Introduction
Hepatocellular carcinoma (HCC) is ranked second
worldwide with regard to tumor-related death (1). In addition, chronic infection through
the hepatitis B virus (HBV) is strongly associated with death
(2). Surgical resection or
transplantation is used as a form of treatment of HCC, when the
disease is detected early. However, in most cases, patients are
diagnosed with HCC at an advanced stage. Consequently, doctors have
limited treatment options because of lack of effective therapies.
Activation of proto-oncogenes and deactivation of cancer-inhibitor
genes play a pivotal role in tumorigenesis (3).
MicroRNAs can suppress the expression of related
genes or induce their degradation at the post-transcriptional level
(4–6).
Approximately 1–5% of animal genes have been predicted to comprise
miRNAs (7). Existing miRNAs have been
identified in a number of organisms. Cell proliferation,
differentiation, and apoptosis are regulated in the
post-transcriptional regulation mechanism by various miRNAs
(8,9).
Therefore, human cancer cells developing from normal cells through
a series of disorders are caused by the aberrant regulation of
miRNAs (10).
The dysregulated expression of many microRNAs,
including miR-21 in HCC, has been reported, which were identified
in genomic analysis (11,12). The up- or downregulation of miR-21
were revealed in abundant malignancies, often overexpressed in
various types of cancer, such as multiple myeloma pathogenesis,
lung cancer (13–15). It has been confirmed that programmed
cell death 4 (PDCD4) as a tumor suppressor may be downregulated by
miR-21 (16). The tumorigenesis of
colorectal carcinoma may also be stimulated by miR-21 at the
post-transcriptional level (17). In
addition, miR-21 controlled breast cancer cell progression by
targeting the PDCD4 factor (18). The
expression of long non-coding RNA (IncRNA) growth arrest-specific 5
(GAS5) can be inhibited by miR-21. Of note, GAS5 is capable of
suppressing miR-21 expression (19).
In HCC, HBV X protein (HBx) downregulates PDCD4 expression via
miR-21 (20). The function of miR-21
was also present in pancreatic, gastric and lung cancer (21–23). The
Kruppel-like family of transcription factors (KLFs), are considered
tumor suppressors. The cell growth of breast cancer was inhibited
via KLF5 (24). In addition, the cell
growth, cell cycle regulation, and angiogenesis can be influenced
by KLF5. In human bladder cancer, Gao et al found that cell
proliferation and tumorigenesis were promoted by KLF5 (25). Downregulation of KLF5 is often
detected in prostate cancer (26).
However, whether the regulation of migration and
invasion in HCC is induced via miR-21 through KLF5 remains to be
elucidated. To analyze the function of miR-21, miR-21 upregulation
in HCC was identified in tumor tissues and cell line in this study
using RT-qPCR. The results showed that in HCC cells, miR-21
overexpression promoted metastasis and invasion. Furthermore, KLF5
as a downstream target of miR-21 was inhibited by miR-21. The
present findings demonstrate a valuable therapeutic biomarker for
HCC differentiation and treatment.
Materials and methods
Tissue samples
Between March, 2013 and May, 2016, 80 patients
selected from People's Hospital of Linyi (Linyi, China), were
included in this study. Following surgical resection, tumor tissues
obtained from the patients, were quickly frozen in liquid nitrogen
and then stored at −80°C until further use. None of the patients
participating in the study underwent chemo- or radiation
therapy.
The present study was approved by the Ethics
Committee of People's Hospital of Linyi. Informed consent was
obtained from all the patients whose tissues were used in this
study.
Cell culture
Three cell lines (Huh 7 and SK-HEP-1) and normal
human hepatocytes (LO-2) were purchased from the American Type
Culture Collection (ATCC; Manassas, VA, USA) and Sciencell (San
Diego, CA, USA), respectively. These cell lines were cultured with
DMEM medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), added with 10% FBS (Biowest, South America Origin, Paris,
France), 100 IU/ml penicillin and 100 µg/ml streptomycin, and
placed in a humidity incubator at 37°C and 5% CO2.
Plasmid construction
In our study, the psiCHECK-2 plasmid (Promega,
Madison, WI, USA) was selected for the dual luciferase reporter
assay. The partial KLF5 gene served as the gene from which the
3′-untranslated regions (UTRs) were taken and cloned for the
appropriate miR-21-binding site. Then, it was linked into the
downstream of the luciferase reporter gene of psiCHECK-2. The
plasmids containing miR-21 or LNA anti-miR-21 were constructed. In
order to knock down the KLF5 gene, KLF5 siRNA was produced.
Transfection
We purchased miR-21 mimic and inhibitor from
GenePharma Co., Ltd. (Suzhou, China) and KLF5 vector from Shanghai
Genechem Co., Ltd. (Shanghai, China). Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) and RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) were used to transfect
miR-21 mimic, miR-21 inhibitor, KLF5 vector or both KLF5 vector and
miR-21 mimic, respectively. For transfection, Huh 7 or SK-HEP-1
cells were seeded in 24- or 6-well plates and then transfected with
the indicated plasmids for 48 h using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. The cells were collected for the following
experiments after 48 h of transfection.
