Introduction
Cervical cancer is the fourth most frequent cause of
mortality in women worldwide (1). It
has a prevalence of 20% in developed countries, whereas there is a
high incidence of invasive cases in developing countries, including
Thailand, due to the lack of cancer prevention strategies, leading
to ~90% cervical cancer-associated mortalities (2–4). This
type of cancer can take from one to numerous years to progress from
a precancerous state to cancer since it is a slow progressive
disease. The precancerous state is known as cervical
intraepithelial neoplasia (CIN) and is categorized according to the
cell abnormality levels (CIN I, II and III) (5). Human papillomavirus (HPV) is the main
etiological factor of cervical cancer (6). It can be classified into many types;
the high-risk HPV types (including HPV 16, 18 and 52) are commonly
associated with cervical cancer.
In the clinic, there are several methods of
treatment for cervical cancer, including surgery (7,8),
chemotherapy (hormone), radiotherapy or various combinations of
these methods (9–12). However, the mortality rate of
cervical cancer is still high (13).
Currently, several screening tests for cervical cancer are
available, including conventional Pap smears, liquid-based cytology
and HPV DNA testing. From previous studies, a combination of
liquid-based cytology and DNA testing has demonstrated a high
sensitivity in cervical cancer screening. However, this technique
requires cervical swabs, which are complicated and highly invasive
when compared with the collection of urine. Therefore, a less
invasive protocol is required for diagnosis. The detection of
urinary proteins is a good alternative that may lead to the
discovery of cancer biomarkers (14).
Proteomic techniques, including two-dimensional gel
electrophoresis, mass spectrometry (MS) and validation by
immunodetection, have been used to identify protein biomarkers of
diseases, especially in different types of cancers (15). Recently, MS-based proteomics has
become an increasingly powerful technique, not only for the
high-throughput identification of large numbers of proteins, but
also for protein quantification (16). Protein quantification can increase
the amount of important information obtained from identified
proteomes, and can compare protein expressions across various types
of samples or treatments. Using MS for comparative protein
quantification typically employs stable isotope incorporation, but
recently comparative quantification by label-free liquid
chromatography (LCn)-MS proteomics data has provided an
alternative approach (17).
The aim of the present study was to employ the
quantitative label-free MS technique to compare the profiles of
protein expression between the urine of normal HPV negative
patients and cervical cancer patients. We did not include
preinvasive diseases as cell abnormality levels (CIN I, II and
III). The results may elucidate the protein-protein interactions
and the potential urinary detection biomarkers of cervical
cancer.
Materials and methods
Collection and preparation of
samples
Cytobrush and urine samples were obtained from
cervical cancer patients and healthy volunteers at the Chulabhorn
Hospital from July 2014 to April 2015. All participants received
detailed information regarding the study objectives and gave
written informed consent. The present study was approved by the
Ethical Review Board of the Chulabhorn Hospital (Bangkok, Thailand;
no. 31/2554). All participants were subsequently performed HPV
genotyping analysis and normal routine cervical cancer screening at
Chulabhorn Hospital. The samples for HPV genotyping were obtained
using a cytobrush by gynecologists or well-trained general
practitioners during pelvic examinations at Chulabhorn Hospital.
The first morning urine samples were collected and fractionated
within 24 h. Each specimen was collected in 10 ml conical tubes and
the protease inhibitor cocktails (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) were added. The detail of the 13 urine samples
from HPV-negative females who were diagnosed as non-cervical cancer
was shown in Table IA. While, the
other 24 urine samples were collected from females with cervical
cancer at different stages (S1B1, SIB2, SIIA, SIIA1, SIIB, SIIIB,
SIVA and SIVB) as shown in Table
IB.
 | Table I.Clinicopathological data of normal
individuals and patients with cervical cancer. |
Table I.
Clinicopathological data of normal
individuals and patients with cervical cancer.
| A, Normal |
|---|
|
|---|
| Sample no. | Age, years | HPV genotype | CA state | Pathology
information |
|---|
| N1 | 47 | Negative | – | – |
| N2 | 35 | Negative | – | – |
| N3 | 49 | Negative | – | – |
| N4 | 49 | Negative | – | – |
| N5 | 52 | Negative | – | – |
| N6 | 52 | Negative | – | – |
| N7 | 42 | Negative | – | – |
| N8 | 44 | Negative | – | – |
| N9 | 40 | Negative | – | – |
| N10 | 44 | Negative | – | – |
| N11 | 53 | Negative | – | – |
| N12 | 55 | Negative | – | – |
| N13 | 64 | Negative | – | – |
|
| B,
Cancer |
|
| Sample
no. | Age,
years | HPV
genotype | CA
state | Pathology
information |
|
|---|
| C1 | 38 | Negative | IB1 | Adenocarcinoma in
situ |
| C2 | 44 | 33 | IB1 | Squamous cell
carcinoma, non-keratinizing moderately differentiated |
| C3 | 51 | 16 | IIA | Squamous cell
carcinoma, non-keratinizing moderately differentiated |
| C4 | 50 | 68 | IIA | Adenocarcinoma,
moderately-poorly differentiated |
| C5 | 56 | Negative | IIA1 | Squamous cell
carcinoma, papillary type |
| C6 | 51 | 59 | IIB | Squamous cell
keratinizing carcinoma |
| C7 | 54 | 16 | IIB | Adenocarcinoma,
endocervical |
| C8 | 64 | 18 | IIB | Squamous cell
carcinoma, non-keratinizing, moderately-poorly |
|
|
|
|
| differentiated |
| C9 | 50 | 33, LR11 | IIB | Squamous cell
carcinoma, keratinizing type, moderately differentiated |
| C10 | 49 | 16 | IIB | Squamous cell
carcinoma |
| C11 | 64 | Negative | IIB | Adenocarcinoma,
endometrial moderately differentiated, metastasis |
| C12 | 42 | 16 | IIIB | Squamous cell
carcinoma, non-keratinizing moderately differentiated |
| C13 | 61 | 16 | IIIB | Squamous cell
carcinoma, non-keratinizing well-moderately |
|
|
|
|
| differentiated |
| C14 | 50 | Negative | IIIB | Squamous cell
carcinoma, keratinizing type |
| C15 | 58 | 31 | IIIB | Squamous cell
carcinoma, non-keratinizing moderately differentiated, |
|
|
|
|
| metastasis |
| C16 | 49 | 52,33 | IVA | Squamous cell
carcinoma, non-keratinizing moderately differentiated |
| C17 | 49 | 18 | IVB | Adenocarcinoma,
Poorly differentiated |
| C18 | 59 | 52 | IVB | Squamous cell
carcinoma, non-keratinizing, moderate differentiated |
| C19 | 59 | 18 | IIB | Adenocarcinoma,
endocervical like, metastasis |
| C20 | 57 | 53 | IB2 | Squamous cell
carcinoma, keratinizing poorly differentiated |
| C21 | 33 | 16 | IIB | Squamous cell
carcinoma, non-keratinizing moderately differentiated |
| C22 | 55 | Negative | IIB | Endometrium biopsy,
poorly differentiated |
| C23 | 60 | 16 | IIB | Squamous cell
carcinoma |
| C24 | 61 | LR 72 | IIB | Adenocarcinoma,
poorly differentiated, metastasis |
Cytology examination
The Bethesda 2001 report system was used as the
standard protocol for classification of all cervical cytology
slides (18). The different grades,
such as squamous cell carcinoma and adenocarcinoma, were determined
using the cervical cytology slides by qualified pathologists via a
normal routine at Chulabhorn Hospital.
HPV genotyping
The Linear array HPV testing (Roche Diagnostics
Indianapolis, IN, USA) was employed to identify HPV genotypes. This
kit was for identification of 37 HPV genotypes including 12
high-risk, 8 probable high-risk and 17 low-risk types. Briefly, 450
bp fragments of the L1 region of HPV were first amplified by
polymerase chain reaction, followed by hybridization using a
reverse line blot system for the simultaneous detection of up to 37
HPV genotypes (i.e., genotypes 6, 11, 16, 18, 26, 31, 33, 35, 39,
40, 42, 45, 51, 52, 53, 54, 55, 56, 58, 59, 61, 62, 64, 66, 67, 68,
69, 70, 71, 72, 73, 81, 82, 83, 84, IS39, and CP6108) (19).
Sample preparation
Urine samples were stored at 4°C and then
centrifuged at 1,120 × g for 5 min at 4°C to remove cell debris.
The supernatants were precipitated with 50% v/v acetone for 16 h
followed by centrifugation at 12,000 × g for 10 min at 4°C. The
pellets were kept at −80°C until further use.
In solution digestion
The pellets of urine samples from 5 normal (N9-N13)
and from 6 cervical cancer patients at different stages (C19-C24)
were resuspended in 50 mM ammonium bicarbonate
(NH4HCO3) separately and the protein
concentration was determined by spectrophotometry using the
Bradford method (20). The samples
were pooled by obtaining 5 µg from each of the 5 normal urine
samples and 5 µg from each of the 6 cancer urine samples. A total
of 5 µg of the pooled samples were separately reduced with 100 mM
DTT (10 mM final concentration) for 5 min at 95°C and alkylated
with 1/10 volume of 200 mM iodoacetamide prior to incubation for 30
min in the dark at room temperature. Digestion was performed
overnight at 37°C by adding 1:50 (w/w) of sequencing grade trypsin
(Promega Corporation, Madison, WI, USA) to the protein in each
sample. The digestion reaction was stopped by adding formic acid to
reach a 1% final concentration and the samples were dried
completely by SpeedVac.
LC-MS/MS analysis
To prepare samples for label-free LC-MS/MS
quantification, the digested samples were dissolved in 0.1% formic
acid in H2O and separated on a nanoACQUITY system
(Waters Corporation, Milford, MA, USA). All pooled urine samples
were run in triplicate. The samples were injected into a
nanoACQUITY UPLC column (1.7 µm BEH, 75 µm × 200 mm C18) at a flow
rate of 300 nl/min. The column temperature was maintained at 40°C.
The LC gradient (1–50% B in 70 min, 50–90% B in 5 min, followed by
15 min on 90% B) was performed using 0.1% formic acid in
H2O as solvent A and 0.1% formic acid in ACN as solvent
B. The eluting peptides were analyzed directly via MS/MS on an
amaZon speed ion trap mass spectrometer (Bruker Corporation,
Billerica, MA, USA) equipped with a captive-electrospray ion
source. The positive mode was used with a spray voltage of 1,300 V
and the capillary temperature was set at 150°C. Mass spectra were
acquired from 400–1,400 m/z using parameters optimized at 922 m/z
with a target of 500,000 set for ion charge control and a maximum
acquisition time of 100 msec. The scan range was 50–3,000 m/z.
