Introduction
Bronchial asthma is one of the most common chronic
diseases throughout the world and its rate of prevalence has
continuously increased over the last decades (1). During the asthmatic process, epithelial
damage is followed by the thickening of asthmatic bronchial walls,
which is mechanistically linked to the local inflammation and the
activation of bronchial fibroblasts (2–4). Local
inflammation induces the differentiation of bronchial fibroblasts
into α-smooth muscle actin (α-SMA)positive
myofibroblasts and increases their longevity (5) through paracrine (for example
TGFβ-dependent) and cell adhesion-dependent mechanisms (3,6,7). A high extracellular matrix-producing
activity and contractility of myofibroblasts (8) participates in bronchial fibrosis,
fibrotic remodeling of airways and lung parenchymal compartments,
and a pulmonary dysfunction. The asthmatic process may also
facilitate the progression of other chronic lung diseases.
Importantly, epidemiologic data show the correlation between the
incidence of asthma and lung cancer in developed countries
(9).
Chronic inflammation and tissue fibrosis have long
been considered to induce phenotypic shifts in cancer cells, such
as epithelial-mesenchymal transition (EMT), which accounts for the
formation of cancer invasive front. In particular,
cancer-associated fibroblasts (CAFs) participate in cancer
progression and metastasis (10,11).
During lung cancer progression, the connective tissue of bronchial
walls provides the scaffold for developing tumors, forms tissue
barriers against immune responses of the host and/or constitutes
the routes for the invasion of cancer cells (12). Accordingly, the activation of
fibroblasts/myofibroblasts, which is characteristic for the
asthmatic process, may facilitate lung cancer progression. However,
the contribution of asthmatic bronchial fibroblasts in systemic
dissemination of lung cancer cells remains unaddressed. We have
previously shown that human bronchial fibroblasts (HBFs) derived
from asthmatic patients (AS HBFs) display relatively high
susceptibility to TGFβ-inducible fibroblast-myofibroblast
transition [FMT; (13–15)]. These heritable pro-fibrotic
properties of AS HBFs prompted us to hypothesize that AS HBFs may
contribute to lung carcinogenesis. To elucidate the links between
their function and the progression of lung cancer, we compared the
behavior of human non-small cell lung cancer A549 cells exposed to
the signals from monolayers of AS HBFs and from HBFs derived from
non-asthmatic (NA) donors.
Materials and methods
Isolation and propagation of primary
HBFs
Primary HBFs were isolated from bronchial biopsies
derived from i) the individuals in whom diagnostic bronchoscopy
ruled out any serious airway pathology, i.e., asthma, chronic
inflammatory lung disease or cancer (NA group; N=5, averaged age,
57.4±9.9 years; FEV1(%)=101.83±14.7) and ii) from the patients with
moderate asthma (AS group; N=5; age, 39±17.6 years; mean duration
of asthma=16.7±12.2 years; FEV1(%)=48.75±0.86). Patients were
treated in the Department of Medicine of Jagiellonian University
(Kraków, Poland) and were in a stable clinical condition. AS HBF
lineages were derived from a 38 years old man/66 years old woman
(asthma duration=10/30 years; FEV1(%)=35/52.5). These lineages
displayed a considerably higher pro-fibrotic potential than their
non-asthmatic counterparts; however, no relationship between their
phenotypic properties and the age/sex of the donors, or with the
development of asthma severity was observed (6,13–15). The present study was approved by the
Jagiellonian University's Ethics Committee (decision no.
122.6120.69.2015) on the basis of written informed consent for the
use of HBF cell lines in basic research, which had previously been
obtained from all donors. For endpoint experiments, HBFs were
harvested between the 5th and 15th passage after the establishment
of primary cultures.
Co-cultures of A549 cells with
HBFs
HBFs were propagated in DMEM supplemented with a 10%
FCS and antibiotic-antimycotic cocktail (both Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) whereas human lung carcinoma A549 cells
(ECACC 86012804) were cultured in RPMI-1640 medium (Sigma-Aldrich;
Merck KGaA) with the same supplements. Cells were incubated in a
humidified atmosphere with 5% CO2 at 37°C to reach 80%
confluence. For ‘open’ co-cultures with HBFs, A549 cells were
seeded on coverslips at the density of 104
cells/cm2 together with HBFs (seeded at 103
cells/cm2) and incubated for 48 h in DMEM supplemented
with a 10% FCS. Alternatively, A549 and HBFs (NA or AS) were seeded
into the chambers of Ibidi µ-Slide 2 Well Co-Culture dish (81806;
Ibidi GmbH, Munich, Germany) for 48 h in 2 ml of the same medium
(i.e., in the conditions allowing only for exchange of the medium
between the chambers; ‘isolated’ co-cultures). Furthermore, A549
cells and HBFs were seeded in the chambers of 2 well silicone
inserts with a defined cell free gap (81176; Ibidi GmbH) cultured
for 6 h and the next 42 h after the removal of silicone insert
(‘confronted’ co-cultures). Media conditioned by HBFs and 3
variants of HBF/A549 co-cultures were collected and kept in −80°C
for endpoint experiments. For the estimation of the effect of
A549/HBF ‘secretomes’ on A549 motility/transmigration, the cells
were incubated in the presence of the mixture of relevant
conditioned medium/fresh medium (1:1 v/v). For 2D invasion assays
A549 cells and HBFs were seeded in the chambers of 2-well silicone
inserts (81176; Ibidi GmbH). The cells were cultured to monolayers
(48 h) and allowed to migrate for the next 48 h after removal of
the silicone insert. Then, the cells were fixed and stained (see
below) to estimate the effect of paracrine/juxtacrine signaling
between A549 cells and HBFs on A549 invasive front formation.