RNA extraction and reverse
transcription-quantitative PCR
TRIzol reagent (Takara, Dalian, China) was used to
isolate total RNA from HCC tissues and three cell lines Huh 7,
SK-HEP-1 and LO-2 cell. The SYBR PrimeScript RT-PCR kit (Takara)
was applied for reverse-transcription of RNA to detect miR-21 and
KLF5. The Roche Light Cycler 480 instrument (Roche, Basel,
Switzerland) was used to perform reverse transcription-quantitative
PCR (RT-qPCR). The snRNA U6 or GAPDH were identified as the
internal control for miRNA or KLF5, respectively. The miR-21 and
KLF5 relative expression ratio was determined using the
2−∆∆Cq algorithm (27).
PCR primers used were as follows: miR-21, forward:
5′-TAGCTTATCAGACTGATG-3′ and reverse: 5′-TGGTGTCGTGGAGTCG-3′; U6,
forward: 5′-CTCGCTTCGGCAGCACA-3′ and reverse:
5′-AACGCTTCACGAATTTGCGT-3′; KLF5 forward:,
5′-ATCGAGATGTTCGCTCGTGC-3′; and reverse,:
5′-TTTAAAGGCAGACACTGAGTCAG-3′; GAPDH forward:
5′-ATCGTCCACCGCAAATGCTTCTA-3′; and reverse:
5′-AGCCATGCCAATCTCATCTTGTT-3′. All the RT-qPCR experiments were
repeated three times.
Western blot analysis
Tissue powder (200 mg) stored at −80°C were lysed in
lysis buffer (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) for
protein extraction. The concentration of the extracted protein was
analyzed using BCA kit (Beyotime Institute of Biotechnology,
Haimen, China). A 20 µg protein sample was loaded in each lane, and
gel electrophoresis was conducted using 12% SDS-PAGE to separate
the extracted proteins, which were subsequently transferred on
nitrocellulose membranes that blocked overnight in Tris-buffered
saline including 0.1% Tween-20 and 5% skimmed milk powder at 4°C.
The membranes were then incubated with the primary antibody: rabbit
monoclonal anti-KLF5 (ab 137676; 1:1,000; Abcam, Cambridge, MA,
USA) at 4°C for 8 h. Rabbit monoclonal to GAPDH (EPR16891; 1:5000;
Abcam) was chosen as the internal reference. Then, they were
incubated with secondary antibody goat anti-rabbit IgG-HRP
(sc-2004; 1:3,000; Santa Cruz Biotechnology, Inc. Santa Cruz, CA,
USA) at room temperature for 1 h. The proteins of interest were
detected by the enhanced chemiluminescence (ECL) detection system
(Sea Biotech, Shanghai, China). Finally, the extracted protein was
quantified using the Bio-Rad GelDoc 2000 instrument (Bio-Rad,
Hercules, CA, USA).
Luciferase reporter analysis
We strived to seek antisense matches of KLF5 3′-UTR
sequences against miR-21 by means of TargetScan. In order to
perform luciferase reporter analysis, 100 ng of the negative
control or miR-21 mimic and psiCHECK-2-KLF5-3′-UTR-WT, or
psiCHECK-2-KLF5-3′-UTR-MT were co-transfected into Huh 7 cell line.
At 48 h after transfection, the cells were gathered and analyzed
via dual-luciferase reporter assay system (Promega, Madison, WI,
USA). Transfections were performed three times.
Transwell migration and invasion
assays
The 24-well Transwell chambers (Millipore
Corporation, Billerica, MA, USA) were used for the cell migration
assay according to the manufacturer's protocol. DMEM medium
containing 5% fetal bovine serum (FBS) was added to the lower
chamber as a chemoattractant. After incubation for 24 h, the cells
adhering to the upper membrane were removed with cotton wool,
whereas the cells that had migrated or invaded through the membrane
were fixed with methanol and stained with 0.1% crystal violet in 5%
CO2 at 37°C for 15 min. For the invasion assay, the
procedure was similar to the cell migration assay, except that
membranes of the upper chambers were pre-coated with 100 ml
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). The cells were
photographed under a phase-contrast microscope (Olympus, Tokyo,
Japan) and counted in five randomly chosen fields. Each experiment
was performed in triplicate.
Statistical analysis
The results are presented as the mean ± standard
deviation (SD). Statistical analysis was conducted using SPSS 17.0
(SPSS, Inc., Chicago, IL, USA). Each experiment was repeated at
least three times. The Student's t-test or Tukey-Kramer post hoc
test after one-way analysis of variance (ANOVA) in SPSS were used
to analyze the differences between the groups. Correlation between
mRNA and miRNA were estimated using the Spearman's correlation
method. P<0.05 was considered to indicate a significant
difference. We generated graphs using GraphPad Prism 6 Software
(GraphPad Software, Inc., La Jolla, CA, USA).
