Introduction
Lung cancer is a leading cause of cancer-associated
mortality worldwide (1,2). Among different types of lung cancer,
non-small cell lung cancer (NSCLC) accounts for >80% of
mortalities (2,3). Although several therapeutic strategies
have been developed, including surgery, chemotherapy and radiation
therapy, the prognosis of NSCLC patients remains poor, and the
5-year survival rate is <15% (4).
NSCLC is difficult to treat due to malignant migration and
metastasis, and the worsening physical conditions seriously affect
the quality of life of patients (5,6).
Therefore, understanding the underlying molecular mechanism of
migration and metastasis in NSCLC may aid the development of more
effective therapeutic strategies.
Prostaglandin E2 (PGE2) is the predominant product
of arachidonic acid metabolism by cyclooxygenase-2 (COX-2). A
number of studies have suggested that PGE2 serves functions in
various biological processes, including inflammation, cell
survival, migration and invasion (7–9). PGE2 has
been demonstrated to bind to E prostanoid receptors on the surface
of the cell membrane and thus regulate signaling pathways,
including cAMP-PKA (10), PI3K/Akt
and Ras/ERK (11), to exert their
function. E-prostanoid 3 (Ep3) is a G protein-coupled receptor that
serves an essential function in various diseases, including
inflammation, hypertension and cancer (12–15). Amano
et al (16) suggested that Ep3
signaling on endothelial cells is essential for the matrix
metalloproteinase (MMP)-9 upregulation that subsequently enhances
tumor metastasis and angiogenesis. Fang et al (17) reported that Actinidia chinensis
Planch root extract (acRoots) inhibits hepatocellular carcinoma
cell invasion and metastasis via the inhibition of EP3 expression,
resulting in decreased activation of vascular endothelial growth
factor (VEGF), epidermal growth factor receptor, MMP2 and MMP9.
Jiang et al (18) demonstrated
that the inhibition of cell growth and induction of apoptosis by
retinoic acid chalcone in colon cancer is mediated by inhibition of
COX-2 expression, and subsequent inhibition of PGE2 and PGE2
receptors. The aforementioned studies suggest that the abnormal
expression of Ep3 serves an important function in a number of
cancer cells, and is associated with cell growth and metastasis.
Yano et al (19) demonstrated
that the expression of Ep3 may be a factor in the PGE2-mediated
activation of the Ras signaling pathway in A549 cells. Yamaki et
al (20) suggested that
PGE2-dependent activation of Src signaling via Ep3 serves an
important function in growth of A549 cells. These results suggest
that Ep3 is involved in PGE2-mediated cellular processes in A549
cells. However, the functional effects and underlying molecular
mechanisms of Ep3 in the development of NSCLC remain to be
elucidated.
A number of studies have suggested that the
regulation of Ep3 in cancer cells may be mediated by numerous
signaling pathways, including extracellular signal-related kinase,
phosphoinositide 3-kinase/protein kinase B and nuclear factor
κ-light-chain-enhancer of activated B cells signaling (21,22). It
has been reported that transforming growth factor (TGF)-β signaling
serves a function in numerous types of cancer by regulating a
variety of cellular events, including proliferation, migration and
apoptosis (23,24). By binding to its receptor, TGF-β is
able to activate Smad2 and Smad3, and initiate their translocation
to the nucleus by forming a trimer with Smad4, to regulate the
expression of TGF-β dependent genes (25,26).
Several investigations have demonstrated that the activation of
TGF-β, and the subsequent phosphorylation of Smad2 and Smad3
promote the invasion and migration of lung cancer cells (27,28),
suggesting that TGF-β/Smad signaling is involved in the regulation
of lung cancer cells. A previous study reported that the inhibition
of Ep3 attenuates pulmonary hypertension through suppression of
Rho/TGF-β1 signaling (29),
suggesting that the regulation of Ep3 may be associated with TGF-β
signaling. Thus, establishing whether TGF-β signaling is involved
in the effects of Ep3 in lung cancer cells is of interest.
In the present study, the expression of Ep3 in NSCLC
tissues and A549 cells was evaluated. The effects of Ep3 on the
cell viability, migration, invasion and apoptosis of A549 cells
were investigated, and the underlying molecular mechanisms of each
were explored. It was hypothesized that the inhibition of Ep3 may
suppress the cell viability, migration and invasion, and promote
cell apoptosis of A549 cells.
Materials and methods
Tissue specimens
A total of 17 NSCLC tissues and corresponding
adjacent normal lung tissues were obtained from patients from
Zhoukou Central Hospital (Zhoukou, China) who underwent curative
resection for NSCLC between August 2015 and October 2016. Among
them, twelve were males, and five were females. Their ages were
between 47 and 68 years, and the mean age was 61 years. All of the
patients provided written informed consent, and the present study
was approved by the Ethics Committee of Zhoukou Central
Hospital.
Cell culture and L-798106
treatment
The A549 NSCLC cell line and HPAEpiC human alveolar
epithelial cell line were purchased from the American Type Culture
Collection (Manassas, VA, USA). The cells were cultured in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and maintained at 37°C in a humidified atmosphere
with 5% CO2. A549 cells were treated with 1 µM Ep3
antagonist L-798106 (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
for 48 h in the following experiments.
Cell transfection
Ep3 siRNA (si-Ep3) and negative control siRNA
(si-control) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). The TGF-β cDNA was cloned into pcDNA 3.1 vector
(Invitrogen; Thermo Fisher Scientific, Inc.). The constructs were
verified by DNA sequencing. The construction and verification of
the pcDNA 3.1-TGF-β plasmids was performed by Generay Biotech Co.,
Ltd (Shanghai, China). Cells were seeded into 6-well, 24-well or
96-well plates according to experimental requirements. When the
cells reached 70–80% confluence, Ep3 siRNA, negative control siRNA
or plasmids were transfected into cells using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturers' protocol. Then, 6 h after transfection, the medium
was changed to RPMI-1640 medium supplemented with 10% FBS.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total mRNA was extracted from tissues and cells
using TRIzol® (Thermo Fisher Scientific, Inc.) according
to the manufacturers' protocol. mRNA was reverse transcribed to
cDNA using a PrimeScript RT reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China) according to the manufacturers' protocol. The
mRNA levels of Ep3 and β-actin were measured using SYBR Premix
Real-Time PCR reagent (Takara Biotechnology Co., Ltd.) according to
the manufacturer's protocol. The primers used in the present study
were as follows: Ep3 forward, 5′-TCCTTCCTAATCGCCGTTC-3′ and
reverse, 5′-CTCCGCTTCAGGTTGTTCAT-3′; β-actin forward,
5′-CCTGGACTTCGAGCAAGAGA-3′ and reverse, 5′-ACTTGCGCTCAGGAGGAGCA-3′.
The PCR conditions were as follows: 10 min at 95°C, followed by 40
cycles of 15 sec at 95°C, 30 sec at 60°C and 30 sec at 72°C, and a
final extension for 5 min at 72°C. The relative expression level of
Ep3 was normalized to β-actin levels. The relative expression
levels were calculated using the 2−ΔΔCt method (30).
