Introduction
Liver cancer has the fourth highest cancer mortality
rate worldwide, and China is a country with a high incidence rate
of liver cancer, where it accounts for ~40% of the total number of
cancer cases and cases of cancer-associated mortality (1,2). Surgery
remains the most effective treatment option for patients with liver
cancer (3,4). The majority of liver cancer cases are
diagnosed at an advanced or unresectable stage; therefore, novel
therapeutic strategies and therapeutic targets are required.
Current studies mainly focus on the pathogenic genes and molecular
mechanisms involved in liver cancer (5,6).
HECT domain and ankyrin repeat containing E3
ubiquitin protein ligase 1 (HACE1) is a member of the HECT
domain-containing E3 ligase family and was originally identified to
be associated with the occurrence of Wilms' tumour (7). Furthermore, additional studies revealed
that lower expression or mutations of HACE1 are associated with
numerous types of human malignant tumours, including breast cancer,
colorectal cancer, lung cancer, liver cancer, gastric cancer and
lymphoma (5,8–12), which
suggests that HACE1 functions as a tumour suppressor. Additional
research concerning HACE1 has focused on a variety of downstream
pathways; for example, HACE1 has been reported to act as a tumour
suppressor by ubiquitinating optineurin (OPTN) and activating
selective autophagy (10).
In mammalian cells, DNA methylation is critical for
the regulation of gene expression and, therefore, serves a pivotal
role in numerous physiological and pathological processes (13). DNA hypermethylation of tumour
suppressor genes silences their expression and contributes to
several types of human cancer (9,14).
Targeted DNA demethylation via the widely used clustered regularly
interspaced short palindromic repeat (CRISPR)-CRISPR-associated
(Cas) system has been widely reported on in recent years (15–17). A
strategy for targeted demethylation of specific genomic loci by
tethering Tet1-CD to the MS2 RNA element-containing single guide
RNA (sgRNA) 2.0 system-guided dCas9 and MS2 bacteriophage coat
proteins was the first sgRNA and Cas9-mediated demethylation system
to be reported (15,17).
In the present study, low HACE1 expression was
identified in human liver cancer cell lines compared with in a
normal liver cell line. Subsequently, HACE1 expression was
activated via targeted DNA demethylation using a two-plasmid
system. Finally, in the present study, increased HACE1 expression
was revealed to inhibit proliferation and activate selective
autophagy in liver cancer cells.
Materials and methods
Plasmid construction
The pdCas9-Tet1-CD and pcDNA3.1-MS2-Tet1-CD plasmids
were provided by Professor Ronggui Hu (Chinese Academy of Sciences,
Shanghai, China). sgRNAs targeting HACE1 were designed using an
online tool (version 1.2; http://crispr.mit.edu/) as previously described
(18). The designed sgRNAs (Table I) were synthesized as
oligonucleotides (Sangon Biotech Co., Ltd., Shanghai, China),
annealed and inserted into the pdCas9-Tet1-CD vector, which was
digested with BbsI.
 | Table I.Sequences of the sgRNA target sites
used in construction of the pdCas9-Tet1-CD expression plasmids. |
Table I.
Sequences of the sgRNA target sites
used in construction of the pdCas9-Tet1-CD expression plasmids.
| sgRNA for HACE1 | Target site sequence
(protospacer adjacent motif region) |
|---|
| sgRNA1 |
5′-GCCCTGGGCGGAGTCACGTTGGG-3′ |
| sgRNA2 |
5′-GCGCCCAGGCCACGCCAACGCGG-3′ |
| sgRNA3 |
5′-TGGGCGTACTCCTAAGCTTCTGG-3′ |
| sgRNA4 |
5′-GAGTACGCCCAGTCGCTGCGTGG-3′ |
| sgRNA5 |
5′-CCTGCCGGGCGGCTTTATGAGGG-3′ |
| sgRNA6 |
5′-CCCTCATAAAGCCGCCCGGCAGG-3′ |
| sgRNA7 |
5′-CGTTGATGATGTATGTTGGCTGG-3′ |
Cell culture and transfection
The human liver cancer cell lines HepG2 and Huh7,
and the normal liver cell line L02 were obtained from the Cell Bank
of the Chinese Academy of Sciences (Shanghai, China). The human
liver cancer cell line Hep3B was purchased from the American Type
Culture Collection (Manassas, VA, USA). All four cell lines have
been authenticated by short tandem repeat profiling within the last
2 years. All cell lines were cultured in Dulbecco's modified
Eagle's medium supplemented with 10% foetal bovine serum, 100 U/ml
penicillin and 100 mg/ml streptomycin (all Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in a 37°C humidified atmosphere
with 5% CO2. For the demethylation experiments, Hep3B or
HepG2 cells (104 cells/well) were seeded into plates and
the pdCas9-Tet1-CD (10 µg) and pcDNA3.1-MS2-Tet1-CD (8 µg) plasmids
were transfected into cells using Lipofectamine® 2000
(Thermo Fisher Scientific, Inc.). The subsequent experiments were
performed 48 h post-transfection. For colony formation assays, cell
lines with stable expression of the demethylation system were
established as previously described (15).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using a total RNA
kit (Tiangen Biotech Co., Ltd., Beijing, China). cDNA was
synthesized using the ReverTra Ace qPCR RT Master Mix (Toyobo Life
Science, Osaka, Japan). The temperature protocol was as follows:
Incubation at 37°C for 15 min, 50°C for 5 min, and then 98°C for 5
min. qPCR was performed to assess the relative abundance of HACE1
mRNA using specific primers (Table
II) and SYBR Green dye (Toyobo Life Science) on an ABI 7500
fast real-time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Thermocycling conditions were as follows:
Initial denaturing at 94°C for 2 min followed by 40 cycles of 95°C
for 15 sec, 58°C for 15 sec and 72°C for 30 sec. The relative
abundance of HACE1 was normalized to that of GAPDH using the
2−ΔΔCq method (19). All
data were obtained from at least three independent experiments.
| Table II.Sequences of the primers used in
RT-qPCR and bisulphite DNA sequencing. |
Table II.
