Introduction
Melanoma, the most aggressive type of skin cancer,
develops from melanocytes and accounts for 4% of all cancer types
diagnosed in adolescents worldwide (1–3). While
the incidence rate of melanoma is lower than other types of skin
cancer, it can grow rapidly and metastasize when it is not detected
and treated early (4–6). Therefore, late-stage melanoma can be
difficult to treat and fatal (7).
The 5-year survival rate ranges between 15 and 60% depending on the
stage of the melanoma (4,8).
It is understood that disordered gene expression
serves an important role in the development of cancer (9). Gene expression profiling can provide
evidence to determine the diagnosis and identify prognostic markers
or novel therapeutic targets in cancer (10,11).
However, a limited number of previous studies have investigated the
role of gene expression by examining alternative splicing events
(9,12,13).
Alternative splicing can lead to the coding of
several types of proteins. During alternative splicing, specific
exons of a gene can be included or excluded and the alternatively
spliced mRNAs are then translated into proteins that exhibit
different biological functions (12–14).
Alternative splicing regulates gene expression and serves a key
role in this process (9). Defects in
mRNA splicing can result in aberrant splicing, in addition to
oncogenic processes (15–17). A number of previous studies have
demonstrated that splicing defects act as promoters of cancer
(16,18–20).
Furthermore, different splicing factors may cause specific
cancer-promoting isoforms.
It has been demonstrated that alternative splicing
exhibits prognostic value in patients with lung (21), ovarian (22) and breast cancer (23), and glioblastoma (24). However, to the best of our knowledge,
a systematic survival analysis of alternative splicing in melanoma
has not been previously performed, and thus is urgently required.
The aim of the present study was to determine prognostic
alternative splicing events in melanoma using RNA sequencing
(RNA-seq) data from The Cancer Genome Atlas (TCGA).
Materials and methods
Alternative splicing data
collection
Melanoma cohort RNA-seq data (25) were downloaded from TCGA data portal
(tcga-data.nci.nih.gov/tcga). Data of
96 melanoma cases with clinicopathological information were
obtained to explore the changes of alternative splicing events in
association with the carcinogenesis and prognosis of melanoma. To
analyze the alternative splicing profiles for each patient, a
SpliceSeq tool (version 2.1) (26),
which is a java application, was used in the melanoma cohort to
evaluate the splicing patterns of mRNA. The percent spliced in
(PSI) value was calculated to quantify alternative splicing events
ranging between 0 and 1 in seven types of alternative splicing
events, including exon skip (ES), alternate terminator (AT),
mutually exclusive (ME) exons, retained intron (RI), alternate
promoter (AP), alternate acceptor (AA) site and alternate donor
(AD) site.
Survival analysis
The melanoma clinical data were downloaded from TCGA
database. Only patients with an overall survival (OS) time >90
days were enrolled in the study. A total of 96 patients with
melanoma were included in the present study. The patients were
divided into two groups (high- and low-risk groups) according to
the median cut-off value (0.078). Kaplan-Meier and Univariate Cox
regression analyses were used to analyze associations between
splicing factor genes and survival, and multivariate Cox regression
was performed to remove non-independent predictors. To compare the
efficiencies of each prediction model, receiver operator
characteristic (ROC) curves were generated using the survival
receiver ROC package (version 1.0.3; cran.r-project.org/web/packages/survivalROC/index.html)
in R software (version 3.3.0; cran.r-project.org/bin/windows/base/old/3.3.0).
To ascertain the predictive ability, the area under the curve (AUC)
was calculated for each model.
Statistical analysis
The UpSet package (version 1.3.3; CRAN.R-project.org/package=UpSetR) in R
software was used to quantitatively analyze the alternative
splicing intersections for all seven types of alternative splicing
events (27). The most significant
alternative splicing events were selected to generate a gene
network using the Reactome FI Cytoscape 3.6.0 plugin to search for
the key hub genes of survival associated with the alternative
splicing genes (28). Cytoscape
3.6.0 was then used to generate correlation plots. Spearman's rank
correlation coefficient was used to analyze the correlation between
the PSI values and the splicing factor gene expression to identify
potential prognostic biomarkers for melanoma.