Western blot analysis
The protein was extracted from tissues and cells
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China) supplemented with a PMSF
protease inhibitor (Sigma-Aldrich; Merck KGaA). Protein
concentration was quantified using a bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). A total of 40
µg protein lysates were separated via 10–12% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes. Membranes
were blocked in 5% bovine serum albumin (Sigma Aldrich; Merck KGaA)
at room temperature for 1 h, and subsequently immunoblotted with
the following primary antibodies according to the recommended
dilution concentration: Anti-Ep3 (1:500; cat. no. P8372;
Sigma-Aldrich; Merck KGaA), anti-caspase-3 (1:1,000; cat. no.
9662), anti-B cell lymphoma (Bcl)-2 (1:1,000; cat. no. 2872),
anti-Bcl-associated × protein (Bax; 1:1,000; cat. no. 2772; all
Cell Signaling Technology, Inc., Danvers, MA, USA), anti-MMP-9
(1:500; cat. no. ab58803; Abcam, Cambridge, UK), anti-VEGF
(1:1,000; cat. no. V6627; Sigma-Aldrich; Meck KGaA), anti-TGF-β1
(1:1,000; cat. no. SAB4502954; Sigma-Aldrich; Merck KGaA),
anti-Smad-2 (1:1,000; cat. no. ab63576), anti-Smad-3 (1:2,000; cat.
no. ab40854), anti-phosphorylated (p)-Smad2 (1:800; cat. no.
ab53100), anti-p-Smad3 (1:2,000; cat. no. ab52903) and anti-GAPDH
(1:2,000; cat. no. ab8245; all Abcam) at 4°C overnight. Then,
membranes were incubated with corresponding horseradish peroxidase
(HRP)-conjugated secondary antibodies: Anti-rabbit IgG (1:10,000;
cat. no. 7074), anti-mouse IgG (1:10,000; cat. no. 7076; both Cell
Signaling Technology, Inc.) and donkey anti-goat IgG (1:10,000;
cat. no. ab6885; Abcam) at room temperature for 1 h. GAPDH was used
as the loading control. The blots were detected using an enhanced
chemiluminescence solution (EMD Millpore, Billerica, MA, USA),
followed by observing the signals under the Molecular Imager
ChemiDoc XRS Gel Imagine System (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The relative intensities of protein bands were
analyzed using Image lab analysis software (version 4.0; Bio-Rad
Laboratories, Inc.).
MTT assay
Following transfection with Ep3 siRNA or treatment
with L-798106 for 48 h, the cells were treated with 100 µg/ml MTT
for 4 h at 37°C, followed by dissolving of the formazan crystals
with 100 µl of dimethyl sulfoxide. The absorbance value was
determined at a wavelength of 490 nm.
Wound-healing assay
Following transfection with Ep3 siRNA for 6 h,
artificial wounds were created by scraping using a sterile 200-µl
pipette tip. In a separate assay, following washing with
phosphate-buffered saline (PBS), the cells were cultured in
RPMI-1640 medium for 48 h. Artificial wounds were created followed
by treatment with L-798106 for 48 h to measure the effects of
L-798106. Images of cell migration were obtained at 0 h and 48 h
using an inverted light microscope (magnification, ×40). The
distance between the two edges of a wound was calculated using
ImageJ software (version 1.8.0; National Institutes of Health,
Bethesda, MD, USA). The width of wound at 0 h was regarded as 100%
and the rate of wound healing=(0 h width of the wound-48 h width of
the wound)/(0 h width of the wound).
Transwell assay
Following transfection with Ep3 siRNA or treatment
with L-798106 for 48 h, the cells were harvested and washed twice
with PBS. A total of 200 ml cell suspension in serum-free medium
(1×105 cells) was seeded in the upper compartment of
Transwell plates (with Matrigel-coated membrane; 8-µm pore size; BD
Biosciences, Franklin Lakes, NJ, USA). In the lower chamber, 600 µl
medium containing 10% FBS was added. After 24 h incubation, cells
remaining on the upper membranes were removed by cotton swabs. The
migrated cells were fixed with 4% paraformaldehyde for 30 min,
stained with 0.4% crystal violet for 20 min at room temperature,
and then counted under an inverted light microscope (magnification,
×200).
Flow cytometry assay
Following transfection with Ep-3 siRNA or treatment
with L-798106 for 48 h, the cells were harvested and stained with
Annexin V/propidium iodide double staining kit (BD Biosciences)
according to the manufacturer's protocol. Apoptotic cells were
assessed by flow cytometry on an FC500 flow cytometer (Beckman
Coulter, Inc., Brea, CA, USA).
NSCLC xenografts
Specific pathogen-free (SPF) athymic nude mice (all
male; age range, 6–8 weeks; weighing 20–25 g), purchased from the
Experimental Animal Center of the Southwest Medical University
(Sichuan, China), were housed and manipulated according to
protocols approved by the Experimental Animal Center of the
Southwest Medical University. They were maintained at a constant
humidity (60±5%) and temperature (20±1°C) and kept on a 12-h
light/dark cycle. All animals were provided with food and water
ad libitum. In order to research tumorigenicity of Ep3 in
vivo, A549 cells were transfected with or without Ep3 siRNA.
Each mouse was subcutaneously inoculated with either
1×107 Ep3 siRNA transfected A549 cells or control cells
(fluorescent-labeled) in 50% Matrigel (BD Biosciences) (Each group
contained 10 mice). Following the development of a palpable tumor,
the tumor volume was monitored every 5 days and assessed by
measuring the two perpendicular dimensions using a caliper with the
formula: (a × b2)/2, where a is the larger and b is the
smaller dimension of the tumor. At 30 days after inoculation, the
mice were sacrificed and tumor weights were assessed. Tumors from
each mouse were randomly selected for immunohistochemical (IHC)
analysis. All animal procedures were performed according to
relevant national and international guidelines and were approved by
the Animal Experimental Ethical Committee.
Immunohistochemistry
NSCLC tumor tissues were fixed in 10% formalin at
4°C for 24 h, and then embedded in paraffin for IHC analysis.
Briefly, 5 µm-thick paraffin sections were deparaffinized in
xylene, and gradually rehydrated in 100, 95 and 75% ethanol. In
order to quench the activity of endogenous peroxidase, the tissue
sections were incubated in 30% H2O2 for 30
min at room temperature. Following antigen retrieval in heated 10
mM citrate buffer for 10 min at room temperature, the tissue
sections were immunostained with mouse anti-human MMP-9 primary
antibody (cat. no. ab58803; 1:500; Abcam) overnight at 4°C. Tissues
were incubated with the corresponding anti-mouse HRP-conjugated
secondary antibody for 1 h at room temperature. Images were viewed
under a light microscope (magnification, ×200).