Results
Upregulation of miR-21 in the HCC
tissues
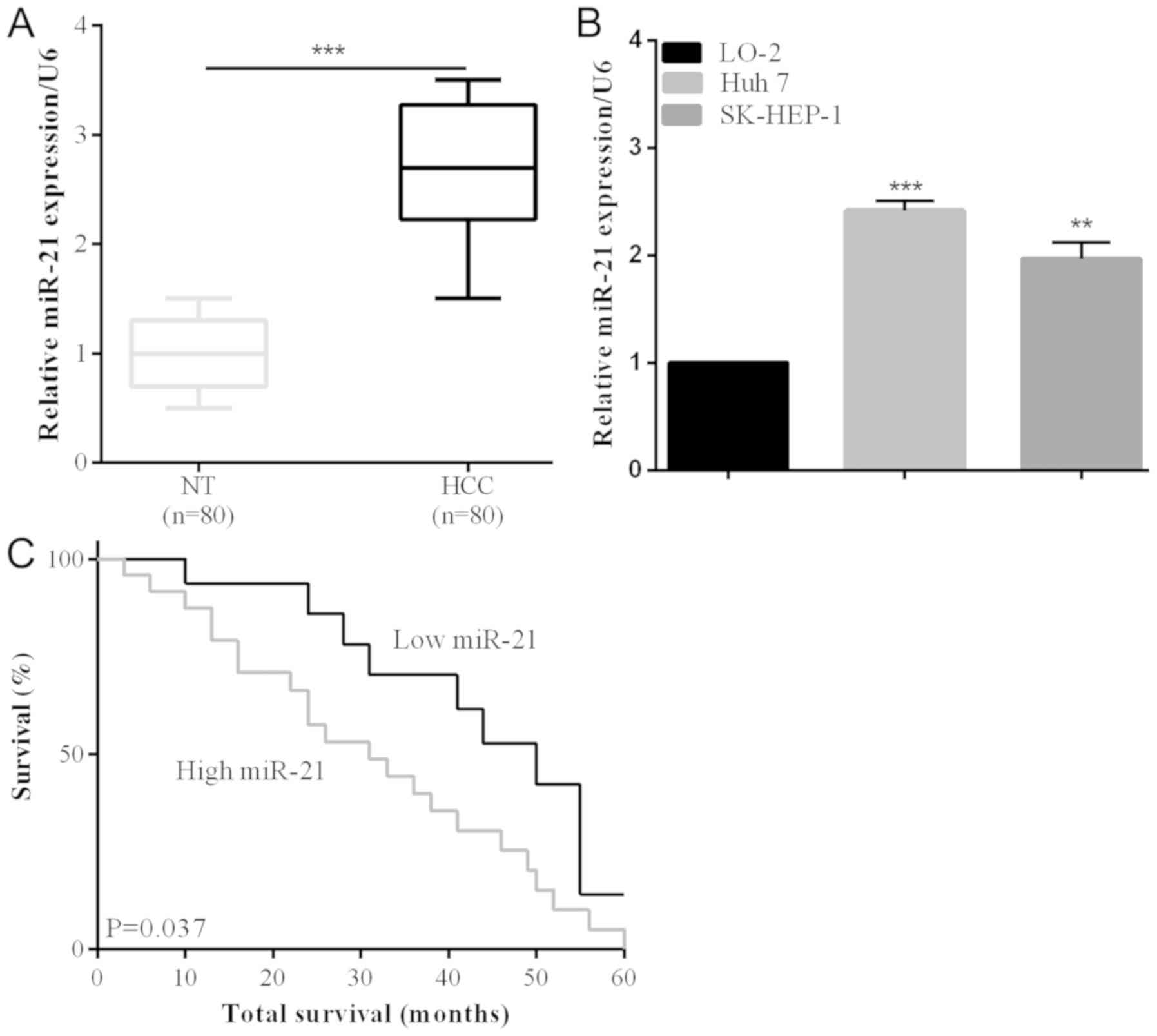
The upregulation of miR-21 was identified in HCC
cancer tissues, as in a previous study (28). Eighty collected human HCC tissues and
their adjacent non-tumorous samples were analyzed using RT-qPCR to
investigate miR-21 expression in this study. miR-21 expression was
significantly increased compared with non-tumorous in HCC tissues
(P<0.001) (Fig. 1A). According to
the RT-qPCR results, miR-121 expression was increased in HCC cell
lines (Huh 7 and SK-HEP-1) compared with LO-2 normal liver cells
(Fig. 1B). The correlation between
clinicopathological characteristics and the miR-21 expression level
was also detected. We observed that the lager tumor size was
associated with an increase of miR-21 expression (P=0.0042)
(Table I). Additionally, poor 5-year
survival rate was significantly associated with miR-21
overexpression (P=0.037) (Fig. 1C),
suggesting a relationship between the prognostic and miR-21
expression level.
 | Table I.miR-21 expression and
clinicopathological characteristics in HCC patients. |
Table I.
miR-21 expression and
clinicopathological characteristics in HCC patients.