Sequences of the primers used in
RT-qPCR and bisulphite DNA sequencing.
| Target gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Primers for
RT-qPCR |
|
|
|
GAPDH |
GAGTCAACGGATTTGGTCGTATTG |
ATTTGCCATGGGTGGAATCATATTG |
|
HACE1 |
GCAAGAAATGGGCAGAAGAAATGTA |
CATCCTCAACATCAACATCACTGAC |
| Primers for
bisulphite |
|
|
| DNA sequencing |
|
|
| HACE1
promoter |
ATAGGGATATAATATAGTTTAA |
AAAAACTATAATTTCCAACTA |
Bisulphite DNA sequencing
Genomic DNA (gDNA) was extracted from cells of the
indicated groups (control cells and cells transfected with sgRNA 1,
4 and 5) using the standard phenol-chloroform extraction method
(20). gDNA was treated with
bisulphite using the CpGenome Turbo Bisulphite Modification kit
(EMD Millipore, Billerica, MA, USA) according to the manufacturer's
protocol. The modified DNA was amplified using Platinum Taq DNA
Polymerase (Thermo Fisher Scientific, Inc.) with the respective
primer sets that recognize bisulphite-modified DNA only (Table II). The cycling parameters were as
follows: Initial denaturing at 94°C for 2 min, followed by 33
cycles of 95°C for 15 sec, 58°C for 15 sec and 72°C for 30 sec.
Subsequently, the PCR products were cloned into the pMD18-T vector
(Takara Bio, Inc., Otsu, Japan) and were sent for Sanger sequencing
by Biosune Biotechnology, Co. (Shanghai, China).
Cell proliferation assay
Cells stably transfected with the demethylation
system were seeded into a 96-well plate (3,000 cells/well). For
this experiment, 6 h post-cell seeding was defined as the 0 h time
point. After 0, 24 or 48 h, the cells were incubated with MTT
solution (cat. no. C0009; Beyotime Institute of Biotechnology,
Haimen, China) for 4 h at 37°C. The formazan product was then
dissolved in dimethyl sulfoxide and quantified
spectrophotometrically at a wavelength of 570 nm using a microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Experiments
were performed in triplicate and repeated three times.
Colony formation assay
Cells stably transfected with the demethylation
system were seeded into 6-well plates (1,000 cells/well). After 7
days, the cells were fixed with 4% paraformaldehyde (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) on ice for 30 min and stained with
0.1% crystal violet (cat. no. C0121; Beyotime Institute of
Biotechnology) for 20 min at room temperature.
IP and immunoblotting
For IP, ~6 million cells transfected with the
demethylation system were lysed in IP buffer (50 mM Tris-HCl, 5 mM
EDTA, 0.1% SDS and 1% Nonidet P-40) supplemented with a protease
inhibitor cocktail (Roche Diagnostics, Basel, Switzerland). Protein
concentration of cell lysates was determined using the Pierce™
Bicinchoninic Acid Protein Assay Kit (Thermo Fisher Scientific,
Inc.) and 2 mg protein of the whole-cell-lysate was incubated with
OPTN antibody (1:100 dilution; cat. no. 10837-1-AP; Proteintech
Group, Inc., Chicago, IL, USA) and protein G agarose beads (cat.
no. 16-266, Merck KGaA) overnight at 4°C. The immunoprecipitants
were enriched and denatured at 100°C for 10 min in 2X SDS-PAGE
loading buffer. The whole-cell-lysate input (20 µg), and
immunoprecipitants were then separated by SDS-PAGE (4%
concentrating gel and 10% separating gel), and transferred onto
polyvinylidene difluoride membranes (Bio-Rad Laboratories, Inc.).
Membranes were blocked in 5% milk in TBS with 0.1% Tween-20 for 1 h
at room temperature. The membranes were incubated at room
temperature for 2 h with primary antibodies against ubiquitin
(1:1,000 dilution; cat. no. sc-47721; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), OPTN (1:2,000 dilution), HACE1 (1:1,000
dilution; cat. no. ab133637; Abcam, Cambridge, UK), GAPDH (1:5,000
dilution; cat. no. 60004-1-Ig; Proteintech Group, Inc.) and
microtubule-associated proteins 1A/1B light chain 3B (LC3; 1:500
dilution; cat. no. L7543; Sigma-Aldrich; Merck KGaA). Membranes
were incubated with horseradish peroxidase(HRP)-conjugated goat
anti-mouse IgG (H+L) or HRP-conjugated goat anti-rabbit IgG (H+L)
secondary antibodies (1:5,000 dilution; cat. nos. G-21040 and
31460; Thermo Fisher Scientific, Inc.) for 1 h at room temperature.
Protein was then labelled by adding 1 ml SuperSignal™ West Pico
PLUS Chemiluminescent Substrate (cat. no. 34577; Thermo Fisher
Scientific, Inc.). The signal was visualized using the Tanon 5200
Imaging System (Tanon Science and Technology Co., Ltd., Shanghai,
China).