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA). The data
were analyzed by one-way analysis of variance followed by
Bonferroni's post hoc test for multiple comparisons. The Student's
t-test was used for comparisons between two groups. Each experiment
was performed in triplicate and the values are presented as the
mean ± standard error of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
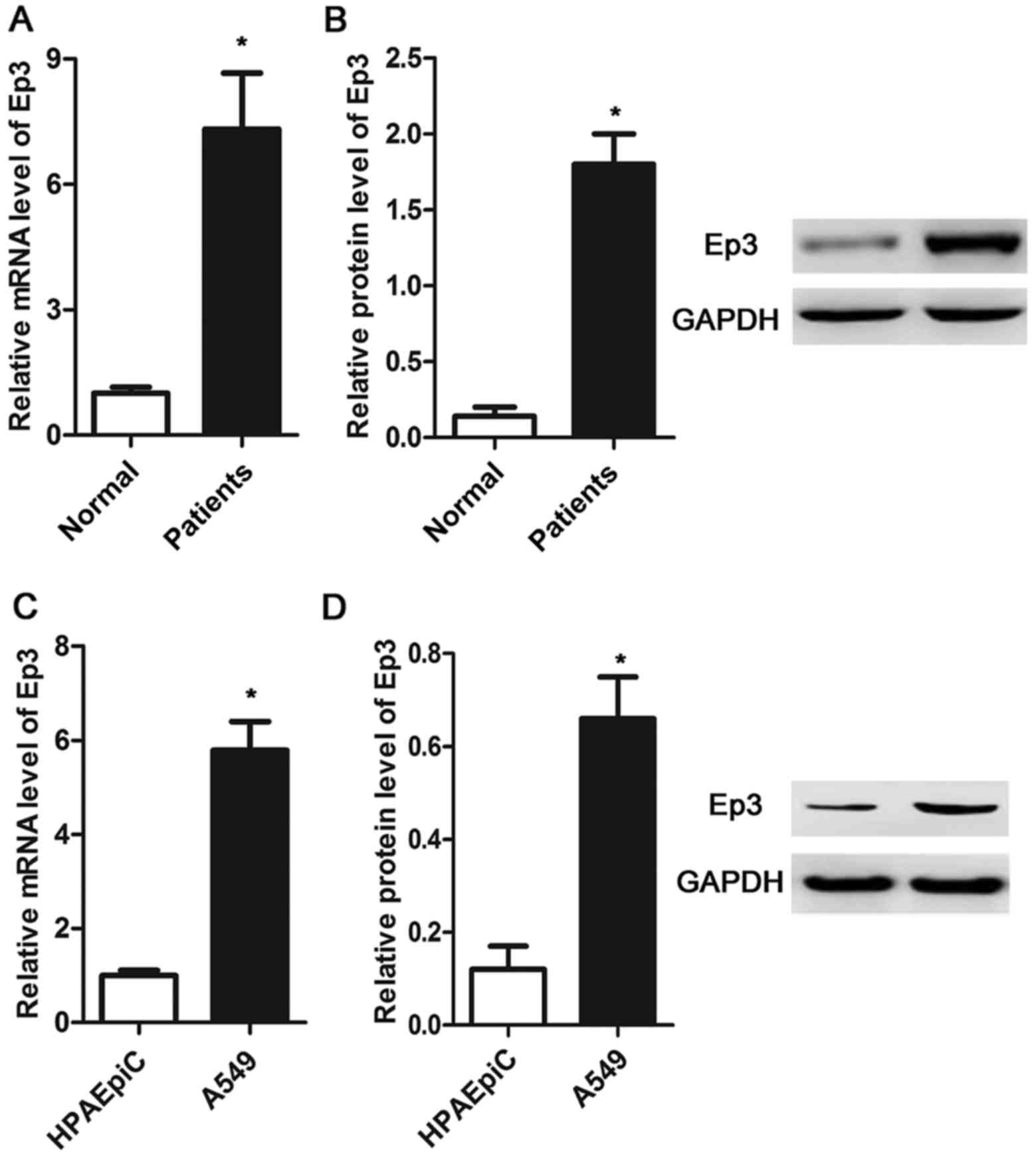
Expression of Ep3 is increased in
NSCLC tissues and A549 cells
The expression of Ep3 was measured by RT-qPCR and
western blotting in NSCLC tissues, A549 cells and HPAEpiC cells. As
presented in Fig. 1A and B, the mRNA
and protein expression levels were significantly increased in the
NSCLC tissues compared with the normal tissues (P<0.05). The
same results were observed in the NSCLC cell line, whereby the
expression of Ep3 was significantly upregulated in A549 cells
compared with HPAEpiC cells (P<0.05; Fig. 1C and D).
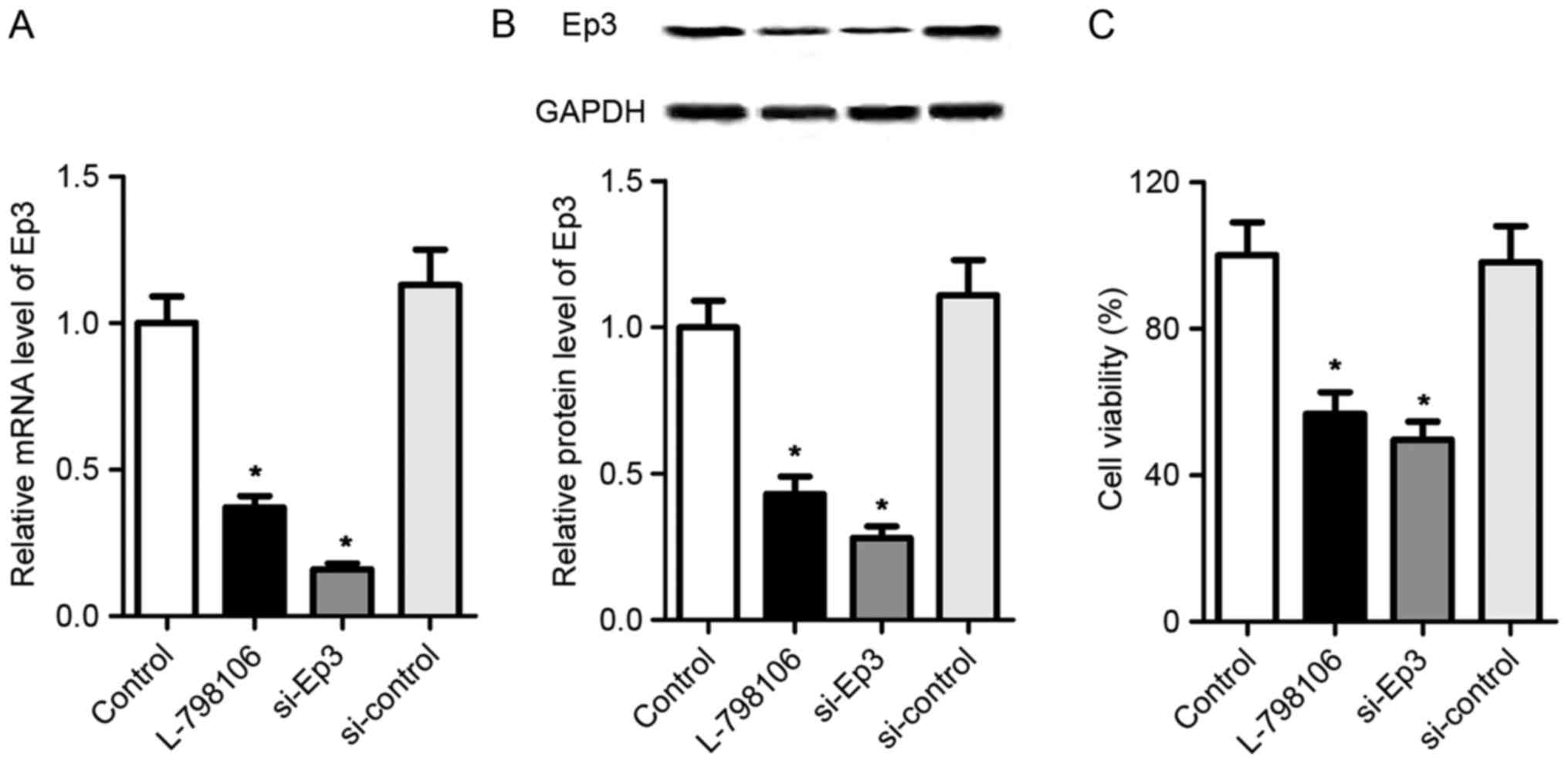
Inhibition of Ep3 inhibits the cell
viability of A549 cells
The elevated expression of Ep3 in NSCLC tissues and
A549 cells suggest that Ep3 may serve an important function in the
development of NSCLC. To explore the biological role of Ep3, A549
cells were transfected with Ep3 siRNA or its inhibitor L-798106 to
suppress the expression of Ep3. As presented in Fig. 2A and B, the mRNA and protein
expression levels of Ep3 were significantly downregulated in
L-798106 and si-Ep3 groups as compared with the control A549 cells
(P<0.05). Subsequently, it was determined whether Ep3 was
involved in the regulation of the cell viability in A549 cells
using an MTT assay. As presented in Fig.
