Introduction
Tumor tissue contains not only cancer cells but also
several stromal components, such as macrophages, immune cells and
fibroblasts. Those cells that are profoundly involved in the
regulation of cancer cell growth. Many studies have reported that
fibroblasts exert important effects on cancer cell growth.
Fibroblast-like stromal cells (stromal cells) contained in tumor
tissues exhibit a myofibroblast phenotype (expressing α-smooth
muscle actin). This population regulates cancer cell growth
(1), cancer cell stemness (2,3), formation
of a premetastatic niches (4) and
drug resistance (5,6), through the secretion of soluble factors.
Thus, interactions between cancer cells and stromal cells
(tumor-stromal cell interactions) are attractive targets for cancer
therapy. Indeed, many studies have reported the presence of
positive regulatory interactions between cancer cells and stromal
cells mediated by secreted factors. However, in a mouse model,
others have shown that stromal cells can have a negative regulatory
impact in a mouse model. Yin et al (7), reported that IL-25 secreted from
tumor-associated fibroblasts suppressed mammary tumor metastasis
and that IL-25 secretion was increased by a lignin derivative.
We have focused on the negative regulation of cancer
cells by stromal cell secreted factors. This approach offers a
novel strategies for discovering new cancer therapeutics. We used a
co-culture system to screen small molecules from natural sources
such as microbial culture media, seeking molecules that modulated
tumor-stromal cell interactions. In additional studies, we used
co-culture systems pairing cancer cells and stromal cells from
several organs. We reported the identification of small molecules
that suppressed cancer cell growth through modulation of stromal
cells. Leucinostatin A and Phthoxazolin A were found by co-culture
screening of prostate cancer cells with prostate stromal cells.
Those agents suppressed cancer cell growth by inhibiting the
expression of insulin-like growth factor-1 (IGF-1) by stromal cells
(8,9).
We also reported that MEK-inhibitor I induced the secretion of
GAPDH by gastric stromal cells, a process that suppressed gastric
cancer cell growth (10). In a recent
study, we found a novel compound, intervenolin (ITV), from the
culture medium of Nocardia sp. Ml96-86F2 (11). ITV inhibited the in vitro
growth of human gastric cancer cells. Importantly, inhibition was
greater when the cancer cells were co-cultured with stromal cells.
Based on this result, we carried out proteomic analysis of
conditioned medium from human gastric fibroblast-like stromal cells
(Hs738), and the results showed that ITV induced TSP-1 secretion
from Hs738 cells.
TSP-1 is a glycoprotein that forms a homo tetramer
in the extracellular microenvironment. It is secreted by several
cell types, including platelets, epithelial cells and fibroblasts.
Rodríguez-Manzaneque et al (12), reported that TSP-1 showed antitumor
activity. In the present study, we report that TSP-1 from
conditioned medium (CM) of Hs738 cells treated with ITV inhibited
the growth of cancer cells through its TSP-1 receptor.
Materials and methods
Cell lines and reagents
Human gastric cancer cell lines MKN-7 and MKN-74
were obtained from the RIKEN cell bank (Tsukuba, Japan). MKN-7 and
MKN-74 stably express a transfected GFP vector as described
(10). Cancer cell lines were
maintained in Dulbecco's Modified Eagle's Medium (DMEM; Nissui,
Tokyo, Japan) supplemented with 10% fetal bovine serum (FBS;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 100 U/ml penicillin
G (Thermo Fisher Scientific, Inc., Waltham, MA, USA), and 100 µg/ml
streptomycin (Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. Hs738. St/Int (Hs738) human gastric stromal cells
(CRL-7869) were obtained from the ATCC (American Type Culture
Collection, Manassas, VA, USA). Stromal cells were maintained in
DMEM supplemented with 10% FBS, 100 U/ml penicillin G, 100 µg/ml
streptomycin, ITH (5 µg/ml insulin, 5 µg/ml transferrin, and 1.4 µM
hydrocortisone), and 5 ng/ml basic FGF (PeproTech, Inc., Rocky
Hill, NJ, USA) at 37°C with 5% CO2 as described
(10). Recombinant human
thrombospondin-1 protein (3074-TH) was purchased from R&D
Systems, Inc., (Minneapolis, MN, USA). Neutralizing antibodies for
TSP-1, A4.1 (mouse monoclonal, MS-418-PABX) and C6.7 (mouse
monoclonal, MS-420-PABX) were purchased from NeoMarkers, Inc.,
(Fremont, CA, USA). ITV was synthesized as described previously
(11).
Preparation of CM from Hs738
Hs738 cells were cultured at 5×104
cells/ml in DMEM supplemented with ITH and 5% FBS. After 1 day, the
medium was replaced with fresh medium including ITV (0.25 µg/ml)
without FBS (serum-free CM). After an additional 4 days of
incubation, CM from Hs738 (Ctrl CM) or ITV-treated Hs738 (ITV CM)
were collected and centrifuged to remove debris. Gastric cancer
cells (3×105 cells/ml) were inoculated in 1 ml of the
75% CM of Hs738 cells or assay medium alone in 35-mm dishes and
cultured for 1 day with dialyzed FBS. The cells were washed with
phosphate-buffered saline (PBS) and the cell lysates were prepared
for western blotting. For cell growth assays, MKN-74 cells were
cultured at 5×103 cells/well in 96-well plates in Hs738
CM at the indicated concentration with dialyzed FBS.
SDS-PAGE and proteomic analysis of
CM
Serum-free CMs were collected from Hs738 cells
treated with or without ITV. Proteins were separated by SDS-PAGE,
and the gel was stained with a PlusOne silver staining kit (GE
Healthcare, Little Chalfont, UK) according to the manual. Bands
showing different patterns in CMs treated with and without ITV were
excised. These excised bands were digested with trypsin, and the
digested peptides were then subjected to LC-MS/MS analysis
(LTQ-Orbitrap; Thermo Fisher Scientific, Inc.). For tryptic in-gel
digestion, the candidate protein bands were cut into small pieces
and destained with 100 µL of a 1:1 solution of 30 mM potassium
ferricyanide (Fujifilm Wako Pure Chemical Industries, Ltd., Tokyo,
Japan) and 100 mM sodium thiosulfate (Fujifilm Wako Pure Chemical
Industries, Ltd.). Reduction was performed by re-swelling the gel
pieces in 100 µL of 10 mM dithiothreitol (Nacalai Tesque, Tokyo,
Japan) in 100 mM NH4HCO3 (Nacalai Tesque)
buffer solution and incubated at 56°C for 1 h. After cooling, the
excess liquid was removed and quickly replaced by the same volume
of freshly prepared 55 mM iodoacetamide in 100 mM
NH4HCO3 solution. The iodoacetamide solution
was removed and the gel pieces were washed with 200 µL 50%
acetonitrite (ACN) in 100 mM NH4HCO3 solution
for 10 min, and the liquid was removed. One band (gel piece)
required 15–20 µL of 0.01 µg trypsin (Trypsin Gold, Mass
Spectrometry Grade; Promega Corporation, Madison, WI, USA) in 100
mM NH4HCO3 digestion buffer. After incubating
the reaction mixture at 37°C overnight, the digestion buffer was
removed and saved in a new tube. The peptides from the gel pieces
were extracted by adding 15–20 µL of 50–95% ACN in 0.1%
trifluoroacetic acid (TFA) aq. and incubating for 15 min with
sonication. Finally, the samples were purified using Solid Phase
Extraction C-TIP (Nikkyo Technos Co., Ltd., Tokyo, Japan) after
adding 25 µL of 2% ACN in 0.1% TFA aq. Samples were kept in −20°C
before being analyzed by LC-MS/MS.
