Clin Cancer Res. 20:878–889.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dhayat SA, Mardin WA, Köhler G, Bahde R,
Vowinkel T, Wolters H, Senninger N, Haier J and Mees ST: The
microRNA-200 family-a potential diagnostic marker in hepatocellular
carcinoma? J Surg Oncol. 110:430–438. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hurteau GJ, Carlson JA, Spivack SD and
Brock GJ: Overexpression of the microRNA hsa-miR-200c leads to
reduced expression of transcription factor 8 and increased
expression of E-cadherin. Cancer Res. 67:7972–7976. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liang J, Zhang Y, Jiang G, Liu Z, Xiang W,
Chen X, Chen Z and Zhao J: MiR-138 induces renal carcinoma cell
senescence by targeting EZH2 and is downregulated in human clear
cell renal cell carcinoma. Oncol Res. 21:83–91. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu D, Niu X, Pan H, Zhou Y, Zhang Z, Qu P
and Zhou J: Tumor-suppressing effects of microRNA-429 in human
renal cell carcinoma via the downregulation of Sp1. Oncol Lett.
12:2906–2911. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Y, Chen D, Li Y, Jin L, Liu J, Su Z, Qi
Z, Shi M, Jiang Z, Ni L, et al: Oncogenic cAMP responsive element
binding protein 1 is overexpressed upon loss of tumor suppressive
miR-10b-5p and miR-363-3p in renal cancer. Oncol Rep. 35:1967–1978.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang K and Xu H: Prognostic value of
meta-signature miRNAs in renal cell carcinoma: An integrated miRNA
expression profiling analysis. Sci Rep. 5:102722015. View Article : Google Scholar : PubMed/NCBI
|