Introduction
T-cell lymphoma accounts for 9.7% of all lymphoid
malignancies, with an incidence rate of 1.17 cases per 100,000
people in Hong Kong (1). T-cell
lymphoma has distinct clinical features and a complicated
epidemiology (2). Despite treatment
with intensive chemotherapy or combined chemotherapy, patients with
T-cell lymphoma continue to exhibit suboptimal outcomes and a poor
disease progression (3,4). The development of effective therapeutic
treatments to improve the prognosis of T-cell lymphoma is
required.
The c-Myc transcription factor is a short-lived
nuclear phosphoprotein that serves a vital role in regulating cell
proliferation, differentiation and apoptosis (5,6). Notably,
overexpression of c-Myc has been associated with the occurrence of
hematopoietic malignancies, including lymphoma and leukemia
(7). Glycogen synthase kinase 3β
(GSK3β) regulates c-Myc levels and activity by phosphorylating
c-Myc at threonine 58 to promote its degradation by the
ubiquitin-proteasome pathway in lymphoma (8). A previous study in acute lymphocytic
leukemia cells revealed that GSK3β exhibits a low affinity to the
stabilized c-Myc oncoprotein (7).
Taken together, these results suggested that downregulation of
c-Myc expression may be an effective strategy for the treatment of
lymphoma.
The phosphatidylinositol 3-kinase/protein kinase B
(PI3K/Akt)/GSK3β signaling pathway serves important regulatory
roles in differentiation, cell survival and apoptosis (9). Akt is a critical downstream factor of
PI3K and is activated through PI3K phosphorylation of Akt at serine
473 (10). Phosphorylated Akt (p-Akt)
subsequently activates GSK3β, its principal physiological
substrate, by phosphorylation of GSK3β at serine 9 (10). Activated Akt supported a survival
signal by phosphorylating apoptotic factors, including GSK3β, and
preventing cells from apoptosis (9,11,12). Previous studies have revealed that the
Akt/GSK3β signaling pathway is activated to promote cell
proliferation and decrease apoptosis in various types of carcinoma
(13). The function of the
p-Akt/GSK3β pathway is to promote proliferation and to decrease
apoptosis. p-Akt is involved in tumor proliferation and a poor
prognosis in several types of human cancer, and it may be an
important factor in the development of lymphomas (14,15).
Dihydroartemisinin (DHA) is a compound isolated from
Artemisia annua L, a Chinese medicinal herb. A few reports,
including our previous study, indicated DHA as a promising drug for
tumor therapy due to its antitumor activity in cancer cells
(16–18). DHA significantly induced c-Myc
oncoprotein degradation in tumor cells overexpressing c-Myc, a
process that involved GSK3β phosphorylation of c-Myc at threonine
58 (19). DHA also suppressed cell
proliferation via the Akt/GSK3β/cyclin D1 signaling pathway in A549
lung cancer cells and promoted the degradation of c-Myc via
increased phosphorylation of c-Myc (19,20).
Based on these studies, we hypothesized that DHA
exhibits antiproliferation activity in T-cell lymphoma cells and
that DHA may mediate c-Myc oncoprotein degradation and downregulate
the Akt/GSK3β signaling pathway in T-cell lymphoma cells.
Therefore, the present study examined the potential anticancer
effect of DHA on T-cell lymphoma cells (Jurkat and HuT-78 cells).
We also investigated the underlying mechanism of DHA was also
investigated in T-cell lymphoma cells and the potential involvement
of the AKT/GSK3β signaling pathway. This study may provide evidence
for DHA as a possible treatment for T-cell lymphoma cells.
Materials and methods
Materials and cell culture
DHA was obtained from Chunyou Biological Technology
Corporation (Shanghai, China) and dissolved in dimethyl sulfoxide
(DMSO; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) as a stock
solution of 8,000 µM at −20°C. RPMI-1640 medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) was used to dilute the
stocked DHA for each experiment.
T-cell lymphoma Jurkat and HuT-78 cell lines were
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China). Jurkat cells were maintained in
RPMI-1640 medium supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), and HuT-78 cells
were cultured in Iscove's modified Dulbecco's medium (Hyclone; GE
Healthcare Life Sciences) containing 20% fetal bovine serum. Medium
was supplemented with 100 U/ml penicillin, 100 mg/ml streptomycin
and 2.5 mg/ml amphotericin B, and cells were incubated at 37°C in a
humidified 5% CO2 atmosphere. The control group
comprised cells not exposed to DHA, while the DHA group comprised
cells treated with DHA.
Cell viability assay
Cell viability assay was evaluated using Cell
Counting kit-8 (CCK-8; Anhui Yiyuan Biotechnology Co., Ltd., Anhui,
China). Jurkat cells were seeded at a density of 8×103
cells/well in 100 µl RPMI-1640 medium supplemented with 10% fetal
bovine serum and HuT-78 cells were seeded at a density of
8×103 cells/well in 100 µl Iscove's modified Dulbecco's
medium containing 20% fetal bovine serum in a 96-well plate
(Costar; Corning Incorporated, Corning, NY, USA), and the cells
were incubated for 24, 36, 48, 60 or 72 h with DHA (0, 5, 10, 20,
40 or 80 µM) added to the culture medium. At each time point, 10 µl
CCK-8 was added to each well and the cells were subsequently
incubated at 37°C for 2 h. Absorbance was detected at 450 nm on a
microplate absorbance reader (Sunrise; Tecan Group Ltd., Männedorf,
Switzerland).
