Introduction
Lung cancer is the most prevalent type of cancer
worldwide, and ~80–85% of this malignancy comprises the non-small
cell lung cancer (NSCLC) (1). NSCLC
is an aggressive type of tumor with a 5-year survival rate of 16%.
Although NSCLC diagnosis is made at an early stage, and despite the
availability of numerous treatments, including surgery, radiation
or adjuvant chemotherapy, ~50% of patients with NSCLC relapse
(2). The development of novel
efficient therapeutic alternatives for NSCLC is therefore
crucial.
Cell migration is a key step in tumor development
and metastasis (3). Novel therapies
that could inhibit cell migration have therefore been considered as
novel approaches for lung cancer treatment. Previous studies have
revealed that vascular endothelial growth factor receptor (VEGFR)
signaling pathway, particularly VEGFR2, serves crucial role in
regulating cancer cell migration (4,5).
Furthermore, it has been reported that phosphorylated VEGFR2
activates downstream Ras homologous (Rho)-GTP hydrolase (GTPase),
induces cytoskeleton filaments remodeling and subsequently
activates tumor cell migration (6,7). In
addition, Ku et al (8)
demonstrated that VEGFR knockdown reduces hepatocellular carcinoma
migration. These findings suggest that VEGFR2 pathway serves an
important role in tumor cell migration.
Numerous plant-derived bioactive compounds,
including xylocoside G, formononetin and camptothecin, have been
isolated from traditional plants and are widely used to treat
cancers (9–11). The tripernoid corosolic acid (CA),
also known as 2α-hydroxyursolic acid, is present in numerous
plants, including Lagerstroemia speciosa L. and Actinidia
chinensis, and has exhibited anticancer ability against various
types of tumor. For example, CA induces apoptosis in multiple cell
types through different pathways as follows: CA induces apoptosis
of the cervix adenocarcinoma HeLa and osteosarcoma MG-63 cell lines
via mitochondrial signaling pathway; CA induces apoptosis of the
stomach carcinoma SNU-601 cell line through adenosine
monophosphate-activated protein kinase activation; and CA induces
apoptosis of the stomach carcinoma NCI-N87 cell line via human
epidermal growth factor receptor 2-downregulated cell cycle arrest
(12–14). However, the anticancer effects of CA
and its underlying mechanisms remain poorly understood in lung
cancer.
In the present study, CA exhibited a weak toxicity
in A549 cells, and decreased A549 cell migration. Results also
demonstrated that CA significantly inhibited VEGFR2 kinase
activity, and downregulated VEGFR2/Ras-related C3 botulinum toxin
substrate 1 (Rac1) binding ability. Data from mice xenograft
demonstrated that CA administration inhibited tumor growth in
vivo. Taken altogether, these results suggested that CA may be
considered as a potential novel therapy for NSCLC, and requires
therefore further investigation.
Materials and methods
Cell line and reagents
The A549 cell line was purchased from National
Infrastructure of Cell Line Resource (Beijing, China) and
maintained in high-glucose Dulbecco's modified Eagle's medium
(DMEM) supplemented with 10% fetal bovine serum (FBS; both Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Cells were
placed at 37°C in a humidified atmosphere containing 5%
CO2. CA, MTT, tyrphostin-SU 1498 (cat. no. SU 1498),
Tween-20, protease inhibitor cocktail (cat. no. P8340) and dimethyl
sulfoxide (DMSO) were obtained from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Recombinant Protein G-Sepharose 4B beads were
purchased from Thermo Fisher Scientific Inc. The rabbit primary
antibodies against GAPDH (cat. no. 2118; 1:5,000), VEGFR2 (cat. no.
2479; 1:2,000) and phosphorylated (p)-VEGFR2 (Tyr1059; cat. no.
3817; 1:1,000) were purchased from Cell Signaling Technology (Cell
Signaling Technology Europe, B.V., Leiden, The Netherlands). Rabbit
antibody against phosphoserine (P-Ser; cat. no. ab9332; 1:5,000)
was purchased from Abcam (Cambridge, UK). Horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG secondary antibody (1:3,000,
cat. no. A-21109) was obtained from Invitrogen (Thermo Fisher
Scientific, Inc.).
Cytotoxicity assay
MTT assay was employed to study CA toxicity on A549
cells as described previously (8,15,16).
Briefly, A549 cells were cultured in 96-well plate at the density
of 5×103 cells/well and treated with 0.1% DMSO (control)
or increasing concentrations of CA (20, 40, 60, 80 and 100 µM, CA
diluted in DMSO) for 48 h. (8,16) A
total of 20 µl MTT (5 mg/ml) was then added to cells for 6 h.
Formazan crystals were dissolved with SDS and absorbance was read
on a microplate reader [590/650 nm (absorbance/reference)
wavelengths].
Transwell migration assay
To determine cell migration rate, a Transwell assay
was employed (8 µm pore; cat. no. 351184; Corning Inc., Corning,
NY, USA). Briefly, A549 cells were serum-starved overnight and
resuspended in 300 µl serum-free DMEM medium containing 0.1% DMSO
(control) or increasing concentrations of CA (0, 1, 2, 4 and 8 µM).