|
|
| miR-21
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | No. | Low n (%) | High n (%) |
P-valuea |
|---|
| Age (year) |
|
|
| 0.768 |
|
<50 | 39 | 18 (46.2) | 21 (53.8) |
|
|
≥50 | 61 | 30 (49.2) | 31 (50.8) |
|
| Sex |
|
|
| 0.519 |
|
Male | 52 | 25 (48.1) | 27 (51.9) |
|
|
Female | 48 | 20 (41.7) | 28 (58.3) |
|
| Tumor size
(cm) |
|
|
| 0.0042 |
|
<5 | 31 | 15 (48.4) | 16 (51.6) |
|
| ≥5 | 69 | 14 (20.3) | 55 (79.7) |
|
| Tumor
differentiation |
|
|
| 0.732 |
| I +
II | 55 | 25 (45.5) | 30 (54.5) |
|
| III +
IV | 45 | 22 (48.9) | 23 (51.1) |
|
| Metastasis |
|
|
| 0.658 |
| No | 41 | 19 (46.3) | 22 (53.7) |
|
|
Yes | 59 | 25 (42.4) | 24 (57.6) |
|
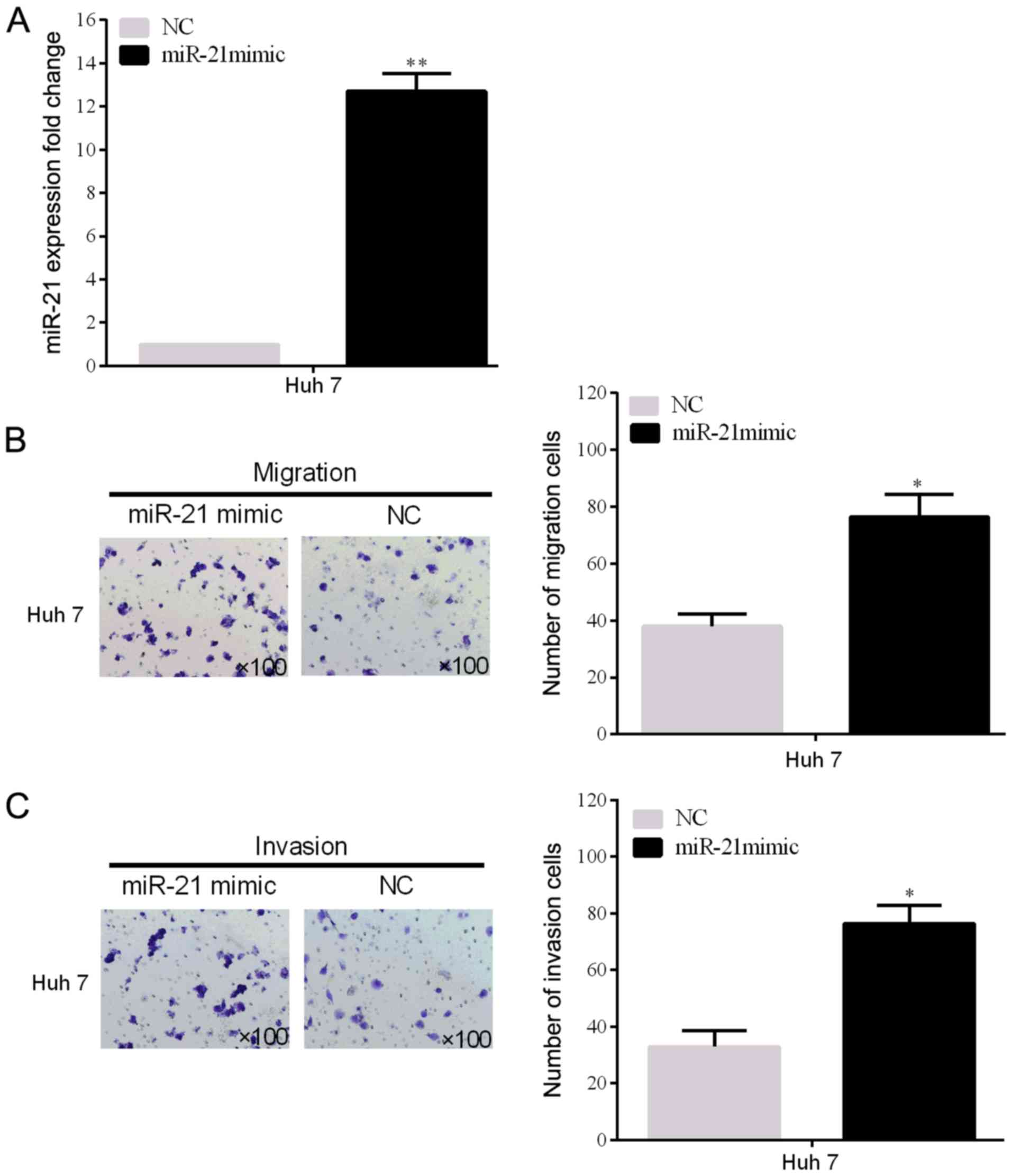
miR-21 overexpression increases the
number of invasive and migrated cells in HCC
To prove the miR-21 biological influence of cell
invasion and migration, miR-21 expression levels of the Huh 7 cell
line transfected with miR-21 mimic plasmid was detected (P<0.05)
(Fig. 2A). In addition, the invasion
and migration rates of the two HCC cell lines (SMMC-7721) were
remarkably promoted by miR-21 overexpression, which were
demonstrated by the Transwell assays (Fig. 2B and C).