LC-ESI MS/MS analysis
LC-ESI MS/MS analysis was performed using a Paradigm
MS4 LC system (Michrom BioResources, Auburn, CA, USA) coupled with
an LTQ Orbitrap™ XL mass spectrometer (Thermo Fisher
Scientific, Inc.). An L-column ODS, 3 µm, 0.1 mm id × 50 mm (CERI,
Tokyo, JAPAN) was used. HPLC was performed using a linear gradient
solvent system (solvent A; 0.1% formic acid in 2% ACN aq., solvent
B; 0.1% formic acid in 90% ACN) from 5 to 45% solvent A at a flow
rate of 0.5 µL/min for 20 min followed by MS/MS equipped with a
CaptiveSpray™ source (Bruker Corporation, Billerica, MA, USA). The
sprayer voltage was set to 1.8 kV, and capillary temperature was
set to 200°C. Parent mass peaks with ranges from m/z 400 to
2,000 were selected for MS/MS analysis by Data Dependent Top3.
Normalized collision energy was fixed at 35. The MS/MS data were
processed using Xcalibur™ 2.2 SP1.48. Peptides were
searched with MASCOT Server 2.5 (Matrix Science, Inc., Boston, MA,
USA). The search parameters were set as follows: cRAP 20100518 (112
sequences; 37,418 residues) and IPI_human (91,464 sequences;
36,355,611 residues) database, enzyme set for trypsin, allowance of
one missed cleavage, variable modifications were
carbamidomethylation at cysteine residues and oxidation at
methionine residues, peptide and MS/MS tolerance was 5 ppm and 0.6
Da, respectively, and the limit of peptide charges state was 2+,
and 3+.
Immunoblotting
Cells (3×105) were cultured in 6-well
plates for the indicated times. The cells were washed twice with
ice-cold PBS containing 100 µM Na3VO4 and
then treated with lysis buffer (20 mM Hepes, pH 7.5, 150 mM NaCl,
1% Triton X-100, 10% glycerol, 1 mM EDTA, 50 mM NaF, 50 mM
β-glycerophosphate, 1 mM Na3VO4, and 25 µg/ml
each of antipain, leupeptin, and pepstatin) (13). Equal amounts of protein extracts were
separated by SDS-PAGE and transferred onto Immobilon transfer
membrane (IPVH304F0, Millipore, Bedford, MA). After incubation with
the primary antibody at room temperature (RT) for 1 h, membranes
were incubated with HRP-conjugated secondary antibody at RT, 30
min. ECL Western Blotting Detection Reagents (RPN2209; GE
Healthcare) were used to visualize the immunoblot signals and
signals were determined by ImageQuant LAS4000 mini (GE Healthcare).
Relative intensities of determined bans were calculated as peak
area using Image J (National Institutes of Health, Bethesda, MD,
USA). The following antibodies from Santa Cruz Biotechnology, Inc.,
(Dallas, TX, USA) were used for primary labeling: Anti-TSP-1
(1:1,000; mouse monoclonal, sc-59887), anti-CDK4 (1:1,000; rabbit
polyclonal, sc-260) and anti-Rb (1:1,000; rabbit polyclonal,
sc-50). Anti-c-Myc antibody (1:1,000; rabbit monoclonal, cat. no.
13987) antibody was purchased from Cell Signaling Technology, Inc.,
(Danvers, MA, USA). Anti-α-tubulin (1:1,000; mouse monoclonal,
T5168) antibody was purchased from Sigma-Aldrich; Merck KGaA.
Anti-rabbit IgG, peroxidase-linked species-specific whole antibody
(from donkey, NA 934; GE Healthcare) or anti-mouse IgG,
peroxidase-linked species-specific whole antibody (from donkey, NA
931; GE Healthcare) were as the secondary antibody.
Cell growth and co-culture
experiments
Cells were inoculated into 96-well plates at
5×103 cells per well in 0.1 ml of DMEM supplemented with
1% D-FBS and ITH. After culture for the indicated number of days,
cell growth was determined using
3-(4,5-Dimethyl-2-lythiazolyl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT; 341-01823; Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) as described (14).
Alternatively, cells were lysed in 10 mM Tris-HCl pH 7.4, 150 mM
NaCl, 0.9 mM CaCl2, and 1% Triton X-100 and growth was
measured by determining GFP fluorescence intensity (excitation at
485 nm and emission at 538 nm, assessed with a Cytation 5 plate
reader; BioTek Instruments, Inc., Winooski, VT, USA). For
co-culture experiments, Hs738 cells were first inoculated into
96-well plates at 5×103 cells/well in 0.1 ml of DMEM
supplemented with 1% D-FBS and ITH. ITV was added to the wells and
the cells were cultured for 2 days. For cultivation with CM alone,
culture medium from Hs738 after 2 days of incubation with ITV was
removed to new wells. Then, 10 µL of a gastric cancer cell
suspension (5×104 cells/ml) in serum-free DMEM were
added to the monolayer of Hs738 cells or only CM and the cells were
further cultured for 3 days. For monoculture of gastric cancer
cells, assay medium alone with ITV was first incubated for 2 days,
and then gastric cancer cells were added as described above and
further cultured for 3 days. The growth of gastric cancer cells was
determined by measuring GFP fluorescence intensity as described
(10). For ‘non-contact’ co-culture
experiments, 0.6 ml of Hs738 cells (5×104 cells/ml) in
DMEM supplemented with 1% D-FBS and ITH (co-culture) or only assay
medium (mono culture) were first added to outer wells of 24-well
cell culture insert plates and 0.1 ml of the assay medium was added
to the inner wells with 0.4 µm pore size membranes (140620; Thermo
Fisher Scientific, Inc.). After 2 days of culture, 10 µL of a
gastric cancer cell suspension (5×103) in serum-free
DMEM was added to the inner wells and the plates were further
cultured for 3 days.