Each sample contained 5×104 cells, which were cultured
in the Transwell upper chamber. The lower chamber was filled with
DMEM supplemented with 10% FBS. Following 16 h incubation, the
lower chamber was isolated and cells were fixed using 4%
paraformaldehyde at room temperature (RT) for 5 min. Cells were
then stained with crystal violet. Results represented the mean of
counting in three different areas under light microscope
(magnification, ×40), according to a previous study (17).
Western blotting and
co-immunoprecipitation (IP)
For western blotting, A549 cells or mouse tumor
tissues was lysed in radioimmunoprecipitation assay buffer (25 mM
Hepes, 150 mM NaCl, 10 mM MgCl2, 1% Nonidet P-40 and 10
mM DTT) and supplemented with protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA). The protein amount in each sample was
determined using NanoDrop 2000 (Thermo Fisher Scientific, Inc.) and
a total of 50 µg protein/lane was separated via SDS-PAGE on a 10%
gel. The proteins were subsequently transferred onto 0.45 µm
polyvinylidene difluoride membrane (Thermo Fisher Scientific, Inc.)
under 400 mA current for 1 h. The membrane was blocked with 5%
non-fat milk powder at RT for 1 h. The membranes were then
incubated with primary antibodies against VEGFR2 and p-VEGFR2
(Tyr1059) at 4°C over night. After incubation with HRP-conjugated
goat anti-rabbit IgG secondary antibody at RT for 1 h, proteins
were detected with Immobilon Western Chemiluminescent HRP Substrate
(ECL) reagent (cat no. WBKLS0500; EMD Millipore) in a dark room
with Kodak film and semi-quantified using ImageJ (release number
1.50i; National Institutes of Health).
For co-IP, A549 cells were treated for 24 h with
0.1% DMSO or increasing concentrations of CA (0, 4 and 8 µM). An
additional well of untreated cells was used as beads only control
(control). Cells were then lysed in reduced RIPA buffer [25 mM
Hepes, 150 mM NaCl, 10 mM MgCl2, 1% Nonidet P-40 and
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA)] and
centrifuged at 7,500 × g for 10 min at 4°C. Supernatant was
collected and protein concentration was determined by NanoDrop
2000. A total of 1 mg protein each group was incubated with
anti-VEGFR2-immobilized beads (1:100) (for experimental groups) or
without primary antibody (for control group) overnight at 4°C. A
quantity of 5% samples from each group was collected as input.
Samples were incubated with PureProteome magnetic beads (cat. no.
LSK MAGA10, EMD Millipore Billerica) for 1 h at 4°C and washed
three times with PBS containing 5% Tween-20 at 4°C. Beads were
collected with magnetic rock after each washing step and boiled at
95°C for 5 min. Samples were eventually detected by western
blotting as aforementioned. P-Ser primary antibody was used for IP
groups while VEGFR2 and GAPDH were used for input. Membranes were
incubated with primary antibodies overnight at 4°C, and with
HRP-conjugated goat anti-rabbit IgG secondary antibody for 2 h at
room temperature. Protein amount was determined as described
above.
Kinase activity assay
Kinase activity assay was performed with the
ADP-Glo™ kinase assay kit (Promega Corporation, Madison,
WI, USA). Increasing concentrations of CA were diluted with kinase
reaction buffer and added to tubes containing 3 ng of kinase insert
domain receptor (KDR; also known as VEGFR2) for 10 min. Then, 0.1
µg/µl enzyme substrate and 10 µM ATP were added to each tube for 1
h. ADP-Glo reagent (25 µl) was then added to each tube for 40 min
at room temperature. Eventually, 50 µl kinase detection buffer was
added to each tube. All samples were transferred onto a 96-well
plate and results were read with a microplate reader.
Rho-GTPase activity assay
A549 cells were treated with 0.1% DMSO (control) or
CA for 24 h and lysed with RIPA buffer. Tumor tissues from mouse
model was collected and lysed in RIPA buffer. Cell lysates (500 µg)
or tissue lysates (1 mg) were incubated with purified
glutathione-S-transferase (GST) fusion protein conjugated with Rac1
binding domain (PAK-PBD; Cytoskeleton, Inc., Denver, CO, USA) at
4°C overnight. MagneGST™ beads (Promega Corporation)
were added to each sample for further protein pull down. Pull down
samples were centrifuged at 12,000 × g for 30 min at 4°C and
detected by western blotting as previously described.
Labeling of mitochondria and
tubulin
A549 cells were cultured in 35 mm plastic dishes and
treated with 0.1% DMSO (control) or CA for 48 h. Cells were treated
with Tubulin Tracker™ and MitoTracker®
(Invitrogen, both diluted 1:1,000 as final concentration) for 5 min
at 37°C. Live cells were directly observed under confocal
microscope (magnification, ×60).