2C, compared with control group, the cells treated with
L-798106 or Ep3 siRNA exhibited significantly lower cell viability
rates (P<0.05). These data suggest that Ep3 deficiency may be
associated with the growth activity of A549 cells.
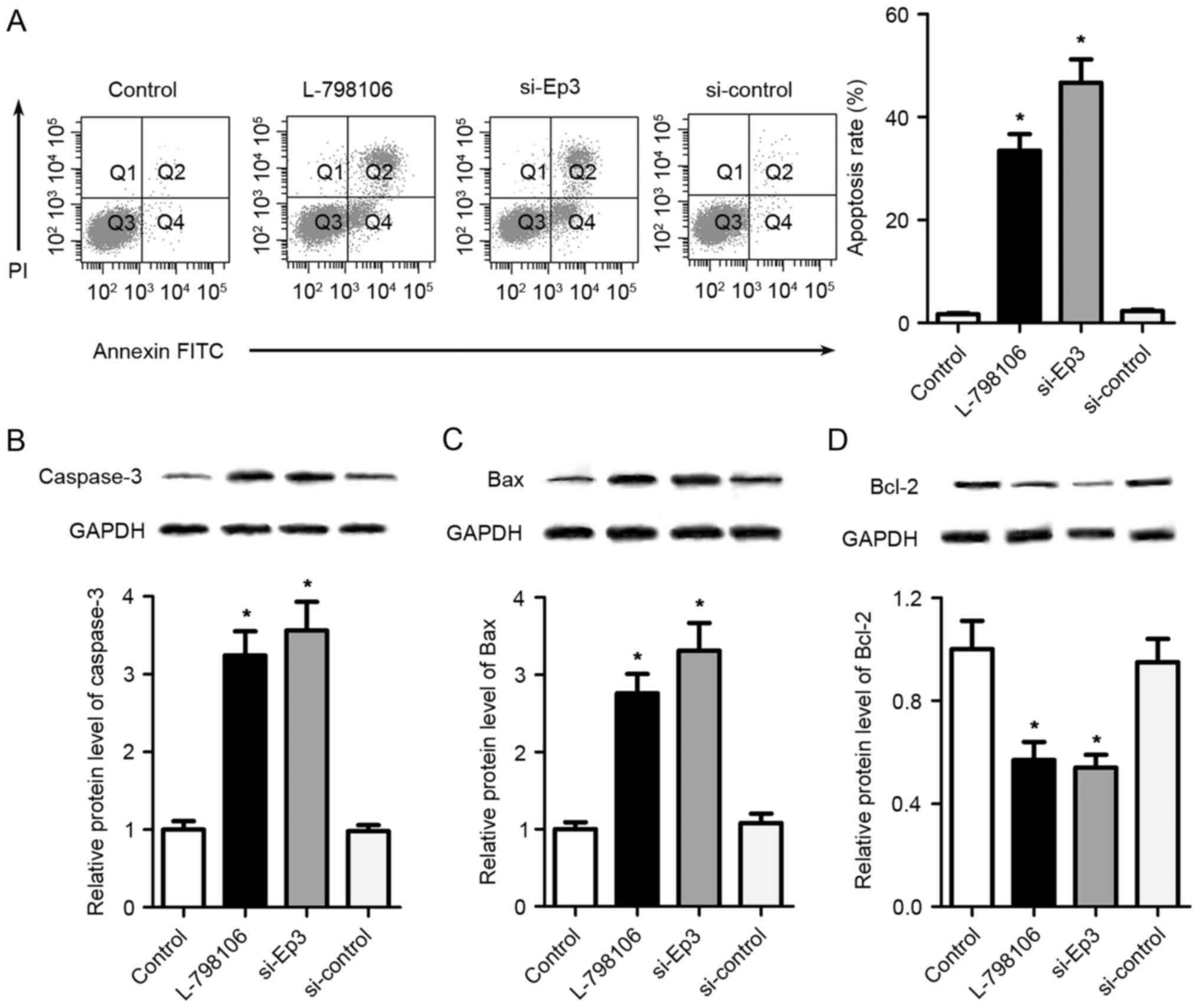
Ep3 deficiency promotes the cell
apoptosis of A549 cells
To further investigate the effects of Ep3 on the
apoptosis of A549 cells, a flow cytometry assay was performed. As
presented in Fig. 3A, the proportion
of apoptotic cells was significantly increased in L-798106 and
si-Ep3 groups compared with control group (P<0.05).
Additionally, western blotting was performed to assess the
expression of apoptosis-associated proteins. As presented in
Fig. 3B-D, compared with the control
group, Ep3 deficiency by L-798106 or Ep3 siRNA significantly
upregulated the expression of caspase-3 and Bax, and downregulated
the expression of Bcl-2 (P<0.05).