Blocking of TSP-1 activity by
neutralizing antibody
Recombinant human TSP-1 (12.5 µg/ml) in DMEM was
incubated with a monoclonal anti-TSP-1 IgM antibody (Clone A4.1 or
C6.7 or not) at 37°C for 1 h. MKN-74 cells were cultured at
5×103 cells/ml in 0.1 ml of above medium with ITH and
dialyzed FBS in a 96-well plate for 3 days. For blocking of CM
activity, CM of Hs738 was prepared with ITV (1 µg/ml) in the
presence of 1% ITH and dialyzed FBS for 3 days. MKN-74 cells were
cultured in CM or DMEM (containing 1% ITH and dialyzed FBS) with
monoclonal anti-TSP-1 IgM antibody (Clone A4.1 or C6.7 or not) for
3 days.
Statistical analysis
All data are representative of at least 3
independent experiments with similar results. Statistical analysis
was carried out using two-tailed Student's t-test or analysis of
variance followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of ITV on the growth of gastric
cancer cells co-cultured with gastric stromal cells
The effect of ITV on gastric cancer cells
co-cultured with gastric stromal cells was evaluated as presented
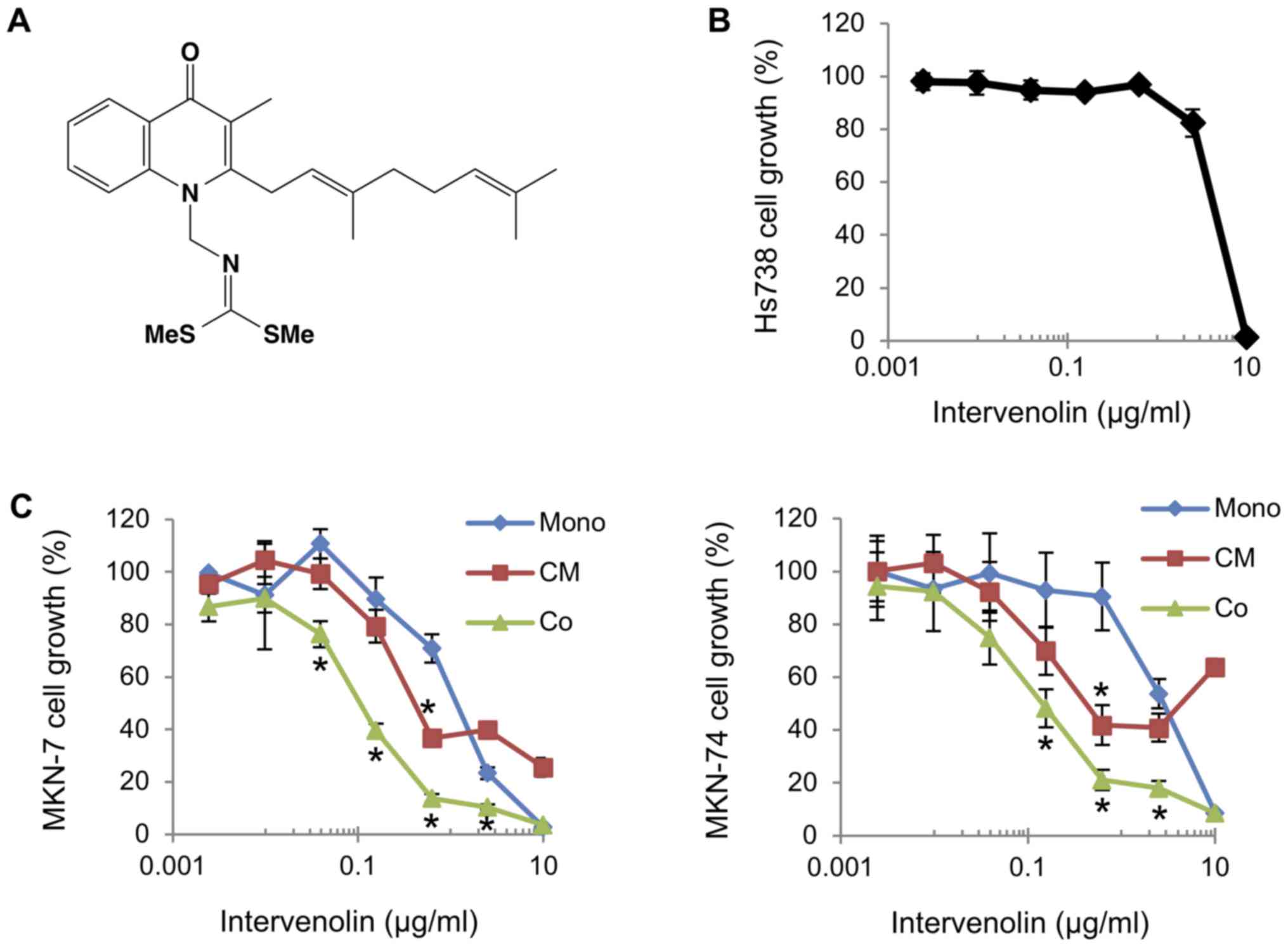
in Fig. 1. The structure of ITV is
shown in Fig. 1A. Human gastric
cancer cells (MKN-7 and MKN-74) were cultured with human gastric
stromal cells (Hs738) in the presence of various concentrations of
ITV (Fig. 1C). ITV inhibited the
growth of MKN-7 and MKN-74 cells cultured alone with
IC50 values of 1.45 and 3.13 µg/ml, respectively.
Importantly, when co-cultured with Hs738, ITV inhibited the growth
of the lines at much lower concentrations (IC50 values
of 0.12 and 0.15 µg/ml respectively; Fig.