Animal xenograft model and in vivo
imaging system (IVIS) Spectrum Imaging
Animal experiments were performed under approved
protocol of the Institutional Animal Care and Use Committee of The
Hainan Medical University. The male NOD/SCID mice (4–6 weeks old;
n=60) were purchased from the Chinese Academy of Sciences, Beijing,
China and taken care of in the Laboratory Animal Center of Hainan
Medical University. They were kept into individually-ventilated
cages, and allowed free access to food and water. A549 cells
(2×106) were labeled with D-luciferin potassium by
Sinochrome (Shanghai, China; cat. no. BC-219-10) and suspended in
200 µl DMEM to reach the final density of 1×107
cells/ml. This cell suspension was eventually injected
subcutaneously into the flanks of each mouse. Two weeks later, a
total of 30 out of the 60 mice successfully grew tumor cells. These
mice subsequently received intraperitoneal (i.p.) daily injection
of 50 µl 10% DMSO (control) (8) or
various concentrations of CA (2, 4 or 8 mg/kg/day) for 14 days.
Each group consisted of 6,8,8 and 8 mice, respectively. Tumor size
was directly visualized by IVIS spectrum imaging system
(PerkinElmer, Inc., Waltham, MA, USA) and fluorescence intensity
was calculated.
Statistical analysis
Statistical analysis was conducted with GraphPad
Prism (version no. 4.0; GraphPad Software, Inc.). All data were
expressed as the means ± standard errors and calculated from at
least three independent experiments. Statistical comparisons were
assessed by using Student's t-test or one-way analysis of variance
followed by a post hoc analysis (Tukey test) when applicable.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CA significantly inhibits cells
migration
To determine the antitumor effects of CA in
vitro, the effect of CA on cell viability and migration were
assessed. A549 cells were treated with control (DMSO) or increasing
concentrations of CA (0–100 µg/ml) for 48 h prior to measuring cell
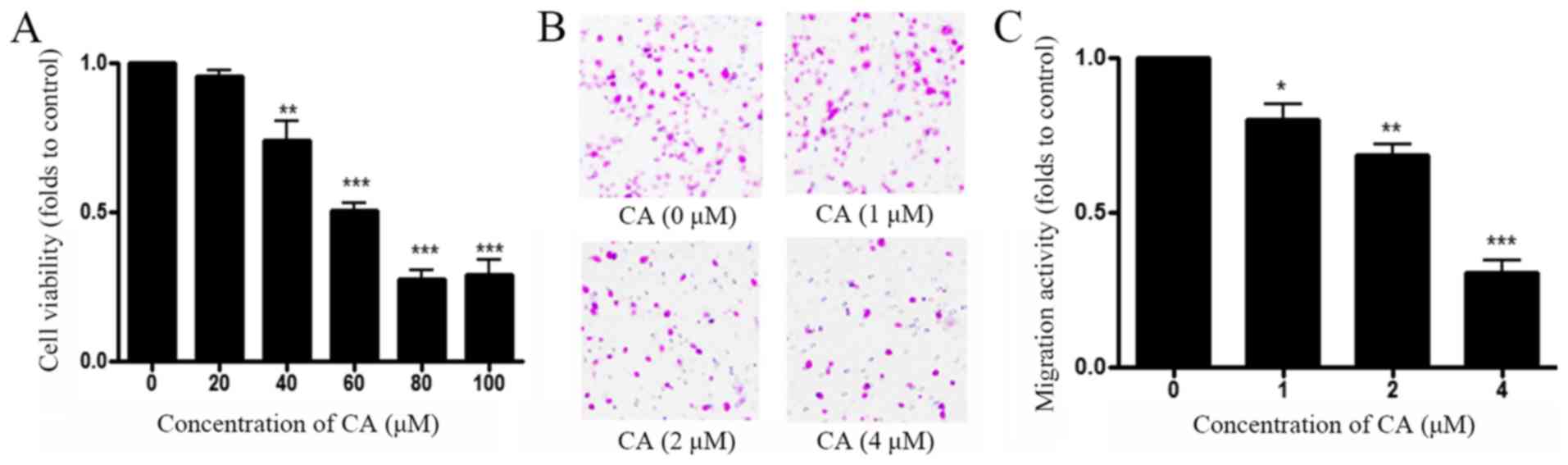
viability with MTT assay. As shown in Fig. 1A, CA decreased A549 cell viability in
a dose-dependent manner, and the half maximal inhibitory
concentration (IC50) was 65 µM. Transwell assay was
performed to measure A549 cell migration following treatment with
CA. Results demonstrated that CA inhibited A549 cell migration in a
dose-dependent manner, and IC50 for migration was 4 µM
(Fig. 1B and C). These results
indicated that CA inhibited more effectively A549 cell migration
than A549 cell viability with a ratio IC50
cytotoxicity/IC50 migration=16.