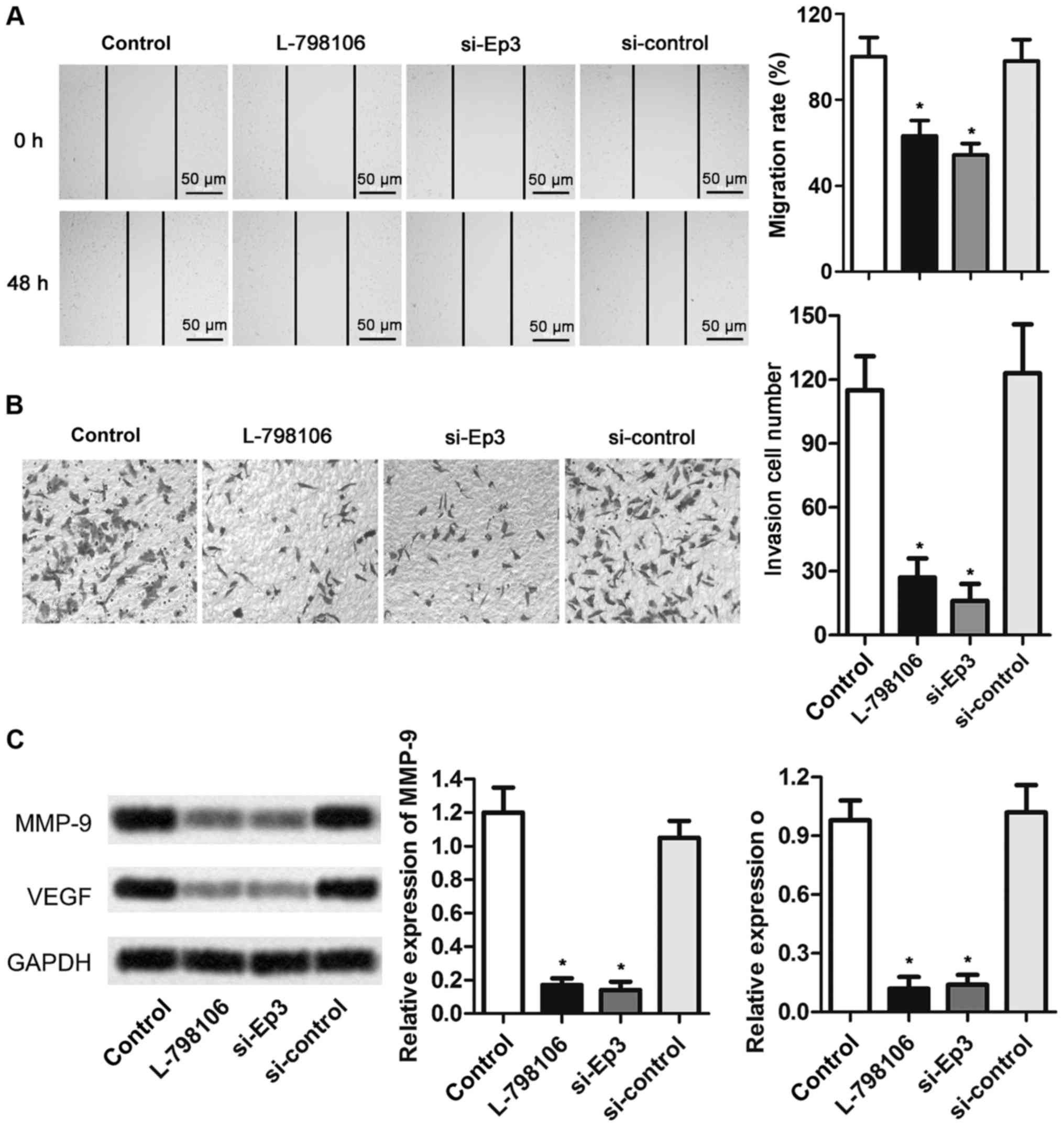
Ep3 deficiency inhibits the cell
migration and invasion of A549 cells
To determine whether Ep3 was involved in the
migration and invasion of A549 cells, wound-healing and Transwell
assays were performed. As presented in Fig. 4, the abilities of cell migration and
invasion in L-798106 and si-Ep3 groups were significantly
suppressed compared with that in control cells (P<0.05).
Furthermore, the expression level of matrix metalloproteinase MMP-9
and VEGF was significantly reduced in L-798106 and si-Ep3 groups
compared with control group (P<0.05). These results suggest that
Ep3 deficiency may be a suppressor of cell motility of NSCLC.
Together, these data suggest that inhibition of Ep3 may be an
effective approach in the treatment of NSCLC due to its functions
in the cell viability, migration, invasion and apoptosis in NSCLC
cells.
Ep3 deficiency suppresses the growth
of A549 cells via inhibition of the TGF-β/Smad signaling
pathway
In the present study, it was demonstrated that Ep3
deficiency was involved in the regulation of cell viability,
migration, invasion and apoptosis of A549 cells, and the underlying
molecular mechanisms were investigated further. A number of studies
have reported that TGF-β/Smad signaling is an important pathway in
the progression of NSCLC (31–33),
therefore whether TGF-β/Smad signaling mediated the roles of Ep3
was further investigated in A549 cells. The expression of
pathway-associated proteins was evaluated using western blotting.
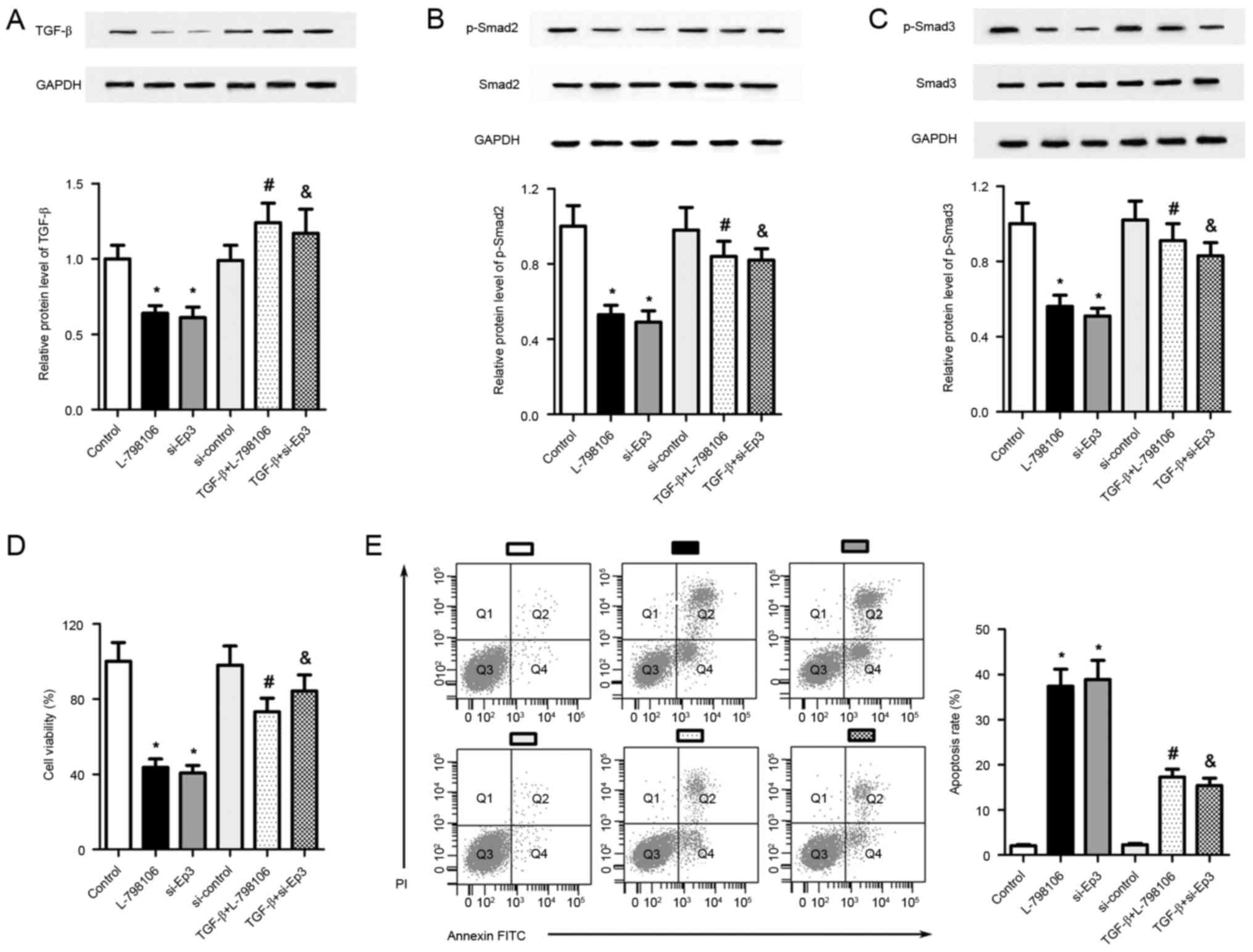
As presented in Fig. 5A-C, treatment
with L-798106 or Ep3 siRNAs significantly inhibited the expression
of TGF-β, p-Smad2 and p-Smad3 inA549 cells, suggesting that
TGF-β/Smad signaling may mediate the functions of Ep3 in A549 cells
(P<0.05). To address the direct involvement of TGF-β/Smad
signaling in the regulation of Ep3 in A549 cells, pcDNA3.1-TGF-β
plasmids were introduced into A549 cells prior to the treatment of
L-798106 or Ep3 siRNA. As presented in Fig. 5A-C, compared with the L-798106 group,
transfection of pcDNA3.1-TGF-β plasmids significantly elevated the
levels of TGF-β, p-Smad2 and p-Smad3 (P<0.05). The same results
were observed in the TGF-β+si-Ep3 group as compared with the si-Ep3
group (P<0.05). The MTT assay revealed that the reduction in
cell viability caused by Ep3 knockdown was partially reversed in
pcDNA3.1-TGF-β-transfected A549 cells (P<0.05; Fig. 5D). Furthermore, compared with the
L-798106 or si-Ep3 group, cell apoptosis was reduced when TGF-β was
overexpressed in these cells (P<0.05; Fig. 5E). These data suggest that cell
viability inhibition and cell apoptosis induced by Ep3 knockdown
were at least partially associated with the inhibition of
TGF-β/Smad signaling.