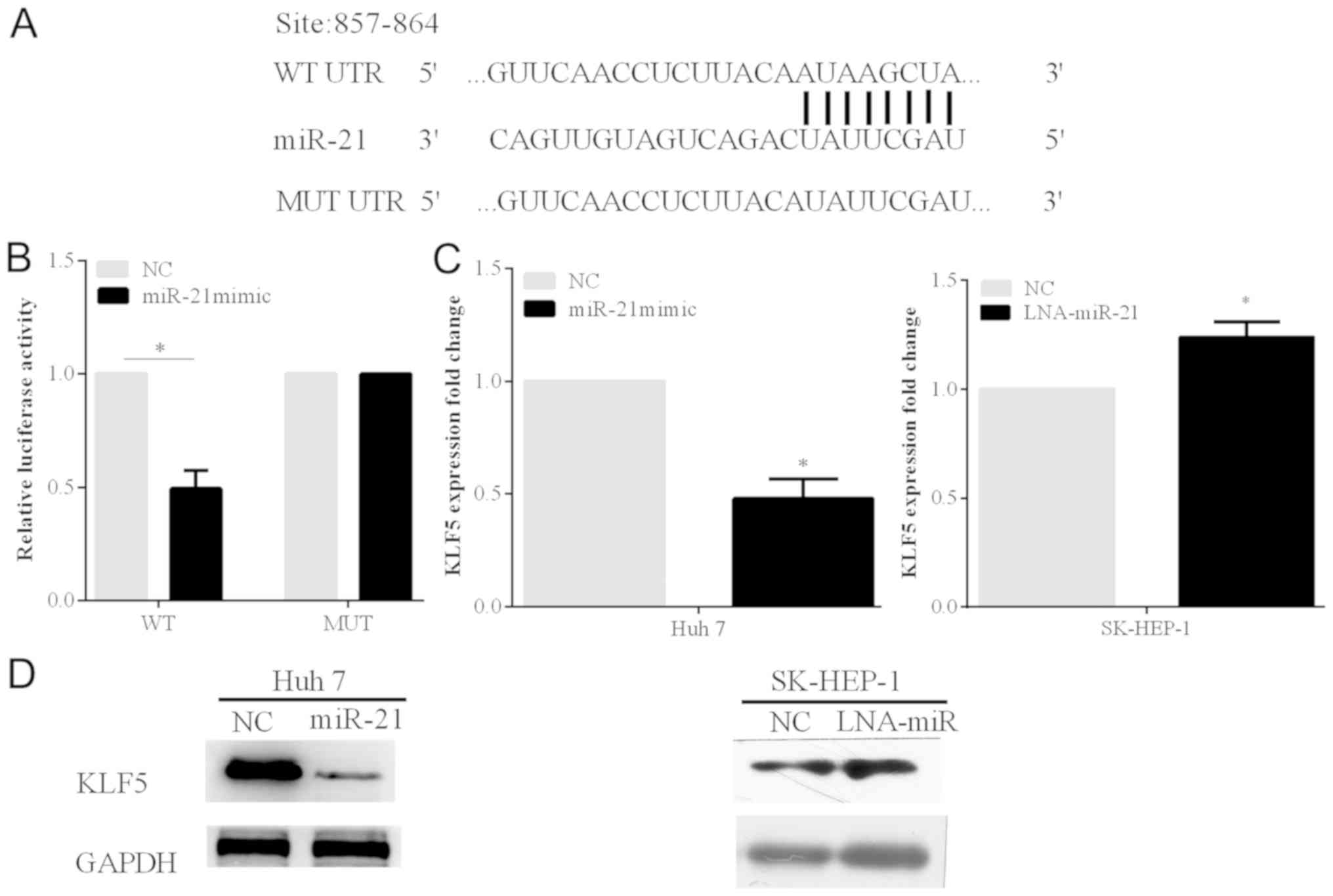
miR-21 inhibits the KLF5 expression
and binds to the 3′-UTR of the KLF5
To identify the potential downstream targets of
miR-21, bioinformatics analysis was carried out on the website
TargetScan (http://www.targetscan.org/). By the online prediction,
KLF5 was selected as the target gene of miR-21. The prediction
showed that KLF5 gene has one binding site to miR-21 at 857–864 bp
in the 3′-UTR (Fig. 3A). In order to
verify the prediction, the luciferase reporter analysis was carried
out using the Huh 7 cell line.
The relative luciferase activity of the cells
transfected with the miR-21 mimic and the psi-KLF5-3′-UTR-WT
plasmid was significantly decreased. On the other hand, when the
miR-21 mimic and psi-KLF5-3′-UTR-MUT plasmid containing mutant
sites at the 3′-UTR of KLF5 were transfected into the cell line,
the reduction extent of the luciferase activity was alleviated
(P<0.05) (Fig. 3B).
Furthermore, we found that the protein and mRNA
expression levels of KLF5 were markedly inhibited by miR-21
overexpression in Huh 7 cells. The protein and mRNA expression
levels of KLF5 were increased following with the silenced miR-21 in
SK-HEP-1 cells (P<0.05) (Fig. 3C and
D).
