Introduction
Colon cancer was the third leading cause of
malignancy-associated deaths in America in 2014, and represents a
major therapeutic challenge (1).
Despite advances in therapy, including surgery and chemotherapy,
tumor recurrence and metastasis cannot yet be effectively prevented
(2). Accumulating evidence suggests
the presence of a small subpopulation of cells, known as cancer
stem cells (CSCs), in colon cancer that exhibit stem-like features,
and promote tumor formation, metastasis and resistance to therapy
(3–6). Therefore, therapies targeting colon
CSCs may be a promising approach to eradicating colon cancer
(7–9).
Higher expression of c-Myc has been found in colon
cancer cells, relative to in normal mucosa, suggesting that c-Myc
is involved in tumor development and progression (10). Recently, c-Myc has been recognized as
an important regulator of stem cell biology (11); thus, it may serve as a link between
malignancy and ‘stemness’. Previous studies have shown that c-Myc
has a key role in regulating the self-renewal and survival of
glioma CSCs, and targeting c-Myc pathways may significantly improve
brain tumor therapies (12). It is
also reported that c-Myc can contribute to the multidrug resistance
profile by dysregulating the transcription of specific ABC
transporter genes in cancer cells (13,14).
However, c-Myc gene regulation mechanisms underlying the
maintenance of self-renewal and drug-resistant properties in colon
CSCs are still unclear.
In a previous study (6), a subpopulation of colon CSCs expressing
a CD133 surface phenotype were isolated from the human HT-29
colonic adenocarcinoma cell line using flow cytometry cell sorting
(FACS). CD133+ cells possess a greater tumor
sphere-forming efficiency in vitro and higher tumorigenic
potential in vivo. Furthermore, CD133+ cells are
endowed with stem cell-like properties, including the expression of
‘stemness’ genes, such as Wnt2, BMI1, Oct3/4, Notch1 and c-Myc, and
the maintenance of self-renewal and differentiation capacities. The
miRNA expression signature of colon CSCs was revealed in our
previous study (6).
In the present study, it was further demonstrated
that c-Myc was consistently overexpressed in colon CSCs.
Additionally, it was shown that downregulation of c-Myc suppressed
the self-renewal of colon CSCs and the growth of xenografts.
Furthermore, it was suggested that the depletion of c-Myc may
enhance the chemosensitivity of colon CSCs through the
downregulation of ABCG2 and ABCB5 expression. The present study
demonstrated that the expression of c-Myc has a crucial role in
maintaining the self-renewal and chemoresistance of colon CSCs.
Materials and methods
Cell culture and colon sphere
formation
The human HT-29 colonic adenocarcinoma cell line was
obtained from the American Type Culture Collection (ATCC; Manassas,
VA, USA) and was maintained in McCoy's 5A medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C
and 5% CO2, and the medium was changed every 3 days.
Cells were passaged at 80% confluence and seeded at 20% confluence
to keep them at optimal proliferating conditions.
For colon sphere formation, single-cell suspensions
were cultured in a DMEM/F-12 basal serum-free medium (Gibco; Thermo
Fisher Scientific, Inc.), containing 2 mM L-glutamine, 1 mg/ml
NaHCO3, 4 µg/ml heparin, 100 µg/ml transferrin, 25 µg/ml
insulin, 30 nM sodium selenite anhydrous and 20 nM progesterone
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), and supplemented
with 20 ng/ml pro-epidermal growth factor (EGF; R&D Systems
China Co., Ltd., Shanghai, China) and 10 ng/ml fibroblast growth
factor 2 (FGF-2; R&D Systems China Co.).
In chemoresistance experiments, colon CSCs and
adherent HT-29 cells were exposed to 50 µM 5-FU (Sigma-Aldrich;
Merck KGaA) or 1.25 µM oxaliplatin (Sigma-Aldrich; Merck KGaA) or
FOLFOX (50 µM 5-FU plus 1.25 µM oxaliplatin) for 72 h, and then
observed by inverted phase-contrast microscope.
FACS
For the isolation of CD133+ and
CD133− populations, single-cell suspensions were
incubated with phycoerythrin (PE)-conjugated anti-human CD133/1
(1:1,000; 130–108, AC133 clone; Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany) and FcR blocking reagent (Miltenyi Biotec GmbH)
in staining solution containing 1% BSA and 2 mM EDTA for 10 min at
4°C. Isotype-matched mouse immunoglobulin served as the control.
Samples were analyzed and sorted with a FACSAria flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA). For the positive and
negative populations, only the top 10% most brightly stained cells
or the bottom 10% most dimly stained cells were selected,
respectively. Following cytofluorimetric sorting, viability was
assessed using trypan blue exclusion; cell purity was controlled
via flow cytometry with an antibody against CD133/2-phycoerythrin
(1:1,000; 130-090-853, 293C3 clone; Miltenyi Biotec GmbH) in
staining solution containing 1% BSA and 2 mM EDTA for 10 min at
4°C.