CA inhibits VEGFR2 kinase
activity
A previous study has reported that VEGF/VEGFR
signaling pathway may be involved in cancer cell migration, and
that VEGFR inhibition could reduce hepatocellular carcinoma cell
migration (8). To determine whether
VEGFR was involved in A549 cell migration, VEGFR pathway activity
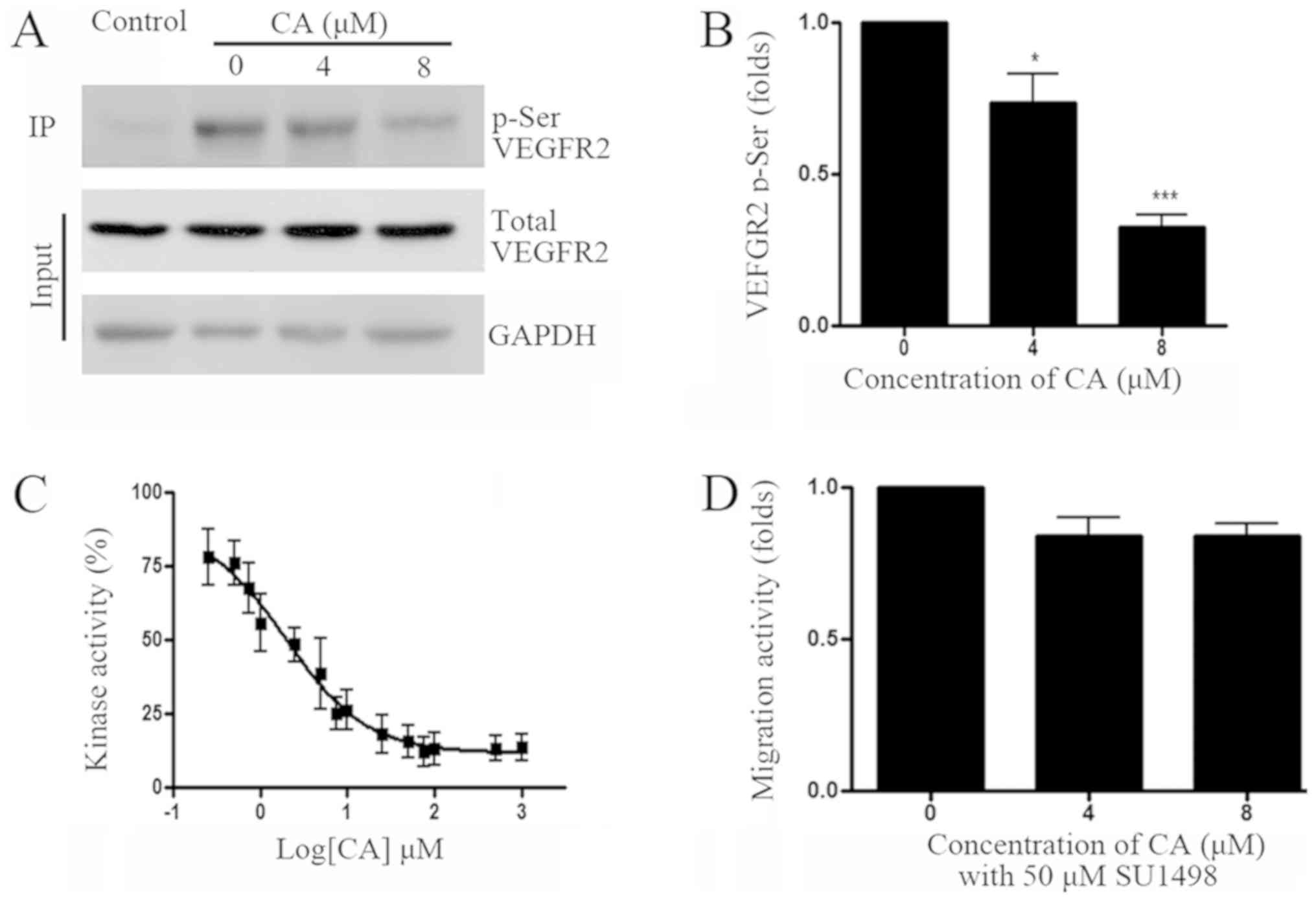
was assessed following A549 cell treatment with CA. Results from
co-IP and western blotting experiments indicated that CA
significantly reduced VEGFR2 phosphorylation. In addition,
determination of VEGFR2 kinase activity demonstrated that 1.95 µM
CA inhibited VEGFR2 activity by 50% (Fig. 2C). To further confirm whether VEGFR2
is involved in A549 cell migration, cell treatment with the VEGFR2
specific inhibitor SU1498 (50 µM) demonstrated that VEGFR2
inhibition may efficiently reduce A549 cell migration (Fig. 2D).