Cell migration and transmigration
tests
Movement of A549 cells in co-cultures with HBFs and
in the conditioned media was registered using Leica DMI6000B
time-lapse videomicroscopy system (Leica Microsystems GmbH,
Wetzlar, Germany) equipped with a temperature/CO2
chamber and Integrated Modulation Contrast (IMC) optics. Sequences
of cell centroid positions were recorded at 300 s time intervals
for 6 h using a dry 20×, NA-0.75 objective (16). The averaged total length of cell
trajectory (i.e., the ‘Distance’ covered by the cells during the
registration time; µm) and the total length of cell displacement
(‘Displacement’, i.e., the distance from the starting point
directly to the cell's final position; µm) were quantified from
cell trajectories with the purpose-designed Hiro program. The
averaged velocity of cell motility (VCM) and velocity of cell
displacement (VCD) were defined as the averaged ‘Distance’/time of
registration and averaged ‘Displacement’/time of registration,
respectively. Cell trajectories (N>50) from no less than three
independent experiments (N>3) were taken for the estimation of
statistical significance. For transmigration assays, A549 cells
were seeded into chambers containing microporous membranes (pore
diameter 8 µm; membrane diameter 6.5 mm; 2422; Corning, NY, USA) at
the density of 300 cells/mm2. The inserts were placed in
the wells of 24-well plates filled with control, HBF and
HBF/A549-conditioned media (1:1 v/v with a fresh medium) and the
cells were allowed to transmigrate for 48 h. The numbers of the
transmigrated cells were determined with Coulter counter after the
next 48 h (17).
Immunofluorescence and
cytofluorimetry
For the immunofluorescence analyses of
Snail-1/α-SMA/Cx43, the cells were fixed in MetOH/Acetone (7:3)
solution for 10 min. in −20°C. The fixed cells were incubated in 3%
BSA solution and incubated with the primary antibodies (mouse
anti-α-SMA IgG; Sigma-Aldrich; Merck KGaA; A2547; 1:300); rabbit
anti-Snail-1 antibody (SAB 4504319; 1:100) and mouse anti-Cx43 IgM
(both Sigma-Aldrich; Merck KGaA; C8093; 1:300) for 1 h. Then, the
cells were incubated with the secondary antibody [Alexa
Fluor® 647-conjugated goat anti-mouse IgG (A21235),
Alexa Fluor® 488-conjugated goat anti-rabbit IgG
(A11008) or Alexa Fluor® 546-conjugated goat anti-mouse
IgM (A21045); all 1:500; Thermo Fisher Scientific, Inc., Waltham,
MA, USA]. Image acquisition was performed with Leica DMI6000B
microscope (DMI7000 version; Leica Microsystems GmbH) equipped with
the Nomarski Interference Contrast (DIC) module. Cytofluorimetric
analyses were performed with LasX software (Leica Microsystems
GmbH) and ImageJ freeware (National Institutes of Health, Bethesda,
MD, USA), respectively (17) to
generate histograms that show the co-localization of
immunofluorescence signals along the scan line.
Statistical analysis
Statistical analysis was performed using one-way
analysis of variance followed by Tukey's honestly significant
difference post-hoc and Statistica Data Miner Software version 13
(StatSoft, Inc., Tulsa, OK, USA). Data are expressed as the mean ±
standard error of the mean. *P<0.05 was considered to indicate a
statistically significant difference.
Results
AS HBFs stimulate the invasive
behavior of A549 cells
To estimate how asthmatic bronchial microenvironment
participates in lung cancer progression, we used an experimental
approach based on the ‘open’ co-cultures of non-small cell lung
cancer (A549) cells with bronchial fibroblasts derived from 2
asthmatic patients (AS HBFs). An activation of A549 cells in the
proximity of both AS HBF lineages was illustrated by the induction
of their motility in the co-cultures (Fig. 1A). Interestingly, AS1 HBFs activated
A549 cell locomotion without any considerable effect on its
efficiency (the cell displacement), whereas the increased velocity
of A549 displacement was observed in the proximity of AS2 HBFs
(Fig. 1C; cf. Table I). Differences in A549 cell reactions
to the signals from the analyzed AS HBF lineages were accompanied
by different behavior of AS and NA HBFs in the proximity of A549
cells. AS HBFs of both lineages were dispersed among A549 cells,
whereas NA HBFs formed compact multicellular clusters (Fig. 1B) and only slightly affected the
motility of A549 cells (Fig. 1D).
Pictures in Fig. 1B illustrate the
morphology of NA and AS HBFs in confluent co-cultures with A549
cells. Some differences in the HBF density result from more
efficient spreading of the fibroblasts from AS group. These
observations suggest that HBFs from asthmatic bronchi can
differentially activate lung cancer cells via paracrine and/or
juxtacrine signaling pathways.