Statistical analysis
All experiments were performed in triplicate. All
values are presented as the means ± standard deviation. One-way
analysis of variance was performed with Tukey's post hoc multiple
comparisons test using GraphPad Prism software (version 5; GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
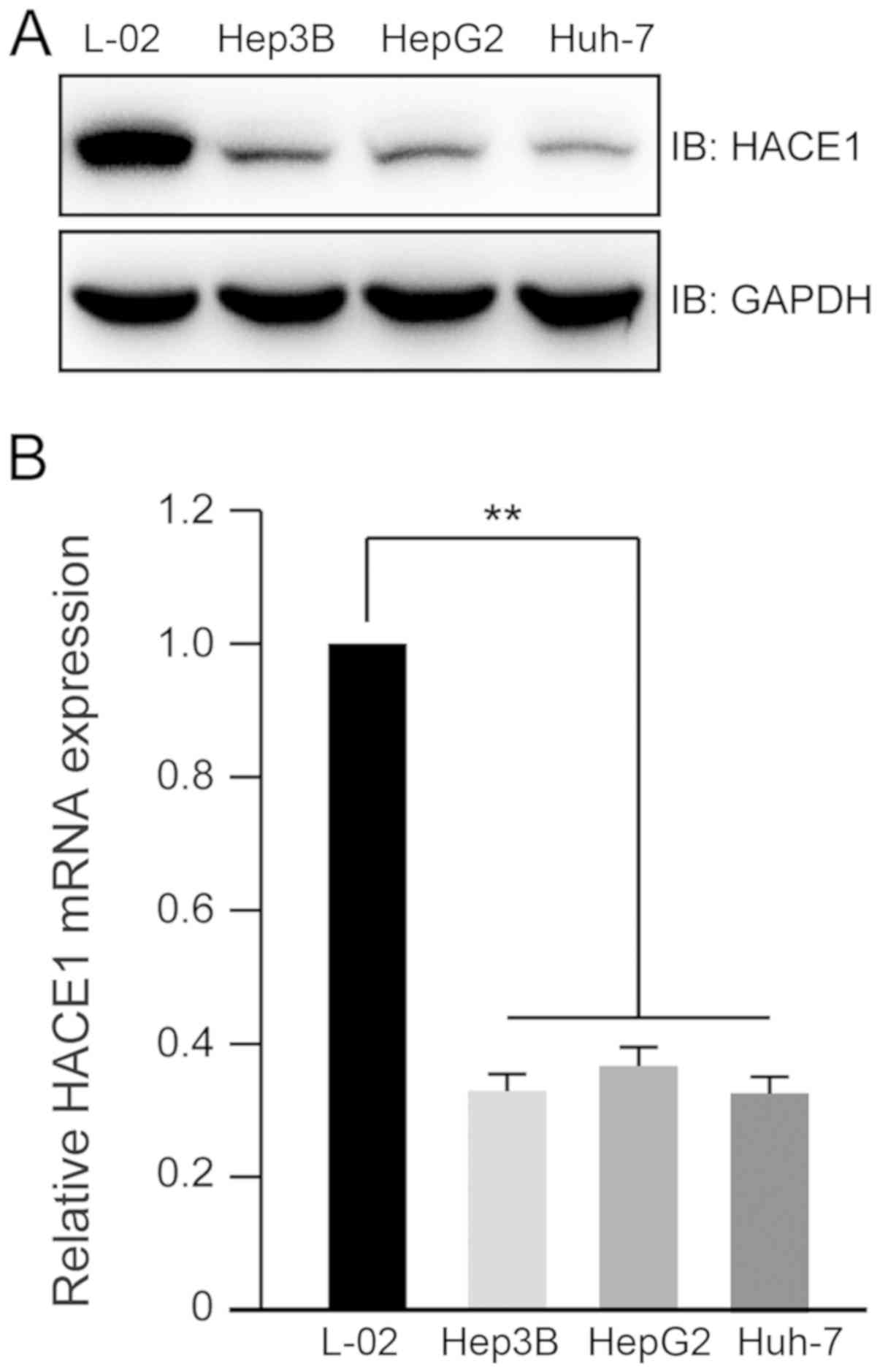
Lower expression of HACE1 in human
liver cancer cell lines
To explore the expression profile of HACE1 in human
liver cancer cell lines, lysates from three liver cancer cell lines
(Hep3B, HepG2 and Huh-7) and one normal liver cell line (L-02) were
prepared. Immunoblot analysis indicated that the protein levels of
HACE1 decreased in the three liver cancer cell lines compared with
in the normal liver cell line (Fig.
1A). The mRNA expression levels of HACE1 in the normal liver
cell line were ~4 times higher than those in the liver cancer cell
lines (Fig. 1B).