Results
Overview of alternative splicing
events in TCGA melanoma cohort
An in-depth analysis of integrated alternative
splicing events was conducted for 96 patients with melanoma with an
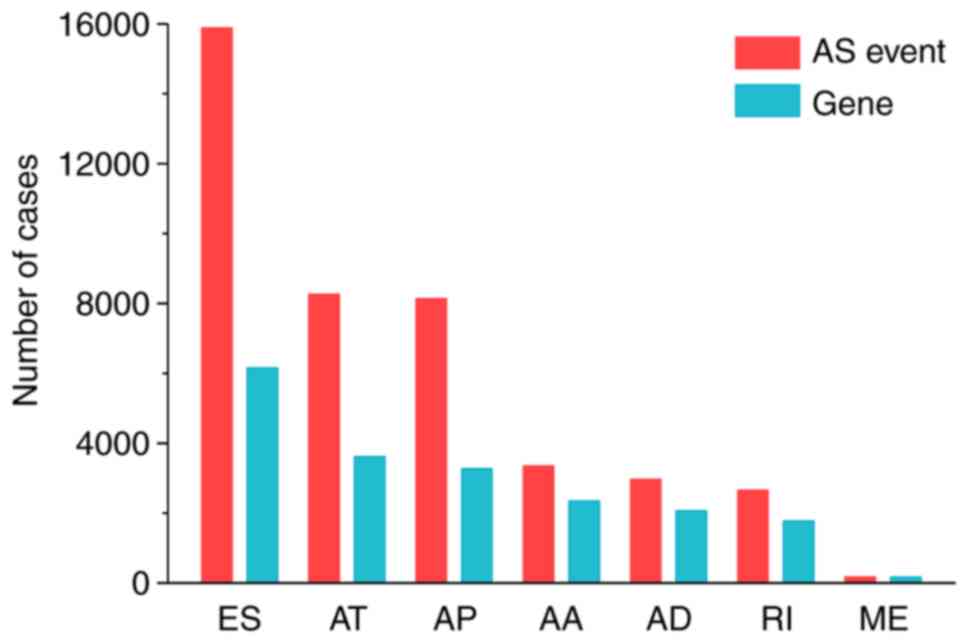
OS time >90 days. In the melanoma cohort, 41,446 mRNA splicing
events were detected in 9,780 genes, which consisted of 15,892 ESs
in 6,160 genes, 8,267 ATs in 3,614 genes, 8,135 APs in 3,273 genes
and 3,315 AAs in 2,350 genes, 2,968 ADs in 2,070 genes, 2,656 RIs
in 1,780 genes and 177 MEs in 175 genes. The results suggested that
several mRNA splicing events occur in one gene and ~38% of the
alternative splicing events were ES events, followed by 19.9% AT
events and 19.6% AP events (Fig.
1).
Survival-associated alternative
splicing events in TCGA melanoma cohort
Every alternative splicing event in TCGA melanoma
cohort was evaluated for its association with survival. The
patients with melanoma were divided into two groups (high- and
low-risk groups) based on the median PSI value. A total of 2,348
survival-associated alternative splicing events (data not shown)
were detected in melanoma (P<0.05). The top ten most significant
events in each type of alternative splicing event were selected.