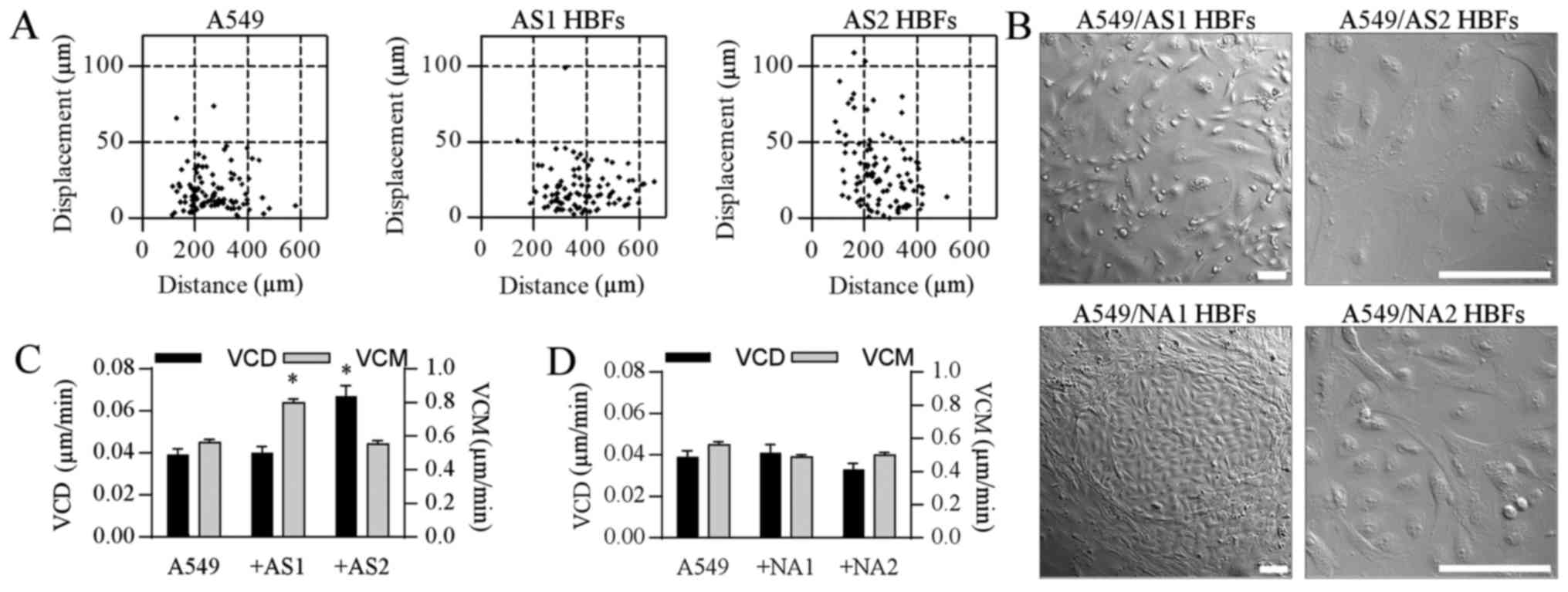 | Figure 1.HBFs stimulate the invasive behavior
of A549 cells. (A) A549 cells and AS HBFs were seeded at a density
of 104 and 103 cells/cm2,
respectively, and were incubated for 48 h in Dulbecco's modified
Eagle's medium supplemented with a 10% fetal calf serum. The
motility of A549 cells was estimated with time-lapse
videomicroscopy and (C) its parameters were quantified in
comparison to control A549 cells. (B) A549/HBF co-cultures were
established as in (A) and cell morphology was visualized with NIC
microscopy. Scale bars, 100 µm. (D) The motility of A549 cells in
co-cultures with NA HBFs estimated by time-lapse videomicroscopy as
in (A). At least 50 cell trajectories were drawn for each condition
and presented in correlative plots. Dot-plots and column charts
present movement parameters at the single cell and population
level, respectively. Data are representative of 3 independent
experiments. *P<0.05 vs. A549. Data are presented as the mean ±
standard error of the mean. Note that the disruption and
penetration of A549 cell monolayers by AS HBFs is correlated with
the induction of A549 motility. HBFs, human bronchial fibroblasts;
AS, asthmatic donors; NA, non-asthmatic donors; VCM, velocity of
cell motility; VCD, velocity of cell displacement; NIC, Nomarski
Interference Contrast. |
 | Table I.Summary of the quantitative data
evaluating the effect of HBFs on A549 cell motility. |
Table I.
Summary of the quantitative data
evaluating the effect of HBFs on A549 cell motility.
| Variant | Distance ± SEM
(µm) | Displacement ± SEM
(µm/min) | CME ± SEM (%) |
|---|
| Control (A549) | 268.797±9.291 | 18.611±1.376 | 6.880±0.723 |
| +CM HBFs |
|
|
|
|
NA1 |
447.073±13.836a | 28.226±2.796 | 7.582±0.900 |
|
NA2 |
417.337±13.776a | 20.420±3.197a | 5.632±0.844 |
|
AS1 | 325.099±9.794 |
34.768±2.932a | 13.805±1.539a |
|
AS2 |
584.146±17.543a | 21.884±2.074 | 5.010±0.768 |
| +CM HBFs
isolated |
|
|
|
|
NA1 | 316.663±11.869 | 27.537±2.269 | 11.206±1.205 |
|
NA2 |
380.407±14.506a | 24.401±2.031 | 8.418±0.947 |
|
AS1 |
416.148±12.914a | 36.541±2.694a | 10.534±1.041 |
|
AS2 |
422.320±16.828a | 41.202±3.436a | 12.924±1.446 |
| +CM A549/HBFs
CONFRONTED |
|
|
|
|
NA1 | 325.500±11.423 | 22.790±2.187 | 7.905±0.851 |
|
NA2 |
463.809±15.997a | 25.114±2.320 | 6.997±0.853 |
|
AS1 |
387.730±14.364a |
44.422±3.688a |
15.099±1.617a |
|
AS2 |
435.521±12.779a |
33.665±3.002a | 9.635±1.177 |
| A549/HBFs
CO-culture |
|
|
|
|
NA1 | 234.485±6.225 | 19.744±1.888 | 9.666±1.117 |
|
NA2 | 239.516±8.294 | 15.692±1.574 | 7.505±0.767 |
|
AS1 |
383.326±10.824a | 19.198±1.381 | 5.556±0.528 |
|
AS2 | 265.195±9.993 |
32.397±2.353a |
15.779±1.665a |
AS HBFs induce the motility of A549
cells via contact-modulated paracrine signaling
Numerous signaling systems have been implicated in
the regulation of cancer cell motility by CAFs (18). In our hands, media conditioned by both
analyzed AS HBF lineages augmented the motility of A549 cells
(Fig. 2A). A549 displaced more
efficiently in the presence of AS1 HBF-conditioned medium, whereas
AS2 HBF secretome had no significant effects on the efficiency of
A549 displacement, even though it induced the movement of A549
cells (Fig. 2C; Table I). An induction of A549 displacement
was also observed in the presence of the media conditioned by AS
HBF/A549 cells co-cultured in the conditions allowing for their
mutual paracrine (i.e., in ‘isolated’ co-cultures) and
paracrine/juxtacrine interactions (i.e., in ‘confronted’
co-cultures). Generally, this effect was stronger than in the
presence of the media conditioned by AS HBFs and ‘open’ AS HBF/A549
co-cultures; however certain differences were seen in A549
responses to AS1 HBF/A549 and AS2 HBF/A549 secretome (Fig. 2C). We also observed nuclear
accumulation of Snail-1 and Cx43 up-regulation in A549 cells
cultured in the media from the ‘isolated’ co-cultures (Fig. 2B). On the other hand, NA HBF/A549
secretomes displayed a considerably lower pro-invasive activity
regardless of the culture conditions (Fig. 2D; Table
I). Collectively, these data show that paracrine/juxtacrine
communication between A549 cells and AS HBFs affects their
secretome, thus regulating A549 invasive behavior.