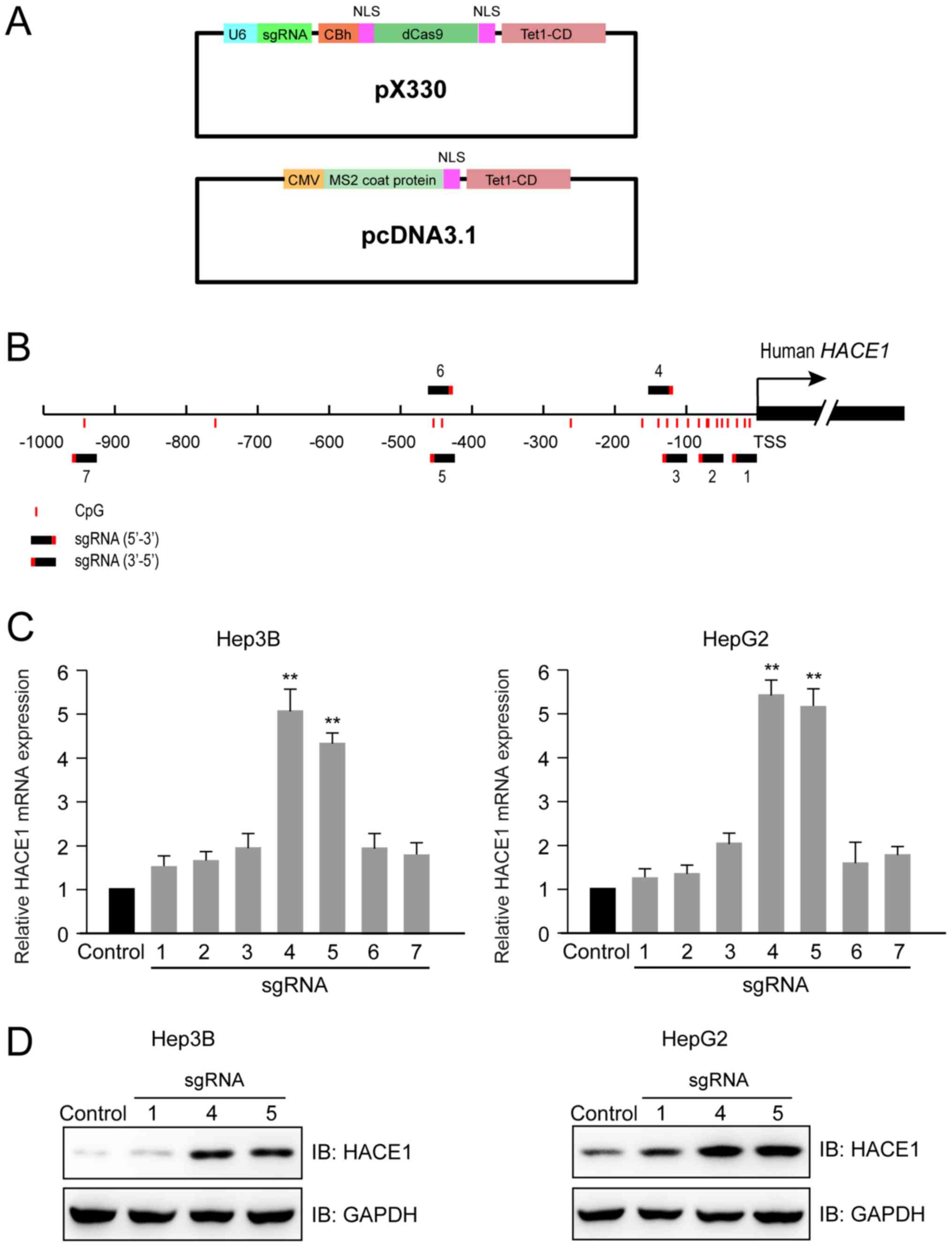
CRISPR-Cas-based HACE1 promoter
demethylation sgRNA design and activity detection
Using the previously described (14) two-plasmid demethylation system
(Fig. 2A), seven sgRNAs were
designed that targeted regions between −1,000 bp and the
transcription start site (TSS) of the human HACE1 gene (Fig. 2B). At 48 h after the transfection of
Hep3B and HepG2 cells with dCas9-Tet1-CD (sgRNAs 1–7) and
MS2-Tet1-CD, two sgRNAs (4 and 5) were identified to increase the
transcription of HACE1 mRNA by 4–5 times compared with the control
group with no sgRNA (Fig. 2C).
Furthermore, immunoblot analysis indicated that the protein levels
of HACE1 were evidently increased in cells transfected with the
aforementioned two sgRNAs (4 and 5) compared with the control group
or sgRNA 1, which had little effect on HACE1 gene expression and
was used as a negative control (Fig.
2D).
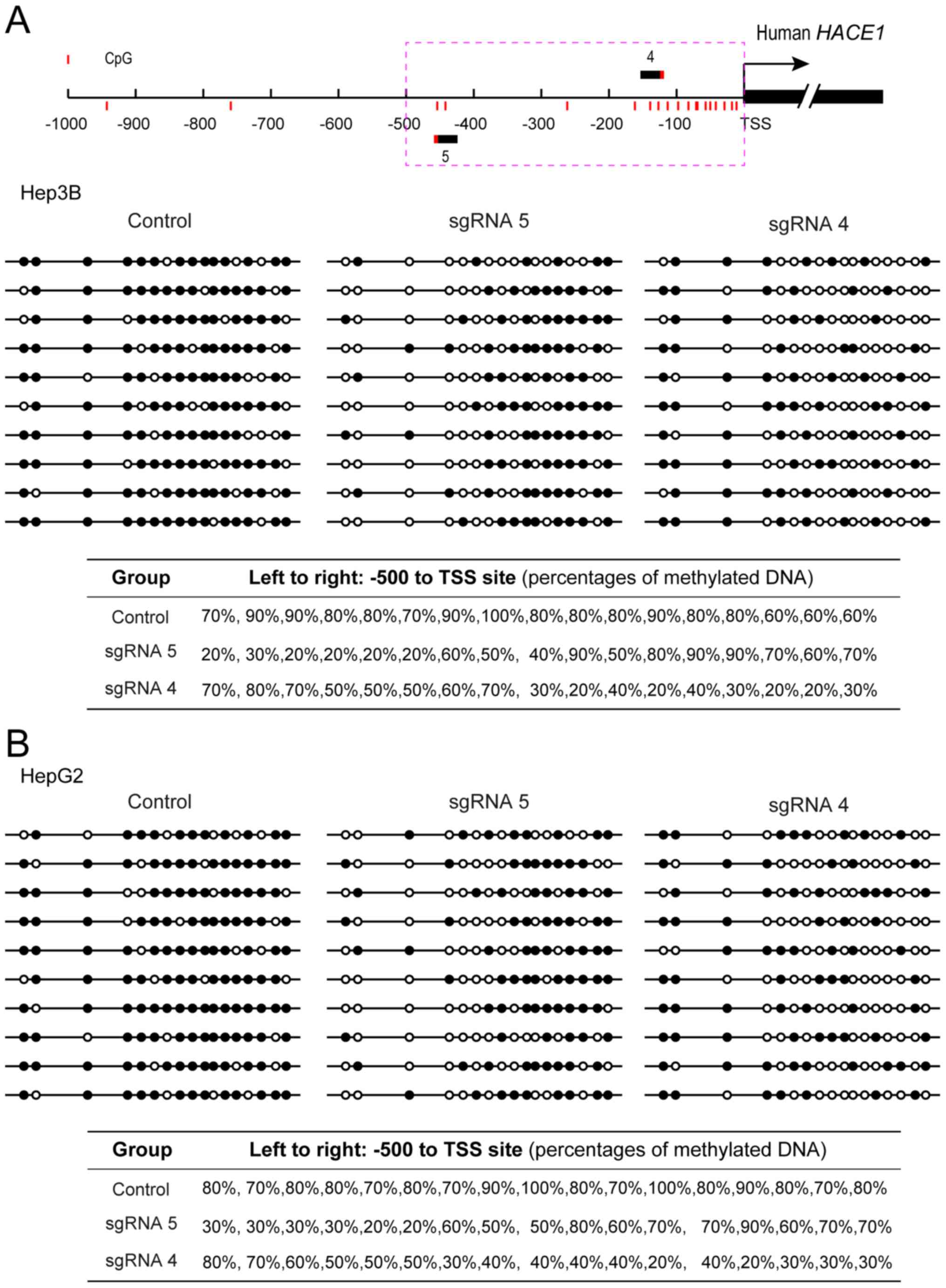
Upregulation of the target HACE1 gene
by specific DNA demethylation
To determine whether the upregulated HACE1 gene
transcription was a direct result of targeted demethylation that