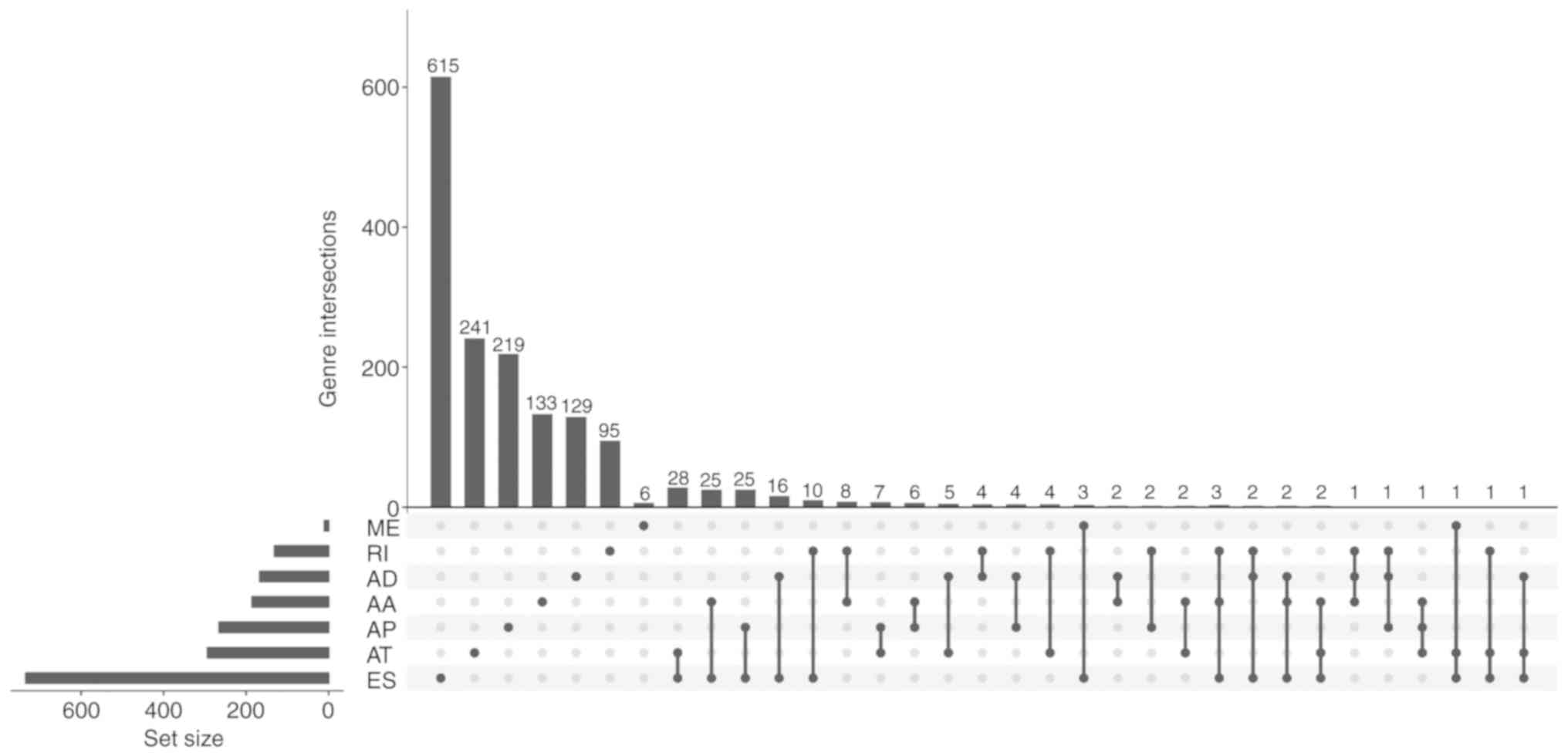
More than one alternative splicing event could occur in one gene,
therefore, an UpSet plot was generated to visualize the
intersecting sets. As demonstrated in Fig. 2, up to three types of splicing event
were identified in one gene; however, the majority of the
survival-associated alternative splicing events were from one gene
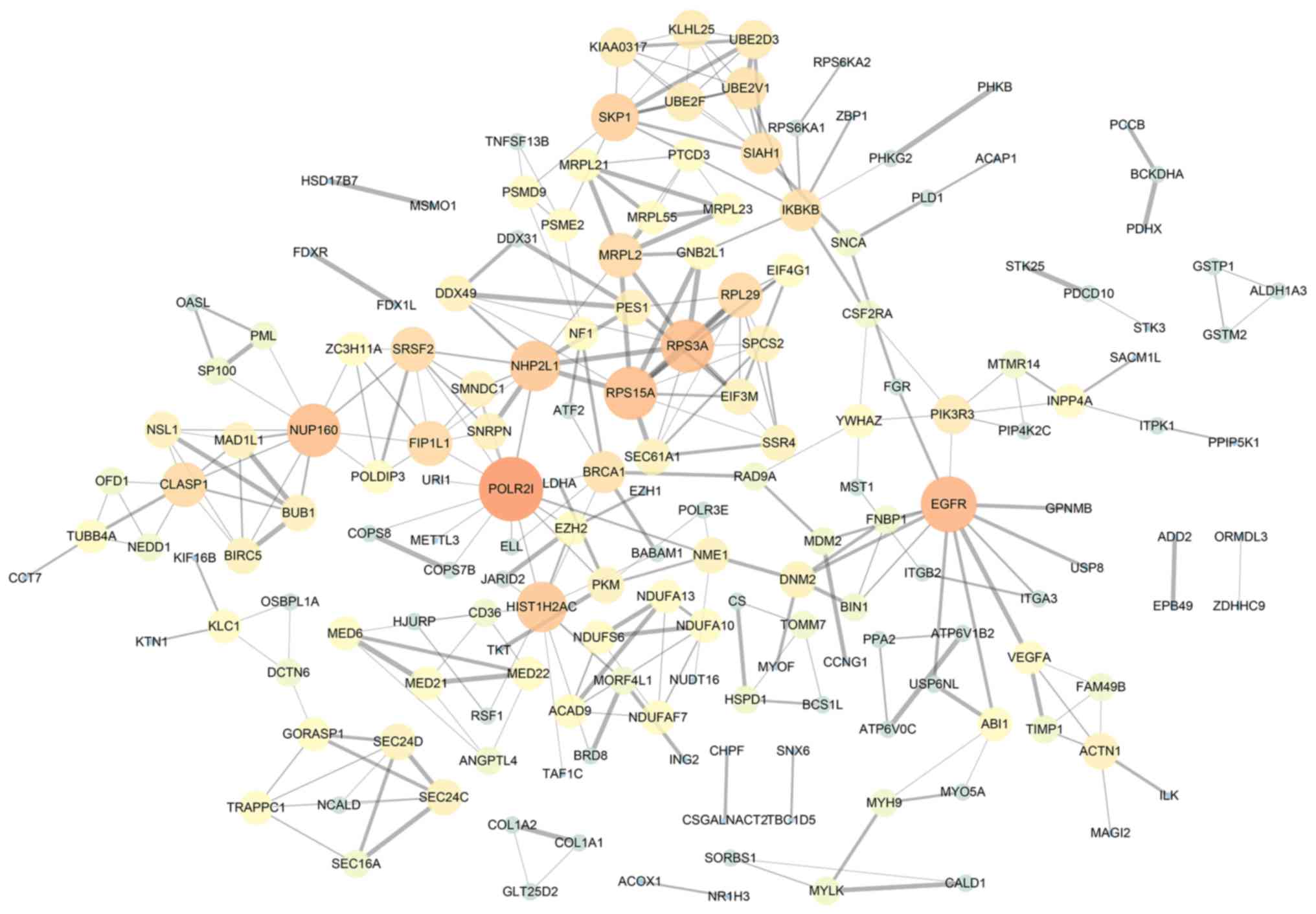
and these were all associated with patient survival. Furthermore, a
gene network was created by Cytoscape analysis. This revealed
important cancer pathways in melanoma by the identification of the
following hub genes: RNA polymerase II subunit I, epidermal growth
factor receptor, nucleoporin 160, ribosomal protein S15A and
ribosomal protein S3A (Fig. 3).
Notably, numerous favorable prognostic factors were identified in
the survival-associated alternative splicing events.