1C). In contrast, the growth of Hs738 cells was inhibited by
ITV treatment only when the concentration 10 µg/ml (Fig. 1B). MKN-7 and MKN-74 were cultured in
CM taken from Hs738 cells treated with ITV (ITV CM). Growth was
also inhibited more strongly than when they were cultured alone
with IC50 values of 0.48 and 0.49 µg/ml, respectively
(Fig. 1C). Moreover, the
growth-inhibitory activity of CM from Hs738 was dependent on the
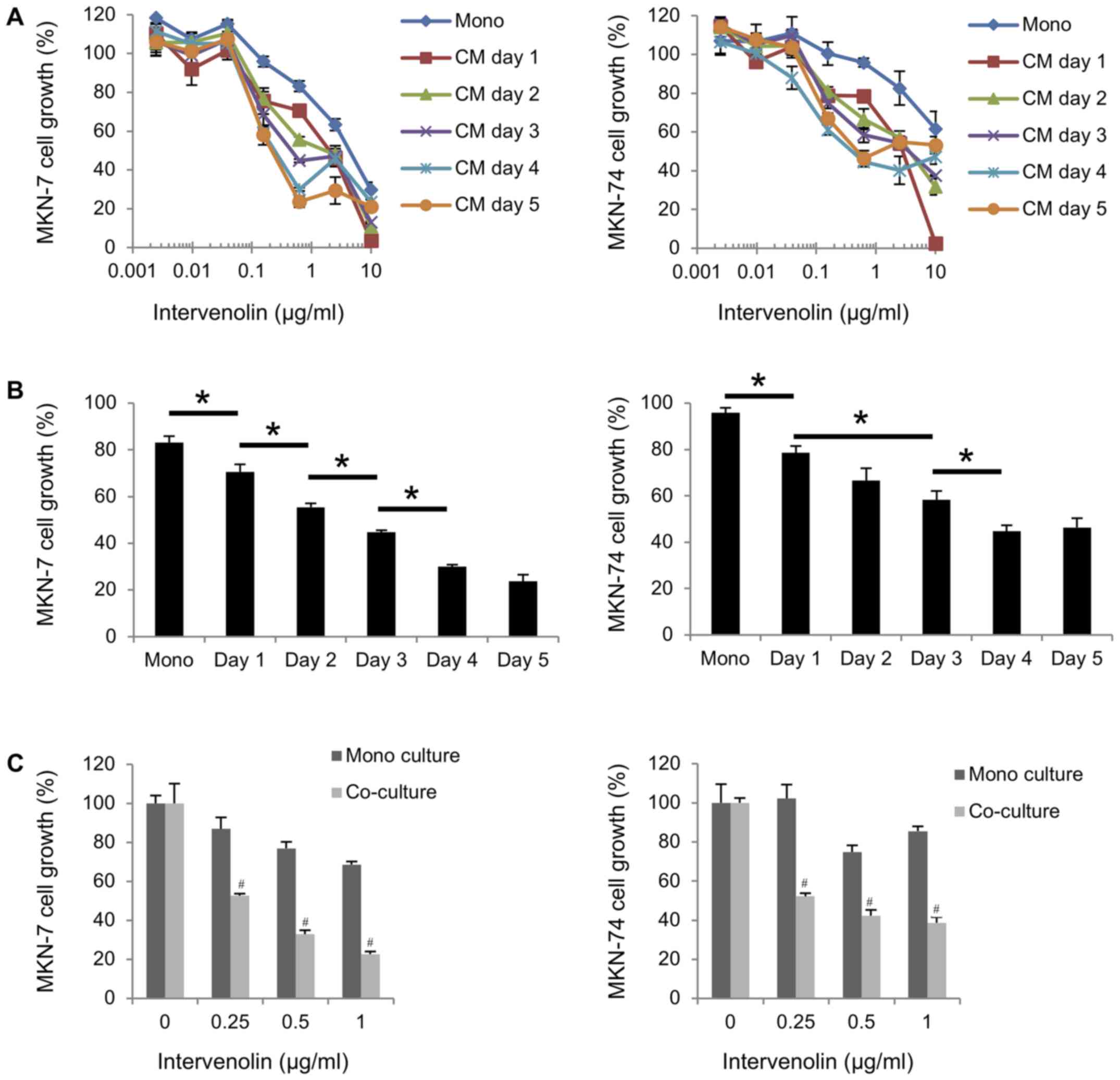
number of days with which the cells were treated with ITV (Fig. 2A and B). To clarify whether the
inhibitory activity of ITV required direct cell-cell contact, we
conducted ‘non-contact’ co-culture experiment. The data showed that
ITV inhibited the growth of gastric cancer cells when they were
co-cultured with Hs738 more strongly than in mono culture
conditions (Fig. 2C). These results
suggested that ITV induced the secretion of anti-cancer factors
from Hs738 into the CM. The cytotoxicity of the highest ITV
concentration (10 µg/ml) for Hs738, MKN-7 and MKN-74 at mono
culture condition was considered to be non-specific and attributed
to the high lipophilicity of ITV (Fig. 1B
and C).
Determination of the anti-cancer
factors in ITV CM of Hs738
We next prepared serum-free CM from Hs738 cells and
carried out proteomic analyses with LC-MS/MS to determine the
anti-cancer factors secreted by ITV-treated HS738 cells. Whereas
the growth of MKN-7 or MKN-74 cells was not changed in Ctrl CM, it
was significantly inhibited in ITV CM, which was dependent on the
concentration of ITV CM. This inhibitory effect of ITV CM for
cancer cell growth was greater than ITV alone or Ctrl CM containing
the same amount of ITV in ITV CM (Fig.
3A). Furthermore, fluorescence microscopy showed growth
inhibition of MKN-7 or MKN-74 cells cultured in ITV CM. The outcome
was similar to the results obtained with the MTT assay in Fig. 3A (Fig.
3B). SDS-PAGE of the CMs with silver staining showed that the
ITV CM contained different proteins than did Ctrl CM, particularly
around 150 kDa. These bands were excised (three bands from Ctrl CM
and five bands from ITV CM; Fig. 3C).
LC-MS/MS analysis revealed that these excised bands were composed
of collagens, but TSP-1 was identified in band No. 5 (Score: 134,
Coverage: 6.2, Mass: 133291). Because collagen band patterns were
changed by ITV treatment, we demonstrated that the protein level of
α-SMA was decreased by ITV treatment (Fig. 3D). Since type I collagen and α-SMA are
myofibroblast markers, these results suggested that ITV might
affect the differentiation of Hs738 cells.