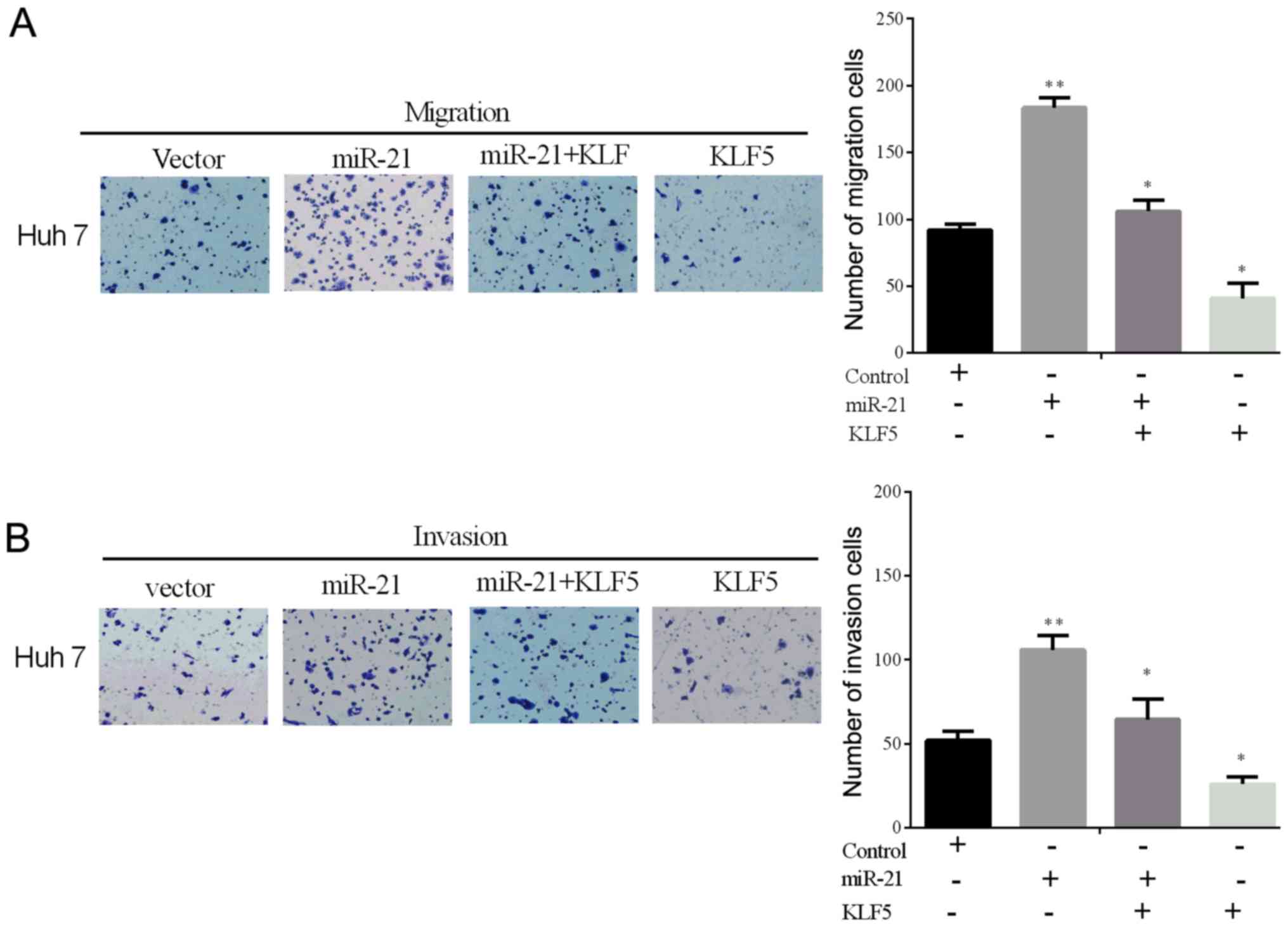
Overexpression of KLF5 alleviates the
miR-21-driven promoting effect on cell line invasion and migration
in vitro
To confirm whether KLF5 overexpression could reverse
the miR-21-driven promoting effect on the cell invasive and
migratory ability in vitro, the expressing plasmid
containing the KLF5 without the 3′-UTR was constructed. The
invasion and migration rates were tested using the Transwell assays
in the selected cell line (Huh 7) transfected with the miR-21 mimic
and the KLF5 lacking the 3′-UTR (Fig. 4A
and B). From the results, we could identify that the
overexpression of KLF5 alleviates the miR-21-driven promoting role
of cell invasion and migration in vitro.
Discussion
A number of onco-miRNAs have been previously
identified, especially miR-21 (28).
The function of miR-21 which is frequently upregulated has been
identified in numerous cancers including colorectal cancer
(29), breast carcinoma (18), pancreatic carcinoma (21), gastric carcinoma (22), lung carcinoma (30,31),
colorectal carcinoma (32) and
hepatocellular carcinoma (33). In
the present study, it was found that the miR-21 expression level
was enhanced in HCC tumor tissues compared to the corresponding
non-tumorous tissues. In addition, the invasive and migratory
ability of the hepatoma cell line in vitro could be promoted
by miR-21 re-expression. At the post-transcriptional level, we
first demonstrated that miR-21 negatively modulated KLF5 by
combining with the 3′-UTR of the tumor suppressor in this
study.
