Introduction
Multiple myeloma is a B-cell malignancy
characterized by abnormal plasmacyte proliferation in the bone
marrow (1). The morbidity
contribution of multiple myeloma is ~1% of all tumors and 10% of
blood system tumors (1,2). Multiple myeloma is the second most
common malignancy of the blood system. An Australian study
identified that the survival time for the majority of cases was
between 5 and 8 years in elderly patients, accounting for 20% of
hematological mortalities (2).
Although various treatments have been tested, including
thalidomide, lenalidomide, bortezomib and hematopoietic stem cell
transplantation, multiple myeloma remains an incurable blood cell
cancer (3,4). Therefore, the development of more
effective antitumor drugs and novel therapeutic strategies for
multiple myeloma is urgently required.
Curcumin is a polyphenol derived from the rhizome of
Curcuma, and has long been used for treating various types
of cancer, including colorectal carcinoma, glioma and lung cancer,
by inhibiting the proliferation and promoting the apoptosis of the
cancer cells (5–9). As a potential cancer inhibitor, curcumin
inhibits almost every phase of tumor development, including
infiltration, colonization and outgrowth. Previous studies have
demonstrated that the antitumor function of curcumin is associated
with a range of genetic and epigenetic alterations of oncogenes and
anti-oncogenes, including the RAC-α serine/threonine-protein kinase
(AKT)/mammalian target of rapamycin (mTOR) pathway components and
downstream signaling molecules (10,11).
mTOR is known as the main regulatory factor of cell
autophagy and apoptosis (10,11), which are involved in the regulation of
various cellular processes. Clinical studies have demonstrated that
the inhibition of mTOR activation can promote cell apoptosis in
several types of cancer, including pancreatic cancer, bladder
cancer and leukemia (12–15). Furthermore, inhibition of the mTOR
signaling pathway can enhance cytotoxicity and induce autophagy in
a possible protective role in K562 leukemia cell lines and in
chronic myeloid leukemia (16).
Resveratrol, a natural phytoalexin, inhibits the mTOR pathway and
induces cell apoptosis and autophagy in multiple myeloma (17).
Previous studies have suggested that curcumin, a
constituent of Curcuma longa, is a potential epigenetic
regulator. This polyphenol inhibits the activity of histone
acetyltransferase, which can lead to inhibition of histone
acetylation (18–20). Curcumin also blocks the catalytic
C1226 site of DNA methyltransferase (DNMT)1 through its chemical
structure to inhibit the activity of the enzyme (21). However, the curcumin-induced
epigenetic regulation of biomarkers in multiple myeloma has not yet
been fully understood, and specifically, whether mTOR serves a role
in epigenetic regulation remains unknown. In the present study,
curcumin was revealed to induce the methylation of the mTOR
promoter, potentially via the upregulation of DNMT3a and
DNMT3b.
Materials and methods
Cell culture and treatment
The NCI-H929 and RPMI-8226 cell lines were purchased
from the American Type Culture Collection bio-resource center
(Manassas, VA, USA). All cells were suspended, cultured and
maintained in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100
µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in a humidified 5% CO2 atmosphere. Curcumin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was dissolved in
dimethylsulfoxide (DMSO; Beyotime Institute of Biotechnology,
Haimen, China) as stock solution (1 mg/ml).
For the proliferation assay, the cells were cultured
in 96-well plates at 5×104 cells/well and treated with
increasing concentrations (0, 5, 10, 15 and 20 µM) of curcumin. At
various time points (0, 24, 48, 72, 96 and 120 h), 10 µl MTT
solution (Beyotime Institute of Biotechnology, Haimen, China) was
added to each well, then the cells were incubated for 4 h at 37°C
in a humidified 5% CO2 atmosphere. The formazan crystals
that formed were dissolved with 100 µl DMSO for 2 h at 37°C. The
absorbance values were measured using the Multiskan™ FC Microplate
Photometer (Thermo Fisher Scientific, Inc.) at a wavelength of 570
nm.
For the apoptosis assay, ~2.5×106 cells
cultured in 6-well plates were treated with 0.1% DMSO or 10 µM
curcumin. Following 24 h of treatment, the cells were collected at
500 × g for 5 min at room temperature, washed and stained using an
Annexin V-Fluorescein Isothiocyanate (FITC)/Propidium Iodide (PI)
kit (BD Biosciences, Franklin Lakes, NJ, USA). Briefly, the cells
were resuspended in 1X binding buffer to a concentration of
106 cells/ml. Volumes of 1 ml were transferred to new
tubes and 5 µl FITC-annexin V was added prior to incubation for 15
min at room temperature, and 5 µl PI solution was added prior to
further incubation for 5 min at room temperature. The entire
process was protected from light. Stained cells were analyzed by
the FACSCanto II flow cytometer and BD FACSDiva software (version
6.1.3; BD Biosciences).
For all other assays, NCI-H929 cells were used as
they were identified to be more sensitive to curcumin compared with
RPMI-8226 cells. Cells were treated with 0.1% DMSO, 10 µM curcumin
or 2.5 µM 5-aza-2′-deoxycytidine (5-aza-CdR), a demethylation
reagent, with 10% FBS-containing medium for 48 h, and were
harvested for DNA, RNA and protein analysis.