MS/MS data were processed by Bruker Compass 1.4 software (Bruker
Corporation).
Data analysis and label-free LC-MS
quantitative profiling
Progenesis label-free LC-MS software version 3.1
(Nonlinear Dynamics, Ltd., Newcastle upon Tyne, UK) was used to
process the raw data obtained from LC-MS/MS and to calculate the
significant changes. The retention time of each sample was aligned
and the reference sample was set up. For all of the replicates,
their retention times were aligned to the established reference and
the intensities of the peak were then normalized. Three criteria
were used to filter all data prior to exporting the MS/MS output
files to identify proteins using Mascot software (www.matrixscience.com), including: i) An analysis of
variance (ANOVA) with Tukey's post hoc test score shown between
experimental groups of P≤0.05; ii) featured mass peaks with +2, +3
and +4 as the charge states; and iii) only data with >1 isotopes
or peptides. From the aforementioned Progenesis software, all of
the exported MS/MS spectra were generated using a Mascot generic
file. Mascot software version 2.4.0 was used for the identification
of peptides. The SwissProt human protein database (Matrix Science,
Ltd., London, UK; www.matrixscience.com) program was set up as follows:
The MS/MS mass tolerance was set at 0.6 Da, peptide mass tolerance
was set to 1.2 Da, carbamidomethylation was set as a fixed
modification and ≤1 missed cleavages were allowed. A false
discovery rate threshold of 1% was applied and identification of
two or more unique peptides and two or more peptides were required
for positive identification, respectively.
Characterization, classification,
functional and protein- protein interactions analysis
Venn Diagrams web tool (http://www.interactivenn.net) was used to compare the
expression of proteins from cervical cancer patients between
identified proteins by LC-MS/MS and the database of Human Proteome
Atlas (www.proteinatlas.org/humanproteome/pathology/cervical+cancer).
The classification and functional analysis of differential
expressed proteins was performed using the PANTHER database
(www.pantherdb.org/). The Gene Ontology
(GO) standard was employed to categorize proteins according to
‘Molecular function’ and ‘Biological process’. The Search Tool for
Retrieval of Interacting Genes/Proteins (STRING) database
(string-db.org) version 9.0 was used to obtain
protein-protein interactions. Protein mode from STRING was
analyzed, and the interactions from upregulated and downregulated
urinary expressed proteins were identified.
Western blot analysis
The expression levels of proteins were analyzed by
western blot analysis. Leucine-rich α-2-glycoprotein (LRG1),
multimerin-1 (MMRN1), S100 calcium-binding protein A8 (S100A8),
serpin B3 (SERPINB3) and cluster of differentiation-44 antigen
(CD44) were selected to be validated for expression. The pellets of
8 normal (N1-N8) and 18 different stages (C1-C18) of cancer urine
samples were resuspended in 50 mM ammonium bicarbonate
(NH4HCO3). A total of 20 µg of urine samples
were diluted in sample buffer (50 mM Tris, pH 6.8, 10% glycerol, 2%
SDS and 0.01% bromophenol blue) and resolved by SDS-PAGE. Following
SDS-PAGE, the proteins were transferred onto 0.20 µm polyvinylidene
difluoride membranes (Pall Life Sciences, Port Washington, NY,
USA). The membranes were blocked with 5% (w/v) non-fat dry milk in
TBST [50 mM Tris, pH 8.0, 150 mM NaCl, and 0.1% Tween-20 (v/v)] for
1 h. Membranes were washed 10 min for 3 times in TBST and incubated
at 4°C overnight with the following primary antibodies: Mouse
anti-LRG1 (1:2,000; ab57992; Abcam, Cambridge, UK), rabbit
anti-MMRN1 (1:2,000; ab130585; Abcam), rabbit anti-S100A8 (1:1,000;
ab92331; Abcam), rabbit anti-CD44 (1:2,000; ab51037; Abcam) and
rabbit anti-SerpinB3 (1:5,000; ab126752; Abcam). Then the membranes
were washed 3 times in TBST, and incubated with horseradish
peroxidase-conjugated anti-mouse (P0260) or anti-rabbit (P0217)
antibodies secondary antibody (Dako; Agilent Technologies, Inc.,
Santa Clara, CA, USA) at room temperature for 1 h. Following
washing 3 times with TBST, the reaction was detected by
chemiluminescence with the WesternBright ECL detection kit
(Advansta Inc., Menlo Park, CA, USA). Darkroom development
techniques were employed to expose and acquire images of the
membranes.
Statistical analysis
The receiver operating characteristic (ROC) curve
and area under the curve (AUC) were used to determine the
performance of the five individual potential biomarkers by
employing GraphPad Prism software version 7 (GraphPad Software,
Inc., La Jolla, CA, USA). The combination of the five potential
biomarkers was also analyzed using logistic regression with SPSS
software version 11 (IBM Corp., Armonk, NY, USA).
Results
Analysis of the altered urinary
proteins expression
The body filtrates from urine such as water,
electrolytes, salts, nitrogenous waste and proteins are derived
from the plasma, kidneys and urogenital tract. The amount of
urinary protein excretion normally is <150 mg/day and needs to
be concentrated for use in experiments (21). Label-free quantitative proteomics is
the best choice for analyzing samples with a low level of proteins.
The pooled urinary samples from 5 normal individuals (N9-N13) and 6
cervical cancer patients (C19-C24) were analyzed by LC-MS/MS, as
shown in Table IIA and B.
Significant changes were calculated using Progenesis label-free
LC-MS software version 3.1. A list of proteins with 2 or more
matched peptides was compiled. A total 133 proteins with 1% FDR
were identified. A total of 60 upregulated and 73 downregulated
urinary proteins were expressed when comparing normal and cervical
cancer samples. Serotransferrin (TF) was observed to be the most
significantly increased protein (2.91-fold, P=0.000004) while
Kininogen-1 (KNG1) was the most significantly decreased protein
(1.73-fold, P=0.000003). The results also revealed that LRG1,
low-density lipoprotein receptor-related protein 2 (LRP2), Myosin
light chain kinase (MYLK), SPARC-like 1, Zinc finger CCHC
domain-containing protein 8, MMRN1 and nectin-2 were notable
proteins that had >2 fold expression while aminopeptidase N had
downregulated protein expression (5.31 fold) and another 10
proteins had between 3.0–4.33 fold altered expression including
protein S100A8, trefoil factor 2, apolipoprotein E (APOE),
apolipoprotein D (APOD), DNA (cytosine-5)-methyltransferase 3B
(DNMT3B), CD44, SERPINB3, macrophage colony-stimulating factor 1
(CSF1), epidymal secretory protein E1 and ribonuclease pancreatic.
The large molecular weight proteins of >100 kDa were obtained
using this method, including LRP2 (521.6 kDa), MYLK (210.7 kDa) and
MMRN1 (138 kDa). These proteins of interest (LRG1, MMRN1, S100A8,
SERPINB3 and CD44) were selected to confirm their expression by
western blot analysis.
| Table II.List of differentially expressed
urinary proteins in normal individuals and patients with cervical
cancer, identified by relative quantitative label-free mass