occurred at a specific HACE1 promoter sequence, the methylation
status of sgRNA-targeted loci (between −500 bp and TSS) was
examined using a bisulphite sequencing approach. As shown in
Fig. 3A and B, expression of the
two-plasmid demethylation system resulted in the removal of methyl
groups from the neighbouring CpG islands of the HACE1 gene promoter
in the Hep3B and HepG2 cell lines. Additionally, the percentages of
methylated CpG for each CpG island are shown.
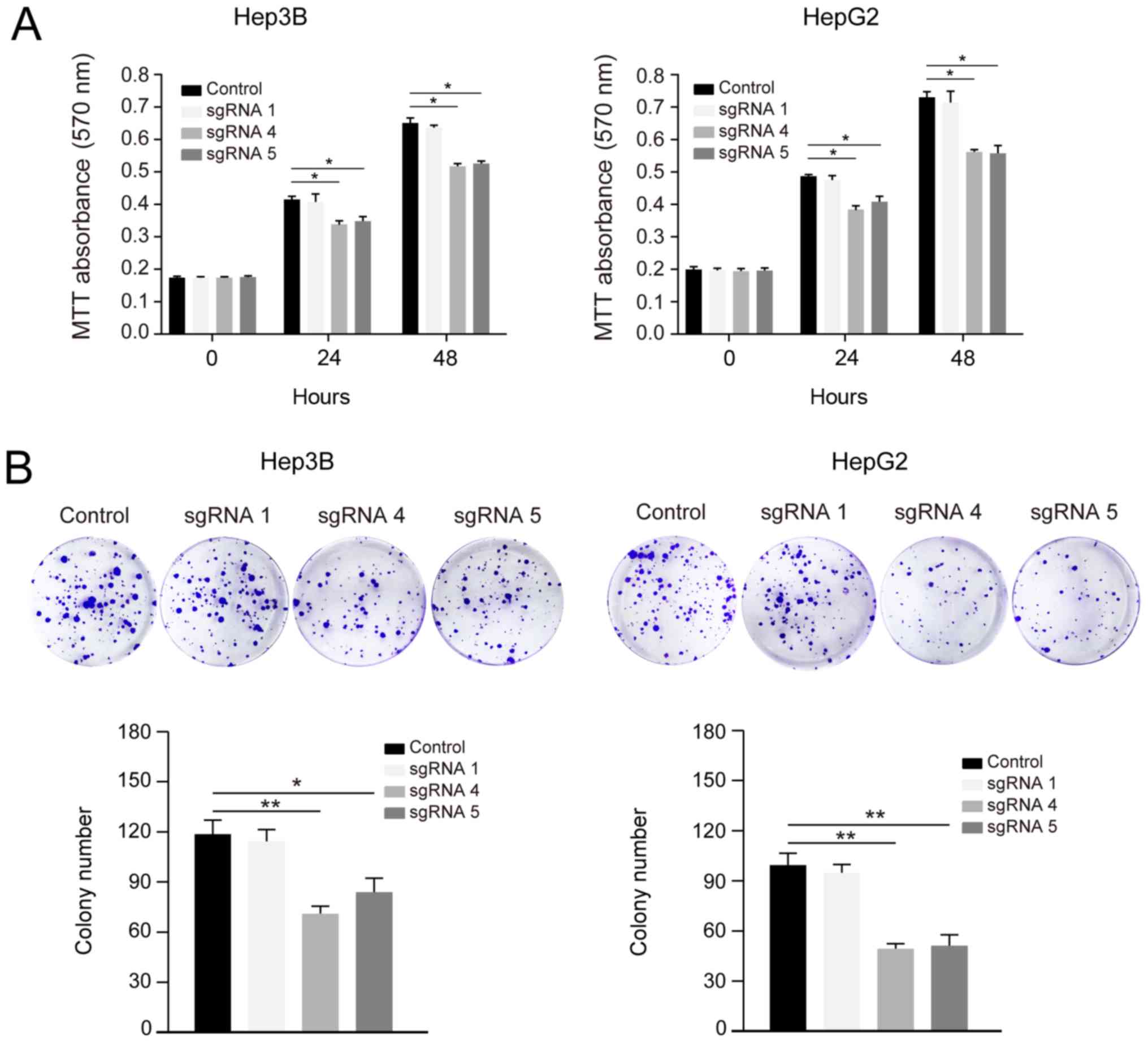
Demethylation of HACE1 inhibits human
liver cancer cell proliferation and colony formation
A previous study reported that HACE1 has a
tumour-suppressive role in hepatocellular carcinoma (5). To evaluate whether CRISPR-Cas-induced
upregulation of HACE1 resulted in physiologically relevant effects,
the proliferation of Hep3B and HepG2 cells was detected by MTT
assays at 24 and 48 h post transfection. sgRNAs 4 and 5-induced
HACE1 upregulation significantly inhibited liver cancer cell
proliferation compared with the control and sgRNA 1 groups
(Fig. 4A).