Prognostic predictors for patients
with melanoma
The OS time of patients with melanoma ranged between
126 and 1,785 days. The top ten most significant events in the
seven types of alternative splicing events were selected to
determine the independent prognostic factors for patients with
melanoma. Multivariate Cox regression analysis with a prognostic
model construction was used to analyze the seven types of
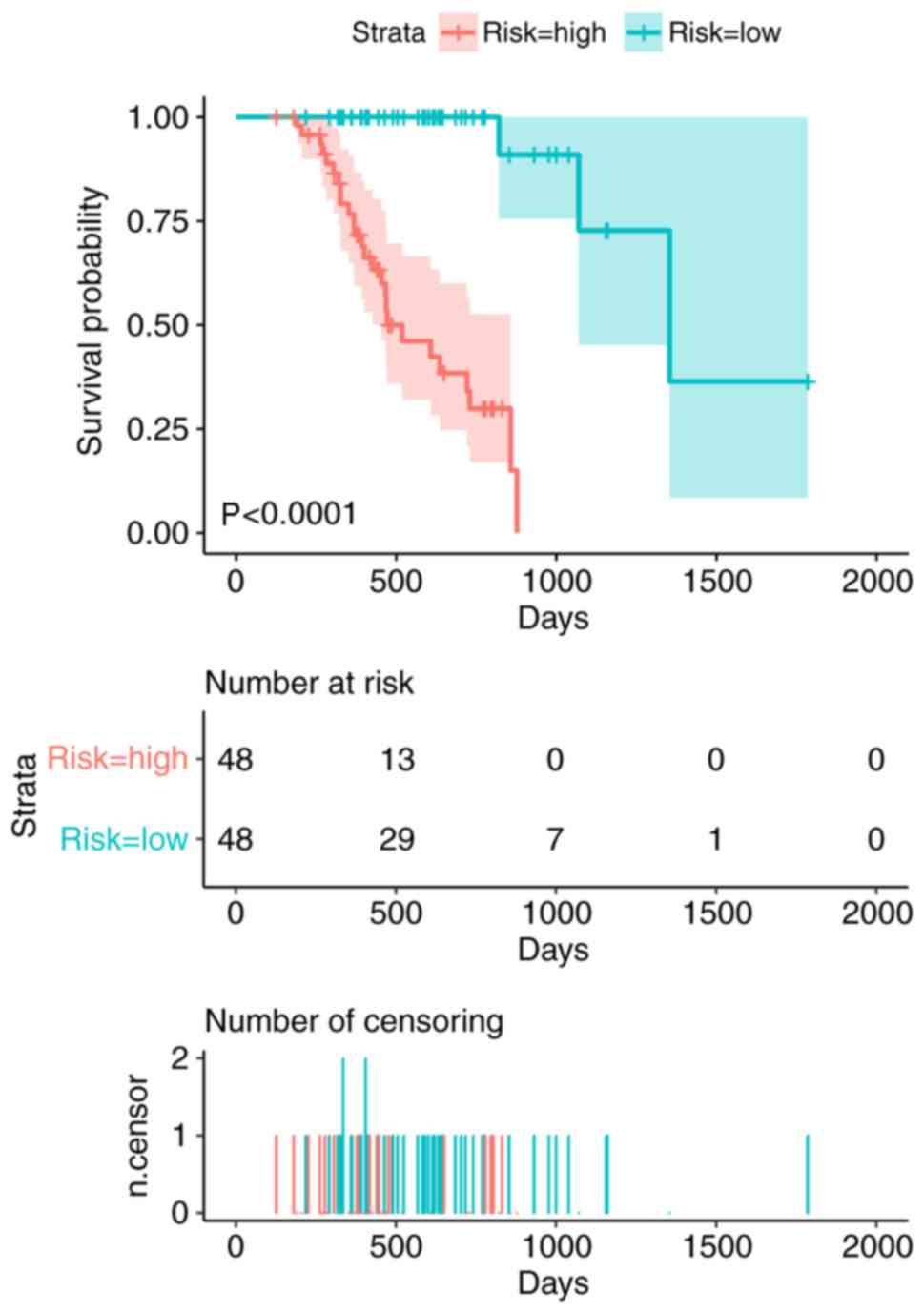
alternative splicing events separately, as demonstrated in Fig. 4. Kaplan-Meier curves of the
prognostic predictors were generated for AA, AD, AP, AT, ES, ME and
RI. The red line indicates the high-risk group and the blue line
indicates the low-risk group. The seven predictors were integrated
for all types of alternative splicing events in order to analyze