spectrometry analysis. |
Table II.
List of differentially expressed
urinary proteins in normal individuals and patients with cervical
cancer, identified by relative quantitative label-free mass
spectrometry analysis.
| A, Upregulated
urinary proteins in patients with cervical cancer with ANOVA P≤0.05
and Max fold change ≥1.5 |
|---|
| No. | Gene name | Accession | Description | Peptide count | Score | ANOVA P-value | Fold-change | MW(kDa)/pI |
|---|
| 1 | TF | TRFE_HUMAN |
Serotransferrin | 15 | 2,566.45 | 0.000004 | 2.91 | 77.06/6.81 |
| 2 | ORM1 | A1AG1_HUMAN | α-1-acid
glycoprotein 1 | 4 | 656.38 | 0.000005 | 2.39 | 23.72/4.93 |
| 3 | LRG1 | A2GL_HUMAN | Leucine-rich
α-2-glycoprotein | 3 | 456.41 | 0.000007 | 2.72 | 38.38/6.45 |
| 4 | LRP2 | LRP2_HUMAN | Low-density
lipoprotein receptor-related protein 2 | 17 | 1,119.49 | 0.000009 | 4.34 | 521.59/4.89 |
| 5 | MYLK | MYLK_HUMAN | Myosin light chain
kinase, smooth muscle | 7 | 288.88 | 0.000013 | 3.28 | 210.71/5.85 |
| 6 | ORM2 | A1AG2_HUMAN | α-1-acid
glycoprotein | 4 | 584.86 | 0.00003 | 2.64 | 23.87/5.03 |
| 7 | PDCD11 | RRP5_HUMAN | Protein RRP5
homolog | 8 | 272.23 | 0.000031 | 2.48 | 208.70/8.99 |
| 8 | APOA1 | APOA1_HUMAN | Apolipoprotein
A-I | 2 | 436.83 | 0.000036 | 2.99 | 30.77/5.56 |
| 9 | FAM109B | SESQ2_HUMAN |
Sesquipedalian-2 | 4 | 35.46 | 0.000048 | 1.8 | 28.33/6.66 |
| 10 | ALB | ALBU_HUMAN | Serum albumin | 30 | 4,048.64 | 0.000063 | 1.98 | 71.31/5.92 |
| 11 | CPM | CBPM_HUMAN | Carboxypeptidase
M | 2 | 111.23 | 0.000337 | 1.95 | 50.93/6.94 |
| 12 | A1BG | A1BG_HUMAN |
α-1B-glycoprotein | 4 | 797.85 | 0.000407 | 2.04 | 54.79/5.56 |
| 13 | SPARCL1 | SPRL1_HUMAN | SPARC-like protein
1 | 9 | 136.75 | 0.000421 | 3.28 | 75.20/4.71 |
| 14 | LYST | LYST_HUMAN |
Lysosomal-trafficking regulator | 12 | 440.97 | 0.000459 | 1.78 | 429.13/6.15 |
| 15 | ZCCHC8 | ZCHC8_HUMAN | Zinc finger CCHC
domain-containing protein 8 | 2 | 50.3 | 0.000489 | 4.78 | 78.57/4.79 |
| 16 | DNAH10 | DYH10_HUMAN | Dynein heavy chain
10, axonemal | 21 | 610.61 | 0.00054 | 1.73 | 514.84/5.64 |
| 17 | CHD6 | CHD6_HUMAN |
Chromodomain-helicase-DNA-binding protein
6 | 14 | 341.78 | 0.000555 | 1.87 | 305.41/5.90 |
| 18 | SYNE1 | SYNE1_HUMAN | Nesprin-1 | 42 | 1,118.05 | 0.000586 | 1.87 | 1011.08/5.37 |
| 19 | HPX | HEMO_HUMAN | Hemopexin | 6 | 727.65 | 0.00061 | 2.29 | 51.67/6.55 |
| 20 | WDFY2 | WDFY2_HUMAN | WD repeat and FYVE
domain-containing protein 2 | 6 | 97.64 | 0.000612 | 2.94 | 45.15/6.46 |
| 21 | SOX6 | SOX6_HUMAN | Transcription
factor SOX-6 | 6 | 137.57 | 0.000684 | 2.14 | 91.92/7.65 |
| 22 | CHGB | SCG1_HUMAN |
Secretogranin-1 | 2 | 73.98 | 0.000973 | 2.99 | 78.27/5.02 |
| 23 | CPE | CBPE_HUMAN | Carboxypeptidase
E | 4 | 37.93 | 0.00107 | 2.19 | 53.15/5.03 |
| 24 | MAP3K7 | M3K7_HUMAN | Mitogen-activated
protein kinase kinase kinase 7 | 8 | 168.58 | 0.00111 | 2.19 | 67.19/6.69 |
| 25 | DCLK1 | DCLK1_HUMAN |
Serine/threonine-protein kinase DCLK1 | 6 | 137.52 | 0.00125 | 1.68 | 82.22/8.84 |
| 26 | GC | VTDB_HUMAN | Vitamin D-binding
protein | 11 | 703.57 | 0.0013 | 1.59 | 54.52/5.40 |
| 27 | KRT17 | K1C17_HUMAN | Keratin, type I
cytoskeletal 17 | 5 | 123.3 | 0.00131 | 2.13 | 48.10/4.97 |
| 28 | – | KV105_HUMAN | Immunoglobulin κ
variable 1–5 | 2 | 82.6 | 0.00149 | 1.74 | 12.78/8.49 |
| 29 | MYO18B | MY18B_HUMAN | Unconventional
myosin-XVIIIb | 21 | 644.74 | 0.00168 | 1.66 | 285.18/6.49 |
| 30 | MACF1 | MACF1_HUMAN | Microtubule-actin
cross-linking factor 1, isoforms 1/2/3/5 | 32 | 996.53 | 0.00173 | 1.82 | 838.30/5.28 |
| 31 | FGG | FIBG_HUMAN | Fibrinogen γ
chain | 3 | 373.78 | 0.00215 | 1.83 | 51.51/5.37 |
| 32 | C3 | CO3_HUMAN | Complement C3 | 10 | 740.32 | 0.00228 | 1.73 | 187.14/6.02 |
| 33 | CP | CERU_HUMAN | Ceruloplasmin | 8 | 1,240.04 | 0.00273 | 1.54 | 122.98/5.44 |
| 34 | AZGP1 | ZA2G_HUMAN |
Zinc-α-2-glycoprotein | 5 | 1,217.5 | 0.00316 | 1.55 | 34.46/5.71 |
| 35 | FGB | FIBB_HUMAN | Fibrinogen
β-chain | 2 | 172.98 | 0.00317 | 1.75 | 55.92/8.54 |
| 36 | PVRL2 | NECT2_HUMAN | Nectin-2 | 3 | 95.27 | 0.00352 | 3.37 | 55.74/4.74 |
| 37 | IL16 | IL16_HUMAN |
Pro-interleukin-16 | 11 | 208.63 | 0.00371 | 1.82 | 141.75/8.34 |
| 38 | UCMA | UCMA_HUMAN | Unique cartilage
matrix-associated protein | 2 | 66.89 | 0.00441 | 1.78 | 16.56/5.47 |
| 39 | PGK2 | PGK2_HUMAN | Phosphoglycerate
kinase 2 | 3 | 92.54 | 0.00489 | 1.95 | 44.79/8.74 |
| 40 | H2BFWT | H2BWT_HUMAN | Histone H2B type
W-T | 3 | 44.94 | 0.00489 | 1.72 | 19.61/10.69 |
| 41 | HP | HPT_HUMAN | Haptoglobin | 3 | 1,055.7 | 0.0061 | 1.72 | 45.20/6.13 |
| 42 | APOA4 | APOA4_HUMAN | Apolipoprotein
A-IV | 6 | 70.31 | 0.00638 | 1.64 | 45.39/5.28 |
| 43 | – | IGHG3_HUMAN | Ig γ-3 chain C
region | 2 | 584.82 | 0.00751 | 1.56 | 41.28/8.23 |
| 44 | R3HCC1L | R3HCL_HUMAN | Coiled-coil
domain-containing protein R3HCC1L | 3 | 156.15 | 0.00789 | 1.53 | 87.88/5.01 |
| 45 | FN1 | FINC_HUMAN | Fibronectin | 12 | 647.44 | 0.00855 | 2 | 262.62/5.46 |
| 46 | HEXA | HEXA_HUMAN | β-hexosaminidase
subunit alpha | 2 | 72.64 | 0.00857 | 1.59 | 61.12/5.04 |
| 47 | AKAP4 | AKAP4_HUMAN | A-kinase anchor
protein 4 | 4 | 151.22 | 0.00885 | 1.76 | 94.47/6.56 |
| 48 | PREX1 | PREX1_HUMAN |
Phosphatidylinositol
3,4,5-trisphosphate-dependent Rac exchanger 1 protein | 8 | 231.71 | 0.02 | 2.19 | 186.20/6.03 |
| 49 | RPS27A | RS27A_HUMAN | Ubiquitin-40S
ribosomal protein S27a | 3 | 63.39 | 0.02 | 1.93 | 17.96/9.68 |
| 50 | AMY2A | AMYP_HUMAN | Pancreatic
α-amylase | 3 | 1,106.07 | 0.02 | 1.89 | 57.70/6.60 |
| 51 | – | KV320_HUMAN | Immunoglobulin κ
variable 3–20 | 2 | 200.67 | 0.02 | 1.77 | 12.55/4.85 |
| 52 | SERPINA7 | THBG_HUMAN | Thyroxine-binding
globulin | 3 | 243.84 | 0.02 | 1.65 | 46.32/5.87 |
| 53 | DNAH3 | DYH3_HUMAN | Dynein heavy chain
3, axonemal | 23 | 639.86 | 0.02 | 1.61 | 470.77/6.04 |
| 54 | MMRN2 | MMRN2_HUMAN | Multimerin-2 | 3 | 79.01 | 0.02 | 1.54 | 104.40/5.56 |
| 55 | AFM | AFAM_HUMAN | Afamin | 6 | 335.44 | 0.03 | 1.65 | 69.06/5.64 |
| 56 | FAM208B | F208B_HUMAN | Protein
FAM208B | 18 | 427.39 | 0.03 | 1.65 | 268.84/5.61 |
| 57 | PTPRS | PTPRS_HUMAN | Receptor-type
tyrosine-protein phosphatase S | 14 | 335.72 | 0.03 | 1.53 | 217.04/6.06 |
| 58 | MMRN1 | MMRN1_HUMAN | Multimerin-1 | 3 | 225.55 | 0.04 | 2.01 | 138.11/8.15 |
| 59 | COL4A2 | CO4A2_HUMAN | Collagen α-2(IV)
chain | 8 | 171.58 | 0.05 | 1.71 | 167.55/8.89 |
| 60 | PLAU | UROK_HUMAN | Urokinase-type
plasminogen activator | 2 | 116.86 | 0.05 | 1.5 | 48.50/8.78 |
|
| B, Downregulated
urinary proteins in patients with cervical cancer with ANOVA P≤0.05
and Max fold change ≥1.5 |
|
| No. | Gene
name |
Accession |
Description | Peptide
count | Score | ANOVA
P-value |
Fold-change |
MW(kDa)/pI |
|
| 1 | KNG1 | KNG1_HUMAN | Kininogen-1 | 13 | 1,338.67 | 0.000003 | 1.73 | n/a |
| 2 | S100A8 | S10A8_HUMAN | Protein
S100-A8 | 2 | 378.86 | 0.000003 | 3.42 | n/a |
| 3 | ANPEP | AMPN_HUMAN | Aminopeptidase
N | 6 | 403.62 | 0.000005 | 5.31 | n/a |
| 4 | APOE | APOE_HUMAN | Apolipoprotein
E | 4 | 61.1 | 0.000027 | 3.04 | n/a |