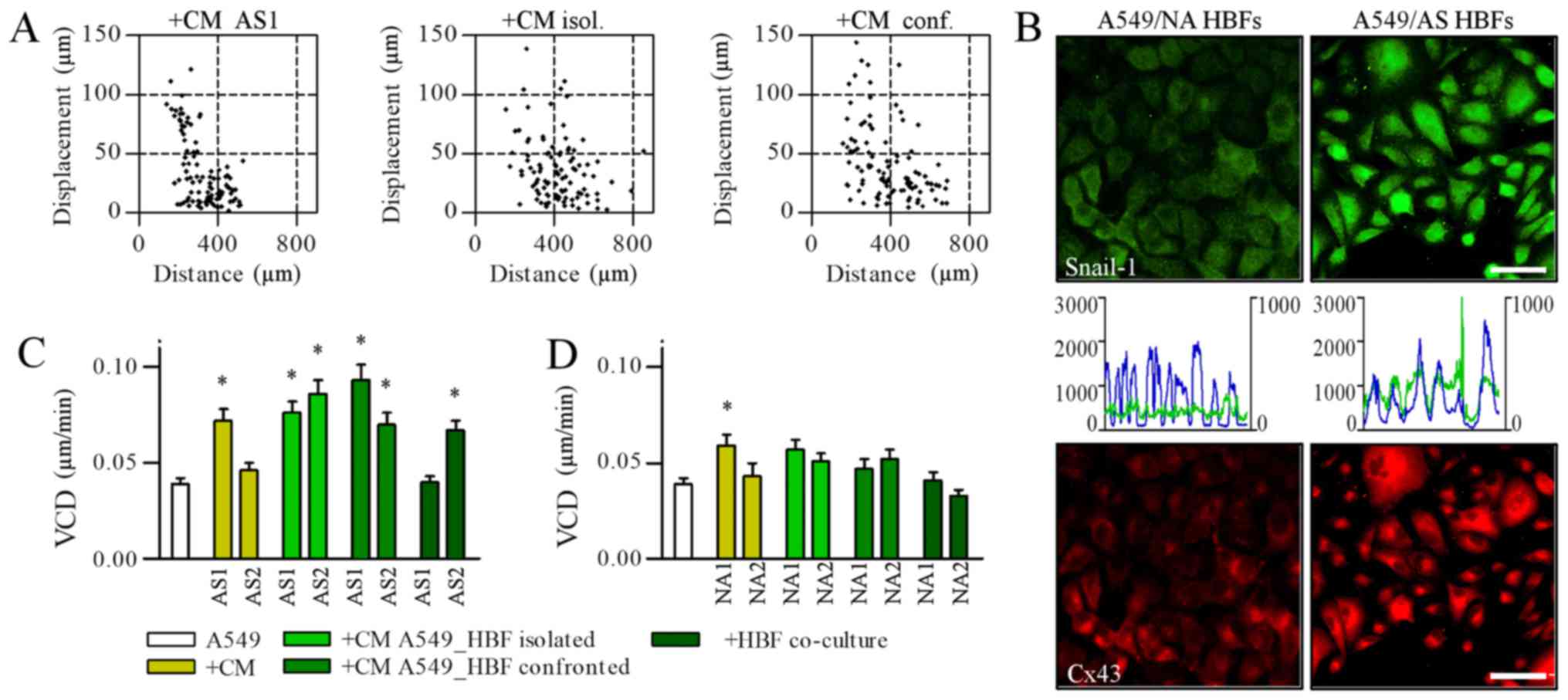 | Figure 2.AS HBFs induce the motility of A549
cells via contact-modulated paracrine signaling. (A) A549 cells
were cultivated in the media conditioned by AS2 HBFs (left),
‘separated’ (middle) and ‘confronted’ AS HBF/A549 co-cultures (1:1
v/v with fresh medium) for 48 h and (C) the parameters of their
motility were analyzed by time-lapse videomicroscopy in comparison
to A549 motility in control conditions and in ‘open’ AS HBF/A549
co-cultures. (B) A549 cells were cultivated in the media
conditioned by ‘separated’ co-cultures of A549 with AS and NA HBFs
(1:1 v/v with fresh medium) for 48 h. Intracellular localization of
Snail-1/Cx43 and co-localization of Snail-1/DNA was visualized with
immunofluorescence and cytofluorimetry, respectively (left
axes/blue line: DNA; right axes/green line: Snail-1). Scale bar, 50
µm; magnification, ×400. (D) The motility of A549 cells in the
presence of the media conditioned by NA HBFs (left), ‘separated’
(middle) and ‘confronted’ NA HBF/A549 co-cultures (1:1 v/v with
fresh medium) was analyzed as in (A). Data are presented as the
mean ± standard error of the mean of 3 independent experiments.