Previous findings have shown that, PDCD4 expression
was downregulated via HBx, which was partially modulated by miR-21
(20). In the present study, we did
not employ HBV and HBx. However, HBV-infected hepatic cells are
involved in the formation of HCC. Therefore, identifying the
correlation between HBx and KLF5 in future investigations is
imperative. miR-21 also has a great impact on the pathogenesis of
atrial fibrosis. In addition, miR-21 inhibition induced by the
antagomir-21 could suppress the fibrosis of atrial fibrillation
post-myocardium infarction (34).
This confirms that miR-21 has various different functions in
different cancer types.
As a member of the Kruppel-like family, KLF5 is
involved in the modulation of various physiological and
pathological processes, KLF5 has been reported to regulate
embryonic development, stress response, cell proliferation and
differentiation, carcinogenesis and cardiovascular remolding
through various regulatory mechanisms, as indicated by previous
findings (35,36). In addition, KLF5 contributes to
bladder cancer angiogenesis via the VEGFA pathway. The GC boxes and
CACCC elements of VEGFA combine with KLF5 to regulate
tumorigenesis. The silence of KLF5 can remarkably inhibit the
expression of VEGFA (25). In our
study, miR-21 regulated KLF5 by binding to its 3′-UTR.
Additionally, HCC cell invasion and migration can be promoted
through the knockdown of KLF5 by siRNA in vitro.
miR-153 reduced KLF5 expression by combining with
its 3′-UTR, as is evident from our results. In addition, the
expression of KLF5, suppressed by miR-153, was verified by the
luciferase reporter assay (37). ERβ
is a nuclear receptor for estrogen (E2). E2 biphasically regulated
KLF5 transcription through ERβ to modulate prostate tumor
progression, which showed that the KLF5 expression was associated
with the progression of prostate tumor (38). Thus, we may develop or identify
effective drugs to prevent the progression of HCC.
In summary, results of the present study show that,
the invasion and migration of HCC cells may be significantly
suppressed by miR-21 by targeting the tumor suppressor KLF5.
Accordingly, the miR-21/KLF5 axis may open up a novel way for
treating HCC, such as tumor invasion and metastasis.
Acknowledgements
Not applicable.
Funding
This research did not receive any specific grant
from funding agencies in the public, commercial, or not-for-profit
sectors.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JW and YC contributed to the conception of the
study; MX (3rd author) contributed significantly to data analysis
and manuscript preparation; XZ performed the data analysis and
wrote the manuscript; YZ and MX (last author) helped perform the
analysis with constructive discussions. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Approval for the study was obtained from the Ethics
Committee of People's Hospital of Linyi (Linyi, China). Informed
consent was obtained from all the patients whose tissues were used
in this study.
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tan A, Yeh SH, Liu CJ, Cheung C and Chen
PJ: Viral hepatocarcinogenesis: from infection to cancer. Liver
Int. 28:175–188. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rana TM: Illuminating the silence:
Understanding the structure and function of small RNAs. Nat Rev Mol
Cell Biol. 8:23–36. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pillai RS, Bhattacharyya SN and Filipowicz
W: Repression of protein synthesis by miRNAs: How many mechanisms?
Trends Cell Biol. 17:118–126. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Berezikov E, Guryev V, van de Belt J,
Wienholds E, Plasterk RH and Cuppen E: Phylogenetic shadowing and
computational identification of human microRNA genes. Cell.
120:21–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Si ML, Zhu S, Wu H, Lu Z, Wu F and Mo YY:
miR-21-mediated tumor growth. Oncogene. 26:2799–2803. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang J, Gusev Y, Aderca I, Mettler TA,
Nagorney DM, Brackett DJ, Roberts LR and Schmittgen TD: Association
of MicroRNA expression in hepatocellular carcinomas with hepatitis
infection, cirrhosis, and patient survival. Clin Cancer Res.
14:419–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang WY, Zhang HF, Wang L, Ma YP, Gao F,
Zhang SJ and Wang LC: miR-21 expression predicts prognosis in
hepatocellular carcinoma. Clin Res Hepatol Gastroenterol.
38:715–719. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pichiorri F, Suh SS, Ladetto M, Kuehl M,
Palumbo T, Drandi D, Taccioli C, Zanesi N, Alder H, Hagan JP, et
al: MicroRNAs regulate critical genes associated with multiple
myeloma pathogenesis. Proc Natl Acad Sci USA. 105:12885–12890.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xia H, Zhang W, Zhang B, Zhao Y, Zhao Y,
Li S and Liu Y: miR-21 modulates the effect of EZH2 on the
biological behavior of human lung cancer stem cells in vitro.
Oncotarget. 8:85442–85451. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu Q, Wang Z, Hu Y, Li J, Li X, Zhou L
and Huang Y: miR-21 promotes migration and invasion by the
miR-21-PDCD4-AP-1 feedback loop in human hepatocellular carcinoma.