Cell apoptosis assay
Jurkat (1×106) were seeded onto a 6-well
plate with 2 ml RPMI-1640 medium supplemented with 10% fetal bovine
serum and HuT-78 (1.5×106) cells were seeded onto a
6-well plate with 2 ml Iscove's modified Dulbecco's medium
containing 20% fetal bovine serum followed by incubation at 37°C
for 48 h. Subsequently, DHA (0 or 15 µM for Jurkat cells and 0 or
30 µM for HuT-78 cells) was added. All cells were resuspended with
cold PBS (Hyclone; GE Healthcare Life Sciences) and centrifuged at
447.2 × g for 5 min at room temperature. Cells were resuspended in
300 µl cold binding buffer (BD Biosciences, Franklin Lakes, NJ,
USA) following the addition of 5 µl Annexin V-fluorescein
isothiocyanate (BD Biosciences) for 15 min in the dark at room
temperature. Propidium iodide (PI; 5 µl; BD Biosciences) was added
and incubated for another 5 min, and another 200 µl binding buffer
was added, followed by a flow cytometer (BD Biosciences) FlowJo
software version 7.6.1 (FlowJo LLC, Ashland, OR, USA) being used to
analyze each sample.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cultured cells
using RNAiso Plus (Takara Bio, Inc., Otsu, Japan), according to the
manufacturer's protocol. Pectrophotometry (Dynamica Scientific
Ltd., Newport Pagnell, UK) was used to determine the quality and
quantity of RNA. Total RNA was reverse transcribed into
single-stranded cDNA using a Prime Script™ RT reagent kit (Takara
Bio, Inc.), according to the manufacturer's protocols. Next, the
reverse transcribed single-stranded cDNA was amplified by SYBR1
Premix Ex Taq II (Takara Bio, Inc.) with a CFX96
spectrofluorometric thermal cycler (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The primers were synthesized by Tsingke
Biological Technology Co., Ltd. (Beijing, China; Table I). Thermocycling conditions included a
pre-incubation stage at 95°C for 30 sec, and 40 cycles of
amplification stage at 95°C for 5 sec and 60°C for 30 sec, and a
melt curve stage at 95°C for 15 sec, 65°C for 60 sec and 95°C for
15 sec. Relative expression of GAPDH mRNA was used as an endogenous
internal control. Gene expression changes was assessed with the
2−ΔΔCq method (21).
 | Table I.Reverse transcription-quantitative
polymerase chain reaction primer sequences. |
Table I.
Reverse transcription-quantitative
polymerase chain reaction primer sequences.
| Gene | Sequence | Size, bp |
|---|
| GAPDH |
| 258 |
|
Forward |
5′-AGAAGGCTGGGGCTCATTTG-3′ |
|
|
Reverse |
5′-AGGGGCCATCCACAGTCTTC-3′ |
|
| c-Myc |
| 204 |
|
Forward |
5′-CTTCTCTCCGTCCTCGGATTCT-3′ |
|
|
Reverse |
5′-GAAGGTGATCCAGACTCTGACCTT-3′ |
|
| Akt |
| 154 |
|
Forward |
5′-TGGACTACCTGCACTCGGAGAA-3′ |
|
|
Reverse |
5′-GTGCCGCAAAAGGTCTTCATGG-3′ |
|
| GSK3β |
| 150 |
|
Forward |
5′-CCGACTAACACCACTGGAAGCT-3′ |
|
|
Reverse |
5′-AGGATGGTAGCCAGAGGTGGAT-3′ |
|
| Bcl-2 |
| 127 |
|
Forward |
5′-ATCGCCCTGTGGATGACTGAGT-3′ |
|
|
Reverse |
5′-GCCAGGAGAAATCAAACAGAGGC-3′ |
|
| Bax |
| 103 |
|
Forward |
5′-TCAGGATGCGTCCACCAAGAAG-3′ |
|
|
Reverse |
5′-TGTGTCCACGGCGGCAATCATC-3′ |
|
Western blot analysis
Cells were washed with PBS and lysed with
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen, China), phenylmethanesulfonyl fluoride
(Beyotime Institute of Biotechnology) and phosphatase inhibitor
cocktail I (MedChemExpress, Monmouth Junction, NJ, USA). Protein
concentrations of each lysate were detected using the bicinchoninic
acid method (Beijing ComWin Biotech Co., Ltd., Beijing, China),
prior to each lysate being boiled at 95°C for 5 min. Equal
quantities (20 µg) of protein were separated by 10 or 12%
SDS-polyacrylamide gel and electrotransferred onto polyvinylidene
difluoride membranes (Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China). The membranes were blocked for 2 h with
5% skimmed milk in TBST at room temperature (Beijing Solarbio
Science & Technology Co., Ltd.), while the membranes
transferred with phosphorylated protein were blocked with 5% bovine
serum albumin (Beijing Solarbio Science & Technology Co., Ltd.)
in TBST at room temperature for 2 h. Each membrane was incubated
with primary antibodies (Abcam, Cambridge, UK; Table II) overnight at 4°C, washed three
times with TBST for 15 min each time, and incubated with secondary
antibody (cat. no. E-AB-1003; Elabscience Biotechnology Co., Ltd.,
Wuhan, China) for 1 h at room temperature. The secondary antibody
was horseradish peroxidase-conjugated goat-anti-rabbit IgG, and the
dilution was 1:2,000. Subsequently, the membranes were washed three
times with TBST for 15 min each time, and the immunoblots were
measured with a Fusion FX7 system (Vilber Lourmat, Marne-la-Vallée,
France) using a chemiluminescent horseradish peroxidase substrate
(EMD Millipore, Billerica, MA, USA). Relative expression of GAPDH
was calculated as an endogenous internal control for protein
loading.
| Table II.An overview of the primary
antibodies. |
Table II.