Western blotting
Treated cells were harvested and lysed using
ice-cold radioimmunoprecipitation assay lysis and extraction buffer
(Thermo Fisher Scientific, Inc.) containing a protein inhibitor
cocktail (cat. no. P2714; Sigma-Aldrich; Merck KGaA). The protein
concentration was determined using a Pierce Rapid Gold
Bicinchoninic Acid Protein assay kit (Thermo Fisher Scientific,
Inc.) and a NanoDrop 2000 instrument (Thermo Fisher Scientific,
Inc.). The proteins (30 µg) were separated by SDS-PAGE (12.5% gel)
and transferred to a polyvinylidene difluoride membrane.
Subsequently, the membrane was blocked with 5% fat-free milk in
Tris-buffered saline and Tween-20, and sequentially incubated
overnight at 4°C with 1:500 diluted primary antibodies and 2 h at
room temperature with 1:2,000 diluted horseradish
peroxidase-conjugated secondary antibodies (ab6721; Abcam,
Cambridge, UK). The signals were detected with the SuperSignal
enhanced chemiluminescence detection system (Thermo Fisher
Scientific, Inc.) and recorded with the Gel Documentation 2000
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Rabbit
polyclonal primary antibodies against human methyl CpG-binding
protein 2 (MECP2; ab2828), DNMT1 (ab19905), DNMT3a (ab2850), DNMT3b
(ab16049) and β-actin (ab8227) were purchased from Abcam.
DNA extraction and bisulfite genomic
sequencing
Genomic DNA was isolated from ~2.5×106
treated cells using the QIAamp DNA Mini kit (Qiagen GmbH, Hilden,
Germany). Bisulfite-modification of the genomic DNA was performed
using the CpGenome™ Fast DNA Modification kit (Merck KGaA). The
promoter region of mTOR, which contains a CpG island
(chr1:11262153-11263153), was divided into two for bisulfite PCR.
Primers for upstream of the first exon of mTOR were as follows:
5′-GTGGTTGTGATAGGTAAAAGATT-3′ (forward) and
5′-AACCTAACACAACCCCTCTAAA-3′ (reverse); primers for downstream of
the first exon of mTOR were as follows: 5′-GAGGGAAGGAGGGTTTTTA-3′
(forward) and 5′-CTTTTAATACAATAATTCCTAAACACC-3′ (reverse). The PCR
products were purified and cloned into TOPO vectors (Invitrogen;
Thermo Fisher Scientific, Inc.) containing a T7 promoter near the
insert site for sequencing, using a T7 primer
(5′-TAATACGACTCACTATAGGG-3′). The overall percentage of the
methylation was calculated by dividing the number of methylated
CpGs by the number of total CpGs. The global DNA methylation of
treated cells was determined using a Methylamp Global DNA
Methylation Quantification kit (EpiGentek, Farmingdale, NY,
USA).
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the treated cells using
the RNeasy Mini kit (Qiagen GmbH). First-strand cDNA was
synthesized from 1 µg total RNA using the SuperScript III
First-Strand Synthesis system for RT-PCR (Invitrogen; Thermo Fisher
Scientific, Inc.). The cDNA was used as the template for qPCR
(ABI7500; Applied Biosystems; Thermo Fisher Scientific, Inc.). The
sequences of the primers used for cDNA amplification were: mTOR
5′-CGCTGTCATCCCTTTATCG-3′ (forward) and 5′-ATGCTCAAACACCTCCACC-3′
(reverse), and β-actin 5′-CTCCATCCTGGCCTCGCTGT-3′ (forward) and
5′-GCTGTCACCTTCACCGTTCC-3′ (reverse). A Power SYBR™ Green Master
kit (Invitrogen; Thermo Fisher Scientific, Inc.) was used to detect
the relative expression of mTOR compared with β-actin using the
ΔΔCq method (22). The thermocycling
conditions were preheating at 50°C for 2 min, denaturation at 95°C
for 10 min, and 40 amplification cycles of denaturation at 95°C for
15 sec, and annealing and extension at 60°C for 60 sec.
Statistical analysis
From the apoptosis, qPCR and western blot assays,
the data are presented as the mean ± standard deviation from 3
independent experiments with duplication of readings. The control
and multiple experimental groups were compared using one-way
analysis of variance with Dunnett's post hoc analysis. The flow
cytometry data between two groups were compared using Student's
t-test. All statistics were using the SPSS software, version 17.0
(SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference.