Small interfering RNA (siRNA)
transfection
siRNA specific to c-Myc (sc-29227; sense,
5′-CAGAAAUGUCCUGAGCAAUUU-3′ and antisense,
5′-AUUGCUCAGGACAUUUCUGUU-3′) and a scrambled siRNA (sc-37007;
sense, 5′-GACUUCAUAAGGCGCAUGCUU-3′ and antisense,
5′-GCAUGCGCCUUAUGAAGUCUU-3′) were obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Sorted CD133+
HT-29 cells (5×105) were seeded into 6-well plates in
SFM. After 24 h, siRNA was transfected into CD133+ cells
at a final concentration of 100 nM using Lipofectamine™ RNAiMAX
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. The cells were collected for a
series of experiments at 48 h after transfection.
In vivo tumorigenicity
Transfected cells (5×105) were
resuspended in 50 µl PBS, and cell aliquots were diluted 1:1 with
growth factor-reduced Matrigel matrix (BD Biosciences) before
injection. A total of 10 BALB/c nude female mice (6 weeks old),
weighing 18–22 g, were purchased from the Shanghai Laboratory
Animal Center of the Chinese Academy of Sciences (Shanghai, China).
The mice were maintained under a 12 h light/dark cycle at an
ambient temperature of 24±1°C in a humidity-controlled environment
in a specific pathogen-free animal facility at the Center for
Animal Experimentation, Medical Institute of Shanghai Jiao Tong
University (Shanghai, China). Sterilized water and γ-irradiated
diet were provided to animals ad libitum. Mice were
subcutaneously injected with post-transfected cells, and the mice
were observed every 2 weeks. The duration of the experiments was 8
weeks following transplantation, after which all mice were
anesthetized with 1% pentobarbital sodium (40 mg/kg) and euthanized
by cervical dislocation. Every mouse had one tumor, which was
excised and measured. Each group consisted of 5 mice.
Western blotting
Protein was extracted with protein extraction kit
(Minute™ Protein Extraction Kit; SD-001; Invent Biotechnologies,
Inc., Plymouth, MN, USA), and the protein concentration was
detected using the bicinchoninic acid (BCA) Protein Assay kit
(Beyotime Institute of Biotechnology, Haimen, China). Subsequently,
20 µg protein was separated via 10% SDS-PAGE, transferred onto a
polyvinylidene fluoride membrane, and blocked in 5% skimmed milk at
20°C for 1 h. The following primary antibodies were added to
membranes at a dilution of 1:1,000 at 4°C for 24 h: Bmi1 (sc-13519;
Santa Cruz Biotechnology, Inc.), ABCG2 (sc-13519; Santa Cruz
Biotechnology, Inc.), ABCB5 (sc-517565; Santa Cruz Biotechnology,
Inc.), Sox2 (sc-365964; Santa Cruz Biotechnology, Inc.), Wnt2
(sc-5208; Santa Cruz Biotechnology, Inc.), Oct3/4 (sc-5279; Santa
Cruz Biotechnology, Inc.), CD133 (130-108-062, AC133 clone;
Miltenyi Biotec GmbH), c-Myc (ab32072; Abcam, Cambridge, MA, USA),
β-actin (ab1801; Abcam) and GAPDH (ab8245; Abcam). The membrane was
then incubated with a horseradish peroxidase-labeled secondary
antibody (1:2,000; A0216; Beyotime Institute of Biotechnology) at
room temperature for 1 h. The membrane was visualized using an
enhanced chemiluminescence detection system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA), and analyzed using a gel analyzer
(Bio-Rad Laboratories, Inc.).
Chemo-sensitivity assays
Chemosensitivity assays were conducted using a Cell
Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan).
Sorted CD133+ HT-29 cells were seeded into 96-well
plates at a density of 4×103 cells/well in 100 µl SFM
overnight, and then transfected with 100 µM of an indicated siRNA.
For the chemosensitivity assay, post-transfected (48 h) cells were
exposed to 5-FU (50 µM), oxaliplatin (1.25 µM) and FOLFOX (50 µM
5-FU plus 1.25 µM oxaliplatin). After incubation for 72 h, a CCK-8
assay was performed, and the survival rate of cells was calculated
as follows: OD treatment/OD control × 100. Experiments were
performed in triplicate.
Migration and invasion assays
Migration and invasion assays were performed in
24-well Transwell chambers with 8-mm pore polycarbonate filter
inserts (Corning Inc., Corning, NY, USA). A total of
5×104 post-transfected cells were seeded on uncoated or
Matrigel-coated inserts in 500 µl serum-free medium for migration
or invasion assays, respectively. The lower chambers were filled
with 500 µl 10% FBS-supplemented DMEM/F12 medium. After 48 h, cells
on the upper side of the filter were removed, and the cells on the
lower surface of the insert were fixed with 4% paraformaldehyde
(500 µl) at room temperature for 20 min, washed twice with PBS (500
µl) and stained with crystal violet (400 µl) at 37°C for 20 min.