An overview of the primary
antibodies.
| Primary
antibody | Specificity | Dilution
factor | Catalog no. |
|---|
| Anti-GAPDH | Monoclonal
rabbit | 1:2,500 | ab9485 |
| Anti-c-Myc | Monoclonal
rabbit | 1:1,000 | ab32072 |
| Anti-c-Myc (phospho
T58) | Monoclonal
rabbit | 1:1,000 | ab185655 |
| Anti-Akt1 | Monoclonal
rabbit | 1:10,000 | ab179463 |
| Anti-Akt1 (phospho
S473) | Monoclonal
rabbit | 1:10,000 | ab81283 |
| Anti-GSK3β | Monoclonal
rabbit | 1:10,000 | ab32391 |
| Anti-GSK3β (phospho
S9) | Monoclonal
rabbit | 1:10,000 | ab75814 |
| Anti-Bcl-2 | Monoclonal
rabbit | 1:2,000 | ab182858 |
| Anti-Bax | Monoclonal
rabbit | 1:2,000 | ab32503 |
Statistical analysis
All experiments were performed at least three times.
Data were analyzed using GraphPad Prism 5.01 (GraphPad Software,
Inc., La Jolla, CA, USA) and are expressed as the mean ± standard
deviation of at least three separate experiments. One-way analysis
of variance (ANOVA) was used to compare the results of cell
viability and cell proliferation, as well as to compare different
time points and concentrations in Jurkat and Hut-78 cells. LSD test
was used following ANOVA. All other results were evaluated using an
independent-samples t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
DHA decreases the viability of Jurkat
and HuT-78 cells
The present study first evaluated the effects of DHA
on the proliferation of the T-cell lymphoma Jurkat and HuT-78 cell
lines using CCK-8 assays. Jurkat and HuT-78 cells were treated with
various concentrations of DHA for different time periods. Cell
morphology analysis revealed that treatment with 15 µM DHA in
Jurkat cells and 30 µM in HuT-78 cells damaged the integrity of the
cell membrane structure and induced a marked reduction in cell
density (Fig. 1A). The CCK-8 results
demonstrated that DHA decreased the viability of Jurkat and HuT-78
cells in a concentration- and time-dependent manner (Fig. 1B and C). Cell viability had
statistical significances in different time points/concentrations
in the Jurkat cell group (all P<0.001). Additionally, cell
viability had statistical significances (P<0.05) except at 36
and 48 h (P=0.53) in HuT 78 cell group. The IC50 values of DHA at
48 h for Jurkat and HuT-78 cells were 16.63 and 33.35 µM,
respectively.
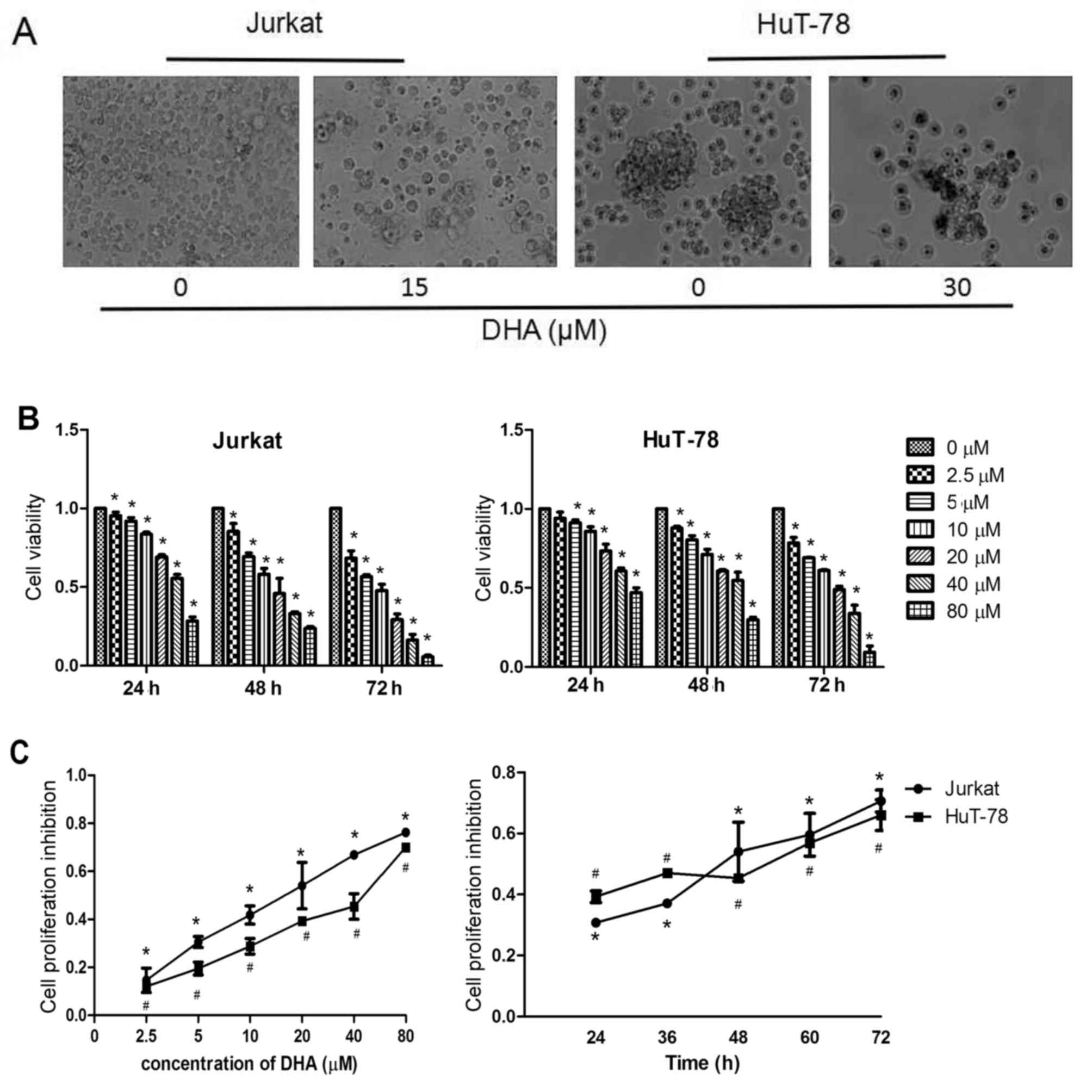 | Figure 1.DHA decreases the viability of Jurkat
and HuT-78 cells. (A) Representative images of Jurkat cells treated
with 0 or 15 µM DHA and HuT-78 cells treated with 0 or 30 µM DHA
for 48 h. (B) Cell viability of Jurkat and HuT-78 cells treated
with different concentrations of DHA for 24, 48 or 72 h.