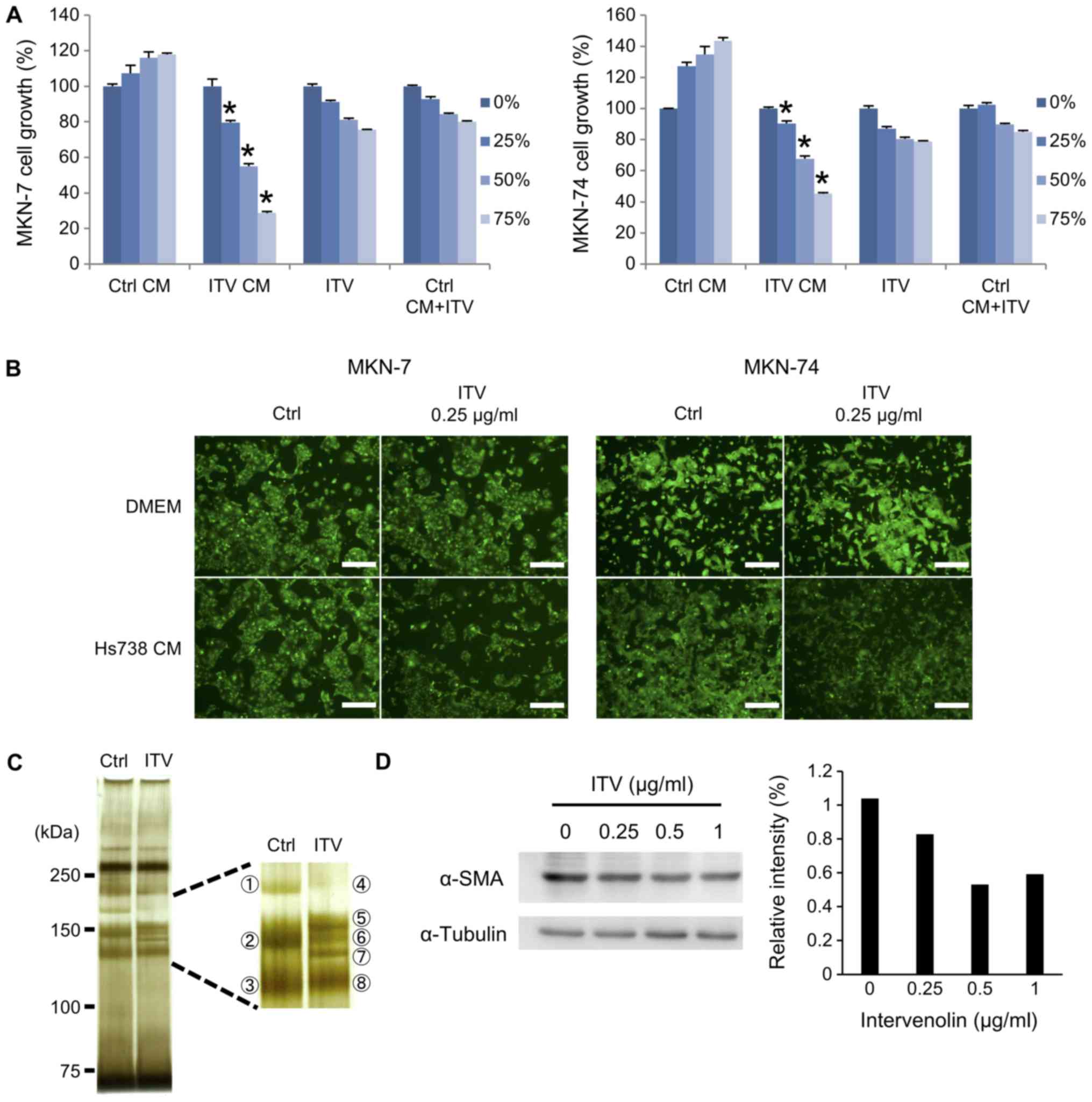 | Figure 3.Determination of the anti-cancer
factors in ITV CM. (A) Hs738 cells were cultured under serum-free
conditions for 4 days to prepare CM. Ctrl CM, ITV CM, ITV (DMEM
contained ITV at the same levels as ITV CM) or Ctrl CM+ITV (Ctrl CM
contained ITV at the same amount of ITV CM) was diluted with DMEM
to become 0, 25, 50, or 75% dilution. MKN-7 or MKN-74 cells were
cultured in the diluted Ctrl CM, ITV CM, ITV or Ctrl CM+ITV for 3
days. Results show means ± SE (n=3). *P<0.05 vs. Ctrl CM. (B)
Representative photomicrographs of cultured gastric cancer cell
lines under fluorescence microscopy. Green: GFP. Scale bar is 200
µm. Upper panels show gastric cancer cell lines cultured in DMEM
with or without ITV (0.25 µg/ml), and lower panels show both cell
lines in Ctrl CM or ITV CM. (C) Equal volumes of Ctrl CM or ITV CM
prepared in serum-free condition were separated with SDS-PAGE and
silver-stained. The right box is an enlarged image of the stained
gel and indicates excised bands. (D) Western blot analysis with
extracts of Hs738 cells cultured with the indicated concentrations
of ITV for 1 day. Relative intensities of determined bands of α-SMA
(normalized with α-tubulin, analyzed by Image J) are shown. Western
blot analyses were repeated at least three time with similar
results. A representative result is shown here. ITV, intervenolin;
CM, conditioned medium; Hs738, human gastric fibroblast-like
stromal cells. |
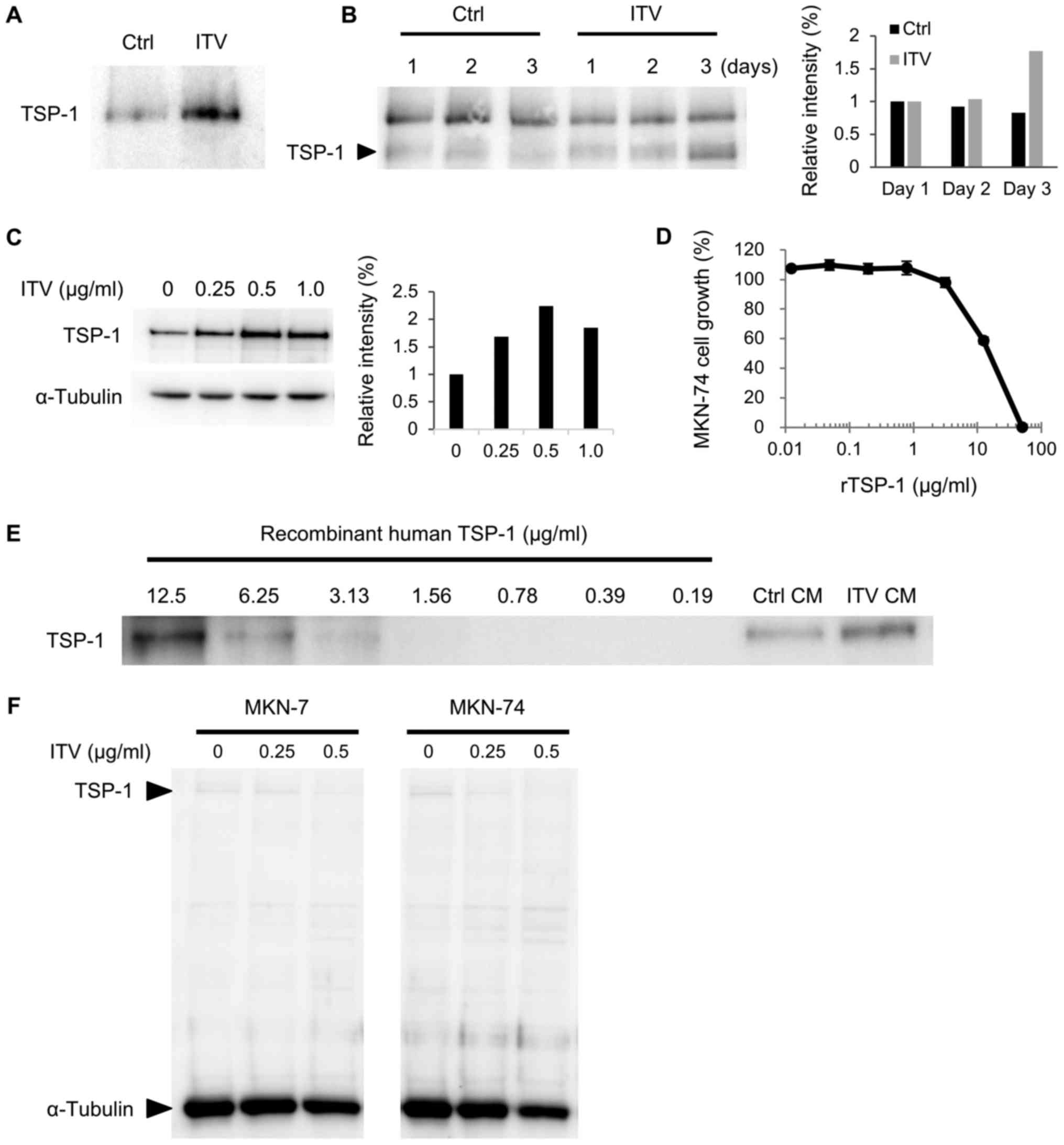
ITV treatment induces TSP-1 secretion
from Hs738 cells
We further analyzed the level of TSP-1 protein in
ITV CM. Western blot analysis showed the amount of TSP-1 was
significantly increased by ITV treatment in serum-free CM that was
used for proteomic analysis (Fig.