The number of stained cells was counted in 3 randomly selected
fields of view under a light microscope using a ×10 objective
(Nikon Corporation, Tokyo, Japan). Assays were performed in
triplicate.
Statistical analysis
SPSS 19.00 software (IBM Corp., Armonk, NY, USA) was
used to analyze the data in the present study. Data are presented
as the mean ± standard deviation and differences between 2 groups
were compared using Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
CD133+ cells exhibit
stem-like features in vitro
It was previously proven that CD133 is a powerful
marker for detecting colon CSC subpopulations in the human HT-29
colonic adenocarcinoma cell line (6). In the present study, CD133+
and CD133− cells were purified from the HT-29 cell line
using FACS. To explore the molecular features of colon CSCs, the
expression of genes involved in stem cell-associated pathways was
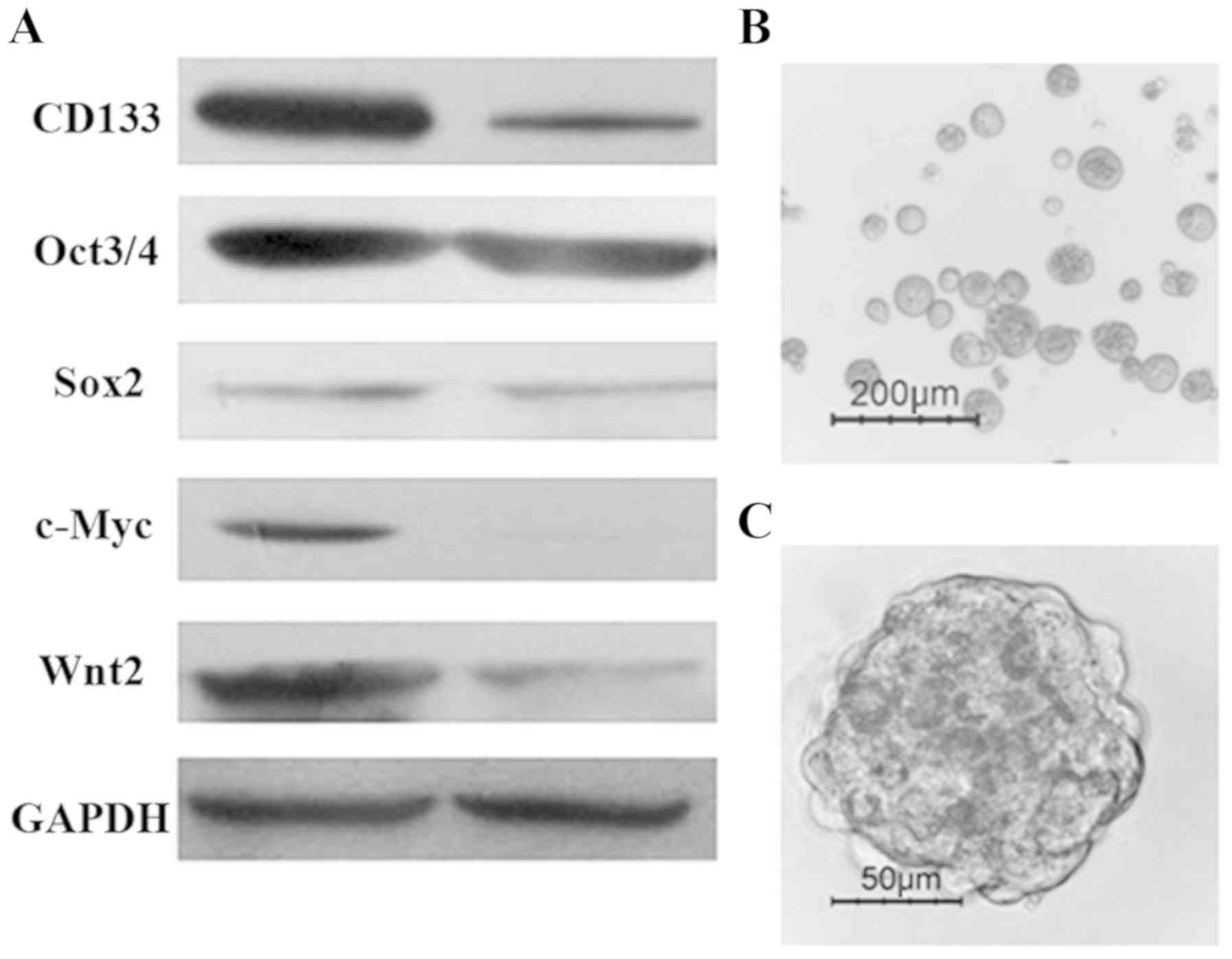
detected by western blotting. As shown in Fig. 1A, the expression of ‘stemness’ genes,
such as c-Myc, Sox2, Oct4, CD133 and Wnt2, was higher in
CD133+ cells compared with that in CD133−
cells. As has been reported previously (6), isolated CD133+ cells were
able to generate tumor spheres when cultured for 10–14 days in
serum-free stem cell medium supplemented with 20 ng/ml EGF and 10
ng/ml FGF-2. Floating sphere-like cellular aggregates were formed,
as shown in Fig. 1B and C.