*P<0.05, compared with the control group. (C) Cell viability of
Jurkat and HuT-78 cells treated with 0, 2.5, 5, 10, 20, 40 or 80 µM
DHA for 48 h. Cell viability of Jurkat (treated with 20 µM DHA) and
HuT-78 cells (treated with 40 µM DHA) after 24, 36, 48, 60 or 72 h.
DHA, dihydroartemisinin. #P<0.001 and *P<0.05,
compared with the control group. |
DHA promotes the apoptosis of Jurkat
and HuT-78 cells
Jurkat cells were treated with 15 µM DHA, and HuT-78
cells were treated with 30 µM DHA for 48 h, and apoptosis was
examined by Annexin V/PI staining. Flow cytometric analyses
demonstrated that 15 µM DHA significantly induced Jurkat cell
apoptosis (P<0.001). Compared with the control group, which
exhibited an apoptosis rate of 1.53%, the apoptosis rate of the DHA
treatment group was increased to 32.44%. The apoptosis rate of the
HuT-78 cells was also significantly increased to 22.81% following
administration of 30 µM DHA compared with 1.65% in the control
cells (P<0.001; Fig. 2A and
B).
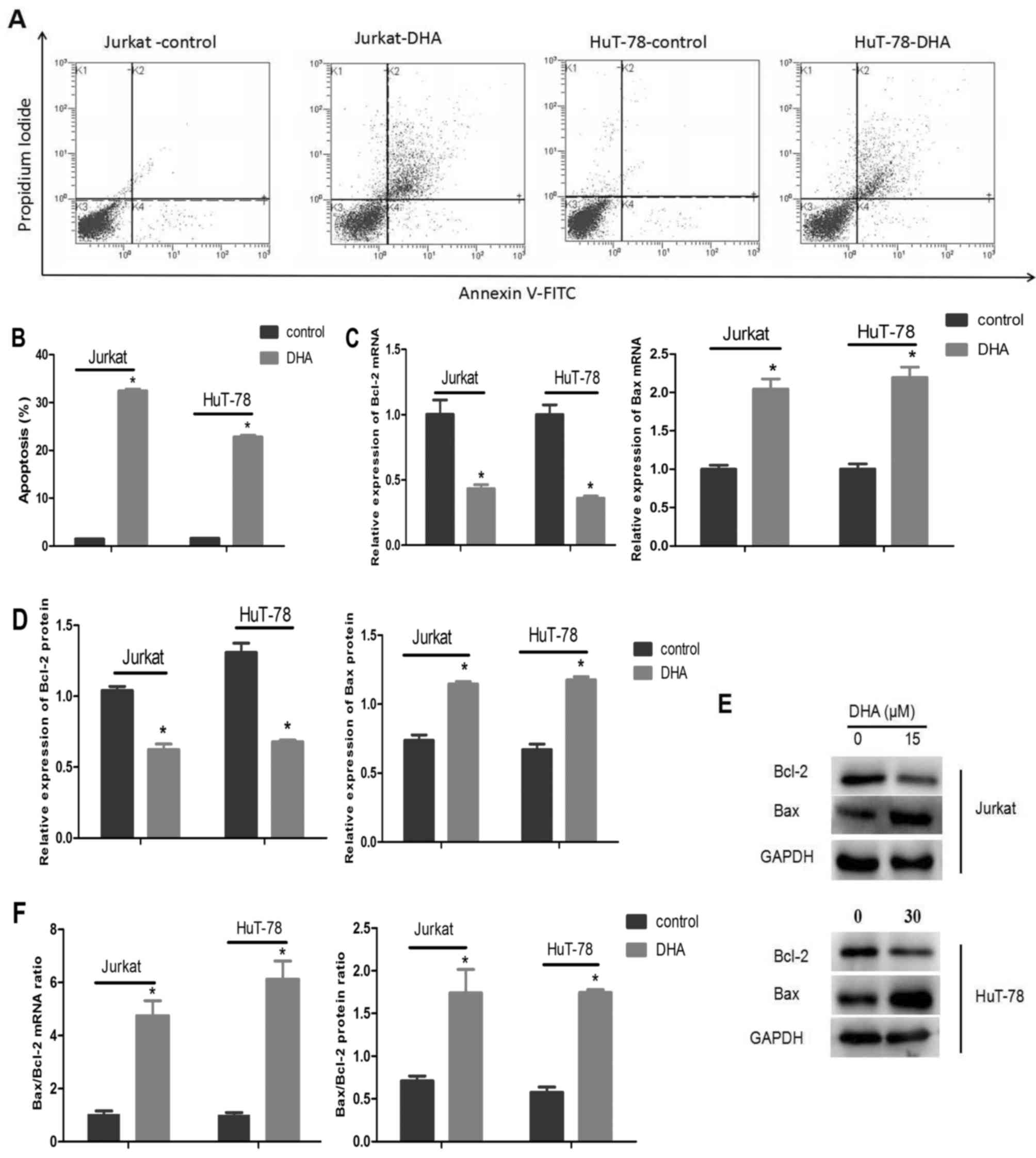 | Figure 2.DHA induces apoptosis in Jurkat and
HuT-78 cells. (A) Jurkat and HuT-78 cells were treated with 15 and
30 µM DHA, respectively, for 48 h and stained with Annexin
V-FITC/propidium iodide. (B) Data from A are presented as the mean
± standard deviation. *P<0.001, compared with the control group.