| 5 | SPP1 | OSTP_HUMAN | Osteopontin | 4 | 801.94 | 0.000047 | 2.81 | n/a |
| 6 | LAMP1 | LAMP1_HUMAN | Lysosome-associated
membrane glycoprotein 1 | 4 | 209.2 | 0.00005 | 2.1 | n/a |
| 7 | APOD | APOD_HUMAN | Apolipoprotein
D | 2 | 373.48 | 0.000052 | 3.62 | n/a |
| 8 | LPHN1 | AGRL1_HUMAN | Adhesion G
protein-coupled receptor L1 | 11 | 164.47 | 0.000056 | 1.69 | n/a |
| 9 | UMOD | UROM_HUMAN | Uromodulin | 11 | 1,580 | 0.000057 | 2.1 | n/a |
| 10 | DNMT3B | DNM3B_HUMAN | DNA
(cytosine-5)-methyltransferase 3B | 2 | 67.24 | 0.000069 | 3.41 | n/a |
| 11 | TFF2 | TFF2_HUMAN | Trefoil factor
2 | 2 | 86.69 | 0.000074 | 4.33 | n/a |
| 12 | ACTBL2 | ACTBL_HUMAN | β-actin-like
protein 2 | 5 | 188.69 | 0.000093 | 1.77 | n/a |
| 13 | CTSD | CATD_HUMAN | Cathepsin D | 7 | 403.41 | 0.000193 | 2.09 | n/a |
| 14 | GAA | LYAG_HUMAN | Lysosomal
α-glucosidase | 7 | 1,018.11 | 0.00022 | 2.33 | n/a |
| 15 | TNC | TENA_HUMAN | Tenascin | 6 | 271.43 | 0.000327 | 1.85 | n/a |
| 16 | CD44 | CD44_HUMAN | CD44 antigen | 4 | 483.38 | 0.000369 | 3.12 | n/a |
| 17 | CSF1 | CSF1_HUMAN | Macrophage
colony-stimulating factor 1 | 2 | 198.48 | 0.000435 | 3.55 | n/a |
| 18 | CD248 | CD248_HUMAN | Endosialin | 5 | 234.21 | 0.000596 | 2.6 | n/a |
| 19 | ARSA | ARSA_HUMAN | Arylsulfatase
A | 4 | 511.64 | 0.000624 | 2.91 | n/a |
| 20 | CTSB | CATB_HUMAN | Cathepsin B | 2 | 264.96 | 0.000639 | 2.14 | n/a |
| 21 | ANK3 | ANK3_HUMAN | Ankyrin-3 | 49 | 1,009.24 | 0.000851 | 1.7 | n/a |
| 22 | PSAP | SAP_HUMAN | Prosaposin | 3 | 361.86 | 0.000852 | 2.17 | n/a |
| 23 | – | LV321_HUMAN | Immunoglobulin
lambda variable 3–21 | 2 | 154.16 | 0.000969 | 2.13 | n/a |
| 24 | BHMT | BHMT1_HUMAN |
Betaine--homocysteine S-methyltransferase
1 | 7 | 289.37 | 0.001 | 1.94 | n/a |
| 25 | COL15A1 | COFA1_HUMAN | Collagen α-1(XV)
chain | 2 | 179.35 | 0.00104 | 2.04 | n/a |
| 26 | IGLL5 | IGLL5_HUMAN | Immunoglobulin
lambda-like polypeptide 5 | 2 | 385.74 | 0.00107 | 1.59 | n/a |
| 27 | CDH13 | CAD13_HUMAN | Cadherin-13 | 4 | 369.61 | 0.00117 | 1.67 | n/a |
| 28 | PTGDS | PTGDS_HUMAN | Prostaglandin-H2
D-isomerase | 5 | 333.35 | 0.00121 | 1.91 | n/a |
| 29 | COL6A1 | CO6A1_HUMAN | Collagen α-1(VI)
chain | 7 | 715.12 | 0.00138 | 2.01 | n/a |
| 30 | FOLR1 | FOLR1_HUMAN | Folate receptor
α | 5 | 190.49 | 0.00142 | 1.81 | n/a |
| 31 | CD55 | DAF_HUMAN | Complement
decay-accelerating factor | 3 | 500.96 | 0.00148 | 1.82 | n/a |
| 32 | NPC2 | NPC2_HUMAN | Epididymal
secretory protein E1 | 2 | 209.65 | 0.00151 | 3.11 | n/a |
| 33 | PROCR | EPCR_HUMAN | Endothelial protein
C receptor | 3 | 291 | 0.00156 | 1.74 | n/a |
| 34 | AHSG | FETUA_HUMAN |
Alpha-2-HS-glycoprotein | 5 | 229.87 | 0.00162 | 1.51 | n/a |
| 35 | ABCB5 | ABCB5_HUMAN | ATP-binding
cassette sub-family B member 5 | 7 | 217.25 | 0.00162 | 2.16 | n/a |
| 36 | CADM4 | CADM4_HUMAN | Cell adhesion
molecule 4 | 5 | 454.21 | 0.00174 | 1.63 | n/a |
| 37 | RNASE1 | RNAS1_HUMAN | Ribonuclease
pancreatic | 4 | 267.25 | 0.00177 | 3.15 | n/a |
| 38 | WFDC2 | WFDC2_HUMAN | WAP four-disulfide
core domain protein 2 | 2 | 237.59 | 0.00215 | 2.21 | n/a |
| 39 | F5 | FA5_HUMAN | Coagulation factor
V | 10 | 340.57 | 0.0023 | 1.65 | n/a |
| 40 | PRNP | PRIO_HUMAN | Major prion
protein | 4 | 159.7 | 0.00244 | 2.18 | n/a |
| 41 | RETN | RETN_HUMAN | Resistin | 2 | 175.19 | 0.00254 | 1.68 | n/a |
| 42 | KLHL4 | KLHL4_HUMAN | Kelch-like protein
4 | 3 | 97.09 | 0.00335 | 2.16 | n/a |
| 43 | SERPING1 | IC1_HUMAN | Plasma protease C1
inhibitor | 6 | 567.51 | 0.00389 | 1.6 | n/a |
| 44 | PARP14 | PAR14_HUMAN | Poly [ADP-ribose]
polymerase 14 | 4 | 210.84 | 0.00415 | 1.68 | n/a |
| 45 | CRYL1 | CRYL1_HUMAN | Lambda-crystallin
homolog | 4 | 84.49 | 0.00432 | 1.6 | n/a |
| 46 | C16orf62 | CP062_HUMAN | UPF0505 protein
C16orf62 | 4 | 94.21 | 0.00456 | 1.89 | n/a |
| 47 | GALNS | GALNS_HUMAN |
N-acetylgalactosamine-6-sulfatase | 4 | 121.68 | 0.00543 | 1.67 | n/a |
| 48 | SERPINA4 | KAIN_HUMAN | Kallistatin | 8 | 133.52 | 0.00559 | 1.71 | n/a |
| 49 | OSCAR | OSCAR_HUMAN |
Osteoclast-associated immunoglobulin-like
receptor | 3 | 189.22 | 0.00559 | 2.14 | n/a |
| 50 | AMBP | AMBP_HUMAN | Protein AMBP | 8 | 1,211.69 | 0.00628 | 1.76 | n/a |
| 51 | CPQ | CBPQ_HUMAN | Carboxypeptidase
Q | 6 | 355.87 | 0.00656 | 1.64 | n/a |
| 52 | ANGPTL2 | ANGL2_HUMAN |
Angiopoietin-related protein 2 | 5 | 195.93 | 0.00678 | 1.55 | n/a |
| 53 | LAIR1 | LAIR1_HUMAN |
Leukocyte-associated immunoglobulin-like
receptor 1 | 4 | 302.08 | 0.00754 | 1.66 | n/a |
| 54 | PIP | PIP_HUMAN | Prolactin-inducible
protein | 2 | 286.9 | 0.00785 | 1.52 | n/a |
| 55 | SPRR3 | SPRR3_HUMAN | Small proline-rich
protein 3 | 5 | 263.95 | 0.00984 | 3.37 | n/a |
| 56 | MXRA8 | MXRA8_HUMAN |
Matrix-remodeling-associated protein
8 | 2 | 188.15 | 0.00991 | 1.6 | n/a |
| 57 | WDFY3 | WDFY3_HUMAN | WD repeat and FYVE
domain-containing protein 3 | 15 | 358.34 | 0.01 | 1.51 | n/a |
| 58 | CDH2 | CADH2_HUMAN | Cadherin-2 | 2 | 329.32 | 0.01 | 1.6 | n/a |
| 59 | ACP2 | PPAL_HUMAN | Lysosomal acid
phosphatase | 6 | 380.73 | 0.01 | 1.62 | n/a |
| 60 | ATG2A | ATG2A_HUMAN | Autophagy-related
protein 2 homolog A | 5 | 91.97 | 0.01 | 1.85 | n/a |
| 61 | CRNN | CRNN_HUMAN | Cornulin | 2 | 277.69 | 0.01 | 2.04 | n/a |
| 62 | MT1G | MT1G_HUMAN |
Metallothionein-1G | 2 | 111.25 | 0.01 | 2.06 | n/a |
| 63 | ESAM | ESAM_HUMAN | Endothelial
cell-selective adhesion molecule | 5 | 79.85 | 0.01 | 2.14 | n/a |
| 64 | TNRC18 | TNC18_HUMAN | Trinucleotide
repeat-containing gene 18 protein | 9 | 266.88 | 0.02 | 1.51 | n/a |
| 65 | APP | A4_HUMAN | Amyloid β A4
protein | 4 | 227.47 | 0.02 | 1.57 | n/a |
| 66 | SLC3A2 | 4F2_HUMAN | 4F2 cell-surface
antigen heavy chain | 4 | 151.92 | 0.02 | 1.58 | n/a |
| 67 | TTR | TTHY_HUMAN | Transthyretin | 4 | 212.41 | 0.02 | 1.64 | n/a |
| 68 | VPS18 | VPS18_HUMAN | Vacuolar protein
sorting-associated protein 18 homolog | 3 | 87.62 | 0.03 | 1.54 | n/a |
| 69 | S100P | S100P_HUMAN | Protein S100-P | 3 | 103.16 | 0.03 | 1.63 | n/a |
| 70 | ZNF805 | ZN805_HUMAN | Zinc finger protein
805 | 3 | 82.25 | 0.03 | 2.83 | n/a |
| 71 | LYVE1 | LYVE1_HUMAN | Lymphatic vessel
endothelial hyaluronic acid receptor 1 | 4 | 396.4 | 0.04 | 1.52 | n/a |
| 72 | RNF113A | R113A_HUMAN | RING finger protein
113A | 4 | 126.64 | 0.04 | 1.82 | n/a |
| 73 | SERPINB3 | SPB3_HUMAN | Serpin B3 | 3 | 108 | 0.04 | 1.9 | n/a |
Characterization, classification,
functional and protein- protein interactions analysis
Venn diagram comparing protein expression from
cervical cancer between identified proteins by LC-MS/MS and the
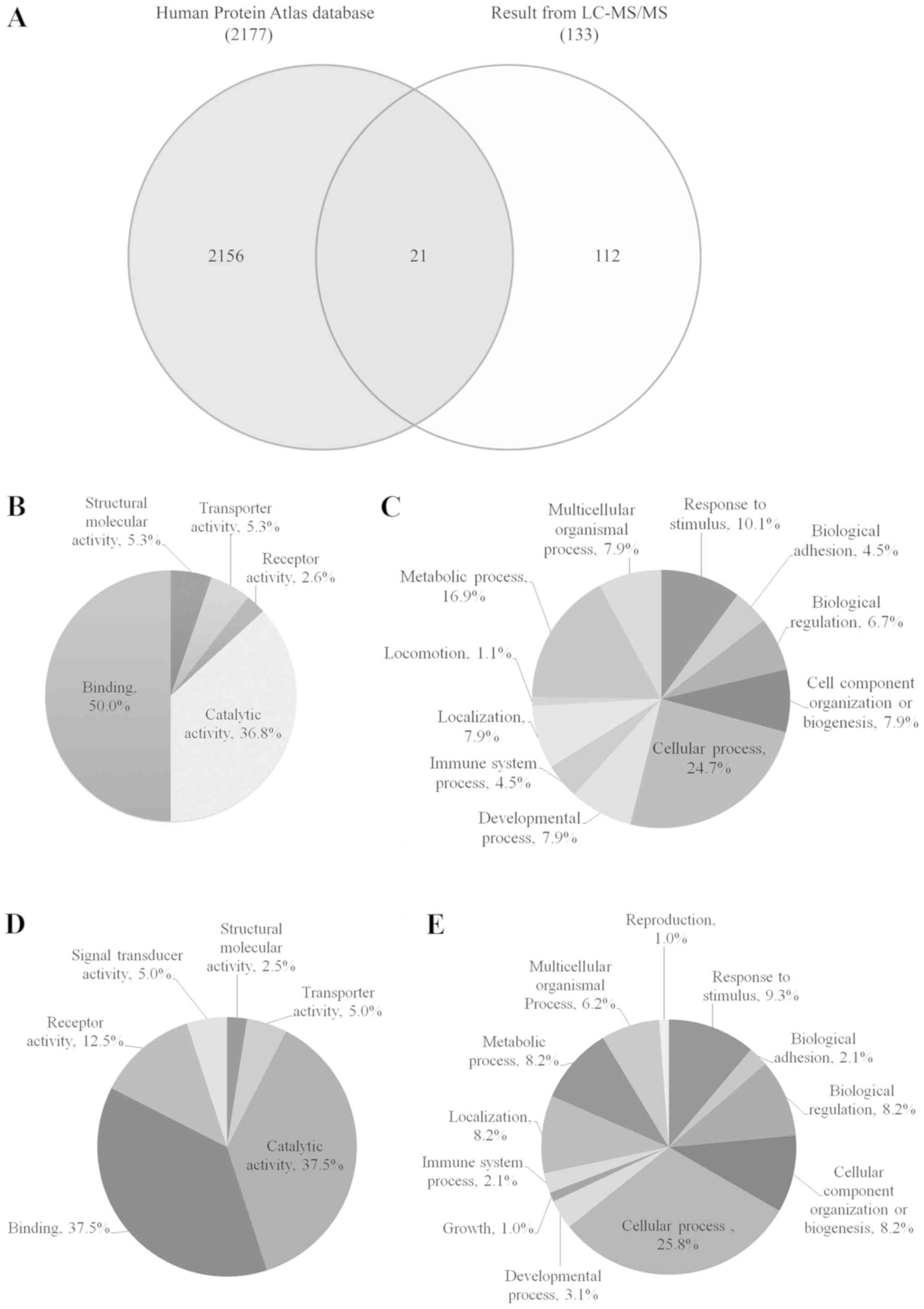
database of Human Protein Atlas was shown in Fig. 1A. Twenty-one identified proteins were
overlapped with The Human Protein Atlas Database, including ABCB5,
AMY2A, ANGPTL2, C3, CD44, CD55, COL6A1, CRNN, CRYL1, DNAH3 ESAM,
GALNS, HEXA, KRT17, MACF1, MXRA8, PGK2, PIP, PTGDS, SERPINB3 and
WFDC2. Interestingly, 112 identified proteins have never been
reported in the database of Human Protein Atlas for cervical
cancer.