Additionally, colon cancer spheres had a larger volume and more
compact sphere structure following the third cell-culture passage
(data not shown). The results demonstrated that CD133+
HT-29 cells exhibit stem-like properties, including the expression
of ‘stemness’ genes and self-renewal capacity.
Depletion of c-Myc attenuates tumor
sphere formation among colon CSCs
Tumor sphere generation in vitro is
indicative of self-renewal potential (5,9). To
investigate whether c-Myc regulates the self-renewal capacity of
colon CSCs, c-Myc expression was downregulated in CD133+
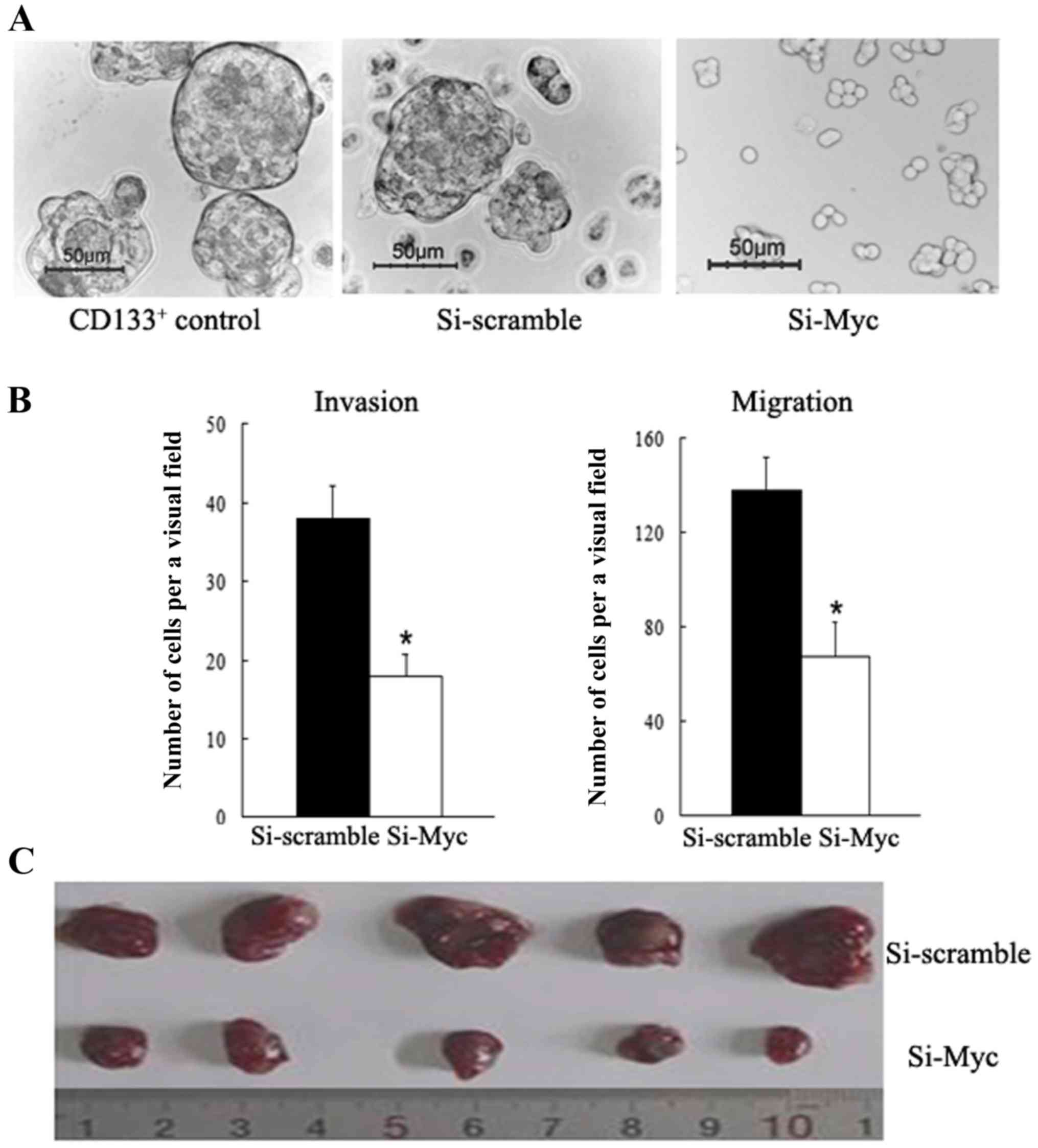
colon CSCs using siRNA, and the results showed that c-Myc-siRNA
cells formed smaller and fewer tumor spheres than the
scramble-siRNA control and CD133+ counterparts when
cultured in serum-free stem cell medium (Fig. 2A). On the other hand, the c-Myc-siRNA
group demonstrated lower Bmi1 expression levels. These data reveal
that c-Myc serves an important role in regulating the self-renewal
ability of colon CSCs cells, partly through regulating Bmi1.