*P<0.05 vs. A549. Note the mobilization of A549 in the presence
of AS HBF/A549-conditioned media, which was correlated with
Snail-1/Cx43 activation. HBFs, human bronchial fibroblasts; AS,
asthmatic donors; NA, non-asthmatic donors; VCD, velocity of cell
displacement; Cx43, connexin43; CM, conditional media. |
AS HBFs selectively modulate the
invasion of A549 cells
To further estimate the biological significance of
juxtacrine/paracrine loops between bronchial fibroblasts and lung
cancer cells, we analyzed the behavior of HBFs and A549 cells at
the confrontation front of their monolayers. When confronted with
A549 cells, NA HBFs formed lateral barrier structures, which were
similar to these seen in the ‘open’ co-cultures (see Fig. 1C) and remained impenetrable to A549
cells (Fig. 3A). In turn we observed
α-SMA up-regulation in AS HBFs. Concomitantly,
α-SMApositive AS HBFs efficiently infiltrated A549
monolayers, which was accompanied by the remodeling of interfaces
between AS HBF and A549 monolayers. Noteworthy, α-SMA expression
was also observed in A549 cells and in AS HBFs exposed to AS
HBF/A549 conditioned media (Fig. 3B).
On the other hand, we did not observe any collective infiltration
of AS HBF continua by A549 cells, even though Snail-1 was
accumulated in A549 nuclei throughout the contact zone with AS HBFs
(Fig. 3C). Instead, small and
scattered clusters of Cx43positive A549 cells were
observed within AS HBF continua (Fig. 3A
and B; inserts). This was accompanied by a relatively high
chemotactic response of A549 cells to the media conditioned by
‘open’ A549/AS HBF co-cultures (Fig.
3D). These data indicate that juxtacrine/paracrine signaling in
A549/HBF co-cultures facilitates the activation of AS HBFs and
secretion of chemotactic factors, which arrest lung cancer cells in
A549/HBF confrontation front. Small sub-populations of resistant
cells can overcome this arrest and migrate through cell-penetrable
interfaces between AS HBF and A549 monolayers.
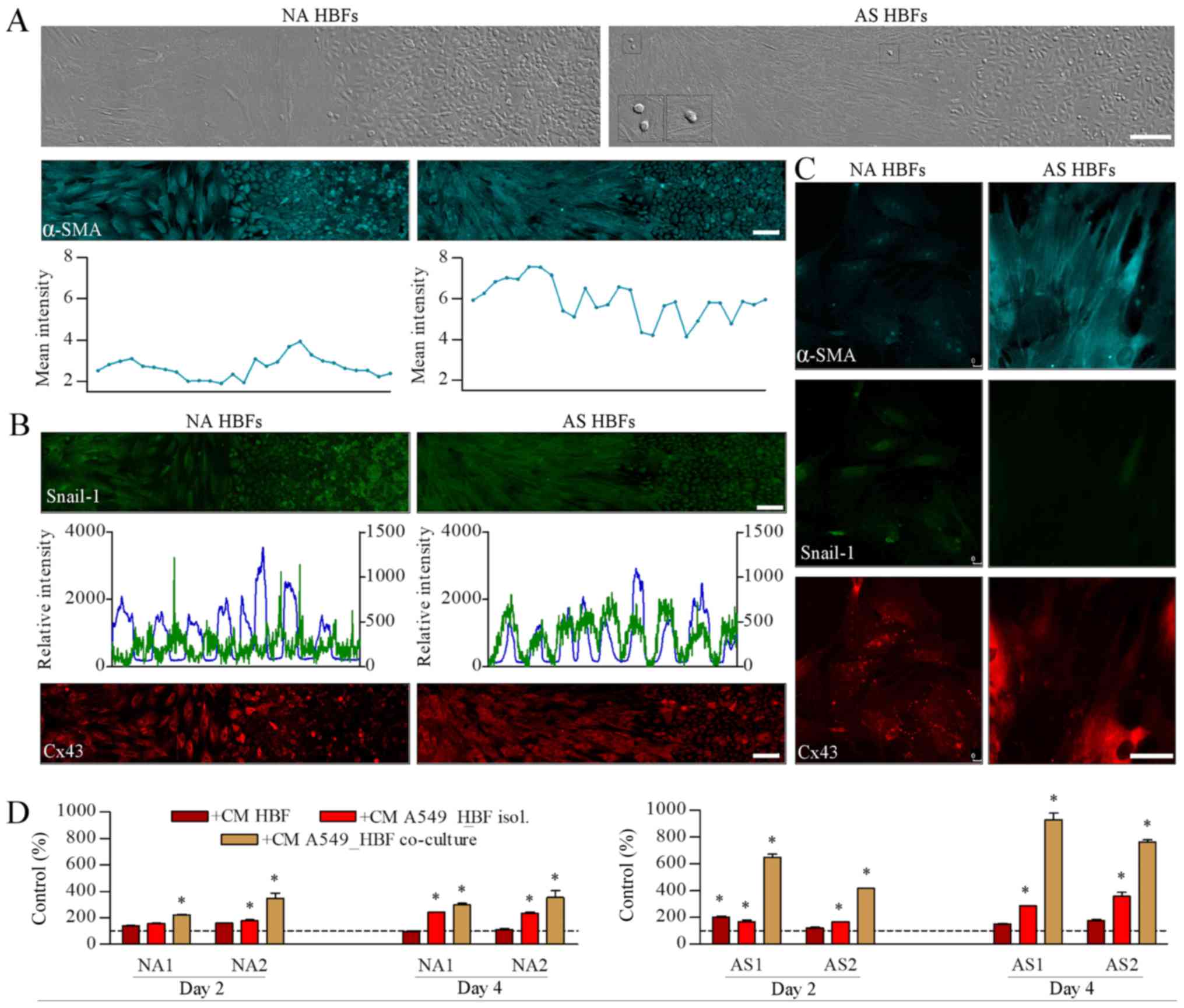 | Figure 3.AS HBFs selectively modulate the
invasion of A549 cells. (A) A549 cells were grown to confluence
with NA HBFs (left-hand panel) or AS HBFs (right-hand panel) in
2-well silicone inserts and co-incubated for 48 h following
diaphragm removal prior to fixation and immunostaining for α-SMA.