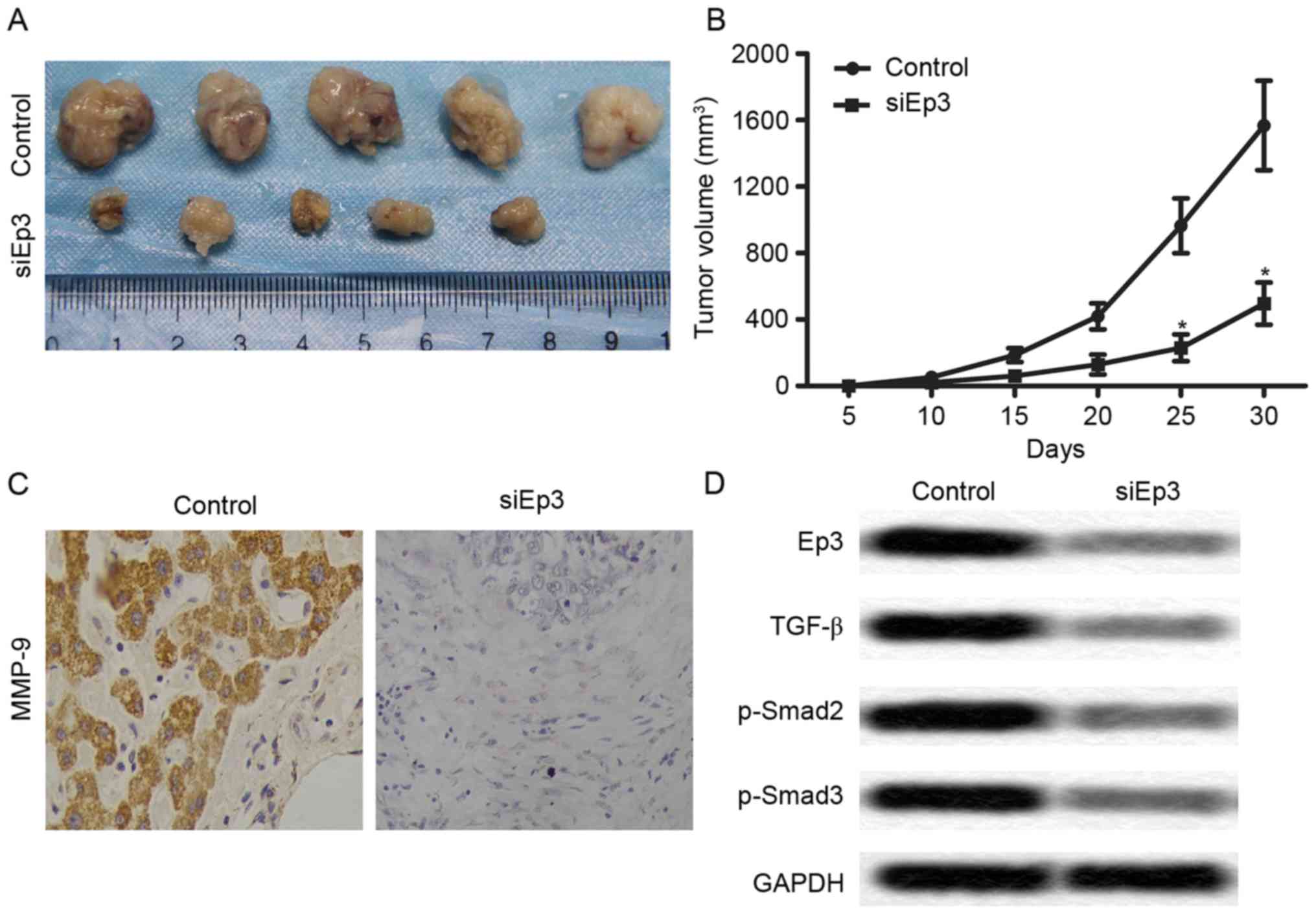
Ep3 deficiency inhibits tumor growth
and metastasis in vivo
To investigate the effects of Ep3 deficiency on
NSCLC cells growth and invasion in vivo, a xenograft mouse
model was established by subcutaneous injection of A549 cells to
SPF nude mice. The mean tumor volume was significantly reduced in
the si-Ep3 group compared with the control group after 25 days
(P<0.05; Fig. 6A and B).
Furthermore, the expression level of MMP-9 was markedly decreased
in the si-Ep3 group compared with the control group using IHC
analysis (Fig. 6C). In addition, the
levels of Ep3, TGF-β, p-Smad2 and p-Smad3 were markedly reduced in
the si-Ep3 group compared with the control group (Fig. 6D). Combined with the results of the
aforementioned experiments, it is hypothesized that Ep3 deficiency
inhibits tumor growth and metastasis in vivo.
Discussion
NSCLC remains a threat to public health worldwide.
Despite advances in therapeutic modalities, little progress has
been made in improving the quality of life and survival in patients
with advanced and metastatic NSCLC. Therefore, an increased
understanding of the underlying molecular mechanism of cancer
proliferation and migration is essential for the development of
novel effective therapeutic strategies against NSCLC. In the
present study, it was identified that inhibition of Ep3
significantly attenuated the viability and migration, and promoted
the apoptosis of A549 cells through suppression of the TGF-β/Smad
signaling pathway, suggesting that inhibition of Ep3 may be a novel
therapeutic strategy for the treatment of NSCLC.