Knockdown of c-Myc inhibits the
invasion and migration potential of colon CSCs
Invasive and migratory abilities are critical
features of CSCs; thus, it was examined whether c-Myc affected
these features in colon cancer. Here, in vitro assays were
conducted to evaluate migratory and invasive capacities. A
Transwell assay found that c-Myc-siRNA cells display a significant
decrease in cell motility relative to their scramble-siRNA
counterparts (P<0.01, Fig. 2B).
Additionally, significantly fewer c-Myc-siRNA cells were able to
invade Matrigel-coated inserts in the Transwell migration chambers
than with the NC counterparts (P<0.01, Fig. 2B). These results indicate that
knockdown of c-Myc inhibits the invasion and migration potential of
colon CSCs, promoting a functional phenotype associated with tumor
aggressiveness.
c-Myc-siRNA suppresses the
tumorigenicity of colon CSCs in vivo
To assess the function of c-Myc with respect to
tumorigenicity in vivo, tumor development experiments were
carried out. Nude mice were subcutaneously injected with
c-Myc-siRNA transfected CD133+ cells or scramble-siRNA
counterparts. As shown in Fig. 2C,
mice in the c-Myc-siRNA group developed much smaller subcutaneous
tumors than those in the scramble-siRNA group. The tumor diameters
of the c-Myc-siRNA group and the scramble-siRNA group were
1.07±0.18 and 0.43±0.13 cm, respectively. In vivo experiment
results illustrated that c-Myc-siRNA attenuates the tumorigenicity
of CD133+ colon CSCs.
Depletion of c-Myc enhances the
chemosensitivity in colon CSCs through the downregulation of ABCG2
and ABCB5 expression
Previous studies have reported that CSCs are widely
resistant to chemotherapeutic drugs (7,8). Herein,
colon CSCs and adherent cells were exposed to 5-FU (50 µM) or
oxaliplatin (1.25 µM) or FOLFOX (50 µM 5-FU plus 1.25 µM
oxaliplatin) for 72 h; as expected, the chemotherapy of HT-29
adherent cells resulted in a significant increase in cell death and
disintegration compared with colon CSCs, as observed via inverted
phase-contrast microscopy (Fig. 3A).
Furthermore, to evaluate the effect of c-Myc on the drug resistance
of colon CSCs, a chemosensitivity assay was conducted. Transfected
cells were treated with the same chemotherapy strategy as
aforementioned. After incubation for further 72 h, the CCK-8 assay
results demonstrated that the survival rates of
c-Myc-siRNA-transfected cells were significantly reduced compared
with those of the scramble-siRNA group (Fig. 3B). High expression of the ATP-binding
cassette and multidrug resistance protein is essential for CSC
chemoresistance (15–18). In the present study, a strong
decrease was found in ABCG2 and ABCB5 expression upon c-Myc
silencing (Fig. 3C). The results
show that c-Myc silencing enhances the chemosensitivity of colon
CSCs through the downregulation of ABCG2 and ABCB5 expression, thus
representing a valid approach for sensitizing colon CSCs to
conventional treatment.