CA inhibits Rac1 activity and disrupts
tubulin organization
Small GTPase activation is an important downstream
event following VEGFR activation. The present study investigated
whether CA treatment could induce downstream Rho-GTP activation in
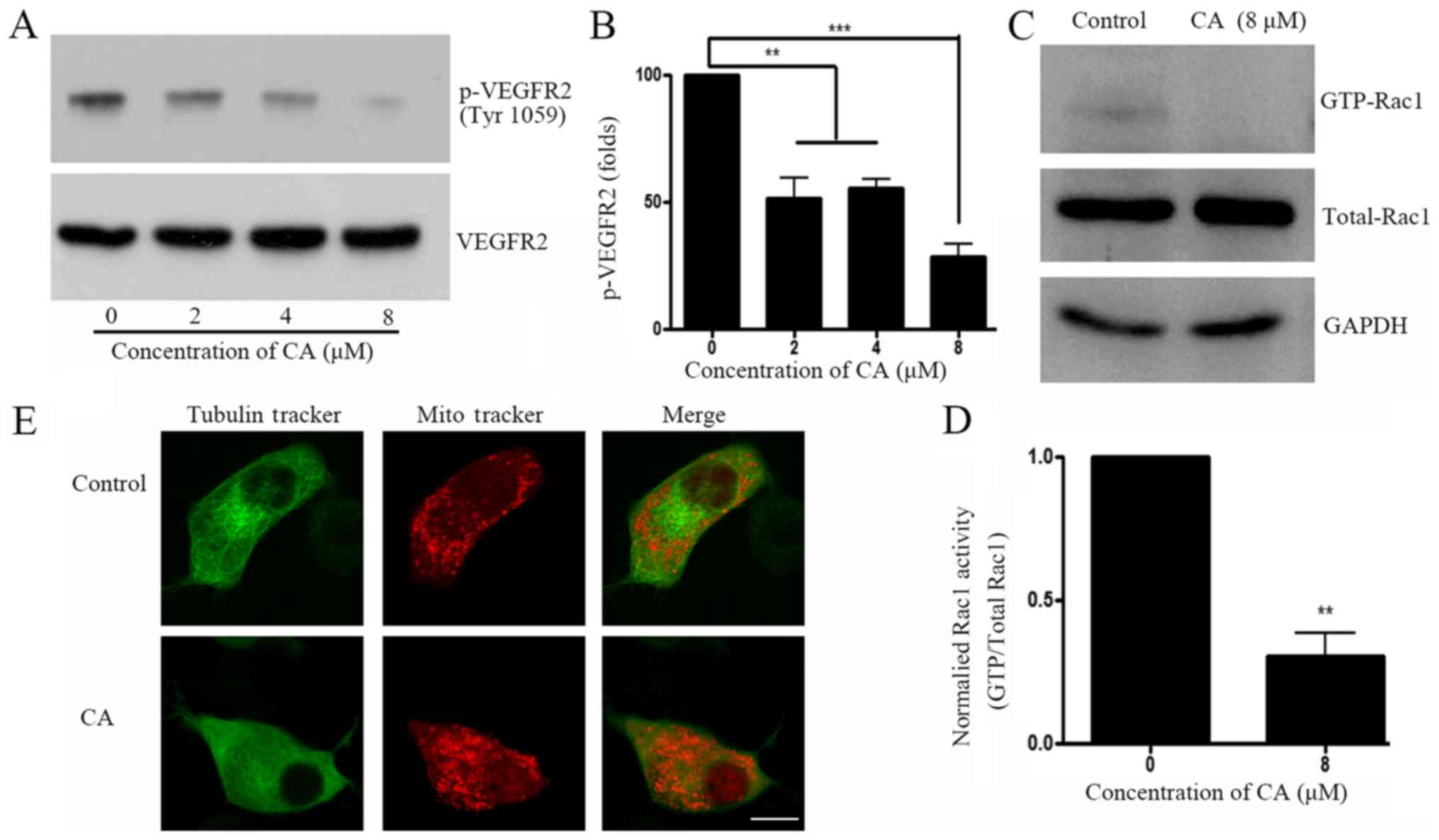
A549 cells. Results demonstrated that CA decreased VEGFR2
phosphorylation at Tyr1059 (Fig. 3A and
B), which is considered as an important phosphorylation site
that elevates downstream Rho-GTPase activity (18). Rho-GTP family activity was further
investigated, and results revealed that GTP-bounded-Rac1, but not
RhoA or cdc42 (data not shown) was significantly downregulated
following CA treatment (Fig. 3C and
D). Since small GTPases have crucial roles on cell skeleton
rearrangement and physiological organization (19), cell staining with Tubulin
Tracker™ and Mitotracker® was performed prior
to analysis with confocal microscopy. Images demonstrated that A549
cell treatment with CA induced tubulin network and mitochondria
distribution disruptions (Fig.
3E).
CA exhibits antitumor effect in
xenograft in vivo model by regulating Rac1 activity
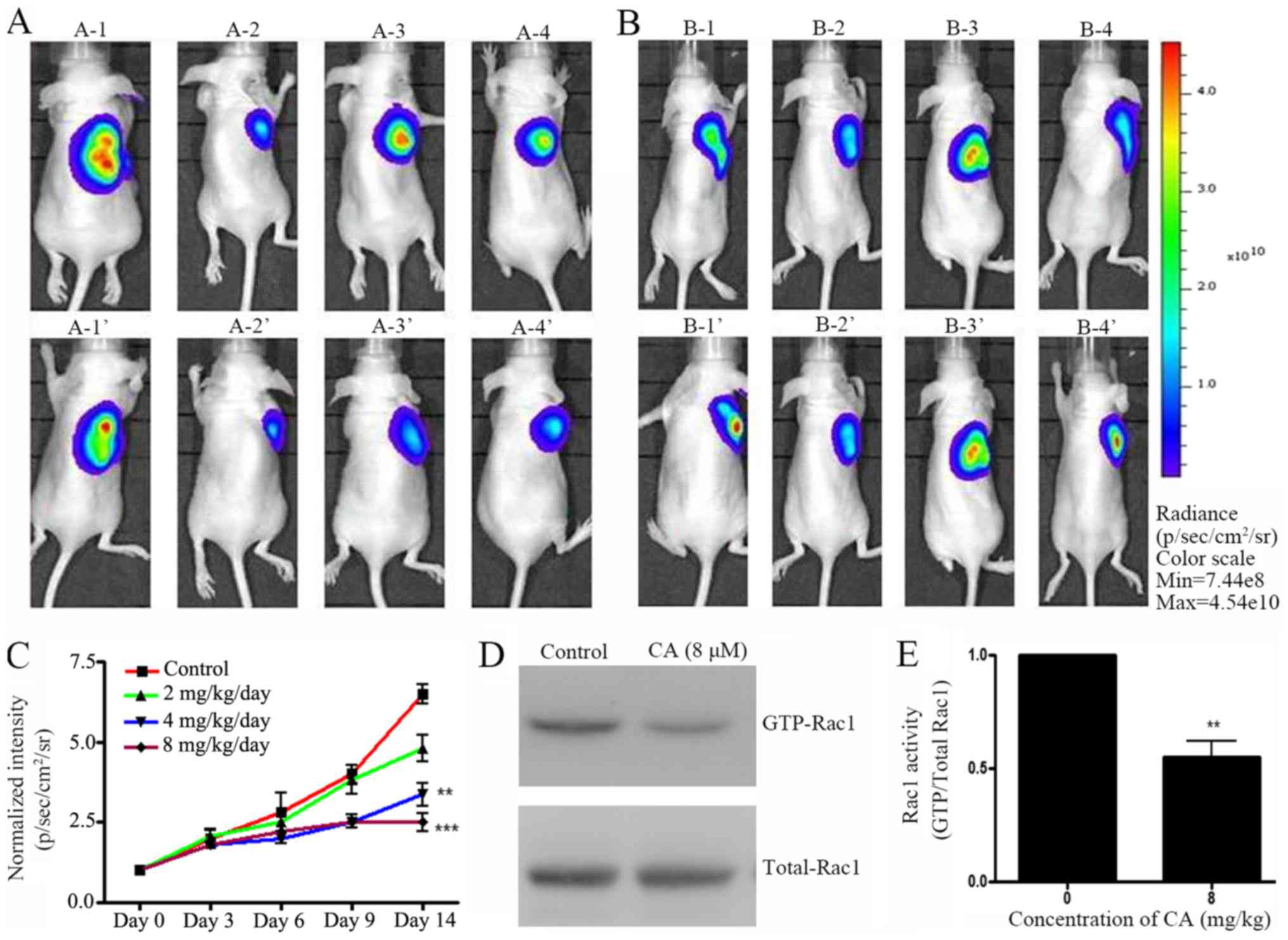
A xenograft mouse model was designed to investigate
the effect of CA in vivo. Pre-labeled A549 cells were
injected in mice and tumor growth was allowed for two weeks.
Preliminary data indicated that mg amount of CA could have an
antitumor effect in vivo (data not shown). Following A459
cell injection, mice were treated with various concentrations of CA
(0, 2, 4 and 8 mg/kg/day) daily (i.p. injection) for two weeks.
IVIS live imaging was employed daily to monitor tumor growth.
Results indicated that 14 days of CA treatment effectively
inhibited A549 cell tumor growth in NOD/SCID mice (Fig. 4A-C). Eventually, mice were sacrificed
and Rac1 activity in tumor tissues was measured. In accordance with
the in vitro results, CA-treated mice exhibited a decrease
in GTP-Rac1 protein level (Fig. 4D and
E). These results suggested that CA mechanisms of action may be
similar in vitro and in vivo.
Discussion