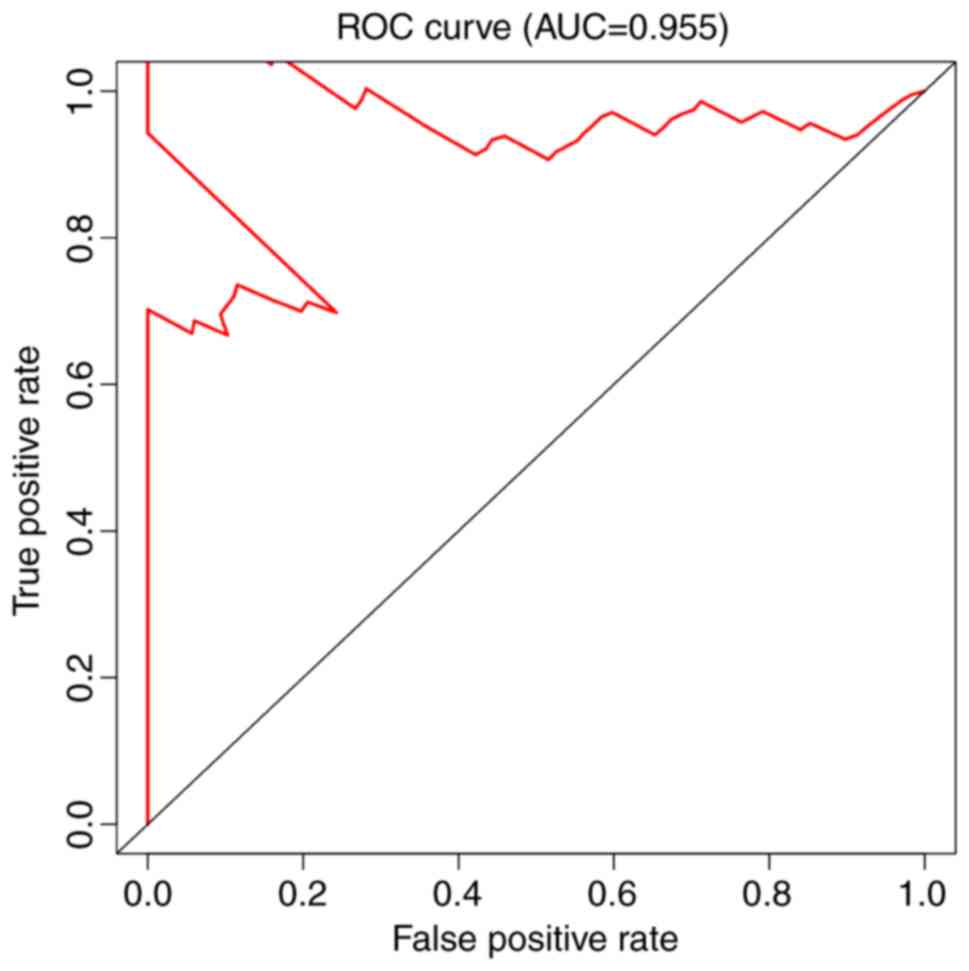
the final prognostic model for patients with melanoma (Fig. 5). ROC curves were generated for the
individual types of splicing event (Fig.
6) and for a combination of all types of splicing events
(Fig. 7), and AUC analysis was used
to evaluate the efficiency of the prognostic models. In the
high-risk and low-risk groups, the median OS times were 472 and
1,354 days, respectively (P<0.0001; Fig. 5). Of all the models, the model with
the AT alternative splicing events exhibited the highest efficiency
for evaluating outcomes for patients with melanoma (Fig. 6D). The AUC value of the AT
alternative splicing events was 0.902, followed by the AP model
with an AUC value of 0.887 (Fig. 6C)
and the RI model with an AUC value of 0.809 (Fig. 6G).