Expression of α-SMA was visualized by immunofluorescence and
cytofluorimetry, respectively. Scale bar, 100 µm; magnification,
×200. Inserts present A549 cells within the HBF monolayer. (B)
Cells were cultivated as in (A) and intracellular localization of
Snail-1/Cx43 and co-localization of Snail-1/DNA were visualized
with immunofluorescence and cytofluorimetry, respectively (left
axes: DNA; right axes: Snail-1). Photomicrographs in (A) and (B)
present panoramic till scans of the interfaces between HBF and A549
monolayers obtained by the composition of two rows of succeeding
images (7 pictures in a row). Scale bars, 100 µm. (C) NA HBFs and
AS HBFs were cultivated in the presence of the media conditioned by
‘separated’ NA HBF/A549 and AS HBF/A549 co-cultures, respectively,
for 48 h and immunostained against α-SMA, Snail-1 and Cx43. Scale
bars, 50 µm; magnification, ×630. (D) A549 cells were seeded onto
microporous membrane, placed in the wells filled with the media
conditioned by A549/HBF co-cultures, and allowed to transmigrate
for 48 h. The number of the transmigrated cells were counted
following 24 h. Data are presented as the mean ± standard error of
the mean of 3 independent experiments. *P<0.05 vs. A549
parameters in control medium. Note the infiltration of AS HBFs into
A549 monolayers, accompanied by the chemotactic activity of the
media from ‘open’ AS HBF/A549 co-cultures and by the presence of
scattered A549 within AS HBF continua. HBFs, human bronchial
fibroblasts; AS, asthmatic donors; NA, non-asthmatic donors; α-SMA,
α-smooth muscle actin; Cx43, connexin43; CM, conditional media. |
Discussion
The contribution of the tissue microenvironment to
the cancer disease is exemplified by the involvement of
endothelial, immune and connective tissue cells in cancer promotion
and progression. These processes are regulated by paracrine and
juxtacrine loops that are locally established between cancer and
stromal cells (19–21). In particular, the interactions between
cancer cells and CAFs participate in the formation of the scaffolds
that sustain the structure of tumor-protective tissue barriers
(22,23). CAFs can also generate signals crucial
for the microevolution and expansion of invasive cancer cell
sub-populations. Our study is the first to suggest the functional
links between lung cancer progression and pro-fibrotic properties
of fibroblasts that reside in asthmatic bronchi.
We have previously shown a high pro-fibrotic
activity of the fibroblasts derived from asthmatic bronchi. In
response to TGFβ, AS HBFs undergo
fibroblast-myofibroblast-transition (FMT), which facilitates
bronchial remodeling and asthmatic process in vivo (13,15,24). Here
we have shown that AS HBFs react to A549 cells and to AS HBF/A549
secretome with α-SMA/Cx43 up-regulation, which is a sign of their
myofibroblastic differentiation (15). Concomitantly, Snail-1/Cx43 activation
and the induction of A549 cell motility was detected in A549 cells
exposed to direct contacts with AS HBFs and to AS HBF/A549
secretome. Snail-1/Cx43-dependent axis has been suggested to
regulate the invasiveness of the prostate (17,25) and
lung cancer cells (26). Therefore,
these observations confirm that paracrine/juxtacrine interactions
between asthmatic CAFs and lung cancer cells contribute to the
phenotypic dynamics at the interface between the cancerous tissue
and bronchial stroma. The lack of the corresponding activation of
NA HBFs and A549 cells in NA HBF/A549 co-cultures suggests the
absence of the corresponding paracrine loops in non-asthmatic
bronchi.
On the other hand, we noticed the differences in the
quantity of motility-related A549 reactions to AS1 and AS2 HBFs.
They can be ascribed to the apparent phenotypic differences between
the discrete AS HBF lineages. In general, AS HBFs lineages derived
from different patients display a very high pro-fibrotic potential
in comparison to their counterparts from NA donors (6,13–15). However, they differ in morphology, a
proliferation rate, susceptibility to TGFβ, and the efficiency of
TGFβ-induced FMT. This is not surprising, since the phenotypic
characteristics of HBF lineages can be interpreted as the snapshots
of the resident cells' characteristics, which may differ between
the patients. A certain diversity of A549 reactions to AS1 and AS2
HBFs may thus illustrate a differential contribution of HBF
lineages to the lung cancer microenvironment in vivo.
Collectively, these data indicate that the asthmatic process may
constitute a local bronchial microenvironment that promotes lung
cancer remodeling and progression.