The up- and downregulated urinary proteins from
Table IIA and B were analyzed by GO
with the PANTHER database system to obtain ‘Molecular function’ and
‘Biological process’ data, as presented in pie charts in Fig. 1B-E. The molecular functions of 60
upregulated proteins from Fig. 1B
were classified into 5 groups: Binding (50%), catalytic activity
(36.8%), structural molecular activity (5.3%), transporter activity
(5.3%) and receptor activity (2.6%), while the biological processes
of these proteins (Fig. 1C) were
classified into 11 groups with the highest percentage observed for
cellular process (24.7%), followed by metabolic process (16.9%) and
response to stimulus (10.1%). The molecular functions of the 73
downregulated proteins, as presented in Fig. 1D, were classified into 6 groups and
were mainly involved in binding (37.5%) and catalytic activity
(37.5%), while the biological processes of these proteins (Fig. 1E) were classified into 12 groups and
were mainly involved in cellular process (25.8%) and response to
stimulus (9.3%).
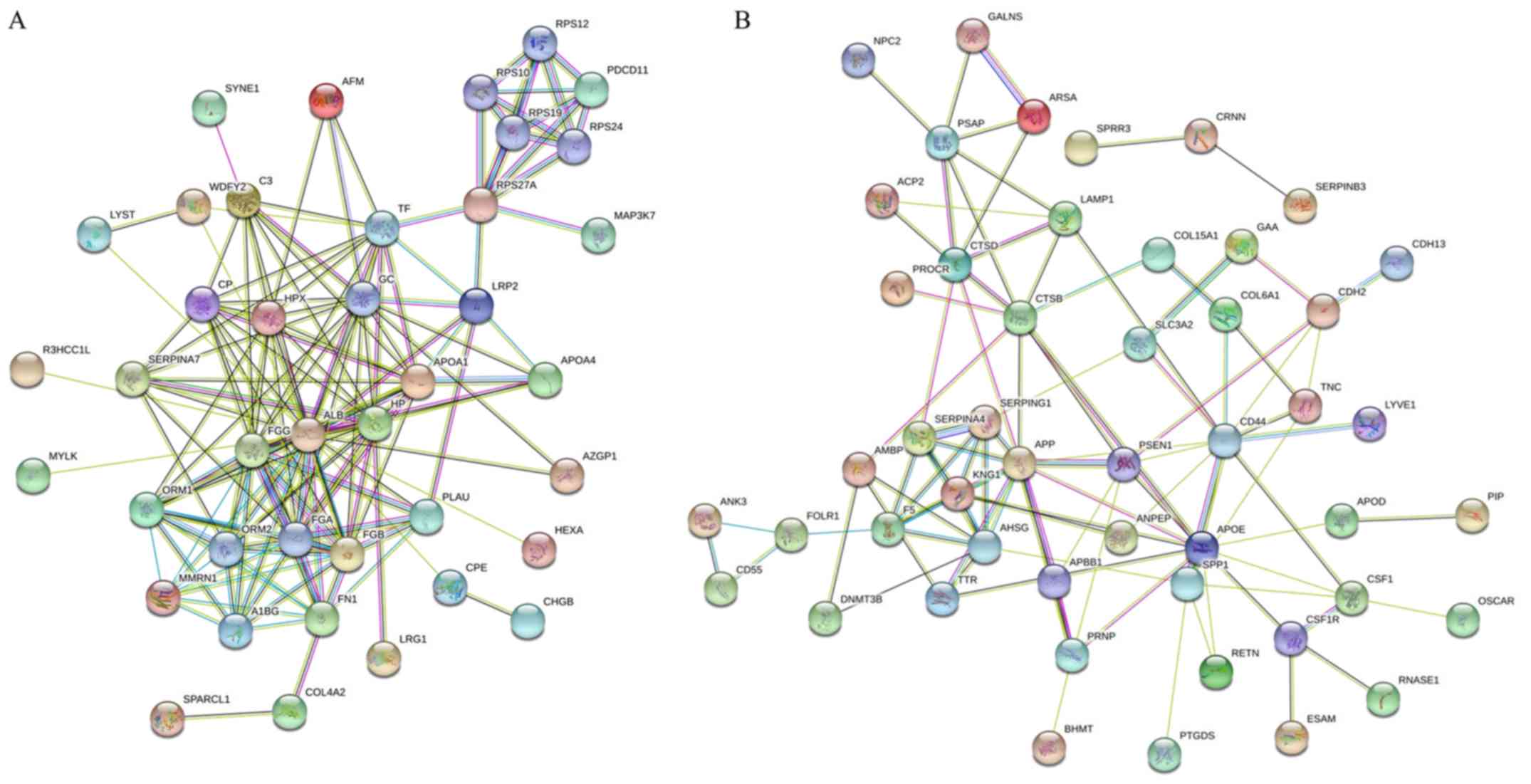
The STRING database was used for analysis of the
protein-protein interactions and was able to predict the similar
functions of proteins by accessing various free resources. The
‘molecular function’, ‘cellular component’ and ‘biological process’
from GO and KEGG pathways enrichment analyses were obtained. The
present study analyzed the 60 and 73 identified proteins that were
up- and downregulated, respectively (Table II), when the pooled urinary samples
of normal individuals and patients with cervical cancer were
compared. The results for the interaction map of the upregulated
proteins was presented in Fig. 2A.
The results for the associations between these proteins revealed
that 40 of the 60 proteins had good interactions. The big cluster
was involved in protein binding functions from molecular function
(GO), metabolic process, blood coagulation, cell-cell adhesion,
regulation of cell morphogenesis and cell motility from biological
process (GO). A total of 19 proteins were involved in binding
including, albumin (ALB), fibrinogen β-chain (FGB), fibrinogen
γ-chain (FGG), fibronectin (FN1), haptoglobin, nesprin-1,
zinc-α-2-glycoprotein, vitamin D-binding protein, TF, LRP2, protein
RRP5 homolog and mitogen-activated protein kinase kinase kinase 7
(MAP3K7) while 12 proteins were involved in blood coagulation, such
as fibrinolysis, and fibrin clot formation including, ALB, FGG,
FGB, FN1, TF, MMRN1, apolipoprotein A, urokinase-type plasminogen
activator, complement C3, ceruloplasmin and lysosomal-trafficking
regulator. There were some proteins of interest that were involved
in multiple functions: For example, MAP3K7 was not only involved in
binding functions but was also involved in cell-cell adhesion and
metabolic process; and microtubule-actin cross-linking factor 1 was
involved in metabolic processes, the regulation of cell
morphogenesis for differentiation and the regulation of anatomical
structure morphogenesis. MMRN1 was also involved in the stress
response and cell adhesion function.
Fig. 2B presents the
STRING analysis of 73 downregulated proteins from the same
comparison. The results revealed that 49 of the 73 proteins had
good interactions. The big cluster was involved in different
protein functions such as stress-response, cell adhesion, leukocyte
migration, wounding response and extracellular matrix organization
from biological process (GO). A total of 20 proteins were involved
in the stress response with fold change range of 2 to 3.62
including DNMT3B (3.41-fold), APOD (3.62-fold), CSF1 (3.55-fold)
and APOE (3.04-fold). Some proteins were involved in multiple
functions: For example, CD44 and S100A8 were involved in all
functions associated with biological process while MMRN1 were
involved in only the stress response and cell adhesion
function.