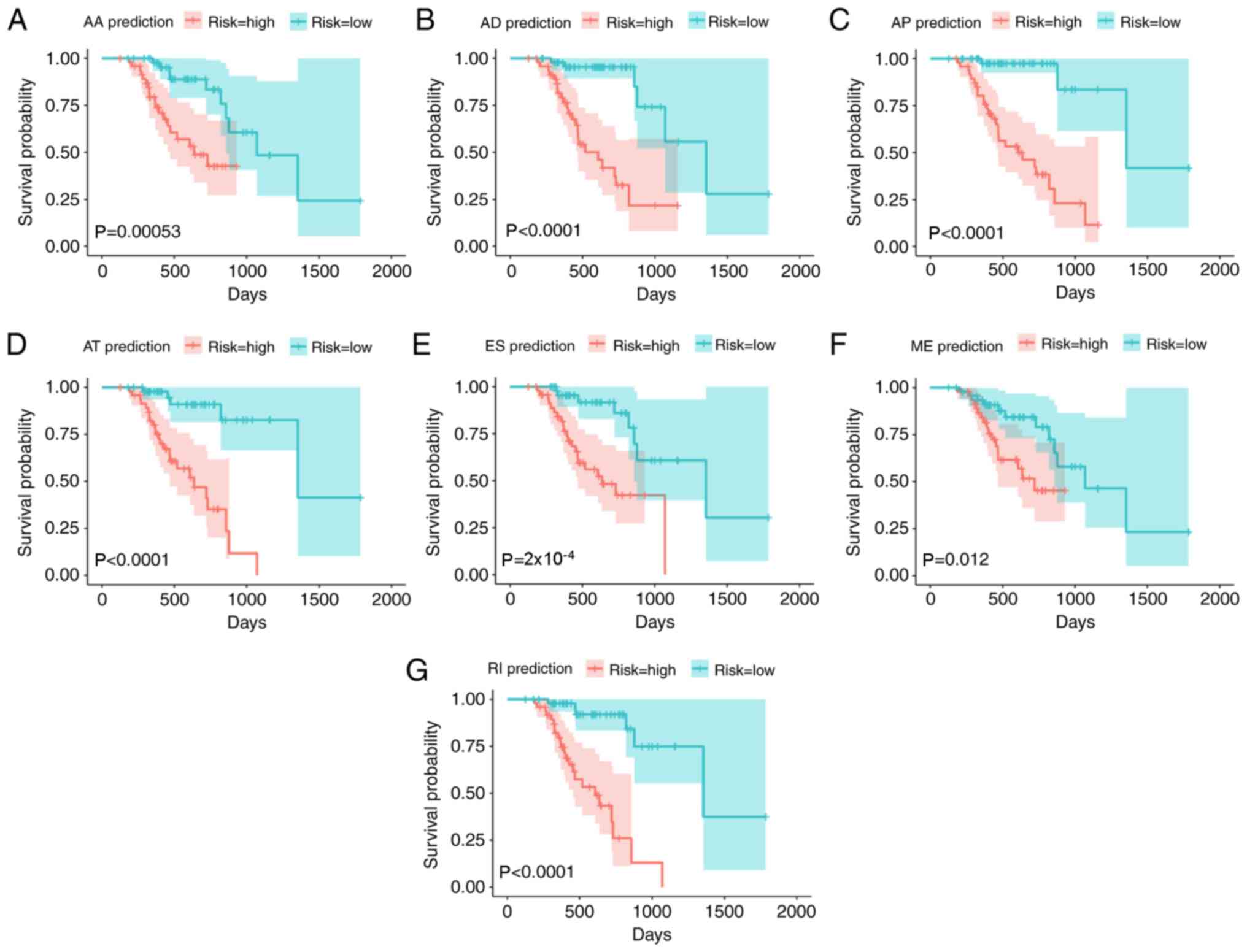 | Figure 4.Kaplan-Meier curves of the prognostic
predictors for patients with melanoma. Kaplan-Meier curves of the
prognostic predictors were generated for each type of alternative
splicing event in melanoma. Kaplan-Meier curves of the prognostic
predictors were generated for (A) AA, (B) AD, (C) AP, (D) AT, (E)
ES, (F) ME and (G) RI. The red line indicates the high-risk group
and the blue line indicates the low-risk group. AA, alternate
acceptor; AD, alternate donor; AP, alternate promoter; AT,