(C) The relative expression of Bcl-2 and Bax mRNA in Jurkat cells
treated with 0 or 15 µM and HuT-78 cells treated with 0 or 30 µM
DHA for 48 h. *P<0.001, compared with the control group. (D)
Quantification of Bcl-2 and Bax levels from immunoblots.
*P<0.001, compared with the control group. (E) Representative
immunoblots of Bcl-2 and Bax in Jurkat and HuT-78 cells treated
with 15 and 30 µM DHA, respectively, for 48 h. (F) The Bax/Bcl-2
mRNA and protein expression ratio in Jurkat and HuT-78 cells
treated with 15 and 30 µM DHA, respectively, for 48 h. *P<0.05,
compared with the control group. DHA, dihydroartemisinin; Bcl-2,
B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
The mRNA and protein expression levels of Bax and
Bcl-2 were also evaluated by RT-qPCR and western blot analysis. In
the two cell lines, Bax mRNA and protein expression was increased
while Bcl-2 mRNA and protein expression was significantly decreased
following exposure to DHA compared with the control group (Fig. 2C-E). In Jurkat cells, DHA induced a
4.72-fold increase in the ratio of Bax/Bcl-2 mRNA compared with the
control group and increased the ratio of Bax/Bcl-2 protein
expression by over 2-fold. In HuT-78 cells treated with DHA, the
ratio of Bax/Bcl-2 mRNA and protein expression increased by
6.08-fold and 3.39-fold, respectively, compared with the control
group (Fig. 2F).
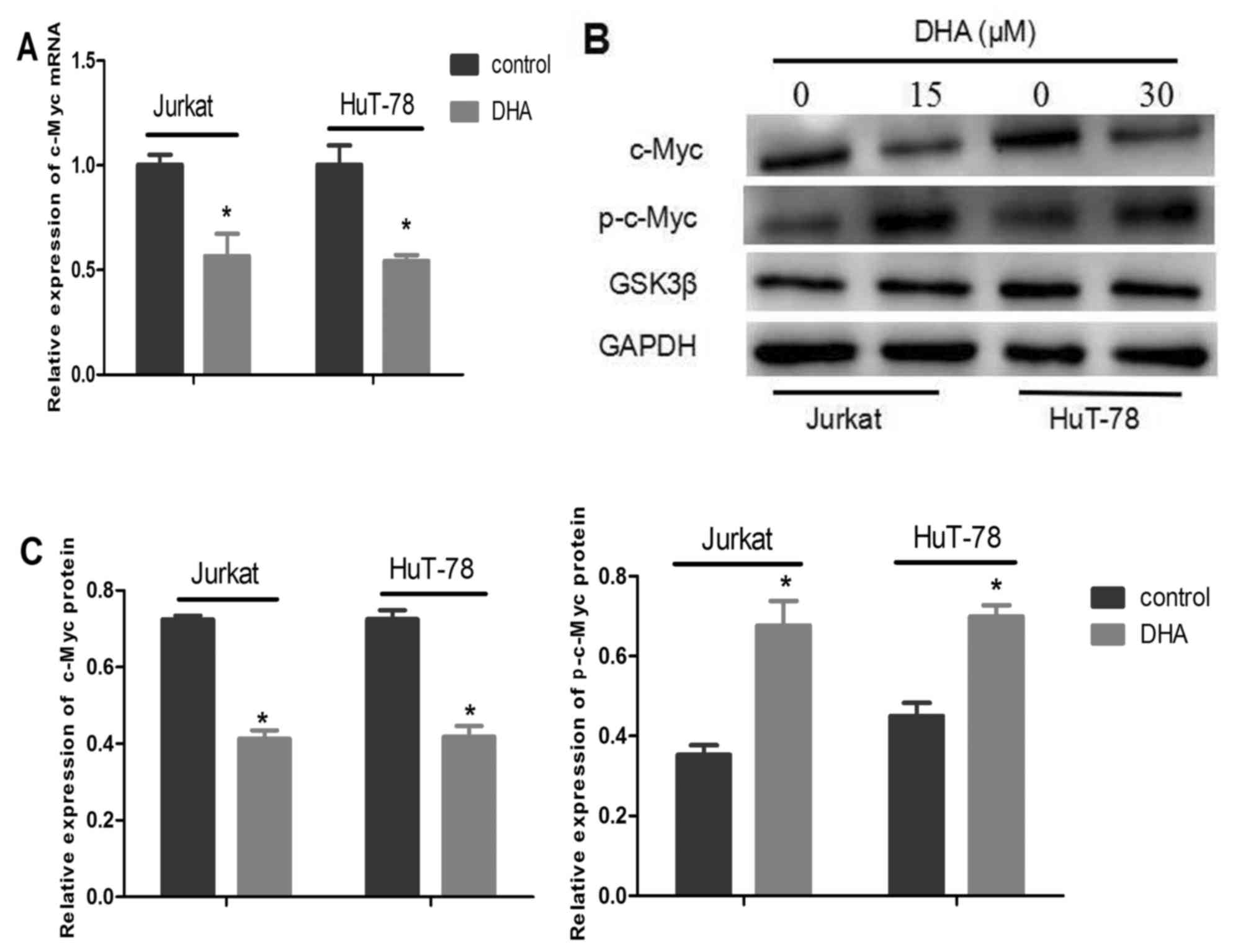
DHA enhances c-Myc phosphorylation at
threonine 58 in Jurkat and HuT-78 cells
To determine whether DHA exerted any effect on c-Myc
mRNA levels, the relative expression of c-Myc mRNA was detected by
RT-qPCR (Fig. 3A). It was revealed
that c-Myc mRNA expression was downregulated in Jurkat cells
treated with 15 µM DHA and HuT-78 cells treated with 30 µM DHA. The
differences were statistically significant compared with the
control groups (P<0.001).