VEGF signaling pathway regulates cell migration,
proliferation and angiogenesis (20). VEGFR2 is a major receptor in VEGF
pathway. VEGFR2-associated signal transduction pathway is
initialized by VEGF/VEGFR2 binding, which can lead to VEGFR2
trans-autophosphorylation on its intracellular tyrosine residues
(21). Two residues in the VEGFR2
active loop domain were identified as Tyr1054 and Tyr1059, of which
phosphorylation is critical for VEGFR2 kinase activity (22). Tyr1054 and Tyr1059 phosphorylation
can induce downstream signal transduction, including p21-activated
kinase (PAK) activation and cytoskeleton remodeling (23). The present study demonstrated that CA
treatment reduced Tyr1059 phosphorylation level of VEGFR2, and
VEGFR2 activity. These results may have highlighted a potential
mechanism of action for the antimigratory effects of CA in A549
cells.
A previous study reported that VEGF-associated cell
migration is dependent of Rac1 activation, and that VEGFR-2
inhibition reduces Rac1 activation (24). Furthermore, VEGFR2
autophosphorylation is critical for VEGFR2-Rac1 pathway activation
(25). In the present study, VEGFR2
phosphorylation level in A549 cell was significantly reduced
following CA treatment with CA. In addition, the combined treatment
of A549 cells with CA and SU1498 did not enhance p-VEGFR2 decrease
compared with CA or SU1498 alone, which suggested that CA may
reduce cell migration through VEGFR2 phosphorylation.
Rho-GTPase activation and subsequent cytoskeleton
rearrangement represent crucial steps for cell migration. For
example, Rac1 regulation induces microtubules rearrangement
(26). With regards to the migration
stage, Rac1 is located at the leading edge of a migrating cell and
regulates actin and microtubule reorganization (27). As previously reported, the
constitutively active form of Rac1 (Q61L) promotes pioneer behavior
in most microtubules, whereas the dominant-negative form of Rac1
(T17N) eliminates these pioneer microtubules (28). In the present study, CA cell
treatment significantly reduced GTP-Rac1 expression, a finding that
has been previously reported (29).