Results
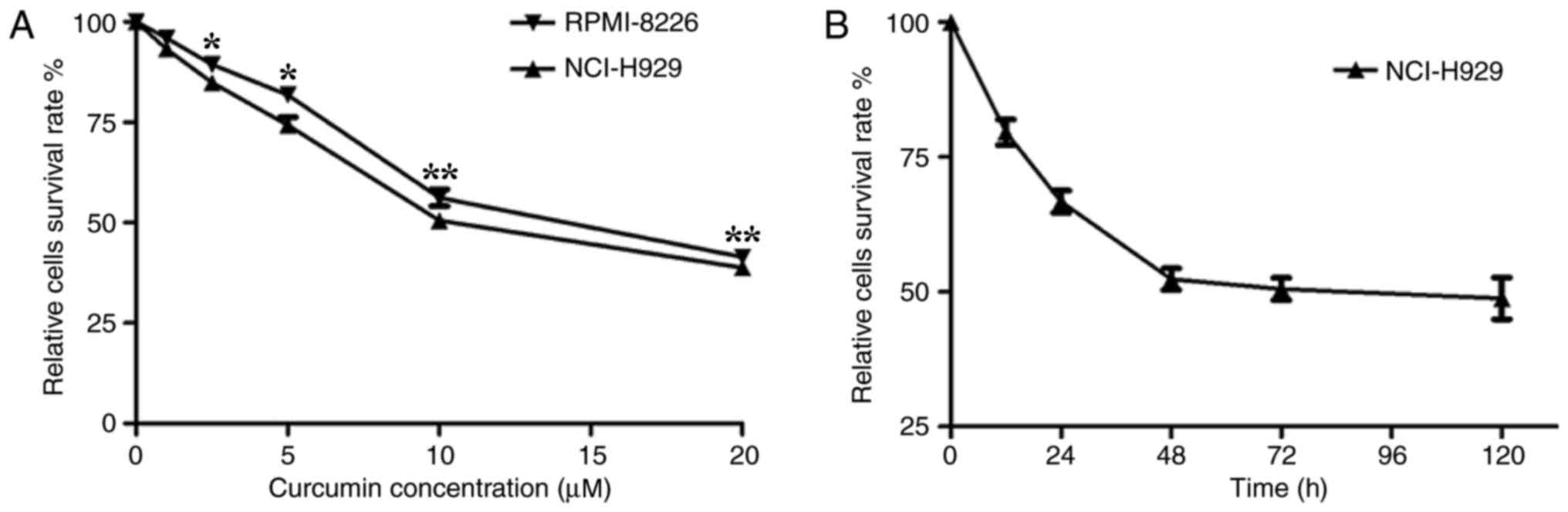
Curcumin suppresses the proliferation
of multiple myeloma cell lines
It has previously been demonstrated that curcumin
influences tumor cell proliferation, apoptosis, migration and
invasion, resulting in an antitumor effect (6–11). To
examine the effective inhibitory concentration (IC) of curcumin in
multiple myeloma cells, an MTT assay was first performed using the
human myeloma NCI-H929 and RPMI-8226 cell lines treated with
various concentrations of curcumin. The results revealed that
curcumin significantly (P<0.05) suppressed the proliferation of
the cells in a dose-dependent manner. The curcumin treatment
resulted in a high level of cytotoxicity when the concentration
reached 5 µM and the highest level was at 20 µM. The half-maximal
IC value of curcumin for these human myeloma cells was ~10 µM
(Fig. 1A). Compared with the
RPMI-8226 cell line, the NCI-H929 cell line was more sensitive to
curcumin. Therefore, the subsequent experiments were performed
using the NCI-H929 cells, whereby 10 µM curcumin was added to the
culture medium for different time periods. The MTT assay
demonstrated that the number of NCI-H929 cells stabilized when
treated with 10 µM curcumin for 48 h or longer (Fig. 1B). These results suggested that the
effect of curcumin on NCI-H929 cells was stable after 48 h,
determining the time periods chosen for the following