To determine whether DHA exhibited any effect on
c-Myc protein levels, c-Myc protein and c-Myc phosphorylated at
threonine 58 expression levels were determined by western blot
analysis. The expression level of phosphorylated c-Myc at threonine
58 was increased following DHA treatment for 2 h, while the total
relative expression of c-Myc oncoprotein was markedly decreased in
Jurkat and HuT-78 cells following treatment with DHA for 48 h
(Fig. 3B and C). The differences in
expression levels were significant compared with the control groups
(P<0.05). DHA did not have a significant effect on the
expression level of GSK3β protein (P>0.05; Fig. 3B).
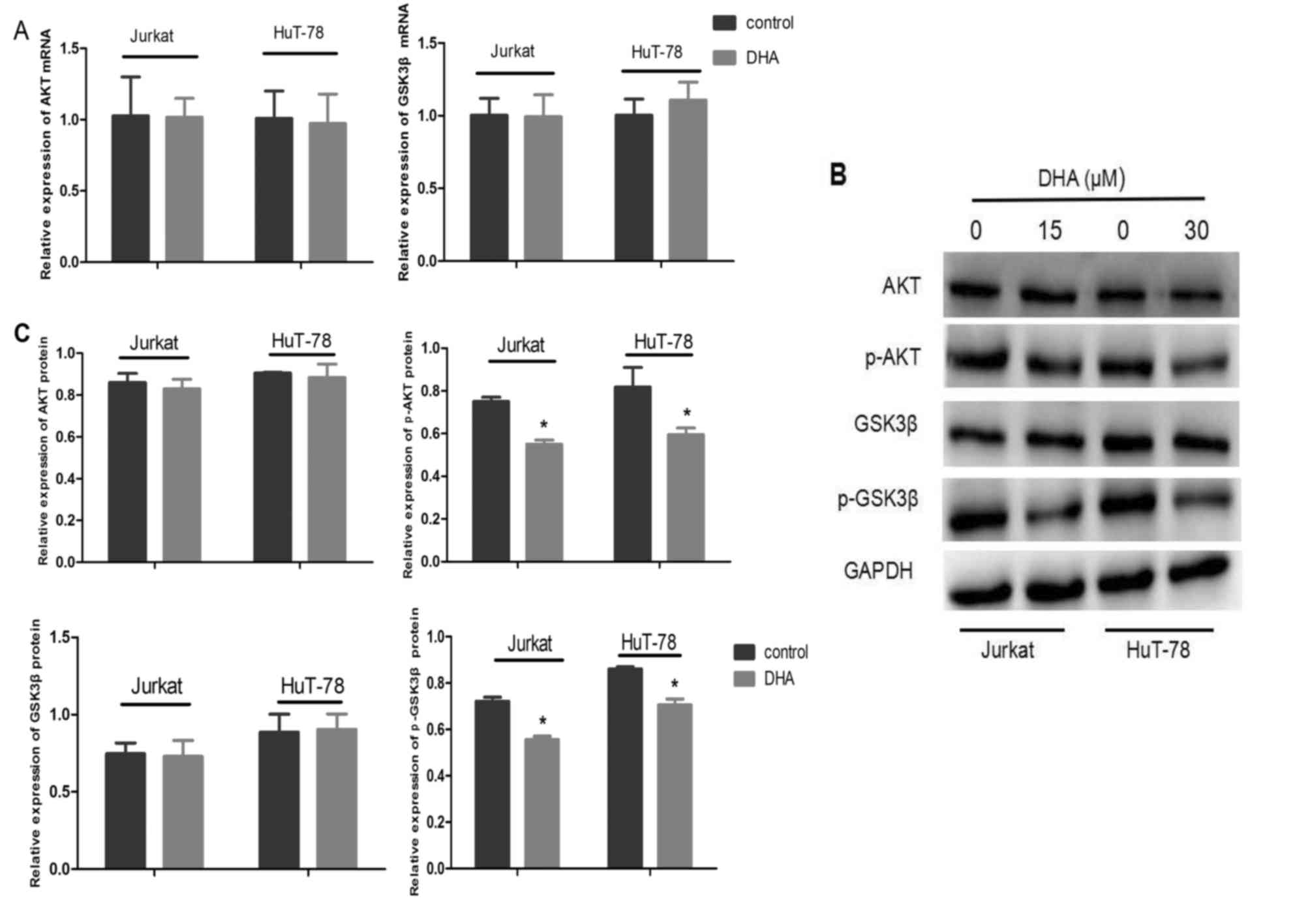
DHA decreases p-Akt and p-GSK3β
expression in Jurkat and HuT-78 cells
To detect the effect of DHA on the Akt/GSK3β
signaling pathway in Jurkat and HuT-78 cells, Akt and GSK3β mRNA
expression levels were examined by RT-qPCR, and Akt, Akt
phosphorylated at serine 473, GSK3β and GSK3β phosphorylated at
serine 9 protein expression levels were examined by western blot
analysis (Fig. 4A-C). DHA had no
effect on the level of Akt and GSK3β mRNA expression or the total
protein expression of Akt and GSK3β (Fig.
4A-C). However, DHA treatment for 6 h resulted in decreased
levels of Akt phosphorylated at serine 473 and GSK3β phosphorylated
at serine 9 (P<0.001; Fig. 4B and
C).
Discussion
T-cell lymphoma is a common hematological
malignancy, and its occurrence is associated with alterations in
critical genes and activation of pivotal signaling pathways
(5). High expression of c-Myc is
associated with the occurrence and development of lymphoma
(7). In addition, the Akt/GSK3β
signaling pathway serves a critical role in the regulation of tumor
cellular processes, including proliferation, differentiation and
apoptosis (7,9). DHA is an effective drug in treating
patients with malaria (22). Recent
studies have revealed that DHA has antitumor activity with few side
effects in various types of tumors in vivo and in
vitro (16–18,23–25). Based
on the results of these aforementioned studies, the present study
investigated the therapeutic role of DHA and its underlying
mechanism in T-cell lymphoma cells.