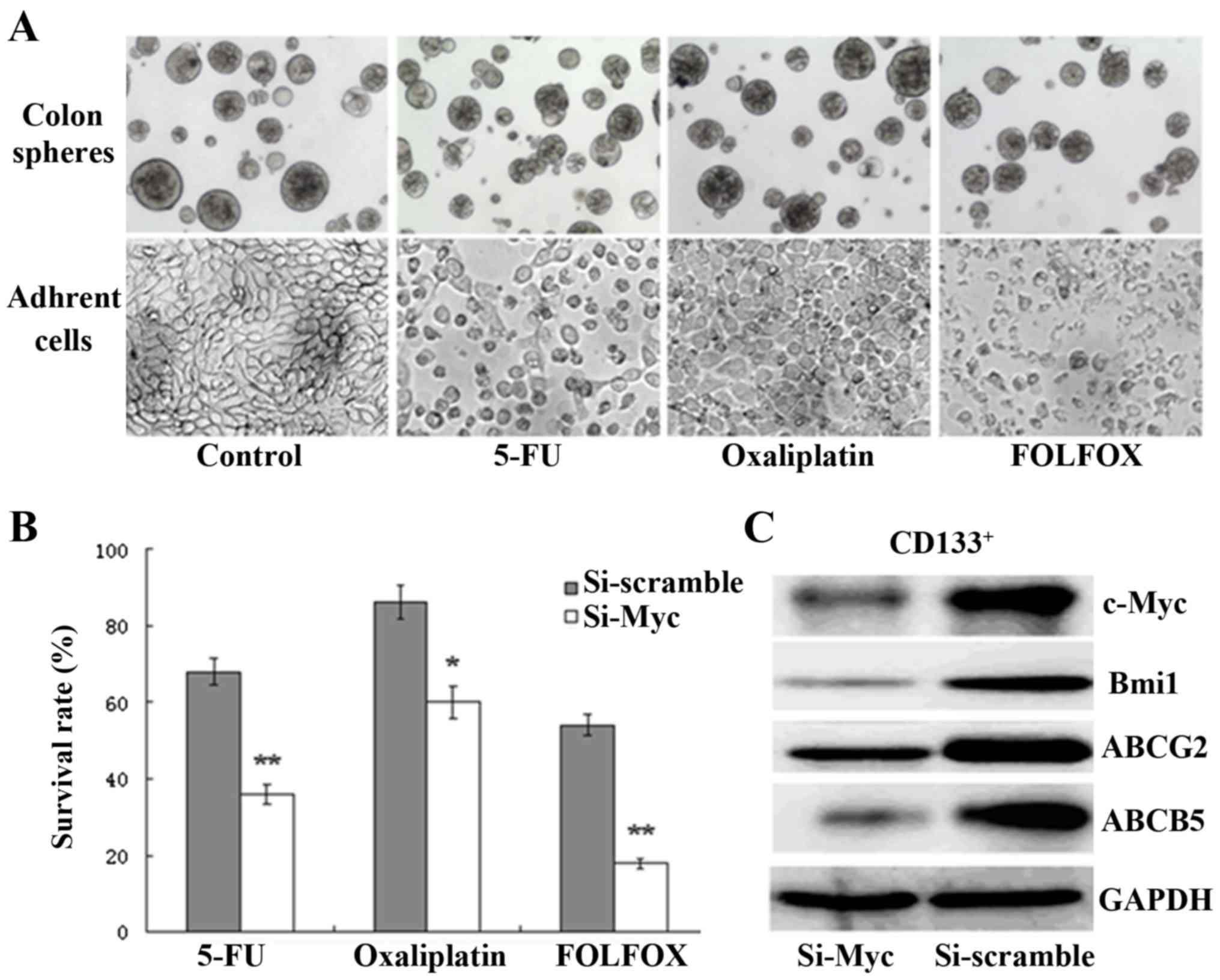 | Figure 3.Depletion of c-Myc enhances the
chemosensitivity of colon CSCs through the regulation of ABCG2 and
ABCB5 expression. (A) Chemotherapy treatment of HT-29 adherent
cells resulted in a significant increase in cell death and
disintegration, compared with colon CSCs, as observed by inverted
phase contrast microscopy (bars, 50 µm). (B) The survival rates of
c-Myc-siRNA-transfected CD133+ cells were significantly
reduced compared with the scramble-siRNA group following treatment
with 5-FU, oxaliplatin or FOLFOX (*P<0.05, **P<0.01). (C)
c-Myc, Bmi1, ABCG-2 and ABCB5 were all downregulated in
CD133+ colon CSCs, as determined by western blotting,
following treatment with c-Myc siRNA. siRNA, small interfering RNA;
CSC, cancer stem cell; FOLFOX, 5-FU plus oxaliplatin. |
Discussion
There is accumulating evidence supporting the fact
that tumors contain a small subpopulation of CSCs, which have a
self-renewing capacity and are responsible for tumor maintenance
and metastasis. In colon cancer, CD133 is regarded as a specific
marker for the isolation and identification of CSCs in primary
colon cancer, and in colon cancer cell lines (3–9). In the
present study, we purified CD133+ colon CSCs from HT-29
cell line by FACS. CD133+ cells have high expression of
‘stemness’ genes, including CD133, Sox2, Oct4, c-Myc and Wnt2; many
of these molecular markers have been previously reported to be
involved in the maintenance of stemness in human CSCs (19). Additionally, it was found that
isolated CD133+ cells were able to generate tumor
spheres when cultivated in serum-free stem cell medium supplemented
with 20 ng/ml EGF and 10 ng/ml FGF-2, which sustained the cells in
an undifferentiated state as sphere-like cellular aggregates. In
addition, it was determined that the secondary passage tumor
spheres have a larger volume and a more compact sphere structure
(data not shown), suggesting that they retained capacity for
self-renewal. The results of the present study demonstrated that
CD133+ cells are endowed with stem cell-like
properties.