experiments.
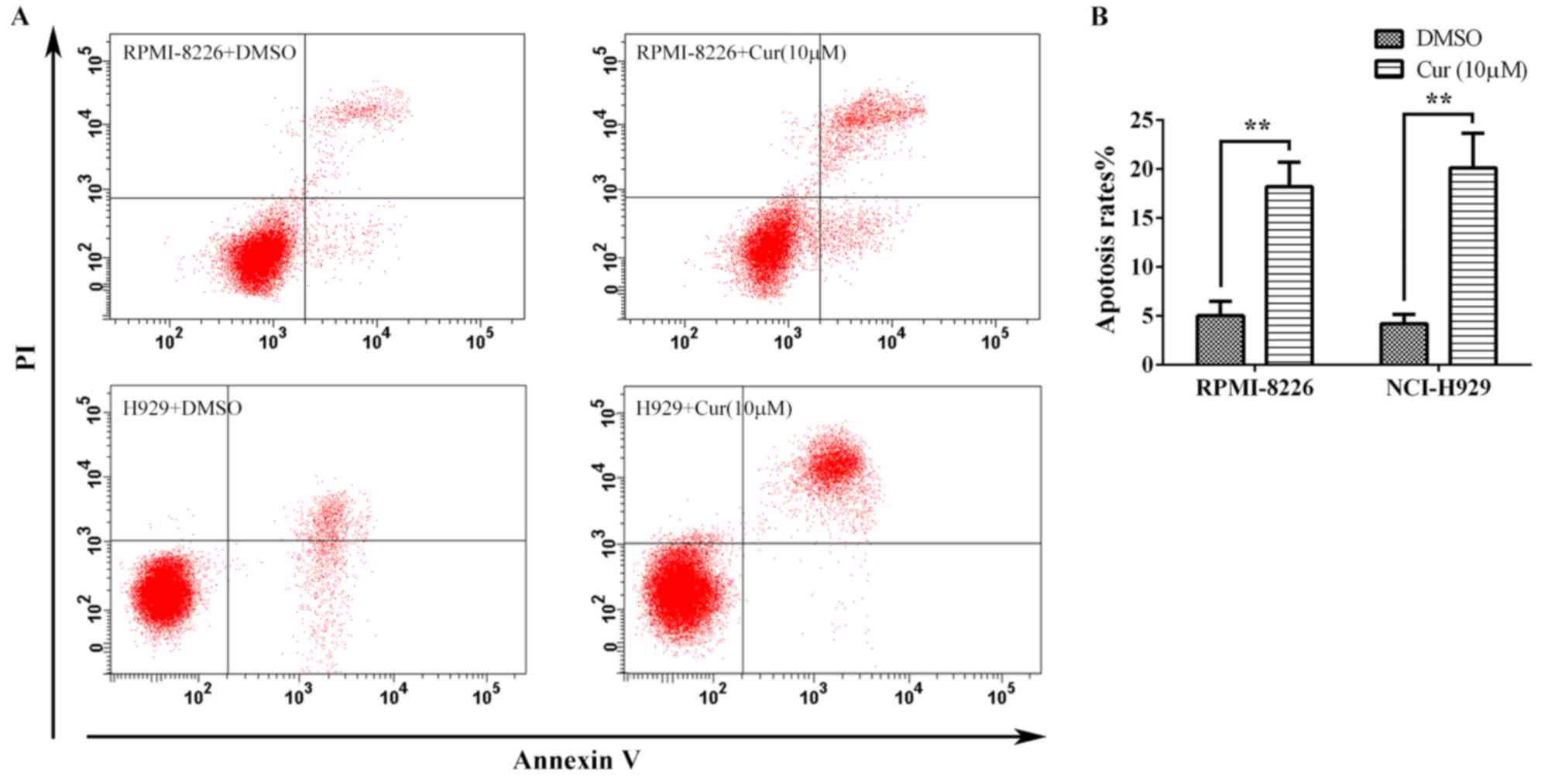
Curcumin induces apoptosis in multiple
myeloma cell lines
To further assess the antitumor effect of curcumin,
the multiple myeloma cells were treated with 10 µM curcumin for 24
h, and the apoptosis was detected by flow cytometry following
Annexin V-FITC/PI staining (Fig. 2A).
The apoptosis rates calculated from the RPMI-8226 and NCI-H929
cells were 18.22±2.50 and 20.13±3.55%, respectively, which were
significant increases compared with the rates for the corresponding
untreated controls (5.03±1.46 and 4.20±0.95%, respectively)
(Fig. 2B).
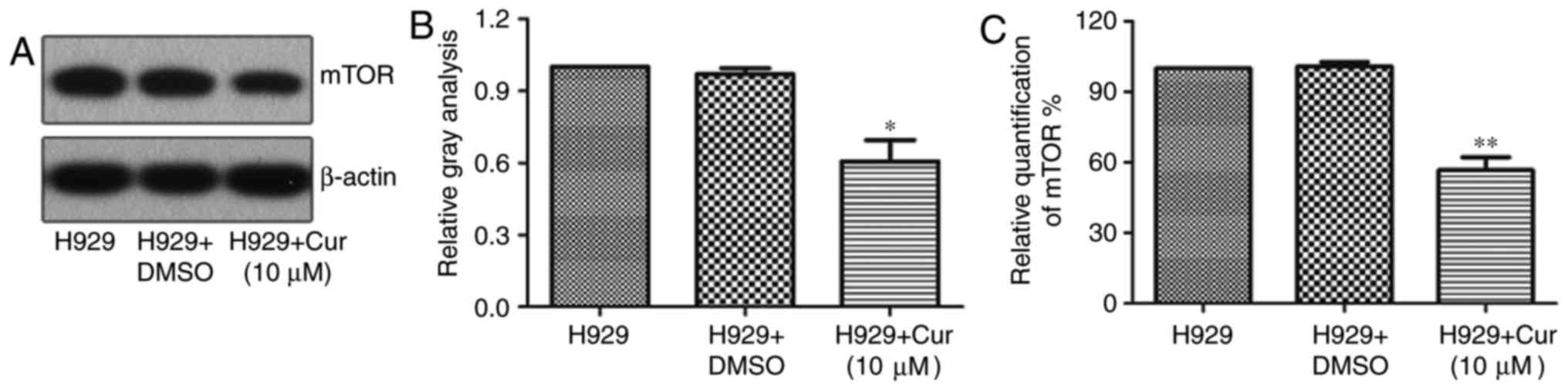
Curcumin inhibits the expression of
mTOR in NCI-H929 cells
In a variety of cancer cells, curcumin can induce
apoptosis or autophagy by downregulating the expression of mTOR
(23,24). To verify whether curcumin treatment
also downregulates the expression of mTOR in the NCI-H929 cell
line, the protein expression of mTOR was examined by western
blotting and was revealed to be 39.34% lower with the curcumin
treatment than with DMSO treatment (Fig.
3A and B). Furthermore, the RT-qPCR results showed that mTOR
mRNA was 43.31% lower in NCI-H929 cells treated with 10 µM curcumin
for 48 h compared with that in the untreated control (Fig. 3C), implying that curcumin inhibits the
expression of mTOR by repressing its transcriptional
regulation.