4A). We also found that TSP-1 in ITV CM increased from day 1 to
day 3 of incubation (Fig. 4B). In
addition, the protein level of intracellular TSP-1 in Hs738 cells
was upregulated by ITV in a dose-dependent manner up to 0.5 µg/ml
of ITV (Fig. 4C). Since the
inhibitory effect of ITV CM was stronger in MKN-74 cells than MKN-7
cells (Fig. 1C), we used the former
for further studies. Recombinant human TSP-1 exhibited inhibitory
activity on the growth of MKN-74 cells (Fig. 4D). Semi-quantitative analysis of TSP-1
in CM by western blot using recombinant TSP-1 as control showed
that ITV CM contained approximately 10 µg/ml of TSP-1 (Fig. 4E). In contrast, the amounts of
intracellular TSP-1 in MKN-7 and MKN-74 cells were very low and
they were not changed by ITV (Fig.
4F). These results suggested that ITV acts on Hs738 cells to
induce the secretion of TSP-1 and that TSP-1 in CM suppresses the
growth of gastric cancer cells.
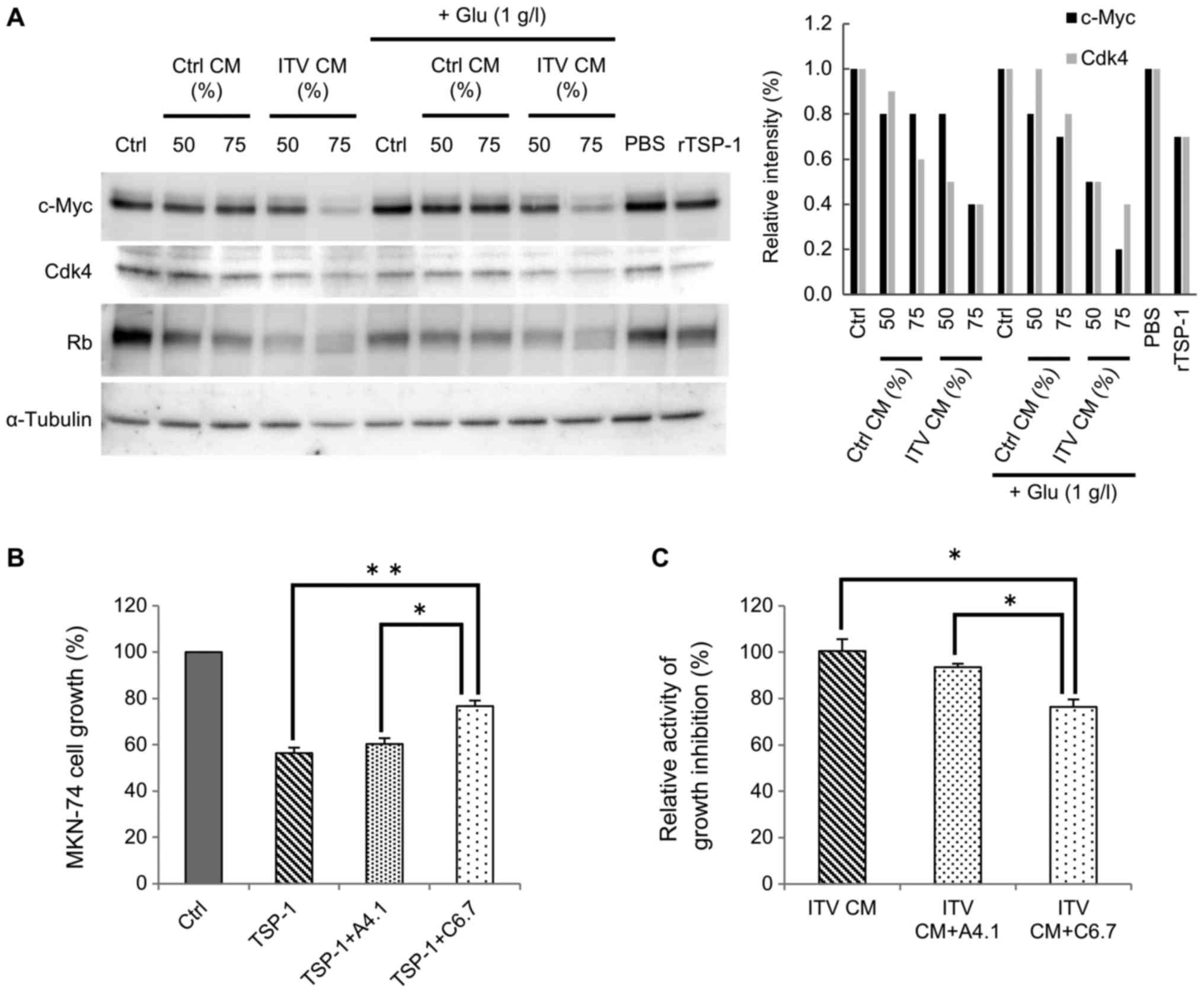
Hs738 CM treated with ITV inhibits
c-Myc and its downstream signaling in cancer cells
Kaur et al (15), reported that TSP-1 regulated c-Myc via
CD47. Thus, we asked whether the inhibition of cancer cell growth
by ITV CM was related to expression of c-Myc in cancer cells.
Western blot analysis of MKN-74 cells that had been cultured in 50%
or 75% CM (Fig. 5A) was performed.
Analysis showed that c-Myc expression in MKN-74 cells was markedly
decreased by ITV CM treatment (Fig.