c-Myc has been proven as one of 4 major factors that
render the reprogramming capability of adult cells into
germline-competent-induced pluripotent stem cells, suggesting that
c-Myc is an important regulator of stem cell biology (11). Previous studies showed that c-Myc
expression may be implicated in tumorigenesis via activating its
downstream target genes (10). These
results suggested that c-Myc may serve as a link connecting
malignancy and ‘stemness’. Herein, it was also demonstrated that
c-Myc expression was upregulated in CD133+ colon CSCs
(Fig. 1A). c-Myc expression was
downregulated in CD133+ colon CSCs by siRNA and examined
to discern whether it would affect the cells. First, it was
observed that treatment with c-Myc siRNA can block sphere formation
(Fig. 2A) inhibit the invasion and
migration potential of CD133+ colon CSCs in vitro
(Fig. 2B). In addition, tumor growth
in vivo was suppressed by c-Myc siRNA (Fig. 2C), and it was found that the Bmi1,
which controls many diverse biological cancer processes such as
differentiation, proliferation, migration, and tumorigenesis
(20), was downregulated in the
c-Myc siRNA group (Fig. 3C). These
data show that c-Myc is essential for survival, and that colon CSC
self-renewal is consistent with the findings in brain CSCs
(12).
Curative treatment failure in patients with cancer
often occurs as a result of intrinsic or acquired resistance of
tumors to chemotherapeutic agents (7). 5-FU, or 5-FU plus oxaliplatin (FOLFOX),
which remains the backbone of colorectal cancer chemotherapeutics,
still shows limited success (2). As
previously reported, CSCs show resistance to conventional
therapies, which may explain why it is difficult to completely
eradicate cancer and why recurrence is an ever-present threat
(7–9). Consistent with these hypotheses, colon
cell spheres were significantly resistant to 5-FU and oxaliplatin
compared with HT-29 adherent cells; even treatment combined with
FOLFOX failed to cause death among treated colon cell spheres
(Fig. 3A). Therapeutic strategies
that specifically target colon CSCs are likely to be effective in
eradicating tumors (21–24). To overcome the resistance to
chemotherapy of colon CSCs, treatment with c-Myc siRNA was used,
and the results showed that c-Myc knockdown can significantly
improve the anti-cancer effects in single or combination-treated
CD133+ colon CSCs in vitro (Fig. 3B).
Understanding how chemoresistance develops, and
eventually how it can be prevented, is crucial to fighting cancer
effectively (23,24). Recent studies have revealed that the
ABC transporter family are involved in multidrug resistance
(15–18). Xie et al (15) observed that ‘side population’ CSCs in
colon cancer expressed high levels of ABCG2 and had a greater
capacity to expel cytotoxic drugs. Furthermore, Porro et al
(14) showed that c-Myc may
contribute to the multidrug resistance profile and malignant
progression of myeloid tumors by dysregulating the transcription of
specific ABC transporter genes. Similarly, Kugimiya et al
(18) reported that c-Myc confers
resistance to 5-FU through regulating ABCB5 expression in human
colon cancer cells. Accordingly, in the present study, it was found
that CD133+ colon CSCs were highly co-expressed with
ABCG2 and ABCB5 (data not shown). Furthermore, it was observed that
there was a significant downregulation of ABCG2 and ABCB5
expression in CD133+ colon CSCs after c-Myc siRNA
treatment was administered (Fig.
3C), thus, enhancing the chemosensitivity of colon CSCs
(Fig. 3B). These findings indicate
that c-Myc silencing sensitizes colon CSCs to chemotherapy-induced
cytotoxicity, at least in part via the downregulation of ABCG2 and
ABCB5, although the molecular mechanisms remain to be fully
clarified; our group is currently planning further study into
this.
In conclusion, the data suggest that c-Myc knockdown
suppresses the self-renewal, tumorigenicity, invasion and drug
resistance of colon CSCs. c-Myc is, therefore, indispensable for
the maintenance of colon CSCs, and targeting c-Myc may be an
effective therapeutic strategy for eliminating colon cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (no. 81372209), and
the Project of Science and Technology of Ningbo City (nos.
2014A610223 and 2017A610153).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HLZ and PW performed all experiments in the present
study. LZ and SDZ analyzed the data, and HLZ and MZL designed all
experiments in the present study.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Lihuili Hospital of Ningbo Medical Center (Ningbo,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thomassen I, van Gestel YR, Lemmens VE and
de Hingh IH: Incidence, prognosis, and treatment options for
patients with synchronous peritoneal carcinomatosis and liver
metastases from colorectal origin. Dis Colon Rectum. 56:1373–1380.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ieta K, Tanaka F, Haraguchi N, Kita Y,
Sakashita H, Mimori K, Matsumoto T, Inoue H, Kuwano H and Mori M:
Biological and genetic characteristics of tumor-initiating cells in
colon cancer. Ann Surg Oncol. 15:638–648. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang H, Li W, Nan F, Ren F, Wang H, Xu Y
and Zhang F: MicroRNA expression profile of colon cancer stem-like
cells in HT-29 adenocarcinoma cell line. Biochem Biophys Res
Commun. 404:273–278. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Anderson EC, Hessman C, Levin TG, Monroe
MM and Wong MH: The role of colorectal cancer stem cells in
metastatic disease and therapeutic response. Cancers. 3:319–339.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang EH and Wicha MS: Colon cancer stem
cells: Implications for prevention and therapy. Trends Mol Med.