Oncol Rep. 27:1660–1668. 2012.PubMed/NCBI
|
|
17
|
Asangani IA, Rasheed SA, Nikolova DA,
Leupold JH, Colburn NH, Post S and Allgayer H: MicroRNA-21 (miR-21)
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in colorectal
cancer. Oncogene. 27:2128–2136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Frankel LB, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Zhu Z, Watabe K, Zhang X, Bai C,
Xu M, Wu F and Mo YY: Negative regulation of lncRNA GAS5 by miR-21.
Cell Death Differ. 20:1558–1568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qiu X, Dong S, Qiao F, Lu S, Song Y, Lao
Y, Li Y, Zeng T, Hu J, Zhang L, et al: HBx-mediated miR-21
upregulation represses tumor-suppressor function of PDCD4 in
hepatocellular carcinoma. Oncogene. 32:3296–3305. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sicard F, Gayral M, Lulka H, Buscail L and
Cordelier P: Targeting miR-21 for the therapy of pancreatic cancer.
Mol Ther. 21:986–994. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang SM, Huang C, Li XF, Yu MZ, He Y and
Li J: miR-21 confers cisplatin resistance in gastric cancer cells
by regulating PTEN. Toxicology. 306:162–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu XG, Zhu WY, Huang YY, Ma LN, Zhou SQ,
Wang YK, Zeng F, Zhou JH and Zhang YK: High expression of serum
miR-21 and tumor miR-200c associated with poor prognosis in
patients with lung cancer. Med Oncol. 29:618–626. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen C, Bhalala HV, Qiao H and Dong JT: A
possible tumor suppressor role of the KLF5 transcription factor in
human breast cancer. Oncogene. 21:6567–6572. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gao Y, Wu K, Chen Y, Zhou J, Du C, Shi Q,
Xu S, Jia J, Tang X, Li F, et al: Beyond proliferation: KLF5
promotes angiogenesis of bladder cancer through directly regulating
VEGFA transcription. Oncotarget. 6:43791–43805. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen C, Bhalala HV, Vessella RL and Dong
JT: KLF5 is frequently deleted and down-regulated but rarely
mutated in prostate cancer. Prostate. 55:81–88. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wagenaar TR, Zabludoff S, Ahn SM, Allerson
C, Arlt H, Baffa R, Cao H, Davis S, Garcia-Echeverria C, Gaur R, et
al: Anti-miR-21 suppresses hepatocellular carcinoma growth via
broad transcriptional network deregulation. Mol Cancer Res.
13:1009–1021. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sazanov AA, Kiselyova EV, Zakharenko AA,
Romanov MN and Zaraysky MI: Plasma and saliva miR-21 expression in
colorectal cancer patients. J Appl Genet. 58:231–237. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lasithiotaki I, Tsitoura E, Koutsopoulos
A, Lagoudaki E, Koutoulaki C, Pitsidianakis G, Spandidos DA,
Siafakas NM, Sourvinos G and Antoniou KM: Aberrant expression of
miR-21, miR-376c and miR-145 and their target host genes in Merkel
cell polyomavirus-positive non-small cell lung cancer. Oncotarget.
8:112371–112383. 2016.PubMed/NCBI
|
|
31
|
Charkiewicz R, Pilz L, Sulewska A,
Kozlowski M, Niklinska W, Moniuszko M, Reszec J, Manegold C and
Niklinski J: Validation for histology-driven diagnosis in non-small
cell lung cancer using hsa-miR-205 and hsa-miR-21 expression by two
different normalization strategies. Int J Cancer. 138:689–697.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu Y, Nangia-Makker P, Farhana L, G
Rajendra S, Levi E and Majumdar AP: miR-21 and miR-145 cooperation
in regulation of colon cancer stem cells. Mol Cancer. 14:982015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li ZB, Li ZZ, Li L, Chu HT and Jia M:
MiR-21 and miR-183 can simultaneously target SOCS6 and modulate
growth and invasion of hepatocellular carcinoma (HCC) cells. Eur
Rev Med Pharmacol Sci. 19:3208–3217. 2015.PubMed/NCBI
|
|
34
|
Adam O, Löhfelm B, Thum T, Gupta SK, Puhl
SL, Schäfers HJ, Böhm M and Laufs U: Role of miR-21 in the
pathogenesis of atrial fibrosis. Basic Res Cardiol. 107:2782012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tetreault MP, Yang Y and Katz JP:
Krüppel-like factors in cancer. Nat Rev Cancer. 13:701–713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dong JT and Chen C: Essential role of KLF5
transcription factor in cell proliferation and differentiation and
its implications for human diseases. Cell Mol Life Sci.
66:2691–2706. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu R, Shi P, Nie Z, Liang H, Zhou Z, Chen
W, Chen H, Dong C, Yang R, Liu S, et al: Mifepristone suppresses
basal triple-negative breast cancer stem cells by down-regulating
KLF5 expression. Theranostics. 6:533–544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nakajima Y, Osakabe A, Waku T, Suzuki T,
Akaogi K, Fujimura T, Homma Y, Inoue S and Yanagisawa J: Estrogen
exhibits a biphasic effect on prostate tumor growth through the
estrogen receptor β-KLF5 pathway. Mol Cell Biol. 36:144–156.
2015.PubMed/NCBI
|