5A). With the suppression of c-Myc, expression levels of CDK4,
whose expression is controlled by c-Myc (16), was also decreased by ITV CM. Note that
the molecular weight of Rb protein was reduced. Those data
indicated the presence of a non-phosphorylated form of this
protein, suggesting that the growth of the cancer cells was likely
inhibited by suppression of the cell cycle. Similar results were
obtained with recombinant human TSP-1 (rTSP-1) treatment. Thus, we
hypothesized that the suppression of cancer cell growth by ITV CM
was due to TSP-1 that was released into the CM. It is known that
the expression level of c-Myc is affected by glucose starvation
(17). However, the same result was
obtained even when extra glucose was added to CM (Fig. 5A).
Next, we tried to block the interaction of TSP-1
with CD36 or CD47 (receptors for TSP-1) (15), using a neutralizing antibody against
each receptor. If CD47 were involved in TSP-1 signaling and cell
cycle arrest, the activity of ITV CM should be cancelled by
neutralizing antibody against CD47. Indeed, the growth inhibitory
activity of rTSP-1 on MKN-74 cells was partially inhibited by C6.7,
a neutralizing antibody for TSP-1 that blocks interaction with
CD47. Interestingly, growth inhibition was not affected by A4.1, a
neutralizing antibody that blocks interaction with CD36 (Fig. 5B). The growth inhibitory activity of
ITV CM was also decreased by C6.7 (Fig.
5C). From these results, we suggest that TSP-1 in CM suppresses
c-Myc via CD47 and inhibits the cell cycle of gastric cancer
cells.
Discussion
In cancer chemotherapy, targeting stromal cells
could be advantageous because it offers a way to circumvent the
problem of tumor cell drug resistance. Moreover, the approach
differs fundamentally from conventional anti-cancer drugs that
directly attack the neoplastic cells. However, negative regulation
of cancer cell growth by stromal cells has not been well explored.
In a previous study, we found that ITV inhibited the growth of
gastric cancer cell lines when they were co-cultured with Hs738, a
gastric stromal cell line. The inhibition was greater than when the
cells were cultured alone (11). From
the results of this co-culture experiment, we suggested that ITV
inhibited cancer cell growth through direct or indirect effects on
the Hs738 cells. To explore this possibility, we first prepared CM
of Hs738 cells treated with ITV and examined its effect on cancer
cells. We found that ITV inhibited cancer cell growth through
indirect effects of Hs738 cells because ITV CM alone possessed
growth inhibitory activity. Additionally, the co-culture
experiments using cell culture inserts showed similar results
indicating that ITV inhibited cancer cell growth through effects on
Hs738 cells without direct cell-cell contact. As shown in Fig. 2A, while 1 day CM from Hs738 treated by
ITV exhibited the strongest activity for inhibition of cancer cell
growth, 2–5 day CM exhibited less stronger activity. For CM
preparation, Hs738 cells died and burst at first 2 day culture with
10 µg/ml of ITV. Thus, it is considered that CM with 10 µg/ml of
ITV contained very low growth inhibitory factors from Hs738 and
non-specific cytotoxicity of ITV (as shown in Fig. 1A) was decreased due to unexpected
interaction with some cell components from dead cells of Hs738.
However, it still has non-specific cytotoxicity of ITV in Day 1 CM
because Hs738 cells are alive at Day 1 even with the highest ITV
concentration. Hence, it is considered that the growth of gastric
cancer cells was strongly inhibited by non-specific cytotoxicity of
the highest ITV in Day 1 CM.
We next tried to identify the anti-cancer factors in
CM prepared from Hs738 cells that were maintained under serum-free
conditions. This approach was taken to minimize proteins from fetal
bovine serum that would otherwise impair proteomic analysis. The
analysis of the CM identified TSP-1 from ITV-treated stromal cells
as a candidate factor. The results also revealed that type I
collagen in ITV CM was decreased or its molecular weight was
shifted. Additionally, we demonstrated that the protein level of
α-SMA was decreased by ITV treatment. Type I collagen and α-SMA are
cancer-associated fibroblast (CAF) markers (18). Since Hs738 cells contain a mixture of
fibroblasts and myofibroblasts, this result suggested that ITV may
interfere with the differentiation of Hs738 cells into the full CAF
phenotype.
Kaur et al (15), reported that MYC gene
expression was suppressed by TSP-1 in Jurkat cells in vitro
whereas the suppression did not occur in CD47-deficient cells.
Rogers et al (19), also
reported that TSP-1-CD47 signaling decreased pulmonary c-Myc. The
oncoprotein c-Myc directly regulates the cell cycle through
transcription of its target genes, including CDK4 (17,20).
Therefore, we postulated that TSP-1 in ITV CM inhibited the growth
of gastric cancer cells through suppression of MYC
expression and the cell cycle. Our results showed that ITV CM
suppressed the expression of c-Myc protein as well as CDK4 protein
in MKN-74 cells. Moreover, the growth inhibitory activities of ITV
CM and rTSP-1 against MKN-74 cells were specifically blocked by
neutralizing antibody against TSP-1, which inhibits the interaction
with CD47. These results suggested that ITV induced TSP-1 secretion
from Hs738 cells and that the secreted TSP-1 suppressed the cell
cycle component in gastric cancer cells via CD47. The anti-TSP-1
neutralizing antibody did not completely block the inhibitory
activity of ITV CM. Thus, the effect of neutralizing antibody at
the concentration used was a little weak because the blockade
ability of the antibody against recombinant TSP-1 is equivalent to
ITV CM. However, there is also a possibility that other factors are
involved in the action of ITV. Lactic acid reportedly acts as a
regulator of TSP-1 expression via AP1, a transcriptional factor
that binds the promoter region of THBS-1 (the TSP-1 gene),
but the mode of action of ITV for induction of TSP-1 remains
unknown (21) and requires further
investigation.
Our data showed that ITV possesses a novel
anti-cancer activity that is exerted through modulation of stromal
cells. Interestingly, it has been found that TSP-1 can promote
invasion, migration and proliferation of breast cancer cells
(22–24). Although we demonstrated that TSP-1
negatively regulated gastric cancer cells via gastric stromal cells
and that it was induced by ITV treatment, it is likely that it is
the balance between positive and negative regulations of cancer
cells that is important for TSP-1 activity.
Acknowledgements
The authors would like to thank Dr. D. Tatsuda and
Mr. M. Amemiya for helpful comments, Dr. R. Sawa and Ms. Y.
Takahashi for analytical measurements and Ms. C. Sakashita for
technical assistance with chemical synthesis [all from Institute of
Microbial Chemistry (BIKAKEN), Tokyo, Microbial Chemistry Research
Foundation].