14:503–509. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fang DD, Kim YJ, Lee CN, Aggarwal S,
McKinnon K, Mesmer D, Norton J, Birse CE, He T, Ruben SM, et al:
Expansion of CD133+ colon cancer cultures retaining stem
cell properties to enable cancer stem cell target discovery. Br J
Cancer. 102:1265–1275. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meyer N and Penn LZ: Reflecting on 25
years with MYC. Nat Rev Cancer. 8:976–990. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Wang H, Li Z, Wu Q, Lathia JD,
McLendon RE, Hjelmeland AB and Rich JN: c-Myc is required for
maintenance of glioma cancer stem cells. PLoS One. 3:e37692008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Porro A, Haber M, Diolaiti D, Iraci N,
Henderson M, Gherardi S, Valli E, Munoz MA, Xue C, Flemming C, et
al: Direct and coordinate regulation of ATP-binding cassette
transporter genes by Myc factors generates specific transcription
signatures that significantly affect the chemoresistance phenotype
of cancer cells. J Biol Chem. 285:19532–19543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Porro A, Iraci N, Soverini S, Diolaiti D,
Gherardi S, Terragna C, Durante S, Valli E, Kalebic T, Bernardoni
R, et al: c-MYC oncoprotein dictates transcriptional profiles of
ATP-binding cassette transporter genes in chronic myelogenous
leukemia CD34+ hematopoietic progenitor cells. Mol
Cancer Res. 9:1054–1066. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xie ZY, Lv K, Xiong Y and Guo WH:
ABCG2-meditated multidrug resistance and tumor-initiating capacity
of side population cells from colon cancer. Oncol Res Treat.
37:666–668, 670–672. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang Y, Hou J, Li G, Song Z, Li X, Yang C,
Liu W, Hu Y and Xu Y: ABCG2 regulates the pattern of self-renewing
divisions in cisplatin-resistant non-small cell lung cancer cell
lines. Oncol Rep. 32:2168–2174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xie N, Mou L, Yuan J, Liu W, Deng T, Li Z,
Jing Y and Hu Z: Modulating drug resistance by targeting
BCRP/ABCG2 using retrovirus-mediated RNA interference. PLoS
One. 9:e1034632014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kugimiya N, Nishimoto A, Hosoyama T, Ueno
K, Enoki T, Li TS and Hamano K: The c-MYC-ABCB5 axis has a pivotal
role in 5-fluorouracil resistance in human colon cancer cells. J
Cell Mol Med. 19:1569–1581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Amini S, Fathi F, Mobalegi J,
Sofimajidpour H and Ghadimi T: The expressions of stem cell
markers: Oct4, Nanog, Sox2, nucleostemin, Bmi, Zfx, Tcl1, Tbx3,
Dppa4, and Esrrb in bladder, colon, and prostate cancer, and
certain cancer cell lines. Anat Cell Biol. 47:1–11. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cao L, Bombard J, Cintron K, Sheedy J,
Weetall ML and Davis TW: BMI1 as a novel target for drug discovery
in cancer. J Cell Biochem. 112:2729–2741. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
James MI, Iwuji C, Irving G, Karmokar A,
Higgins JA, Griffin-Teal N, Thomas A, Greaves P, Cai H, Patel SR,
et al: Curcumin inhibits cancer stem cell phenotypes in ex vivo
models of colorectal liver metastases, and is clinically safe and
tolerable in combination with FOLFOX chemotherapy. Cancer Lett.
364:135–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Todaro M, Alea MP, Di Stefano AB,
Cammareri P, Vermeulen L, Iovino F, Tripodo C, Russo A, Gulotta G,
Medema JP, et al: Colon cancer stem cells dictate tumor growth and
resist cell death by production of interleukin-4. Cell Stem Cell.
1:389–402. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kreso A, van Galen P, Pedley NM,
Lima-Fernandes E, Frelin C, Davis T, Cao L, Baiazitov R, Du W,
Sydorenko N, et al: Self-renewal as a therapeutic target in human
colorectal cancer. Nat Med. 20:29–36. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zeuner A, Todaro M, Stassi G and De Maria
R: Colorectal cancer stem cells: From the crypt to the clinic. Cell
Stem Cell. 15:692–705. 2014. View Article : Google Scholar : PubMed/NCBI
|