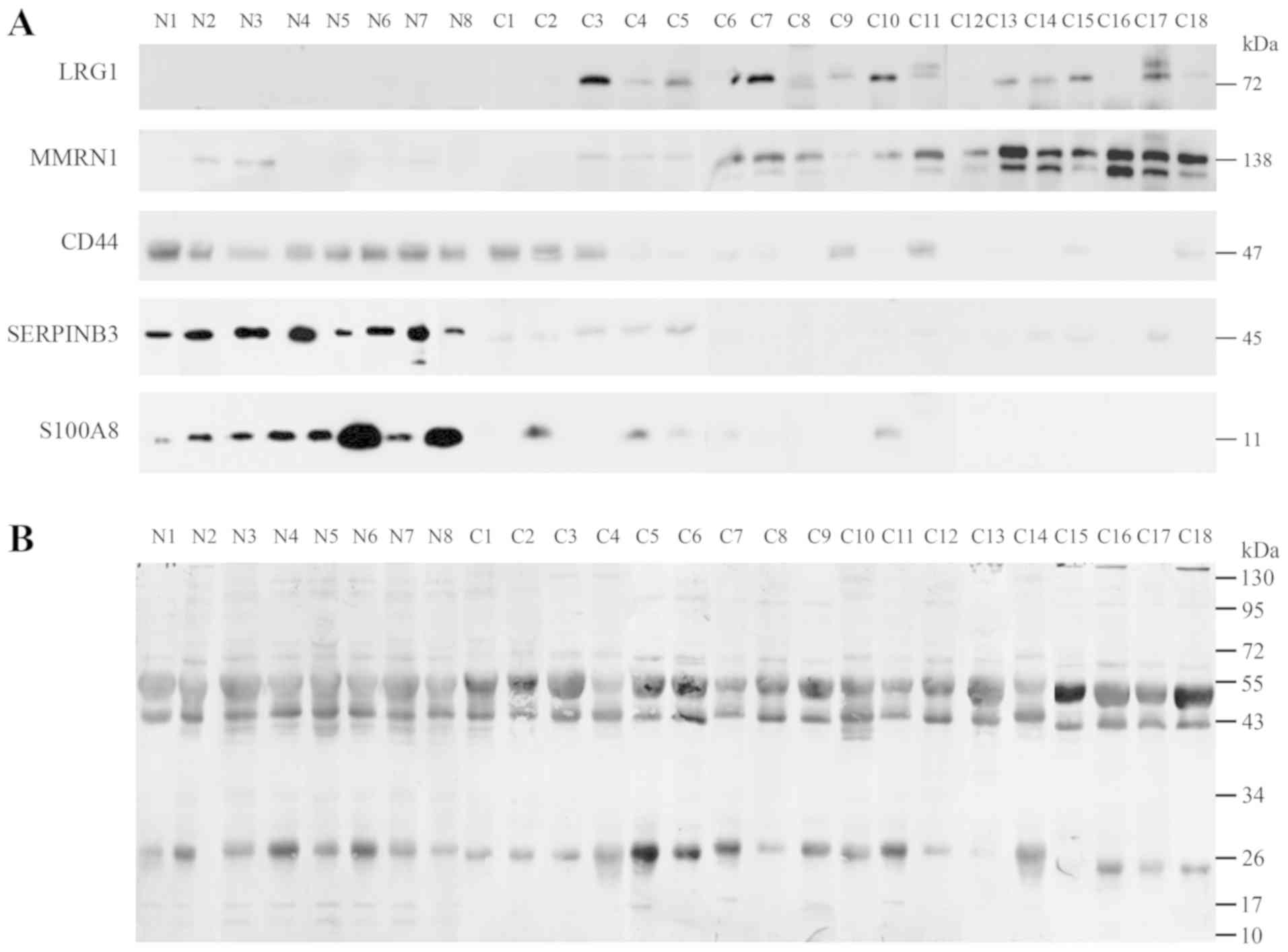
Validation of the identified potential
urine markers
Notable proteins were selected to confirm the
association with the urine of normal individuals and patients with
different stages of cervical cancer including 8 normal (N1-N8), 2
samples of stage IB1 (C1-C2), 9 samples of stage II (C3-C11), 4
samples of stage IB1 (C12-C15) and 3 samples of stage IVB
(C16-C18), as determined by western blot analysis. The expression
of LRG1, MMRN1, S100A8, SERPINB3 and CD44 were confirmed as shown
in Fig. 3. LRG1 and MMRN1 were
upregulated in stages IIB, IIIB and IVB of cervical cancer while
S100A8, SERPINB3 and CD44 had reduced expression in stage I and in
some stage II cancer samples. Notably, CD44 was highly expressed in
normal and stage IB1, and had low expression in stages II, III and
IV.
Sensitivity of potential
biomarkers
The results from immunodetection by western blot
analysis revealed that LRG1, MMRN1, CD44, S100A8 and SERPINB3 could
be potential biomarkers for cervical cancer. All five proteins were
tested individually and in combination for sensitivity and
specificity using logistic regression on SPSS software and ROC
curves, respectively. The ROC curves were plotted for these
proteins to discriminate between cervical cancer (all stages) and
control urine samples (Fig. 4). The
results showed the AUC of LRG1, MMRN1, CD44, S100A8 and SERPINB3
were 0.993, 0.785, 0.938, 0.931 and 0.986, respectively. LRG1 and
SERPINB3 had the highest AUC among all 5 proteins. The combination
of the 5 biomarker panel was calculated and the value of the AUC
was increased to 1 (100% sensitivity, 87.5% specificity), which was
better than LRG1 or SERPINB3 individually.
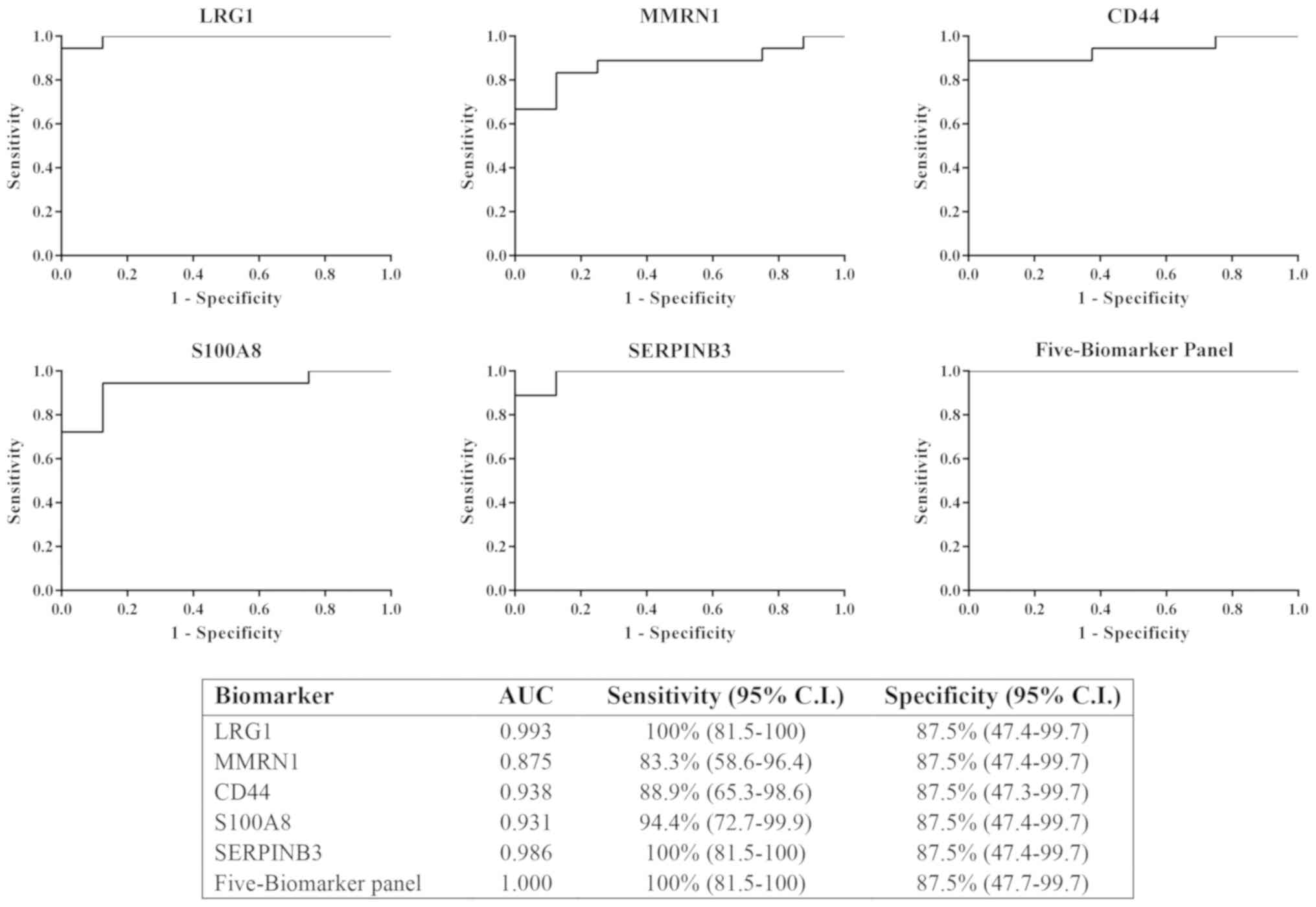 | Figure 4.ROC curve of the 5 individual
potential biomarkers (LRG1, MMRN1, CD44, S100A8 and SERPINB3) was
calculated to determine the area under the curve, sensitivity (95%
CI) and specificity (95% CI). The ROC for the 5-biomarker panel was
analyzed by combining the 5 potential biomarkers. ROC, receiver
operating characteristic; LRG1, leucine-rich α-2-glycoprotein;
MMRN1, multimerin-1; SERPINB3, serpin B3; CD44, cluster of
differentiation 44 antigen; CI, confidence interval; AUC, area
under the curve. |
Discussion
Proteomics is a useful technique to identify for
cancer biomarkers. Urinary proteomic analysis for biomarker
discovery of cervical cancer has been reported previously (15). There have also been reports in
cervical cancer cell lines of the deregulation of
cytoskeletal-associated proteins (22). Two-dimensional gel electrophoresis
followed by MS has been used for a decade and non-gel based-MS has
now become popular so label-free shotgun proteomics has been a good
choice to produce very effective results for the global profiles of
the samples. In the present study, quantitative label-free MS
analysis was employed to identify the differences in the expression
of urinary proteins in pooled cancer and normal samples. Progenesis
QI software was used to analyze the differential expression between
these pooled samples. The following criteria was used in this
experiment: i) An ANOVA score ≤0.05; ii) proteins with ≥2 matched
peptides; and iii) a Mascot score of ≥40. The present study
identified a total of 133 proteins (1% FDR).
Blood coagulation and fibrinolysis is one of the
main functions in the urinary proteins of cervical cancer. The
proteins of interest were coagulation factor V, FGB, FGG, KNG1,
MMRN1, LRP2 and MYLK. Following validation via western blot
analysis with 8 normal urine samples, (2 of stage I, 9 of stage II,
4 of stage III and 3 of stage IV separately), the proteins LRG1,
MMRN1, S100A8, SERPINB3 and CD44 were revealed to be potential
multiple biomarkers for cervical cancer. The upregulation of LRG1
and MMRN1 in stages II, III and IV and the downregulation of
S100-A8 and SERPINB3 in stages I, II, III and IV were of interest.