In the present study, DHA inhibited the
proliferation of T-cell lymphoma cells in a concentration- and
time-dependent manner. The results of the present study were
consistent with those of previous studies in Jurkat and ovarian
cancer cells (16,18). When different concentrations of DHA
were compared, it was revealed that Jurkat cells were more
sensitive to DHA than HuT-78 cells. These results indicated that
DHA may be a promising drug for treatment in T-lymphoma cells.
However, the effect of DHA on the patient samples and xenograft
mouse model in T-cell lymphoma were not assessed in the present
study; therefore, we will further investigate the effect of DHA in
future in vivo studies.
The results of the present study demonstrated that
DHA reduced c-Myc protein expression by two potential mechanisms.
To begin with, DHA treatment caused a decrease in the c-Myc mRNA
level, resulting in a decreased c-Myc protein expression.
Additionally, DHA enhanced the phosphorylation of c-Myc at
threonine 58, which may trigger c-Myc oncoprotein degradation in
T-cell lymphoma cells by the ubiquitin proteasome system. A
previous study revealed that DHA promoted c-Myc oncoprotein
degradation in HL-60 and HCT116 cells (19). This suggested that DHA-induced
phosphorylation of c-Myc at threonine 58 targets c-Myc for
degradation. Taken together, the results of the present study
suggested that DHA suppressed c-Myc protein expression at the
transcriptional and post-transcriptional level, leading to
inhibition of cell proliferation and induction of cell
apoptosis.
The Akt/GSK3β signaling pathway serves an important
role in the regulation of cell proliferation and apoptosis in
cancer. The Akt/GSK3β signaling pathway is activated upon
phosphorylation of Akt and GSK3β at serine 473 and serine 9,
respectively, and promotes proliferation and inhibits apoptosis.
Previous studies have demonstrated that DHA could inhibit the
Akt/GSK3β signaling pathway in lung cancer and glioma cells
(23,25). To further elucidate the mechanism of
DHA treatment in T-cell lymphoma cells, the present study
investigated the effect of DHA on the Akt/GSK3β signaling pathway.
The results of the present study revealed that DHA could suppress
the Akt/GSK3β signaling pathway through a process that involved
reduction of p-Akt and p-GSK3β expression levels, but did not
impact GSK3β steady-state levels.
The Bcl-2 protein is an anti-apoptotic factor that
serves a critical role in apoptosis, whereas Bax functions as a
pro-apoptotic effector (26). The
results of the present study demonstrated that DHA-induced
apoptosis was accompanied by the downregulation of anti-apoptotic
Bcl-2 expression and the upregulation of pro-apoptotic Bax
expression, suggesting that DHA exerts its effects of inducing
apoptosis by modulating Bax/Bcl-2 expression levels in T-cell
lymphoma cells.
Previous studies have demonstrated that Bcl-2 could
abrogate c-Myc-induced apoptosis (27), and that c-Myc may synergize with Akt
in cell growth and proliferation (28). These studies indicated that c-Myc
cooperates with Akt and Bcl-2 to promote tumor progression. These
results combined with those of the present study further suggested
that the DHA-induced apoptotic effect may also involve DHA reducing
Bcl-2 expression and relieving its inhibition of c-Myc. DHA also
reduces the expression of c-Myc and p-Akt, leading to the
suppression of Jurkat and HuT-78 cell growth and proliferation.
In conclusion, the results of the present study
suggested that DHA decreased c-Myc protein expression levels at the
transcriptional level and triggered c-Myc degradation by enhancing
the phosphorylation of c-Myc at threonine 58 to inhibit cell
proliferation and increase cell apoptosis. Furthermore, DHA blocked
cell proliferation by suppressing the Akt/GSK3β signaling pathway
and reducing c-Myc protein expression, and DHA also induced
apoptosis by increasing the ratio of Bax/Bcl-2 in T-cell lymphoma
cells. The results of the present study aid in clarifying the
mechanism of DHA antitumor activity in T-cell lymphoma cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Plan of Traditional Chinese Medicine Development
Projects, Shandong, China (2017–197).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SW, WW, XZ, CZ and HZ conceived and designed the
experiments. WW, XZ, LS and YC performed the experiments and
acquired the data. LS analyzed and interpreted the data regarding
the cell viability assay. YC analyzed and interpreted the data
regarding the cell apoptosis assay. SW, XZ and WW analyzed and
interpreted the data regarding RT-qPCR and western blot analysis.
SW, WW and XZ drafted the manuscript. SW, CZ and HZ revised the
manuscript critically for important intellectual content.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declared that they have no conflict of
interest.
References
|
1
|
Bassig BA, Au WY, Mang O, Ngan R, Morton
LM, Ip DK, Hu W, Zheng T, Seow WJ, Xu J, et al: Subtype-specific
incidence rates of lymphoid malignancies in Hong Kong compared to
the United States, 2001–2010. Cancer Epidemiol. 42:15–23. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Uckun FM, Gaynon PS, Sensel MG, Nachman J,
Trigg ME, Steinherz PG, Hutchinson R, Bostrom BC, Sather HN and
Reaman GH: Clinical features and treatment outcome of childhood
T-lineage acute lymphoblastic leukemia according to the apparent
maturational stage of T-lineage leukemic blasts: A children's
cancer group study. J Clin Oncol. 15:2214–2221. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aifantis I, Raetz E and Buonamici S:
Molecular pathogenesis of T-cell leukaemia and lymphoma. Nat Rev
Immunol. 8:380–390. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Advani RH, Ansell SM, Lechowicz MJ, Beaven
AW, Loberiza F, Carson KR, Evens AM, Foss F, Horwitz S, Pro B, et
al: A phase II study of cyclophosphamide, etoposide, vincristine
and prednisone (CEOP) Alternating with Pralatrexate (P) as front
line therapy for patients with peripheral T-cell lymphoma (PTCL):
Final results from the T-cell consortium trial. Brit J Haematol.