The demethylation systems containing sgRNAs 4 and 5
were stably expressed in Hep3B and HepG2 cells; subsequently, these
cells underwent colony formation assays. Decreased colony numbers
were observed in the sgRNA 4 and 5 groups compared with in the
control and sgRNA 1 groups (Fig.
4B). These data suggested that CRISPR-Cas-based upregulation of
HACE1 resulted in physiologically relevant effects and inhibited
liver cancer cell proliferation.
Demethylation of HACE1 promotes OPTN
ubiquitination and autophagy in liver cancer cells
Ubiquitination of the autophagy receptor OPTN by
HACE1 has been demonstrated to activate selective autophagy,
resulting in tumour suppression in lung cancer (10). In the present study, it was
investigated if this phenomenon could also occur in liver cancer
cells. The ubiquitination of OPTN markedly increased when cells
were transfected with demethylation system vectors containing
sgRNAs 4 or 5 (Fig. 5). LC3 is a
central protein in the autophagy pathway, where it functions in
autophagy substrate selection and autophagosome biogenesis. As a
result, LC3 is the most widely used marker of autophagosomes. The
ratio of LC3 II to LC3 I clearly increased when Hep3B and HepG2
cells were transfected with demethylation system vectors containing
sgRNAs 4 or 5 compared with the control and sgRNA 1 groups
(Fig. 5). Collectively, these
results suggested that increased expression of HACE1 by targeted
demethylation may promote OPTN ubiquitination and autophagy
activity in liver cancer cells.
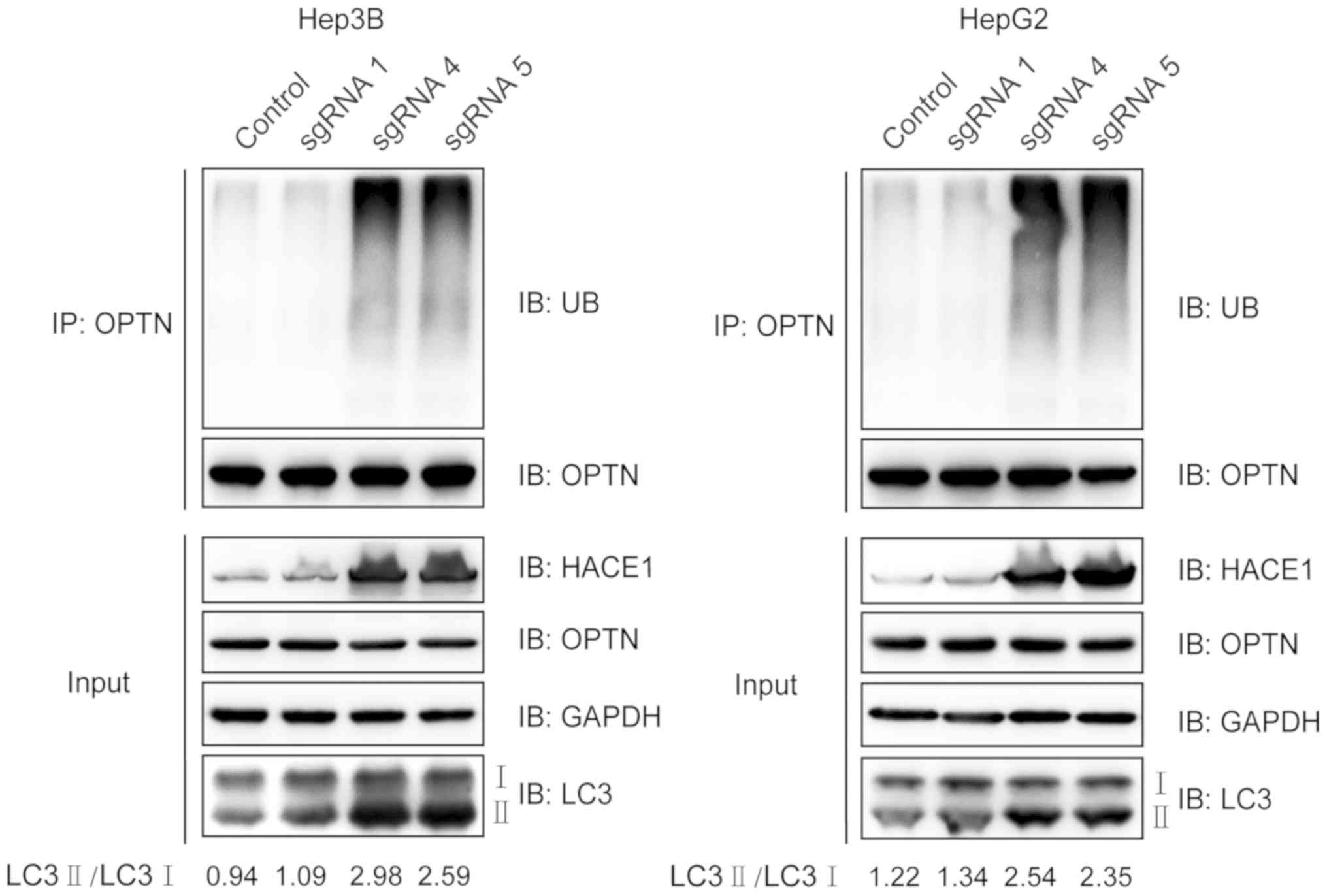 | Figure 5.Demethylation of HACE1 promotes OPTN
ubiquitination and autophagy. sgRNA-mediated HACE1 demethylation
promoted OPTN ubiquitination. Hep3B or HepG2 cells were
co-transfected with sgHACE1 (1, 4 and 5)-guided dCas9-Tet1-CD and
MS2-Tet1-CD for 48 h. Subsequently, cell lysates were
immunoprecipitated with an anti-OPTN antibody and subjected to
immunoblotting. LC3 II/I expression was also detected. Inputs
refers to whole cell lysates, and GAPDH was used as a loading
control. Cas, clustered regularly interspaced short palindromic
repeats-associated; HACE1, HECT domain and ankyrin repeat
containing E3 ubiquitin protein ligase 1; IB, immunoblotting; LC3,
microtubule-associated proteins 1A/1B light chain 3B; OPTN,
optineurin; sgRNA, single guide RNA; UB, ubiquitin. |
Discussion
HACE1 downregulation has been identified in numerous
types of cancer, including hepatocellular carcinoma, breast cancer,
colorectal cancer, gastric cancer and leukaemia (5,11).
Previous studies have demonstrated that decreased expression or
deletion of HACE1 caused by HACE1 methylation or ubiquitination is
associated with the occurrence and invasion of various types of
carcinoma (18,21–23).
HACE1 is a candidate tumour suppressor gene and a potential
therapeutic target for several types of human cancer, including