In addition, CD44 was highly expressed in normal samples and stage
IB1, and had reduced expression in stages II, III and IV. Following
the immunodetection results, ROC curve and logistic regression
analysis were employed to analyze all 5 potential biomarkers. The
results of the individual test for discriminatory ability using ROC
analysis revealed the highest AUC of 0.993 and 0.986 for LRG1 and
SERPINB3, respectively. When the panel of 5 biomarkers was
combined, the value of the AUC was increased from ~0.99 in 2
potentially good biomarkers (LRG1 and SERPINB3) to 1 with 100%
sensitivity and 87.5% specificity. On the other hand, combination
of the panel of 5 biomarkers improved the distinguished power
between two diagnostic groups (normal/cancer). Since an individual
biomarker is potentially a good diagnostic marker for cervical
cancer, the panel of 5 biomarkers is a better option for the
identification of cervical cancer patients.
To the best of our knowledge, MMRN1, for the first
time, was revealed to be highly expressed in urinary samples of
cervical cancer. This protein was also shown to be an isoform-1 of
MMRN1 due to the spectrum of peptides belonging to this isoform;
for example, KIENLTSAVNSLNFIIK (residue 677–693) and
NTDNIIYPEEYSSCSR (residue 1,032–1,047) belong to isoform-1. The
MMRN-1 precursor has a molecular weight of 135.75–136 Da. This
protein was reported to be a potential serum biomarker in multiple
myeloma as it decreased in patients with cancer (23), and was downregulated in non-small
cell lung cancer (24). MMRN1 was
also revealed to serve an important role in the storage and
stabilization of factor V in platelets and was reported to have a
function in the extracellular matrix or as an adhesive protein.
LRG1, which serves a role in cell survival, human
malignancies, proliferation and inhibits apoptosis, was upregulated
in various types of cancers including hepatocellular carcinoma,
pancreatic cancer and ovarian cancer (25–27). The
expression of LRG1 was 2.72-fold upregulated in the urine of
patients with cervical cancer when compared with normal samples; to
the best of our knowledge, this is the first time this protein has
been associated with this type of cancer. The immunodetection of
the expression of LRG1 was present only in the cervical cancer
stages II to IV.
The expressions of S100A8, CD44 and SERPINB3 were
downregulated when compared with the urine of normal and different
stages of cancer samples as determined by label free MS and
confirmation via western blotting.
Protein S100A8/A9 or calprotactin, a member of the
S100 family, is a heterogenous multimer of the subunit S100A8 and
S100A9. It is an EF-hand calcium-binding protein and has been shown
to be involved in cell cycle regulation, differentiation, invasion
and extracellular matrix cell adhesion (28,29).
This protein expression is upregulated in various types of cancers
by differential regulation in malignancies and is dependent on the
origin of cells and tissues. The adenocarcinomas were diagnosed via
the occurrence of tumorigenesis. Examples of these tissues include
prostate (30), gastric (31), thyroid (32), colorectal (33), ovarian (34), bladder (35) and pancreatic ductal adenocarcinoma
(36). By contrast, downregulated
expression of S100A8/A9 was observed in tissues of squamous cell
carcinomas of head and neck (HNSCC) (28), cervical (37), oral, oropharyngeal, esophageal and
nasopharyngeal (38,39). Loss of S100A8/A9 in tumors was
reported to be strongly associated with higher grades, poor
squamous differentiation and poorer survival of patients with HNSCC
(40). The present study
demonstrated no S100A8 expression in cancer patients, which was in
agreement with an earlier report (28,40,41).
CD44 was first observed on the surface of
granulocytes, T-lymphocytes and cortical thymocytes by Dalchau
et al (42). It is a protein
of the multifunctional family of the type I transmembrane and is
involved in the adhesion, migration and metastasis of cells. The
changes in CD44 expression serve a role in malignant tumors. A
previous study reported that the loss of CD44 expression predicted
higher advanced stages of oral squamous cell carcinoma (43). The reduction of CD44 expression was
reported to be involved in high invasiveness of tongue SCC and lead
to the metastasis of cervical lymph node (44). The expression of CD44 in the urine of
bladder cancer and urothelial carcinoma patients was also reported
previously and further studies were performed in the combination
with other proteins (45,46). The present results revealed that
there was a lower expression of CD44 in the urine samples of stages
II, III and IV cervical cancer patients but high expression was
observed in normal and stage I patients. Some studies have reported
CD44 as a biomarker for cervical cancer from tissue samples
(47) and they revealed that a lower
expression of CD44 was an indicator of high invasiveness of tumors
by increasing the rate of cervical cancer lymph node metastasis.
The mediation of CD44 on constitutive type I receptor signaling was
also observed in cervical cancer cells (48).
SERPINB3, a serine protease inhibitor member, has
been detected in many types of cells including leukocytes, normal
epithelium and tumors of epithelial origins for example epithelium
of cervix uterine, tonsils, esophagus and tongue. SERPINB3 was also
expressed in the amniotic fluid, saliva and respiratory secretions
of healthy people (49). SERPINB3
has been found in the serum of patients with many types of SCC
including HNSCC (50) and in
patients with psoriasis (51). It is
involved in the regulation of apoptosis via many mechanisms
(52). In Matrigel assays, when
cancer cell invasion was decreased, SERPINB3 was overexpressed
(53). The present results revealed
that SERPINB3 was overexpressed in all 9 normal urine samples but
had lower expression in stage I samples, and in 3 out of 9 stage II
samples and some of stage III, 2 out of 5 samples of stage III and
1 out of 3 samples of stage IV.
Our study aimed to demonstrate different urinary
protein profile between normal HPV negative and cervical cancer
patients, so we did not include preinvasive diseases as CIN1, CIN2
or CIN3. We evaluated the potential biomarkers (MMRN1, LRG1,
S100A8, SERPINB3 and CD44 using immunodetection with different
stages (stage I to stage IV) of cervical cancer. For further study,
we will collect all types of samples including precancerous state
and confirm whether these proteins could detect in early stage of
cervical cancer or not.
In conclusion, quantitative MS analysis could
identify the up- and downregulated proteins when comparing the
urine of normal individuals and cervical cancer patients. To the
best of our knowledge, MMRN1 and LRG1 were detected for the first
time in cervical cancer. MMRN1, LRG1, S100A8, SERPINB3 and CD44
were selected for validation by western blot analysis. The results
revealed that MMRN1 and LRG1 were upregulated, while S100A8,
SERPINB3 and CD44 were downregulated in cancer patients. ROC
analysis demonstrated that LRG1 and SERPINB3 could be used alone,
while these 5 proteins could also be combined to detect the
occurrence of cervical cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Chulabhorn
Research Institute (grant no. BC 2008-02) and Chulabhorn Royal
Academy (grant no. 31/2554).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DC and KWa performed the experiments including mass
spectrometry, progenesis analysis, western blot analysis and data
analysis, and helped with manuscript preparation. CS and CW
designed the study, developed the methodology and interpreted the
results. PS, SK and PDNA made substantial contributions to the
analysis and interpretation of data, wrote part of manuscript and
performed the literature search. KS, JC and CV performed the
statistical and bioinformatics analyses. DC, KWa, PS, CW, KS, JC
and CV prepared the figures. CS wrote the manuscript and prepared
the original draft. JS supervised the quality of the study design
and gave final approval of the manuscript prior to submission. NS
performed the clinical data collection and provided the HPV
genotyping data. NK and NP performed the patient recruitment
procedures, as well as the surgical operations and treatments on
the enrolled subjects. KWi performed the urine sample analyses and
data collection. WU was involved in sample and data collection. TS
performed the histological examination and provided the pathology
information. JS, CA and NMP provided technical assistance and
contributed to the study design. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical Review
Board of the Chulabhorn Hospital (no. 31/2554), and written
informed consent was obtained from the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Ervik M,
Dikshit R, Eser S, Mathers CD, Rebelo M, Parkin DM, Forman D and
Bray F: GLOBOCAN 2012 v1.0, cancer incidence and mortality
worldwide: IARC cancer base No. 11 [Internet] (Lyon, France).
International Agency for Research on Cancer. 2013.http://globocan.iarc.frDecember 10–2016
|
|
3
|
von Knebel Doeberitz M, Reuschenbach M,
Schmidt D and Bergeron C: Biomarkers for cervical cancer screening:
The role of p16(INK4a) to highlight transforming HPV infections.
Expert Rev Proteomics. 9:149–163. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ostör AG: Natural history of cervical
intraepithelial neoplasia: A critical review. Int J Gynecol Pathol.
12:186–192. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Van Keer S, Pattyn J, Tjalma WAA, Van
Ostade X, Ieven M, Van Damme P and Vorsters A: First-void urine: A
potential biomarker source for triage of high-risk human
papillomavirus infected women. Eur J Obstet Gynecol Reprod Biol.
216:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ng HT, Yen MS, Chao KC, Chen CY and Yuan
CC: Radical hysterectomy: Past, present, and future. Eur J Gynaecol
Oncol. 26:585–588. 2005.PubMed/NCBI
|
|
8
|
Hoffman MS: Extent of radical
hysterectomy: Evolving emphasis. Gynecol Oncol. 94:1–9. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Umanzor J, Aguiluz M, Pineda C, Andrade S,
Erazo M, Flores C and Santillana S: Concurrent
cisplatin/gemcitabine chemotherapy along with radiotherapy in
locally advanced cervical carcinoma: A phase II trial. Gynecol
Oncol. 100:70–75. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tanaka T, Kokawa K and Umesaki N:
Preoperative chemotherapy with irinotecan and mitomycin for FIGO
stage IIIb cervical squamous cell carcinoma: A pilot study. Eur J
Gynaecol Oncol. 26:605–607. 2005.PubMed/NCBI
|
|
11
|
Linghu H, Xu XR, Mei YY, Tang JY, Tang LD
and Sun T: Response of early stage bulky cervical squamous
carcinoma to preoperative adjuvant chemotherapy. Chin Med Sci J.