The potential significance of AS HBFs for cancer
development in vivo was also emphasized by their invasive
behavior in the proximity of A549 cells. AS HBFs failed to form
lateral barrier structures that are characteristic for their
non-asthmatic counterparts; instead, they collectively infiltrated
A549 monolayers (4). On the other
hand, we observed a relatively low translocation of A549 in
co-cultures with AS HBFs and the lack of collective infiltration of
AS HBF continua by A549 cells. This somewhat unexpected observation
can be interpreted in terms of a strong chemotactic activity of the
factors preferentially secreted by AS HBFs/A549 cells within the
contact zone. It suggests that combined juxtacrine/paracrine
interactions between AS HBFs and A549 cells counteract their
chemodynamic effect on A549 cells. These observations also confirm
the modulating effect of juxtacrine signaling on the
quality/quantity of integrated AS HBF/A549 secretome. Noteworthy,
scattered A549 cells were seen within AS HBF monolayers beyond AS
HBFs/A549 confrontation zones. This is consistent with our previous
report on the heterogeneity of A549 invasive potential (26). It shows that small sub-populations of
chemotaxis-resistant A549 cells can still colonize more distant
regions of asthmatic bronchi.
Epidemiologic association between asthma and the
risk of lung cancer formation is a controversial matter (9,27). For the
first time we have shown that the microenvironment of asthmatic
airways promotes the establishment of signaling loops between
bronchial fibroblasts and lung cancer cells. This observation
remains in concordance with the reports on intercellular signaling
between cancer cells and CAFs during cancer progression (18,28,29).
Accordingly, the infiltration of lung tumors by CAFs, which is
induced by paracrine loops between asthmatic CAFs and lung cancer
cells, may stabilize the structure of lung tumors. Chemotactic
arrest of lung cancer cells, enforced by the gradients of
chemotactic signals generated at the contact zone between tumor
cell mass and stroma, can further strengthen this effect.
Concomitant activation of cancer cells may stimulate local
remodeling of cancerous tissue. However, small sub-populations of
invasive, chemotaxis-resistant lung cancer cells, which penetrate
the compromised stromal barriers, can form new chemotactic loci and
prompt collective invasion of other cancer cells. Further research
based on the more comprehensive spectrum of lung cancer cell
lineages is necessary to verify this hypothesis, to elucidate a
biological significance of the expression and nuclear accumulation
of α-SMA in A549 cells and to identify the elements of AS HBF/A549
secretome, which are responsible for lung cancer cell activation.
However, our data suggest the role of the asthmatic bronchial
microenvironment and chronic inflammation in the formation and
stabilization of the invasive front(s) of lung tumors. Accordingly,
the potential mechanistic links between the asthmatic process and
lung cancer progression justify the inclusion of asthma into the
long list of potential prognostic markers in the lung cancer
therapy.
Acknowledgements
The authors would like to thank Mr. Dawid Wnuk
(Department of Cell Biology, Faculty of Biochemistry, Biophysics,
and Biotechnology, Jagiellonian University) for their technical
assistance.
Funding
Faculty of Biochemistry, Biophysics and
Biotechnology of Jagiellonian University is a partner of the
Leading National Research Center (KNOW) supported by the Ministry
of Science and Higher Education. The present study was financially
supported by the Student Research Grant Program at Faculty of
Biochemistry, Biophysics and Biotechnology, Jagiellonian University
(grant no. KNOW 5/2016) and partially by the Polish National
Science Centre (grant no. UMO-2015/19/D/NZ3/00273).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DR designed the study, obtained the data, performed
data analysis and prepared the manuscript. FR and KR obtained the
data and performed data analysis. JCa conducted data analysis and
prepared the manuscript. TW analyzed the data and prepared the
manuscript. MM designed the study. JCz conceived and designed the
study, performed data analysis and prepared the final manuscript
for publication.
Ethics approval and consent to
participate
The present study was approved by the Jagiellonian
University's Ethics Committee (decision no. 122.6120.69.2015) and
written informed consent was obtained for the use of HBF cell lines
in basic research, which had previously been obtained from all
donors.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HBF
|
human bronchial fibroblasts
|
|
Cx43
|
connexin43
|
|
α-SMA
|
α-smooth muscle actin
|
|
EMT
|
epithelial-mesenchymal transition
|
|
FMT
|
fibroblast-myofibroblast
transition
|
|
CAFs
|
cancer-associated fibroblasts
|
|
CM
|
conditional media
|
|
CME
|
coefficient of movement efficiency
|
References
|
1
|
Anandan C, Nurmatov U, van Schayck OC and
Sheikh A: Is the prevalence of asthma declining? Systematic review
ofepidemiological studies. Allergy. 65:152–167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Al Muhsen S, Johnson JR and Hamid Q:
Remodeling in asthma. J Allergy Clin Immunol. 128:451–462. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Halwani R, Al Muhsen S, Al Jahdali H and
Hamid Q: Role of transforming growth factor-β in airway remodeling
in asthma. Am J Respir Cell Mol Biol. 44:127–133. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ingram JL, Huggins MJ, Church TD, Li Y,
Francisco DC, Degan S, Firszt R, Beaver DM, Lugogo NL, Wang Y, et
al: Airway fibroblasts in asthma manifest an invasive phenotype. Am
J Respir Crit Care Med. 183:1625–1632. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Darby IA, Laverdet B, Bonté F and
Desmoulière A: Fibroblasts and myofibroblasts in wound healing.