liver cancer. The results of the present study indicated that
demethylation of the HACE1 promoter enhanced its expression, and
inhibited proliferation and colony formation of liver cancer cells.
However, a previous study demonstrated that HACE1 promotes melanoma
cell migration and adhesion in vitro and that it is required
for mouse lung colonization by melanoma cells in vivo
(24). These findings indicated that
whether HACE1 acts as a tumour suppressor gene or oncogene may
depend on the cancer type.
HACE1 was first studied in Wilms' tumour and was
later identified to be frequently lost or downregulated in a
variety of tumours. Its role in tumour suppression has been
extensively investigated, and a number of studies have indicated
that HACE1 can restrain reactive oxygen species generation, control
cell fate by regulating TNF receptor superfamily member 1A, and
impede tumour growth by accelerating the ubiquitination of Rac
family small GTPase 1 (11,25,26).
Notably, it has been reported that HACE1 acts as a tumour
suppressor by ubiquitinating OPTN, and that it activates selective
autophagy (10). The present study
indicated that demethylation of the HACE1 promoter may lead to OPTN
ubiquitination and elevated protein levels of LC3 II.
A previous study indicated that the dCas9-based
demethylation system has non-additive effects (14). In the present study, the two-plasmid
demethylation system efficiently removed the methyl groups from
neighbouring CpG islands on the HACE1 gene promoter in liver cancer
cells.
In conclusion, further efforts are required to apply
the dCas9-based demethylation system in animal models to
investigate liver cancer that is aetiologically caused by HACE1
gene hypermethylation. Activating the expression of HACE1 may be a
promising approach for anticancer therapy.
Acknowledgements
The authors would like to thank Professor Ronggui Hu
(Institute of Biochemistry and Cell Biology, Shanghai Institutes
for Biological Sciences, Chinese Academy of Sciences) for providing
the demethylation system plasmids (pdCas9-Tet1-CD and
pcDNA3.1-MS2-Tet1-CD) and technical support.
Funding
This study was supported by research grants from the
National Natural Science Foundation Youth Fund of China (grant no.
81702622) and Liaoning Province Doctoral Startup Fund (grant no.
201501022).
Availability of data and materials
All data generated or analysed during this study are
included in the published article.
Authors' contributions
ZY and ZL conceived and designed the experiments.
ZY, YL and TH performed the experiments, collected the data and
analysed the results. ZY and ZL wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clinicians. 65:87–108. 2015. View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Waller LP, Deshpande V and Pyrsopoulos N:
Hepatocellular carcinoma: A comprehensive review. World J Hepatol.