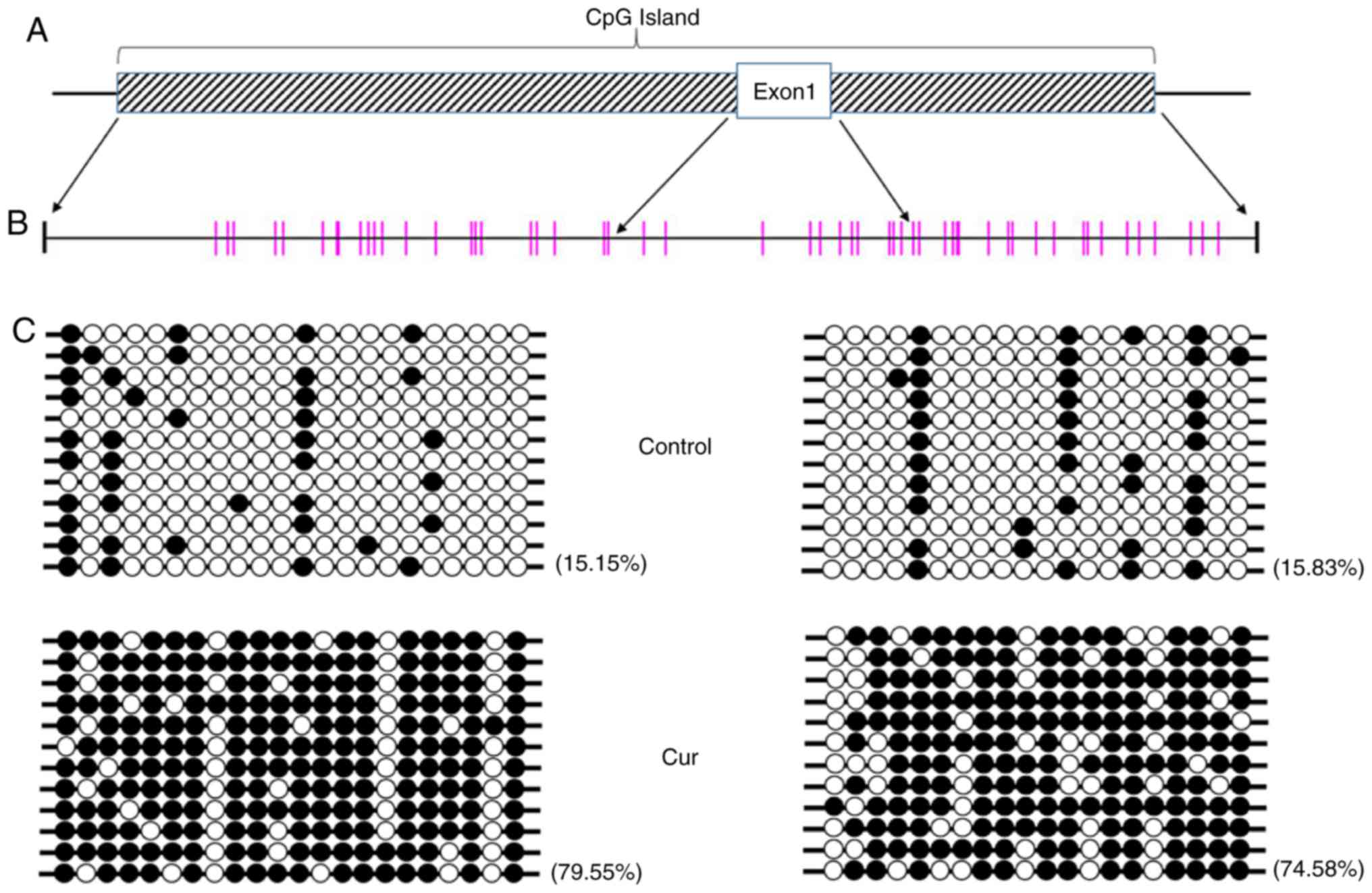
Curcumin treatment leads to
hypermethylation of the mTOR promoter region
Curcumin has the potential for DNA methylation
regulation, and the transcription of associated genes may be
directly affected when DNA methylation is altered (25,26). To
further determine whether a change in methylation levels was
induced by curcumin, the DNA methylation status of the whole genome
of NCI-H929 cells treated with 10 µM curcumin was determined using
the Global DNA Methylation Quantification Ultra kit, and no
significant difference was noted compared with the control group
(data not shown), indicating that no significant change in DNA
methylation on the genomic level occurs following curcumin
treatment. However, further bisulfide sequencing revealed that the
CpG island in the mTOR promoter was in a hypermethylated state
(Fig. 4).
Curcumin upregulates the expression of
DNMT3
Alterations in DNA methylation are mediated by
DNMTs. In order to reveal which DNMT participates in the
curcumin-induced hypermethylation of the mTOR promoter, the
NCI-H929 cells were treated with curcumin and DNA methylation
inhibitor 5-aza-CdR for 48 h, and changes in the expression levels
of various DNMTs were detected by western blotting (Fig. 5). The levels of the DNMTs were
decreased to various degrees in the NCI-H929 cells treated with
5-aza-CdR. In the curcumin-treated cells, the expression of DNMT3a
and DNMT3b, two key methylation transferases of de novo
methylation, was significantly upregulated compared with that in
the control group. However, there were no significant changes in
the expression levels of maintenance methylase DNMT1 and
methylation-binding protein MECP2 following treatment with
curcumin.