19:116–119. 2004.PubMed/NCBI
|
|
12
|
Candelaria M, Garcia-Arias A, Cetina L and
Dueñas-Gonzalez A: Radiosensitizers in cervical cancer. Cisplatin
and beyond. Radiat Oncol. 1:152006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goto T, Kino N, Shirai T, Fujimura M,
Takahashi M and Shiromizu K: Late recurrence of invasive cervical
cancer: Twenty years' experience in a single cancer institute. J
Obstet Gynaecol Res. 31:514–519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anderson NG, Anderson NL and Tollaksen SL:
Proteins of human urine. I. Concentration and analysis by
two-dimensional electrophoresis. Clin Chem. 25:1199–1210.
1979.PubMed/NCBI
|
|
15
|
Aobchey T, Niamsup H, Siriaree S, Sookkheo
B, Boonyapranai K and Chen ST: Proteomic analysis of candidate
prognostic urinary marker for cervical cancer. J Proteomics
Bioinform. 6:245–251. 2013. View Article : Google Scholar
|
|
16
|
Cox J, Hein MY, Luber CA, Paron I, Nagaraj
N and Mann M: Accurate proteome-wide label-free quantification by
delayed normalization and maximal peptide ratio extraction, termed
MaxLFQ. Mol Cell Proteomics. 13:2513–2526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wong JW and Cagney G: An overview of
label-free quantitation methods in proteomics by mass spectrometry.
Methods Mol Biol. 604:273–283. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Apgar BS, Zoschnick L and Wright Jr TC:
The 2001 Bethesda system terminology. Am Fam Physician.
68:1992–1998. 2003.PubMed/NCBI
|
|
19
|
Kantathavorn N, Mahidol C, Sritana N,
Sricharunrat T, Phoolcharoen N, Auewarakul C, Teerayathanakul N,
Taepisitpong C, Saeloo S, Sornsamdang G, et al: Genotypic
distribution of human papillomavirus (HPV) and cervical cytology
findings in 5906 Thai women undergoing cervical cancer screening
programs. Infect Agent Cancer. 10:72015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beetham R and Cattell WR: Proteinuria:
Pathophysiology, significance and recommendations for measurement
in clinical practice. Ann Clin Biochem. 30:425–434. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pappa KI, Lygirou V, Kontostathi G,
Zoidakis J, Makridakis M, Vougas K, Daskalakis G, Polyzos A and
Anagnou NP: Proteomic analysis of normal and cancer cervical cell
lines reveals deregulation of cytoskeleton-associated proteins.
Cancer Genomics Proteomics. 14:253–266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang HT, Tian EB, Chen YL, Deng HT and
Wang QT: Proteomic analysis for finding serum pathogenic factors
and potential biomarkers in multiple myeloma. Chin Med J (Engl).
128:1108–1113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Välk K, Vooder T, Kolde R, Reintam MA,
Petzold C, Vilo J and Metspalu A: Gene expression profiles of
non-small cell lung cancer: Survival prediction and new biomarkers.
Oncology. 79:283–292. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He X, Wang Y, Zhang W, Li H, Luo R, Zhou
Y, Liao CL, Huang H, Lv X, Xie Z and He M: Screening differential
expression of serum proteins in AFP-negative HBV-related
hepatocellular carcinoma using iTRAQ-MALDI-MS/MS. Neoplasma.
61:17–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Furukawa K, Kawamoto K, Eguchi H, Tanemura
M, Tanida T, Tomimaru Y, Akita H, Hama N, Wada H, Kobayashi S, et
al: Clinicopathological significance of leucine-rich
α2-glycoprotein-1 in sera of patients with pancreatic cancer.
Pancreas. 44:93–98. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu J, Xie X, Nie S, Buckanovich RJ and
Lubman DM: Altered expression of sialylated glycoproteins in
ovarian cancer sera using lectin-based ELISA assay and quantitative
glycoproteomics analysis. J Proteome Res. 12:3342–3352. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khammanivong A, Wang C, Sorenson BS, Ross
KF and Herzberg MC: S100A8/A9 (calprotectin) negatively regulates
G2/M cell cycle progression and growth of squamous cell carcinoma.
PLoS One. 8:e693952013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Silva EJ, Argyris PP, Zou X, Ross KF and
Herzberg MC: S100A8/A9 regulates MMP-2 expression and invasion and
migration by carcinoma cells. Int J Biochem Cell Biol. 55:279–287.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hermani A, Hess J, De Servi B, Medunjanin
S, Grobholz R, Trojan L, Angel P and Mayer D: Calcium-binding
proteins S100A8 and S100A9 as novel diagnostic markers in human
prostate cancer. Clin Cancer Res. 11:5146–5152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang L, Chang EW, Wong SC, Ong SM, Chong
DQ and Ling KL: Increased myeloid-derived suppressor cells in
gastric cancer correlate with cancer stage and plasma S100A8/A9
proinflammatory proteins. J Immunol. 190:794–804. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ito Y, Arai K, Nozawa R, Yoshida H,
Hirokawa M, Fukushima M, Inoue H, Tomoda C, Kihara M, Higashiyama
T, et al: S100A8 and S100A9 expression is a crucial factor for
dedifferentiation in thyroid carcinoma. Anticancer Res.
29:4157–4161. 2009.PubMed/NCBI
|
|
33
|
Duan L, Wu R, Ye L, Wang H, Yang X, Zhang
Y, Chen X, Zuo G, Zhang Y, Weng Y, et al: S100A8 and S100A9 are
associated with colorectal carcinoma progression and contribute to
colorectal carcinoma cell survival and migration via Wnt/β-catenin
pathway. PLoS One. 8:e620922013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ott HW, Lindner H, Sarg B, Mueller-Holzner
E, Abendstein B, Bergant A, Fessler S, Schwaerzler P, Zeimet A,
Marth C and Illmensee K: Calgranulins in cystic fluid and serum
from patients with ovarian carcinomas. Cancer Res. 63:7507–7514.
2003.PubMed/NCBI
|
|
35
|
Minami S, Sato Y, Matsumoto T, Kageyama T,
Kawashima Y, Yoshio K, Ishii J, Matsumoto K, Nagashio R and Okayasu
I: Proteomic study of sera from patients with bladder cancer:
Usefulness of S100A8 and S100A9 proteins. Cancer Genomics
Proteomics. 7:181–189. 2010.PubMed/NCBI
|
|
36
|
El Gammal AT, Sturm JH, Pinnschmidt HO,
Hofmann BT, Bellon E, Ghadban T, Melling NT, Bachmann KA, Izbicki
J, Bockhorn M, et al: Protein S100A8/A9: A potential new biomarker
for pancreatic diseases. Int J Clin Endocrinol Metab. 3:23–28.
2017. View Article : Google Scholar
|
|
37
|
Tugizov S, Berline J, Herrera R, Penaranda
ME, Nakagawa M and Palefsky J: Inhibition of human papillomavirus
type 16 E7 phosphorylation by the S100 MRP-8/14 protein complex. J
Virol. 79:1099–1112. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kong JP, Ding F, Zhou CN, Wang XQ, Miao
XP, Wu M and Liu ZH: Loss of myeloid-related proteins 8 and
myeloid-related proteins 14 expression in human esophageal squamous
cell carcinoma correlates with poor differentiation. World J
Gastroenterol. 10:1093–1097. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang J, Cai Y, Xu H, Zhao J, Xu X, Han YL,
Xu ZX, Chen BS, Hu H, Wu M and Wang MR: Expression of MRP14 gene is
frequently down-regulated in Chinese human esophageal cancer. Cell
Res. 14:46–53. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Argyris PP, Slama ZM, Ross KF,
Khammanivong A and Herzberg MC: Calprotectin and the initiation and
progression of head and neck cancer. J Dent Res. 97:674–682. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Khammanivong A, Sorenson BS, Ross KF,
Dickerson EB, Hasina R, Lingen MW and Herzberg MC: Involvement of
calprotectin (S100A8/A9) in molecular pathways associated with
HNSCC. Oncotarget. 7:14029–14047. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dalchau R, Kirkley J and Fabre JW:
Monoclonal antibody to a human leukocyte-specific membrane
glycoprotein probably homologous to the leukocyte-common (L-C)
antigen of the rat. Eur J Immunol. 10:737–744. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kosunen A, Pirinen R, Ropponen K, Pukkila
M, Kellokoski J, Virtaniemi J, Sironen R, Juhola M, Kumpulainen E,
Johansson R, et al: CD44 expression and its relationship with
MMP-9, clinicopathological factors and survival in oral squamous
cell carcinoma. Oral Oncol. 43:51–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mostaan LV, Khorsandi MT, Sharifian SM,
Shandiz FH, Mirashrafi F, Sabzari H, Badiee R, Borghei H and
Yazdani N: Correlation between E-cadherin and CD44 adhesion
molecules expression and cervical lymph node metastasis in oral
tongue SCC: Predictive significance or not. Pathol Res Pract.
207:448–451. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Urquidi V, Kim J, Chang M, Dai Y, Rosser
CJ and Goodison S: CCL18 in a multiplex urine-based assay for the
detection of bladder cancer. PLoS One. 7:e377972012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Arville B, O'Rourke E, Chung F, Amin M and
Bose S: Evaluation of a triple combination of cytokeratin 20, p53
and CD44 for improving detection of urothelial carcinoma in urine
cytology specimens. Cytojournal. 10:252013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao S, Zhou Y, Jiang J, Yuan L and Xue M:
CD44 affects the expression level of FOS-like antigen 1 in cervical
cancer tissues. Mol Med Rep. 9:1667–1674. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wobus M, Kuns R, Wolf C, Horn LC, Köhler
U, Sheyn I, Werness BA and Sherman LS: CD44 mediates constitutive
type I receptor signaling in cervical carcinoma cells. Gynecol
Oncol. 83:227–234. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kato H: Expression and function of
squamous cell carcinoma antigen. Anticancer Res. 16:2149–2153.
1996.PubMed/NCBI
|
|
50
|
Sun Y, Sheshadri N and Zong WX: SERPINB3
and B4: From biochemistry to biology. Semin Cell Dev Biol.
62:170–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hamanaka S, Ujihara M, Numa F and Kato H:
Serum level of squamous cell carcinoma antigen as a new indicator
of disease activity in patients with psoriasis. Arch Dermatol.
133:393–395. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vidalino L, Doria A, Quarta S, Zen M,
Gatta A and Pontisso P: SERPINB3, apoptosis and autoimmunity.
Autoimmun Rev. 9:108–112. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nakashima T, Yasumatsu R, Kuratomi Y,
Masuda M, Kuwano T, Toh S, Umezaki T, Cataltepe S, Silverman GA and
Komune S: Role of squamous cell carcinoma antigen 1 expression in
the invasive potential of head and neck squamous cell carcinoma.
Head Neck. 28:24–30. 2006. View Article : Google Scholar : PubMed/NCBI
|