Increased expression of COX-2 and the consequent
upregulation of PGE2 have been implicated in the pathogenesis of
several types of cancer, including colon, breast, and lung cancer
due to their roles in regulating cell growth and invasion (34–38). Ep3
is a receptor via which PGE2 exerts its cellular effects and has
been reported to be involved in the progression of lung cancer
(20,23,24,39).
However, the effects and detailed molecular mechanisms of Ep3 in
lung cancer remain unresolved. In the present study, it was
identified that Ep3 expression was significantly upregulated in
NSCLC lung tissues and A549 cells, suggesting that Ep3 may function
as a tumor promoter in NSCLC. To investigate the effects of Ep3 in
the development of NSCLC, A549 cells were transfected with Ep3
siRNA or treated with Ep3 inhibitor L-798106 to downregulate the
expression of Ep3. MTT assays, wound-healing assays and Transwell
assays revealed that pharmacological inhibition with L-798106 and
Ep3 siRNA transfection significantly reduced the cell viability,
and the migration and invasion abilities of A549 cells.
Furthermore, flow cytometry analysis demonstrated that the
proportion of apoptotic cells was significantly increased when A549
cells were treated with L-798106 and Ep3 siRNA. Additionally, the
expression levels of several apoptosis-associated proteins were
assessed using western blotting, and the results revealed that the
protein levels of Caspase-3 and Bax had were significantly
upregulated, whereas the expression of Bcl-2 was significantly
downregulated when the expression of Ep3 was inhibited by L-798106
or Ep3 siRNA. These results demonstrated that decreased Ep3
expression may be a factor contributing to inhibition of the
development of NSCLC, including suppressing proliferation and
invasion, and promoting apoptosis of lung cancer cells.
The activation of TGF-β signaling, as well as the
subsequent phosphorylation of Smad2 and Smad3 have been reported to
serve an important function in the regulation of expression of
numerous genes, and thus regulates cellular responses, including
proliferation, migration and apoptosis, in various types of cancer,
including lung cancer (40,41). This suggests that TGF-β/Smad signaling
may be associated with the development of lung cancer. Therefore,
approaches to harbor this signaling may be valuable for treating
lung cancer. A previous study reported that inhibition of Ep3
attenuates pulmonary hypertension through suppression of Rho/TGF-β1
signaling (33), indicating that the
regulation of Ep3 may be associated with TGF-β signaling. It was
therefore investigated whether TGF-β signaling is involved in the
effects of Ep3 in lung cancer cells. In the present study,
treatment with L-798106 or Ep3 siRNA significantly inhibited the
expression of TGF-β, p-Smad2 and p-Smad3 in A549 cells, suggesting
that TGF-β/Smad signaling is involved in the regulation of Ep3 in
A549 cells. To further confirm these results, pcDNA3.1-TGF-β
plasmids were introduced into A549 cells prior to treatment with
L-798106 or Ep3 siRNA. MTT and flow cytometric assays revealed that
the inhibition of cell viability and the promotion of cell
apoptosis induced by Ep3 knockdown were partially reversed by the
overexpression of TGF-β. These data suggest that the Ep3-mediated
biological effects in A549 cells are at least partially mediated by
the TGF-β/Smad signaling pathway.
In conclusion, the results of the present study
suggest that inhibition of Ep3 attenuates the viability and
migration, and promotes the apoptosis of A549 cells, which was
associated with the suppression of TGF-β/Smad signaling. The
current study provides a novel insight into the underlying
molecular mechanism associated with Ep3-mediated effects in NSCLC
cells, and suggests that targeting the Ep3/TGF-β/Smad signaling
pathway provides novel therapeutic strategies for the treatment of
NSCLC. Further studies are required to verify these conclusions
in vivo.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL and YL made substantial contributions to the
study conception and design. LL and DY acquired data and performed
analysis and interpretation of the data. LL drafted the manuscript.
YL revised the manuscript. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
All of the patients provided written informed
consent and approval was provided by the Ethics Committee of
Zhoukou Central Hospital for the human studies. The animal studies
were approved by the Experimental Animal Center of the Southwest
Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Travis WD, Travis LB and Devesa SS: Lung
cancer. Cancer. 75:191–202. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Novello S, Barlesi F, Califano R, Cufer T,
Ekman S, Levra MG, Kerr K, Popat S, Reck M, Senan S, et al:
Metastatic non-small-cell lung cancer: ESMO Clinical Practice
Guidelines for diagnosis, treatment and follow-up. Ann Oncol.
27:v1–v27. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu G, Pei F, Yang F, Li L, Amin AD, Liu
S, Buchan JR and Cho WC: Role of autophagy and apoptosis in
non-small-cell lung cancer. Int J Mol Sci. 18:E3672017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Payandeh M, Sadeghi M, Sadeghi E and
Aeinfar M: The survival of lung cancer based on type of treatment
in non-small-cell lung cancer: A review of literatureInternational
Congress on Anti-Cancer Treatment. Paris, France: 2015
|
|
5
|
Lim BJ, Jung SS, Choi SY and Lee CS:
Expression of metastasis-associated molecules in non-small cell
lung cancer and their prognostic significance. Mol Med Rep.
3:43–49. 2010.PubMed/NCBI
|
|
6
|
Müllertidow C, Diederichs S, Bulk E, Pohle
T, Steffen B, Schwäble J, Plewka S, Thomas M, Metzger R, Schneider
PM, et al: Identification of metastasis-associated receptor
tyrosine kinases in non-small cell lung cancer. Cancer Res.
65:1778–1782. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schoenberger SD, Kim SJ, Sheng J, Rezaei
KA, Lalezary M and Cherney E: Increased prostaglandin E2 (PGE2)
levels in proliferative diabetic retinopathy, and correlation with
VEGF and inflammatory cytokines. Invest Ophthalmol Vis Sci.
53:5906–5911. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chell S, Kaidi A, Kadi A, Williams AC and
Paraskeva C: Mediators of PGE2 synthesis and signalling downstream
of COX-2 represent potential targets for the prevention/treatment
of colorectal cancer. Biochim Biophys Acta. 1766:104–119.
2006.PubMed/NCBI
|
|
9
|
Li Z, Zhang Y, Wanju K and Yehia D: PGE2
promotes renal carcinoma cell invasion through activated RalA.
Oncogene. 32:1408–1415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim CH, Park YG, Noh SH and Kim YK: PGE2
induces the gene expression of bone matrix metalloproteinase-1 in
mouse osteoblasts by cAMP-PKA signaling pathway. Int J Biochem Cell
Biol. 37:375–385. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dajani OF, Meisdalen K, Guren TK, Aasrum
M, Tveteraas IH, Lilleby P, Thoresen GH, Sandnes D and
Christoffersen T: Prostaglandin E2 upregulates EGF-stimulated
signaling in mitogenic pathways involving Akt and ERK in
hepatocytes. J Cell Physiol. 214:371–380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goulet JL, Pace AJ, Key ML, Byrum RS,
Nguyen M, Tilley SL, Morham SG, Langenbach R, Stock JL, McNeish JD,
et al: E-prostanoid-3 receptors mediate the proinflammatory actions
of prostaglandin E2 in acute cutaneous inflammation. J Immunol.