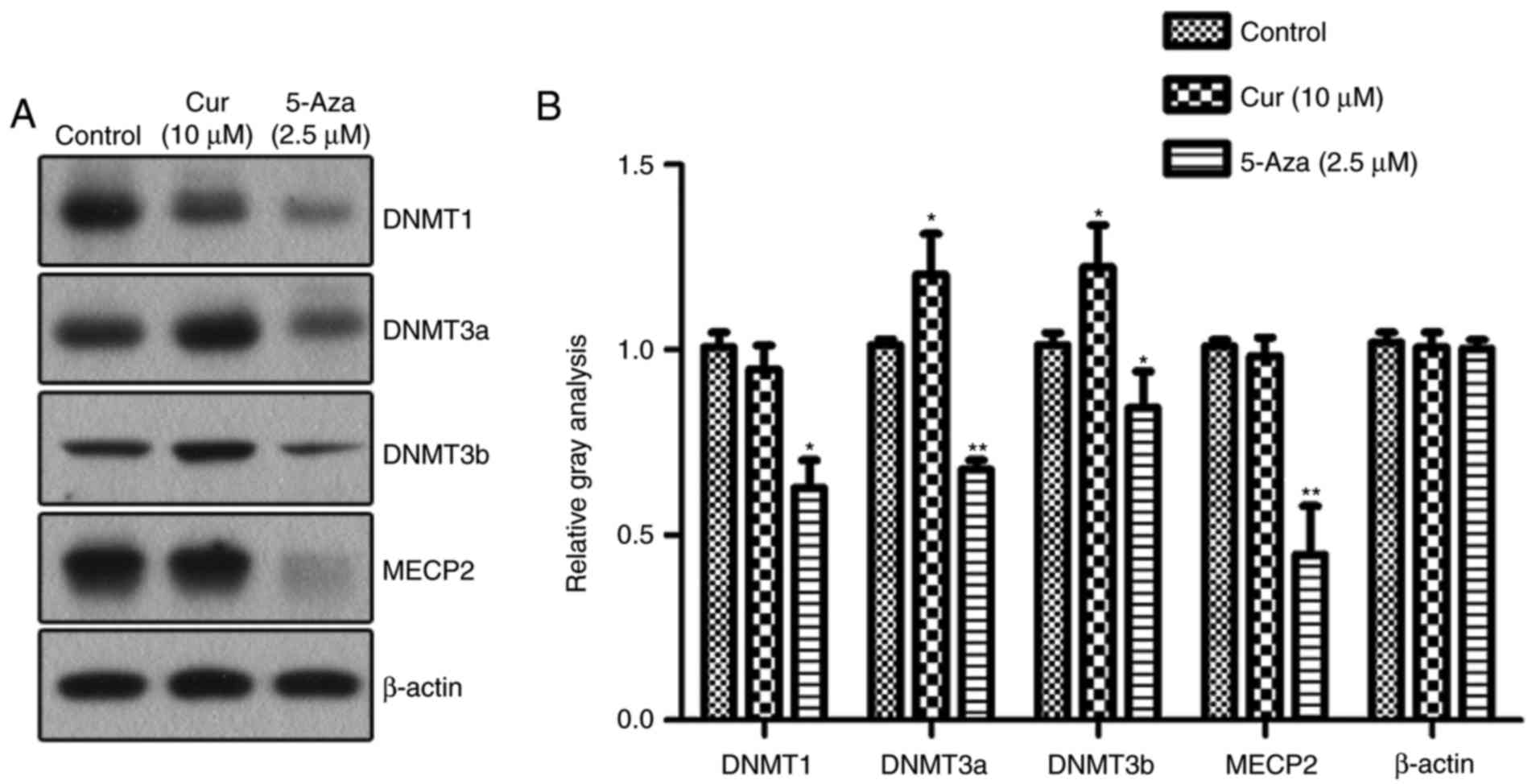 | Figure 5.Expression of DNMTs induced by
curcumin. (A) Western blotting detected the expression levels of
DNMT1, DNMT3a, DNMT3b and MECP2 in NCI-H929 cells following various
treatments. (B) Quantitative analysis of the protein levels of
DNMT1, DNMT3a, DNMT3b and MECP2, normalized to β-actin (n=3).
*P<0.05 and **P<0.01, vs. the control group, as calculated
using analysis of variance. DNMT, DNA methyltransferase; MECP2,
methyl CpG binding protein 2; Cur, curcumin; 5-Aza,
5-aza-2′-deoxycytidine. |
Discussion
The results of the present study revealed that the
expression of mTOR and its promoter methylation in myeloma cells
were altered by curcumin, and that this hypermethylation may
potentially have been mediated by the upregulation of DNMT3.
Curcumin was able to induce apoptosis in 50% of the myeloma cells
when its concentration was increased to 10 µM. Investigation of the
effect of curcumin on normal bone marrow cells was not performed;
however, these results suggest that curcumin may be used in
anti-multiple myeloma treatment. Notably, the results indicated
that there were no widespread changes in genomic DNA methylation
induced by curcumin in NCI-H929 cells, in accordance with the
results from colorectal cells in a study by Link et al
(25). The present study focused on
mTOR, a key factor that activates apoptosis and autophagy pathways,
rather than performing macroscopic genetic clustering analysis.
Lower expression of mTOR and higher promoter methylation were
observed, which may be due to changes in DNMT3 expression.
Curcumin is a plant polyphenol extracted from the
roots of a plant from the Curcuma genus, and has numerous
pharmacological effects, including antitumor, anti-inflammatory,
antioxidant and antibacterial properties (27,28).