Funding
The present study was supported by JSPS KAKENHI
(grant no. JP15K06888) and the Project for Cancer Research And
Therapeutic Evolution (P-CREATE) from the Japan Agency for Medical
Research and development, AMED (grant no. 17cm0106213h0002).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JY and KM designed the experiments. HA and TW
synthesized and provided ITV. JY performed the study and analyzed
the data. KM reviewed the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ITV
|
intervenolin
|
|
CM
|
conditioned media
|
|
ITV CM
|
the CM taken from human gastric
fibroblast-like Hs738 cells treated with ITV
|
|
Ctrl CM
|
CM from non-treated-Hs738
|
|
TSP-1
|
thrombospondin-1
|
|
stromal cells
|
fibroblast-like stromal cells
|
|
α-SMA
|
α-smooth muscle actin
|
References
|
1
|
Subramaniam KS, Tham ST, Mohamed Z, Woo
YL, Adenan Mat NA and Chung I: Cancer-associated fibroblasts
promote proliferation of endometrial cancer cells. PLoS One.
8:e689232013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vermeulen L, De Sousa E, Melo F, van der
Heijden M, Cameron K, de Jong JH, Borovski T, Tuynman JB, Todaro M,
Merz C, Rodermond H, et al: Wnt activity defines colon cancer stem
cells and is regulated by the microenvironment. Nat Cell Biol.
12:468–476. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Quante M, Tu SP, Tomita H, Gonda T, Wang
SS, Takashi S, Baik GH, Shibata W, Diprete B, Betz KS, et al: Bone
marrow-derived myofibroblasts contribute to the mesenchymal stem
cell niche and promote tumor growth. Cancer Cell. 19:257–272. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Malanchi I, Santamaria-Martinez A, Susanto
E, Peng H, Lehr HA, Delaloye JF and Huelsken J: Interactions
between cancer stem cells and their niche govern metastatic
colonization. Nature. 481:85–89. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wilson TR, Fridlyand J, Yan Y, Penuel E,
Burton L, Chan E, Peng J, Lin E, Wang Y, Sosman J, et al:
Widespread potential for growth-factor-driven resistance to
anticancer kinase inhibitors. Nature. 487:505–509. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumari N, Dwarakanath BS, Das A and Bhatt
AN: Role of interleukin-6 in cancer progression and therapeutic
resistance. Tumour Biol. 37:11553–11572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin SY, Jian FY, Chen YH, Chien SC, Hsieh
MC, Hsiao PW, Lee WH, Kuo YH and Yang N: Induction of IL-25
secretion from tumour-associated fibroblasts suppresses mammary
tumour metastasis. Nat Commun. 7:113112016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawada M, Inoue H, Usami I and Ikeda D:
Phthoxazolin A inhibits prostate cancer growth by modulating
tumor-stromal cell interactions. Cancer Sci. 100:150–157. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kawada M, Inoue H, Ohba S, Masuda T,
Momose I and Ikeda D: Leucinostatin A inhibits prostate cancer
growth through reduction of insulin-like growth factor-I expression
in prostate stromal cells. Int J Cancer. 126:810–818.
2010.PubMed/NCBI
|
|
10
|
Kawada M, Inoue H, Ohba S, Yoshida J,
Masuda T, Yamasaki M, Usami I, Sakamoto S, Abe H, Watanabe T, et
al: Stromal cells positively and negatively modulate the growth of
cancer cells: Stimulation via the PGE2-TNFα-IL-6 pathway and
inhibition via secreted GAPDH-E-cadherin interaction. PLoS One.
10:e01194152015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kawada M, Inoue H, Ohba S, Hatano M,
Amemiya M, Hayashi C, Usami I, Abe H, Watanabe T, Kinoshita N, et
al: Intervenolin, a new antitumor compound with anti-Helicobacter
pylori activity, from Nocardia sp. Ml96-86F2. J Antibiot (Tokyo).
66:543–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rodriguez-Manzaneque JC, Lane TF, Ortega
MA, Hynes RO, Lawler J and Iruela-Arispe Ml: Thrombospondin-1
suppresses spontaneous tumor growth and inhibits activation of
matrix metalloproteinase-9 and mobilization of vascular endothelial
growth factor. Proc Natl Acad Sci USA. 98:12485–12490. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawada M, Masuda T, Ishizuka M and
Takeuchi T: 15-Deoxyspergualin inhibits Akt kinase activation and
phosphatidylcholine synthesis. J Biol Chem. 277:27765–27771. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fukazawa H, Mizuno S and Uehara Y: A
microplate assay for quantitation of anchorage-independent growth
of transformed cells. Anal Biochem. 228:83–90. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaur S, Soto-Pantoja DR, Stein EV, Liu C,
Elkahloun AG, Pendrak Ml, Nicolae A, Singh SP, Nie Z, Levens D, et
al: Thrombospondin-1 signaling through CD47 inhibits self-renewal
by regulating c-Myc and other stem cell transcription factors. Sci
Rep. 3:16732013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
de Marval Miliani PL, Macias E, Rounbehler
R, Sicinski P, Kiyokawa H, Johnson DG, Conti CJ and
Rodriguez-Puebla Ml: Lack of cyclin-dependent kinase 4 inhibits
c-myc tumorigenic activities in epithelial tissues. Mol Cell Biol.
24:7538–7547. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu S, Yin X, Fang X, Zheng J, Li L, Liu X
and Chu L: c-MYC responds to glucose deprivation in a
cell-type-dependent manner. Cell Death Discov. 1:150572015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Togo S, Polanska UM, Horimoto Y and Orimo
A: Carcinoma-associated fibroblasts are a promising therapeutic
target. Cancers. 5:149–169. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rogers NM, Sharifi-Sanjani M, Yao M,
Ghimire K, Bienes-Martinez R, Mutchler SM, Knupp HE, Baust J,
Novelli EM, Ross M, et al: TSP1-CD47 signaling is upregulated in
clinical pulmonary hypertension and contributes to pulmonary
arterial vasculopathy and dysfunction. Cardiovasc Res. 113:15–29.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schmidt EV: The role of c-myc in cellular
growth control. Oncogene. 18:2988–2996. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seliger C, Leukel P, Moeckel S, Jachnik B,
Lottaz C, Kreutz M, Brawanski A, Proescholdt M, Bogdahn U,
Bosserhoff AK, et al: Lactate-modulated induction of THBS-1
activates transforming growth factor (TGF)-beta2 and migration of
glioma cells in vitro. PLoS One. 8:e789352013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Albo D, Rothman VL, Roberts DD and
Tuszynski GP: Tumour cell thrombospondin-1 regulates tumour cell
adhesion and invasion through the urokinase plasminogen activator
receptor. Br J Cancer. 83:298–306. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hyder SM, Liang Y and Wu J: Estrogen
regulation of thrombospondin-1 in human breast cancer cells. Int J
Cancer. 125:1045–1053. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yee KO, Connolly CM, Duquette M,
Kazerounian S, Washington R and Lawler J: The effect of
thrombospondin-1 on breast cancer metastasis. Breast Cancer Res
Treat. 114:85–96. 2009. View Article : Google Scholar : PubMed/NCBI
|