172:535–544. 2016. View Article : Google Scholar
|
|
5
|
Kwong YL: T-cell Lymphoma forum. Exp Rev
Anticancer Ther. 10:493–498. 2010. View Article : Google Scholar
|
|
6
|
Pelengaris S, Khan M and Evan G: c-MYC:
More than just a matter of life and death. Nat Rev Cancer.
2:764–776. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Malempati S, Tibbitts D, Cunningham M,
Akkari Y, Olson S, Fan G and Sears RC: Aberrant stabilization of
c-Myc protein in some lymphoblastic leukemias. Leukemia.
20:1572–1581. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gregory MA, Qi Y and Hann SR:
Phosphorylation by glycogen synthase kinase-3 controls c-Myc
proteolysis and subnuclear localization. J Biol Chem.
278:51606–51612. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pap M and Cooper GM: Role of glycogen
synthase kinase-3 in the phosphatidylinositol 3-kinase/Akt cell
survival pathway. J Biol Chem. 273:19929–19932. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hetman M, Cavanaugh JE, Kimelman D and Xia
Z: Role of glycogen synthase kinase-3 beta in neuronal apoptosis
induced by trophic withdrawal. J Neurosci. 20:2567–2574. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shankar SL, Krupski M, Parashar B, Okwuaka
C, O'Guin K, Mani S and Shafit-Zagardo B: UCN-01 alters
phosphorylation of Akt and GSK3beta and induces apoptosis in six
independent human neuroblastoma cell lines. J Neurochem.
90:702–711. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Querfeld C, Rizvi MA, Kuzel TM, Guitart J,
Rademaker A, Sabharwal SS, Krett NL and Rosen ST: The selective
protein kinase C beta inhibitor enzastaurin induces apoptosis in
cutaneous T-cell lymphoma cell lines through the AKT pathway. J
Invest Dermatol. 126:1641–1647. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang L, Wu SN and Xing D: High fluence
low-power laser irradiation induces apoptosis via inactivation of
Akt/GSK3β signaling pathway. J Cell Physiol. 226:588–601. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cai Q, Deng H, Xie D, Lin TX and Lin TY:
Phosphorylated AKT protein is overexpressed in human peripheral
T-cell lymphomas and predicts decreased patient survival. Clin
Lymph Myeloma Leuk. 12:106–112. 2012. View Article : Google Scholar
|
|
15
|
Cicenas J: The potential role of Akt
phosphorylation in human cancers. Int J Biol Marker. 23:1–9. 2008.
View Article : Google Scholar
|
|
16
|
Wang QY, Wu S, Zhao X, Zhao C, Zhao H and
Huo L: Mechanisms of dihydroartemisinin and
dihydroartemisinin/holotransferrin cytotoxicity in T-cell lymphoma
cells. PLoS One. 10:e01373312015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang CZ, Zhang H, Yun J, Chen GG and Lai
PB: Dihydroartemisinin exhibits antitumor activity toward
hepatocellular carcinoma in vitro and in vivo. Biochem Pharmacol.
83:1278–1289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Greenshields AL, Shepherd TG and Hoskin
DW: Contribution of reactive oxygen species to ovarian cancer cell
growth arrest and killing by the anti-malarial drug artesunate. Mol
Carcinogen. 56:75–93. 2017. View
Article : Google Scholar
|
|
19
|
Lu JJ, Meng LH, Shankavaram UT, Zhu CH,
Tong LJ, Chen G, Lin LP, Weinstein JN and Ding J:
Dihydroartemisinin accelerates c-MYC oncoprotein degradation and
induces apoptosis in c-MYC-overexpressing tumor cells. Biochem
Pharmacol. 80:22–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liao K, Li J and Wang Z:
Dihydroartemisinin inhibits cell proliferation via
AKT/GSK3β/cyclinD1 pathway and induces apoptosis in A549 lung
cancer cells. Int J Clin Exp Pathol. 7:8684–8691. 2014.PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta CT) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mohamed AO, Hamid Abdel MM, Mohamed OS,
Elkando NS, Suliman A, Adam MA, Elnour FAA and Malik EM: Efficacies
of DHA-PPQ and AS/SP in patients with uncomplicated Plasmodium
falciparum malaria in an area of an unstable seasonal transmission
in Sudan. Malar J. 16:1632017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Du W, Pang C, Xue Y, Zhang Q and Wei X:
Dihydroartemisinin inhibits the Raf/ERK/MEK and PI3K/AKT pathways
in glioma cells. Oncol Lett. 10:3266–3270. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hou J, Wang D, Zhang R and Wang H:
Experimental therapy of hepatoma with artemisinin and its
derivatives: In vitro and in vivo activity, chemosensitization, and
mechanisms of action. Clin Cancer Res. 14:5519–5530. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen T, Li M, Zhang R and Wang H:
Dihydroartemisinin induces apoptosis and sensitizes human ovarian
cancer cells to carboplatin therapy. J Cell Mol Med. 13:1358–1370.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heath-Engel HM, Chang NC and Shore GC: The
endoplasmic reticulum in apoptosis and autophagy: Role of the BCL-2
protein family. Oncogene. 27:6419–6433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fanidi A, Harrington EA and Evan GI:
Cooperative interaction between c-myc and bcl-2 proto-oncogenes.
Nature. 359:554–556. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yeh ES, Belka GK, Vernon AE, Chen CC, Jung
JJ and Chodosh LA: Hunk negatively regulates c-myc to promote
Akt-mediated cell survival and mammary tumorigenesis induced by
loss of Pten. Proc Natl Acad Sci USA. 110:6103–6108. 2013.
View Article : Google Scholar : PubMed/NCBI
|