173:1321–1326. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen L, Miao Y, Zhang Y, Dou D, Liu L,
Tian X, Yang G, Pu D, Zhang X, Kang J, et al: Inactivation of the
E-prostanoid 3 receptor attenuates the angiotensin II pressor
response via decreasing arterial contractility. Arterioscler Thromb
Vasc Biol. 32:3024–3032. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miyata Y, Ohba K, Matsuo T, Watanabe S,
Hayashi T, Sakai H and Kanetake H: Tumor-associated stromal cells
expressing E-prostanoid 2 or 3 receptors in prostate cancer:
Correlation with tumor aggressiveness and outcome by angiogenesis
and lymphangiogenesis. Urology. 81:136–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Reader J, Holt D and Fulton A:
Prostaglandin E 2 EP receptors as therapeutic targets in breast
cancer. Cancer Metastasis Rev. 30:449–463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Amano H, Ito Y, Suzuki T, Kato S, Matsui
Y, Ogawa F, Murata T, Sugimoto Y, Senior R, Kitasato H, et al:
Roles of a prostaglandin E-type receptor, EP3, in upregulation of
matrix metalloproteinase-9 and vascular endothelial growth factor
during enhancement of tumor metastasis. Cancer Sci. 100:2318–2324.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang T, Hou J, He M, Wang L, Zheng M, Wang
X and Xia J: Actinidia chinensis Planch root extract (acRoots)
inhibits hepatocellular carcinoma progression by inhibiting EP3
expression. Cell Biol Toxicol. 32:499–511. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang C, Wang Q, Xu Z, Li WS, Chen C, Yao
XQ and Liu FK: Cyclooxygenase-2 knockdown using retinoic acid
chalcone (RAC), a promising therapeutic strategy for colon cancer.
Am J Cancer Res. 5:2012–2021. 2015.PubMed/NCBI
|
|
19
|
Yano T, Zissel G, Muller-Qernheim J, Shin
Jae S, Satoh H and Ichikawa T: Prostaglandin E-2 reinforces the
activation of Ras signal pathway in lung adenocarcinoma cells via
EP3. FEBS Lett. 518:154–158. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamaki T, Endoh KM, Miyahara M, Nagamine
I, Thi Thu, Huong N, Sakurai H, Pokorny J and Yano T: Prostaglandin
E2 activates Src signaling in lung adenocarcinoma cell via EP3.
Cancer Lett. 214:115–120. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Israel DD and Regan JW: EP(3) prostanoid
receptor isoforms utilize distinct mechanisms to regulate ERK 1/2
activation. Biochim Biophys Acta. 1791:238–245. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang P, Zhu F, Lee NH and Konstantopoulos
K: Shear-induced interleukin-6 synthesis in chondrocytes: Roles of
E Prostanoid (EP) 2 and EP3 in cAMP/protein KINASE A- and
PI3-K/Akt-dependent NF-κB activation. J Biol Chem. 285:24793–24804.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levy L and Hill CS: Alterations in
components of the TGF-beta superfamily signaling pathways in human
cancer. Cytokine Growth Factor Rev. 17:41–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Massagué J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Massagué J and Wotton D: Transcriptional
control by the TGF-beta/Smad signaling system. EMBO J.
19:1745–1754. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Massagué J: How cells read TGF-beta
signals. Nat Rev Mol Cell Biol. 1:169–178. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zu L, Xue Y and Wang J, Fu Y, Wang X, Xiao
G, Hao M, Sun X, Wang Y, Fu G and Wang J: The feedback loop between
miR-124 and TGF-β pathway plays a significant role on non-small
cell lung cancer metastasis. Carcinogenesis. 37:333–343. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Yi J, Chen X, Zhang Y, XU M and
Yang Z: The regulation of cancer cell migration by lung cancer
cell-derived exosomes through TGF-β and IL-10. Oncol Lett.
11:1527–1530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu A, Zuo C, He Y, Chen G, Piao L, Zhang
J, Xiao B, Shen Y, Tang J, Kong D, et al: EP3 receptor deficiency
attenuates pulmonary hypertension through suppression of Rho/TGF-β1
signaling. J Clin Invest. 125:1228–1242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang JX, Zhai JF, Yang XT and Wang J:
MicroRNA-132 inhibits migration, invasion and
epithelial-mesenchymal transition by regulating TGFβ1/Smad2 in
human non-small cell lung cancer. Eur Rev Med Pharmacol Sci.
20:3793–3801. 2016.PubMed/NCBI
|
|
32
|
Da C, Liu Y, Zhan Y, Liu K and Wang R:
Nobiletin inhibits epithelial-mesenchymal transition of human
non-small cell lung cancer cells by antagonizing the TGF-β1/Smad3
signaling pathway. Oncol Rep. 35:2767–2774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Li M, Hu M, Wei P and Zhu W: BAMBI
overexpression together with β-sitosterol ameliorates NSCLC via
inhibiting autophagy and inactivating TGF-β/Smad2/3 pathway. Oncol
Rep. 37:3046–3054. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Prescott SM and Fitzpatrick FA:
Cyclooxygenase-2 and carcinogenesis. Biochim Biophys Acta.
1470:M69–M78. 2000.PubMed/NCBI
|
|
35
|
Dufour M, Faes S, Dormond-Meuwly A,
Demartines N and Dormond O: PGE2-induced colon cancer growth is
mediated by mTORC1. Biochem Biophys Res Commun. 451:587–591. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bocca C, Ievolella M, Autelli R, Motta M,
Mosso L, Torchio B, Bozzo F, Cannito S, Paternostro C, Colombatto
S, et al: Expression of Cox-2 in human breast cancer cells as a
critical determinant of epithelial-to-mesenchymal transition and
invasiveness. Exp Opin Ther Targets. 18:121–135. 2014. View Article : Google Scholar
|
|
37
|
Sharma S, Yang SC, Zhu L, Reckamp K,
Gardner B, Baratelli F, Huang M, Batra RK and Dubinett SM: Tumor
cyclooxygenase-2/prostaglandin E2-dependent promotion of FOXP3
expression and CD4+ CD25+ T regulatory cell activities in lung
cancer. Cancer Res. 65:5211–5220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pan J, Yang Q, Shao J, Zhang L, Ma J, Wang
Y, Jiang BH, Leng J and Bai X: Cyclooxygenase-2 induced β1-integrin
expression in NSCLC and promoted cell invasion via the
EP1/MAPK/E2F-1/FoxC2 signal pathway. Sci Rep. 6:338232016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Amano H, Hayashi I, Endo H, Kitasato H,
Yamashina S, Maruyama T, Kobayashi M, Satoh K, Narita M, Sugimoto
Y, et al: Host prostaglandin E2-EP3 signaling regulates
tumor-associated angiogenesis and tumor growth. J Exp Med.
197:221–232. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shen JL, Yan CH, Liu Y, Yan XQ, Zhang XL,
Jin Y, Zhang KF, Sang ZF, Zhang GY, Li P and Fu SB: Studies of
TGF-β/Smads expression in lung cancer. Yi Chuan Xue Ba. 30:681–686.
2003.(In Chinese).
|
|
41
|
Liu RY, Zeng Y, Lei Z, Wang L, Yang H, Liu
Z, Zhao J and Zhang HT: JAK/STAT3 signaling is required for
TGF-β-induced epithelial-mesenchymal transition in lung cancer
cells. Int J Oncol. 44:1643–1651. 2014. View Article : Google Scholar : PubMed/NCBI
|