Curcumin may affect cell transcription and regulate apoptosis and
autophagy by modulating multiple cell signals, including the
nuclear factor-κB, phosphatidylinositol-3-kinase/AKT pathway, the
Janus tyrosine kinase/signal transducer and activator of
transcription (STAT) signaling transduction pathway and STAT3
(23,29,30). A
previous experimental and epidemiological study have proposed that
curcumin may alter the DNA methylation status of tumor cells
(25); however, its ability to
regulate DNA methylase in myeloma cells remains unknown. The
present study systematically examined the effect of curcumin on
DNMTs in multiple myeloma NCI-H929 cells. To detect the epigenetic
regulatory effect of curcumin, 5-aza-CdR was used as a positive
control for comparison. Curcumin did not inhibit the expression of
methyl-DNA binding protein MECP2 and the maintenance methylase
DNMT1 in the NCI-H929 cells, in accordance with the results of the
study by Shu et al (31) in
LNcaP cells, suggesting that curcumin has no effect on the
maintenance of methylation. It has been reported that curcumin is
an inhibitor of DNMT1 and may cause a decrease in the overall DNA
methylation level in the MV4-11 lymphoma cell line (21). However, in the present study, curcumin
was not observed to have an effect on the DNMT1 expression in
NCI-H929 cells, but rather it led to an increase in the expression
of DNMT3a and DNMT3b. These differences may be due to several
factors, including the types of cell lines, the curcumin
concentration and the duration of treatment.
In summary, the present study demonstrated that the
downregulation of mTOR was associated with hypermethylation of its
promoter following treatment with curcumin, which may occur through
regulating the expression of DNMT3. It may be concluded that
curcumin possesses anti-multiple myeloma activity, which is
different from that of chemotherapeutic drugs, including 5-aza-CdR,
that cause changes in the overall level of genomic DNA methylation.
The precise sites of DNMT3a and DNMT3b that regulate the mTOR
promoter and affect its expression should be identified and
verified in future studies.
Acknowledgements
The authors would like to thank Mrs. Baoshan Huang
(Wenzhou Medical University, Wenzhou, Zhejiang, China) for
assisting with the western blot analysis, and Mr. Lingyun Li, Mr.
Zhenqiang Huang and Mr. Youfa Ding (Clinical Laboratory, Lishui
People's Hospital, Lishui, Zhejiang, China) for their assistance
with the experiments.
Funding
The present study was supported by Public Projects
of Lishui (grant nos. 2014JYZB03 and 2014JYZB16) provided by the
Lishui Technology Bureau.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BF designed the research. JC, YY, HZ, JJ and CQ
performed the experiments. JC, TZ, YY and CQ analyzed the data. BF
and JC contributed reagents/materials. JC was a major contributor
in writing the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
mTOR
|
mammalian target of rapamycin
|
|
DMNT
|
DNA methyltransferase
|
|
MECP2
|
methyl CpG-binding protein 2
|
References
|
1
|
Dimopoulos MA and Terpos E: Multiple
myeloma. Ann Oncol. 21 Suppl 7:vii143–vii150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Quach H, Prince HM and Spencer A: Managing
multiple myeloma in the elderly: Are we making progress? Expert Rev
Hematol. 4:301–315. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mateos MV, Masszi T, Grzasko N, Hansson M,
Sandhu I, Pour L, Viterbo L, Jackson SR, Stoppa AM, Gimsing P, et
al: Impact of prior therapy on the efficacy and safety of oral
ixazomib-lenalidomide-dexamethasone vs.
placebo-lenalidomide-dexamethasone in patients with
relapsed/refractory multiple myeloma in TOURMALINE-MM1.
Haematologica. 102:1767–1775. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moreau P, Masszi T, Grzasko N, Bahlis NJ,
Hansson M, Pour L, Sandhu I, Ganly P, Baker BW, Jackson SR, et al:
Oral Ixazomib, Lenalidomide, and Dexamethasone for Multiple
Myeloma. N Engl J Med. 374:1621–1634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Howells LM, Mitra A and Manson MM:
Comparison of oxaliplatin- and curcumin-mediated antiproliferative
effects in colorectal cell lines. Int J Cancer. 121:175–183. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jin G, Yang Y, Liu K, Zhao J, Chen X, Liu
H, Bai R, Li X, Jiang Y, Zhang X, et al: Combination curcumin and
(−)-epigallocatechin-3-gallate inhibits colorectal carcinoma
microenvironment-induced angiogenesis by JAK/STAT3/IL-8 pathway.
Oncogenesis. 6:e3842017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weissenberger J, Priester M, Bernreuther
C, Rakel S, Glatzel M, Seifert V and Kögel D: Dietary curcumin
attenuates glioma growth in a syngeneic mouse model by inhibition
of the JAK1,2/STAT3 signaling pathway. Clin Cancer Res.
16:5781–5795. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zanotto-Filho A, Braganhol E, Klafke K,
Figueiró F, Terra SR, Paludo FJ, Morrone M, Bristot IJ, Battastini
AM, Forcelini CM, et al: Autophagy inhibition improves the efficacy
of curcumin/temozolomide combination therapy in glioblastomas.
Cancer Lett. 358:220–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ye M and Zhang J and Zhang J, Miao Q, Yao
L and Zhang J: Curcumin promotes apoptosis by activating the
p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer.
Cancer Lett. 357:196–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Srivastava RK, Chen Q, Siddiqui I, Sarva K
and Shankar S: Linkage of curcumin-induced cell cycle arrest and
apoptosis by cyclin-dependent kinase inhibitor p21(/WAF1/CIP1).