Considering microtubules are one of the key cytoskeleton components
(30) and closely regulated
mitochondrial distribution (31,32), the
disrupted mitochondrial distribution would be a result of
microtubule abnormal organization and could reflect cytoskeleton
structure disruption. The current study further tested whether
microtubule structure was disrupted by CA treatment. CA treatment
could reduce tubulin density and as a consequence, the
mitochondrial distribution changed from arranged along cytoplasmic
microtubules (33) to scattered in
the perinuclear space. This result indicated CA treatment may
disrupt tubulin distribution and organization. Since CA inhibits
hepatocellular carcinoma cell migration via inhibition of the
VEGFR2-FAK pathway and downstream actin rearrangement (8), data from the present study strongly
suggests that CA may inhibit lung cancer A549 cell migration
through cytoskeleton disruption.
The present study has a number of limitations. Due
to the lack of information regarding the effect of CA in
vivo, lower concentrations of CA were tested in mice in order
to minimize a potential cytotoxicity effect. Preliminary data
supported that low concentration of CA may exhibit antitumor
effects in vivo (data not shown). However, due to the
resolution limit of the IVIS system, we can only use a moderate CA
concentration (mg/kg) to show a difference here. It is important to
further verify whether concentrations lower than 1 mg/kg exhibits a
similar effect. Eventually, although no changes in the body weight
or daily activities of the mice were observed following treatment
with a high dose of CA (8 mg/kg/day), it could not be excluded that
CA may have an effect on the behavior of the mice. Behavioral tests
could be employed to confirm the safety of CA in clinical use.
In conclusion, results from the present study
demonstrated that CA treatment exhibited a potential anticancer
activity in the A549 cell line and in in vivo. Particularly,
CA reduced the migration rate of A549 cells, inhibited VEGFR2
kinase activity, disrupted tubulin remodeling and reduced tumor
growth in vivo. In addition, CA exhibited its most potent
anticancer effects in the micromolar concentration range. In
conclusion, the present study suggests that CA may be considered as
a potential chemotherapeutic agent for lung cancer therapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BL designed the experiment and wrote the manuscript.
YL and QW performed the experiments and collected the data. FaL and
FuL analyzed the data and made the figures.
Ethics approval and consent to
participate
Animal experiments were performed under approved
protocol of the Institutional Animal Care and Use Committee of The
Hainan Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jaffe N: Osteosarcoma: Review of the past,
impact on the future. The American experience. Cancer Treat Res.
152:239–262. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Smeland S, Bruland OS, Hjorth L, Brosjö O,
Bjerkehagen B, Osterlundh G, Jakobson A, Hall KS, Monge OR, Björk O
and Alvegaard TA: Results of the scandinavian sarcoma group XIV
protocol for classical osteosarcoma: 63 patients with a minimum
follow-up of 4 years. Acta Orthop. 82:211–216. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martin R, Carvalho-Tavares J, Ibeas E,
Hernández M, Ruiz-Gutierrez V and Nieto ML: Acidic triterpenes
compromise growth and survival of astrocytoma cell lines by
regulating reactive oxygen species accumulation. Cancer Res.
67:3741–3751. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reyes-Zurita FJ, Rufino-Palomares EE,
Lupiáñez JA and Cascante M: Maslinic acid, a natural triterpene
from Olea europaea L., induces apoptosis in HT29 human colon-cancer
cells via the mitochondrial apoptotic pathway. Cancer Lett.
273:44–54. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lamalice L, Houle F and Huot J:
Phosphorylation of Tyr1214 within VEGFR-2 triggers the recruitment
of Nck and activation of Fyn leading to SAPK2/p38 activation and
endothelial cell migration in response to VEGF. J Biol Chem.
281:34009–34020. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ku CY, Wang YR, Lin HY, Lu SC and Lin JY:
Corosolic acid inhibits hepatocellular carcinoma cell migration by
targeting the VEGFR2/Src/FAK pathway. PLoS One. 10:e01267252015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prydz K, Vuong TT and Kolset SO:
Glycosaminoglycan secretion in xyloside treated polarized human
colon carcinoma Caco-2 cells. Glycoconj J. 26:1117–1124. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li T, Zhao X, Mo Z, Huang W, Yan H, Lin Z
and Ye Y: Formononetin promotes cell cycle arrest via
downregulation of Akt/Cyclin D1/CDK4 in human prostate cancer
cells. Cell Physiol Biochem. 34:1351–1358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jeansonne DP, Koh GY, Zhang F,
Kirk-Ballard H, Wolff L, Liu D, Eilertsen K and Liu Z:
Paclitaxel-induced apoptosis is blocked by camptothecin in human
breast and pancreatic cancer cells. Oncol Rep. 25:1473–1480.
2011.PubMed/NCBI
|
|
12
|
Lee MS, Lee CM, Cha EY, Thuong PT, Bae K,
Song IS, Noh SM and Sul JY: Activation of AMP-activated protein
kinase on human gastric cancer cells by apoptosis induced by
corosolic acid isolated from Weigela subsessilis. Phytother Res.