Clin Cosmet Investig Dermatol. 7:301–311. 2014.PubMed/NCBI
|
|
6
|
Michalik M, Pierzchalska M, Wlodarczyk A,
Wójcik KA, Czyż J, Sanak M and Madeja Z: Transition of asthmatic
bronchial fibroblasts to myofibroblasts is inhibited by cell-cell
contacts. Respir Med. 105:1467–1475. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Agarwal SK: Integrins and cadherins as
therapeutic targets in fibrosis. Front Pharmacol. 5:1312014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Desmoulière A, Chaponnier C and Gabbiani
G: Tissue repair, contraction, and the myofibroblast. Wound Repair
Regen. 13:7–12. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rosenberger A, Bickeböller H, McCormack V,
Brenner DR, Duell EJ, Tjønneland A, Friis S, Muscat JE, Yang P,
Wichmann HE, et al: Asthma and lung cancer risk: A systematic
investigation by the International Lung Cancer Consortium.
Carcinogenesis. 33:587–597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Attieh Y and Vignjevic DM: The hallmarks
of CAFs in cancer invasion. Eur J Cell Biol. 95:493–502. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuzet SE and Gaggioli C: Fibroblast
activation in cancer: When seed fertilizes soil. Cell Tissue Res.
365:607–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fujita A, Kameda Y and Goya T:
Clinicopathology of stromal invasion in lung adenocarcinoma. Pathol
Int. 59:1–6. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Michalik M, Soczek E, Kosinska M, Kosińska
M, Rak M, Wójcik KA, Lasota S, Pierzchalska M, Czyż J and Madeja Z:
Lovastatin-induced decrease of intracellular cholesterol level
attenuates fibroblast-to-myofibroblast transition in bronchial
fibroblasts derived from asthmatic patients. Eur J Pharmacol.
704:23–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wójcik KA, Skoda M, Koczurkiewicz P, Sanak
M, Czyż J and Michalik M: Apigenin inhibits TGF-β1 induced
fibroblast-to-myofibroblast transition in human lung fibroblast
populations. Pharmacol Rep. 65:164–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paw M, Borek I, Wnuk D, Ryszawy D,
Piwowarczyk K, Kmiotek K, Wójcik-Pszczoła KA, Pierzchalska M,
Madeja Z, Sanak M, et al: Connexin43 controls the myofibroblastic
differentiation of bronchial fibroblasts from patients with asthma.
Am J Respir Cell Mol Biol. 57:100–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Piwowarczyk K, Paw M, Ryszawy D,
Rutkowska-Zapała M, Madeja Z, Siedlar M and Czyż J:
Connexin43high prostate cancer cells induce endothelial
connexin43 up-regulation through the activation of intercellular
ERK1/2-dependent signaling axis. Eur J Cell Biol. 96:337–346. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ryszawy D, Sarna M, Rak M, Szpak K,
Kędracka-Krok S, Michalik M, Siedlar M, Zuba-Surma E, Burda K,
Korohoda W, et al: Functional links between Snail-1 and Cx43
account for the recruitment of Cx43-positive cells into the
invasive front of prostate cancer. Carcinogenesis. 35:1920–1930.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Erdogan B and Webb DJ: Cancer-associated
fibroblasts modulate growth factor signaling and extracellular
matrix remodeling to regulate tumor metastasis. Biochem Soc Trans.
45:229–236. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Condeelis J and Pollard JW: Macrophages:
Obligate partners for tumor cell migration, invasion, and
metastasis. Cell. 124:263–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Baran B, Bechyne I, Siedlar M, Szpak K,
Mytar B, Sroka J, Laczna E, Madeja Z, Zembala M and Czyz J: Blood
monocytes stimulate migration of human pancreatic carcinoma cells
in vitro: The role of tumour necrosis factor-alpha. Eur J Cell
Biol. 88:743–752. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fidler IJ, Kim SJ and Langley RR: The role
of the organ microenvironment in the biology and therapy of cancer
metastasis. J Cell Biochem. 101:927–936. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bremnes RM, Al Shibli K, Donnem T, Sirera
R, Al-Saad S, Andersen S, Stenvold H, Camps C and Busund LT: The
role of tumor-infiltrating immune cells and chronic inflammation at
the tumor site on cancer development, progression, and prognosis:
Emphasis on non-small cell lung cancer. J Thorac Oncol. 6:824–833.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bremnes RM, Dønnem T, Al Saad S, Al-Shibli
K, Andersen S, Sirera R, Camps C, Marinez I and Busund LT: The role
of tumor stroma in cancer progression and prognosis: Emphasis on
carcinoma-associated fibroblasts and non-small cell lung cancer. J
Thorac Oncol. 6:209–217. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sarna M, Wojcik KA, Hermanowicz P, Wnuk D,
Burda K, Sanak M, Czyż J and Michalik M: Undifferentiated bronchial
fibroblasts derived from asthmatic patients display higher elastic
modulus than their non-asthmatic counterparts. PLoS One.
10:e01168402015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Czyż J, Szpak K and Madeja Z: The role of
connexins in prostate cancer promotion and progression. Nat Rev
Urol. 9:274–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bechyne I, Szpak K, Madeja Z and Czyż J:
Functional heterogeneity of non-small lung adenocarcinoma cell
sub-populations. Cell Biol Int. 36:99–103. 2011. View Article : Google Scholar
|
|
27
|
Qu YL, Liu J, Zhang LX, Wu CM, Chu AJ, Wen
BL, Ma C, Yan XY, Zhang X, Wang DM, et al: Asthma and the risk of
lung cancer: A meta-analysis. Oncotarget. 8:11614–11620. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Langley RR and Fidler IJ: Tumor cell-organ
microenvironment interactions in the pathogenesis of cancer
metastasis. Endocr Rev. 28:297–321. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Erdogan B, Ao M, White LM, Means AL,
Brewer BM, Yang L, Washington MK, Shi C, Franco OE, Weaver AM, et
al: Cancer-associated fibroblasts promote directional cancer cell
migration by aligning fibronectin. J Cell Biol. 216:3799–3816.
2017. View Article : Google Scholar : PubMed/NCBI
|