7:2648–2663. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Waghray A, Murali AR and Menon KN:
Hepatocellular carcinoma: From diagnosis to treatment. World J
Hepatol. 7:1020–1029. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gao ZF, Wu YN, Bai ZT, Zhang L, Zhou Q and
Li X: Tumor-suppressive role of HACE1 in hepatocellular carcinoma
and its clinical significance. Oncol Rep. 36:3427–3435. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Villanueva A, Minguez B, Forner A, Reig M
and Llovet JM: Hepatocellular carcinoma: Novel molecular approaches
for diagnosis, prognosis, and therapy. Annu Rev Med. 61:317–328.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Anglesio MS, Evdokimova V, Melnyk N, Zhang
L, Fernandez CV, Grundy PE, Leach S, Marra MA, Brooks-Wilson AR,
Penninger J and Sorensen PH: Differential expression of a novel
ankyrin containing E3 ubiquitin-protein ligase, Hace1, in sporadic
Wilms' tumor versus normal kidney. Hum Mol Genet. 13:2061–2074.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goka ET and Lippman ME: Loss of the E3
ubiquitin ligase HACE1 results in enhanced Rac1 signaling
contributing to breast cancer progression. Oncogene. 34:5395–5405.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hibi K, Sakata M, Sakuraba K, Shirahata A,
Goto T, Mizukami H, Saito M, Ishibashi K, Kigawa G, Nemoto H and
Sanada Y: Aberrant methylation of the HACE1 gene is frequently
detected in advanced colorectal cancer. Anticancer Res.
28:1581–1584. 2008.PubMed/NCBI
|
|
10
|
Liu Z, Chen P, Gao H, Gu Y, Yang J, Peng
H, Xu X, Wang H, Yang M, Liu X, et al: Ubiquitylation of autophagy
receptor optineurin by HACE1 activates selective autophagy for
tumor suppression. Cancer Cell. 26:106–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen YL, Li DP, Jiang HY, Yang Y, Xu LL,
Zhang SC and Gao H: Overexpression of HACE1 in gastric cancer
inhibits tumor aggressiveness by impeding cell proliferation and
migration. Cancer Med. 7:2472–2484. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Y, de Reynies A, de Leval L, Ghazi
B, Martin-Garcia N, Travert M, Bosq J, Brière J, Petit B, Thomas E,
et al: Gene expression profiling identifies emerging oncogenic
pathways operating in extranodal NK/T-cell lymphoma, nasal type.
Blood. 115:1226–1237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Robertson KD: DNA methylation and human
disease. Nature reviews. Genetics. 6:597–610. 2005.PubMed/NCBI
|
|
14
|
Sakata M, Kitamura YH, Sakuraba K, Goto T,
Mizukami H, Saito M, Ishibashi K, Kigawa G, Nemoto H, Sanada Y and
Hibi K: Methylation of HACE1 in gastric carcinoma. Anticancer Res.
29:2231–2233. 2009.PubMed/NCBI
|
|
15
|
Xu X, Tao Y, Gao X, Zhang L, Li X, Zou W,
Ruan K, Wang F, Xu GL and Hu R: A CRISPR-based approach for
targeted DNA demethylation. Cell Discov. 2:160092016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morita S, Noguchi H, Horii T, Nakabayashi
K, Kimura M, Okamura K, Sakai A, Nakashima H, Hata K, Nakashima K
and Hatada I: Targeted DNA demethylation in vivo using
dCas9-peptide repeat and scFv-TET1 catalytic domain fusions. Nat
Biotechnol. 34:1060–1065. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu XS, Wu H, Krzisch M, Wu X, Graef J,
Muffat J, Hnisz D, Li CH, Yuan B, Xu C, et al: Rescue of fragile X
syndrome neurons by DNA methylation editing of the FMR1 gene. Cell.
172:979–992 e976. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu X, Li C, Gao X, Xia K, Guo H, Li Y, Hao
Z, Zhang L, Gao D, Xu C, et al: Excessive UBE3A dosage impairs
retinoic acid signaling and synaptic plasticity in autism spectrum
disorders. Cell Res. 28:48–68. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Smith-Ravin J, England J, Talbot IC and
Bodmer W: Detection of c-Ki-ras mutations in faecal samples from
sporadic colorectal cancer patients. Gut. 36:81–86. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mettouchi A and Lemichez E: Ubiquitylation
of active Rac1 by the E3 ubiquitin-ligase HACE1. Small GTPases.
3:102–106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gacon G, Mettouchi A and Lemichez E: The
tumor suppressor HACE1 targets Rac1 to ubiquitin-mediated
proteasomal degradation. Med Sci (Paris). 28:39–41. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lachance V, Degrandmaison J, Marois S,
Robitaille M, Génier S, Nadeau S, Angers S and Parent JL:
Ubiquitylation and activation of a Rab GTPase is promoted by a
beta(2)AR-HACE1 complex. J Cell Sci. 127:111–123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
El-Hachem N, Habel N, Naiken T, Bzioueche
H, Cheli Y, Beranger GE, Jaune E, Rouaud F, Nottet N, Reinier F, et
al: Uncovering and deciphering the pro-invasive role of HACE1 in
melanoma cells. Cell Death Differ. 25:2010–2022. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cetinbas N, Daugaard M, Mullen AR, Hajee
S, Rotblat B, Lopez A, Li A, De Berardinis RJ and Sorensen PH: Loss
of the tumor suppressor Hace1 leads to ROS-dependent glutamine
addiction. Oncogene. 34:4005–4010. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tortola L, Nitsch R, Bertrand MJM, Kogler
M, Redouane Y, Kozieradzki I, Uribesalgo I, Fennell LM, Daugaard M,
Klug H, et al: The tumor suppressor hace1 is a critical regulator
of TNFR1-mediated cell fate. Cell Rep. 16:34142016. View Article : Google Scholar : PubMed/NCBI
|