24:1857–1861. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cai X, Zhang H, Tong D, Tan Z, Han D, Ji F
and Hu W: Corosolic acid triggers mitochondria and
caspase-dependent apoptotic cell death in osteosarcoma MG-63 cells.
Phytother Res. 25:1354–1361. 2011.PubMed/NCBI
|
|
14
|
Horlad H, Fujiwara Y, Takemura K, Ohnishi
K, Ikeda T, Tsukamoto H, Mizuta H, Nishimura Y, Takeya M and
Komohara Y: Corosolic acid impairs tumor development and lung
metastasis by inhibiting the immunosuppressive activity of
myeloid-derived suppressor cells. Mol Nutr Food Res. 57:1046–1054.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu Y, Zhou L, Sun M, Zhou T, Zhong K, Wang
H, Liu Y, Liu X, Xiao R, Ge J, et al: Xylocoside G reduces
amyloid-β induced neurotoxicity by inhibiting NF-κB signaling
pathway in neuronal cells. J Alzheimers Dis. 30:263–275. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee HS, Park JB, Lee MS, Cha EY, Kim JY
and Sul JY: Corosolic acid enhances 5-fluorouracil-induced
apoptosis against SNU-620 human gastric carcinoma cells by
inhibition of mammalian target of rapamycin. Mol Med Rep.
12:4782–4788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun M and Zhang H: Par3 and aPKC regulate
BACE1 endosome-to-TGN trafficking through PACS1. Neurobiol Aging.
60:129–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen TT, Luque A, Lee S, Anderson SM,
Segura T and Iruela-Arispe ML: Anchorage of VEGF to the
extracellular matrix conveys differential signaling responses to
endothelial cells. J Cell Biol. 188:595–609. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Sun M, Yang H, Tian X, Tong Y,
Zhou T, Zhang T, Fu Y, Guo X, Fan D, et al: Hypoxia-inducible
factor-1α mediates up-regulation of neprilysin by histone
deacetylase-1 under hypoxia condition in neuroblastoma cells. J
Neurochem. 131:4–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garrett TA, Van Buul JD and Burridge K:
VEGF-induced Rac1 activation in endothelial cells is regulated by
the guanine nucleotide exchange factor Vav2. Exp Cell Res.
313:3285–3297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vazgiourakis VM, Zervou MI, Eliopoulos E,
Sharma S, Sidiropoulos P, Franek BS, Myrthianou E, Melissourgaki M,
Niewold TB, Boumpas DT and Goulielmos GN: Implication of VEGFR2 in
systemic lupus erythematosus: A combined genetic and structural
biological approach. Clin Exp Rheumatol. 31:97–102. 2013.PubMed/NCBI
|
|
22
|
Roskoski R Jr: VEGF receptor
protein-tyrosine kinases: Structure and regulation. Biochem Biophys
Res Commun. 375:287–291. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takahashi T, Yamaguchi S, Chida K and
Shibuya M: A single autophosphorylation site on KDR/Flk-1 is
essential for VEGF-A-dependent activation of PLC-gamma and DNA
synthesis in vascular endothelial cells. EMBO J. 20:2768–2778.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meissner M, Michailidou D, Stein M,
Hrgovic I, Kaufmann R and Gille J: Inhibition of Rac1 GTPase
downregulates vascular endothelial growth factor receptor-2
expression by suppressing Sp1-dependent DNA binding in human
endothelial cells. Exp Dermatol. 18:863–869. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clegg LW and Mac Gabhann F: Site-specific
phosphorylation of VEGFR2 is mediated by receptor trafficking:
Insights from a computational model. PLoS Comput Biol.
11:e10041582015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singleton PA, Dudek SM, Chiang ET and
Garcia JG: Regulation of sphingosine 1-phosphate-induced
endothelial cytoskeletal rearrangement and barrier enhancement by
S1P1 receptor, PI3 kinase, Tiam1/Rac1, and alpha-actinin. FASEB J.
19:1646–1656. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wittmann T, Bokoch GM and Waterman-Storer
CM: Regulation of leading edge microtubule and actin dynamics
downstream of Rac1. J Cell Biol. 161:845–851. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shin OH and Exton JH: Differential binding
of arfaptin 2/POR1 to ADP-ribosylation factors and Rac1. Biochem
Biophys Res Commun. 285:1267–1273. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li XQ, Tian W, Liu XX, Zhang K, Huo JC,
Liu WJ, Li P, Xiao X, Zhao MG and Cao W: Corosolic acid inhibits
the proliferation of glomerular mesangial cells and protects
against diabetic renal damage. Sci Rep. 6:268542016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang P and MacRae TH: Molecular
chaperones and the cytoskeleton. J Cell Sci. 110:1431–1440.
1997.PubMed/NCBI
|
|
31
|
Frederick RL and Shaw JM: Moving
mitochondria: Establishing distribution of an essential organelle.
Traffic. 8:1668–1675. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Woods LC, Berbusse GW and Naylor K:
Microtubules are essential for mitochondrial dynamics-fission,
fusion, and motility-in dictyostelium discoideum. Front Cell Dev
Biol. 4:192016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heggeness MH, Simon M and Singer SJ:
Association of mitochondria with microtubules in cultured cells.
Proc Natl Acad Sci USA. 75:3863–3866. 1978. View Article : Google Scholar : PubMed/NCBI
|