Cell Cycle. 6:2953–2961. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shinojima N, Yokoyama T, Kondo Y and Kondo
S: Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in
curcumin-induced autophagy. Autophagy. 3:635–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao P, Maira SM, García-Echeverría C and
Hedley DW: Activity of a novel, dual PI3-kinase/mTor inhibitor
NVP-BEZ235 against primary human pancreatic cancers grown as
orthotopic xenografts. Br J Cancer. 100:1267–1276. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nassim R, Mansure JJ, Chevalier S, Cury F
and Kassouf W: Combining mTOR inhibition with radiation improves
antitumor activity in bladder cancer cells in vitro and in vivo: A
novel strategy for treatment. PLoS One. 8:e652572013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tasian SK, Teachey DT, Li Y, Shen F,
Harvey RC, Chen IM, Ryan T, Vincent TL, Willman CL, Perl AE, et al:
Potent efficacy of combined PI3K/mTOR and JAK or ABL inhibition in
murine xenograft models of Ph-like acute lymphoblastic leukemia.
Blood. 129:177–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zoncu R, Efeyan A and Sabatini DM: mTOR:
From growth signal integration to cancer, diabetes and ageing. Nat
Rev Mol Cell Biol. 12:21–35. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suangtamai T and Tanyong DI: Diallyl
disulfide induces apoptosis and autophagy via mTOR pathway in
myeloid leukemic cell line. Tumour Biol. 37:10993–10999. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang FM, Galson DL, Roodman GD and Ouyang
H: Resveratrol triggers the pro-apoptotic endoplasmic reticulum
stress response and represses pro-survival XBP1 signaling in human
multiple myeloma cells. Exp Hematol. 39:999–1006. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Balasubramanyam K, Varier RA, Altaf M,
Swaminathan V, Siddappa NB, Ranga U and Kundu TK: Curcumin, a novel
p300/CREB-binding protein-specific inhibitor of acetyltransferase,
represses the acetylation of histone/nonhistone proteins and
histone acetyltransferase-dependent chromatin transcription. J Biol
Chem. 279:51163–51171. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kang J, Chen J, Shi Y, Jia J and Zhang Y:
Curcumin-induced histone hypoacetylation: The role of reactive
oxygen species. Biochem Pharmacol. 69:1205–1213. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang SK, Cha SH and Jeon HG:
Curcumin-induced histone hypoacetylation enhances
caspase-3-dependent glioma cell death and neurogenesis of neural
progenitor cells. Stem Cells Dev. 15:165–174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Xie Z, Jones W, Pavlovicz RE, Liu
S, Yu J, Li PK, Lin J, Fuchs JR, Marcucci G, et al: Curcumin is a
potent DNA hypomethylation agent. Bioorg Med Chem Lett. 19:706–709.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo Y, Shan Q, Gong Y, Lin J, Shi F, Shi R
and Yang X: Curcumin induces apoptosis via simultaneously targeting
AKT/mTOR and RAF/MEK/ERK survival signaling pathways in human
leukemia THP-1 cells. Pharmazie. 69:229–233. 2014.PubMed/NCBI
|
|
24
|
Zhao G, Han X, Zheng S, Li Z, Sha Y, Ni J,
Sun Z, Qiao S and Song Z: Curcumin induces autophagy, inhibits
proliferation and invasion by downregulating AKT/mTOR signaling
pathway in human melanoma cells. Oncol Rep. 35:1065–1074. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Link A, Balaguer F, Shen Y, Lozano JJ,
Leung HC, Boland CR and Goel A: Curcumin modulates DNA methylation
in colorectal cancer cells. PLoS One. 8:e577092013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu P, Huang R, Xiong YL and Wu C:
Protective effects of curcumin against liver fibrosis through
modulating DNA methylation. Chin J Nat Med. 14:255–264.
2016.PubMed/NCBI
|
|
27
|
Agrawal DK and Mishra PK: Curcumin and its
analogues: Potential anticancer agents. Med Res Rev. 30:818–860.
2010.PubMed/NCBI
|
|
28
|
Naksuriya O, Okonogi S, Schiffelers RM and
Hennink WE: Curcumin nanoformulations: A review of pharmaceutical
properties and preclinical studies and clinical data related to
cancer treatment. Biomaterials. 35:3365–3383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marquardt JU, Gomez-Quiroz L, Camacho
Arreguin LO, Pinna F, Lee YH, Kitade M, Dominguez MP, Castven D,
Breuhahn K, Conner EA, et al: Curcumin effectively inhibits
oncogenic NF-κB signaling and restrains stemness features in liver
cancer. J Hepatol. 63:661–669. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu A, Huang JJ, Jin XJ, Li JP, Tang YJ,
Huang XF, Cui HJ, Xu WH and Sun GB: Curcumin suppresses
invasiveness and vasculogenic mimicry of squamous cell carcinoma of
the larynx through the inhibition of JAK-2/STAT-3 signaling
pathway. Am J Cancer Res. 5:278–288. 2014.PubMed/NCBI
|
|
31
|
Shu L, Khor TO, Lee JH, Boyanapalli SS,
Huang Y, Wu TY, Saw CL, Cheung KL and Kong AN: Epigenetic CpG
demethylation of the promoter and reactivation of the expression of
Neurog1 by curcumin in prostate LNCaP cells. AAPS J. 13:606–614.
2011. View Article : Google Scholar : PubMed/NCBI
|