alternate terminator; ES, exon skip; ME, mutually exclusive; RI,
retained intron. |
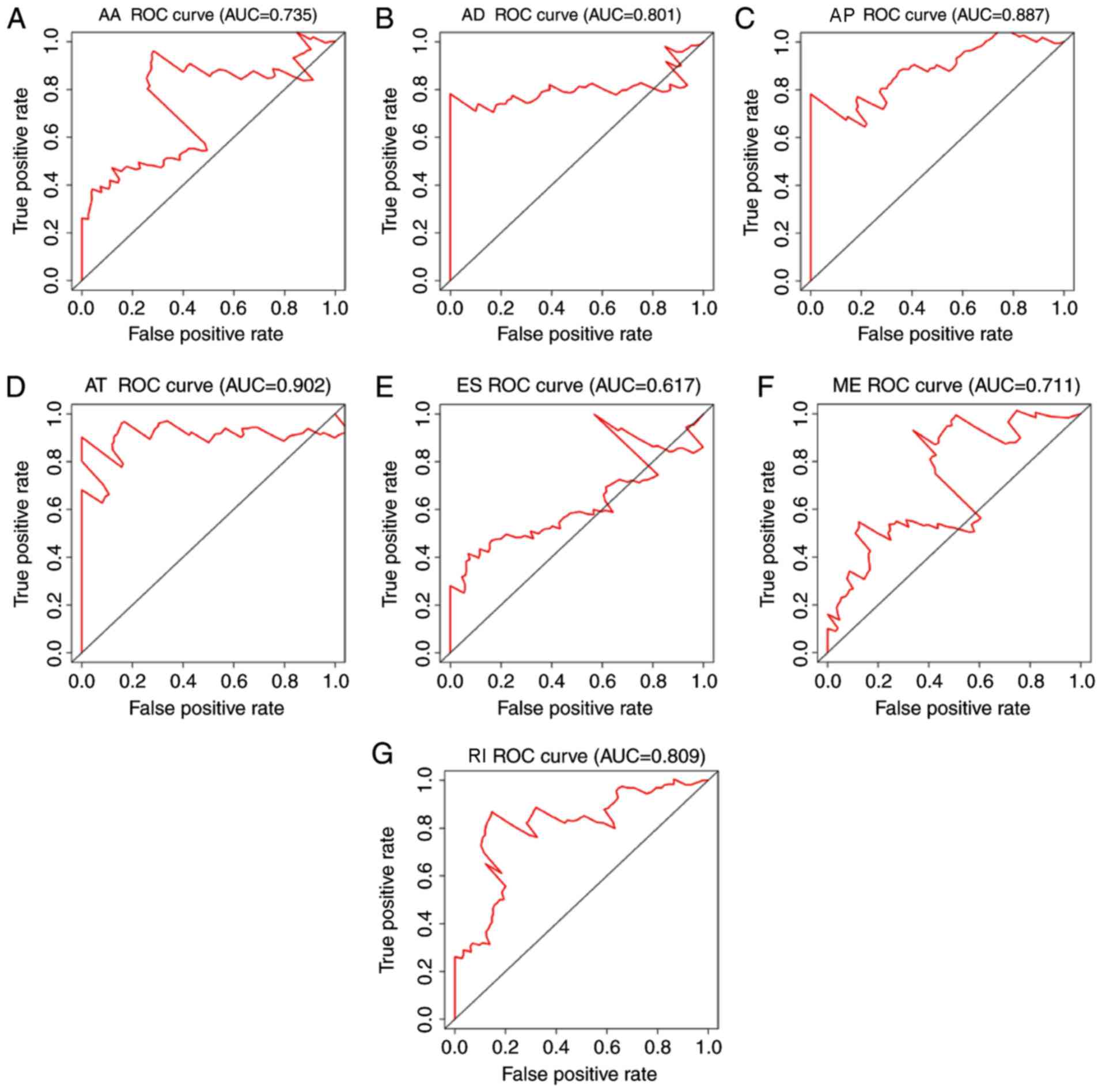 | Figure 6.ROC curves with AUC of the prognostic
predictors for each type of alternative splicing event in melanoma.
ROC curves with AUC of the prognostic predictors for (A) AA, (B)
AD, (C) AP, (D) AT, (E) ES, (F) ME and (G) RI are presented. ROC,
receiver operating characteristic; AUC, area under the curve; ES,
exon skip; AT, alternate terminator; AP, alternate promoter; AA,
alternate acceptor; AD, alternate donor; RI, retained intron; ME,
mutually exclusive. |
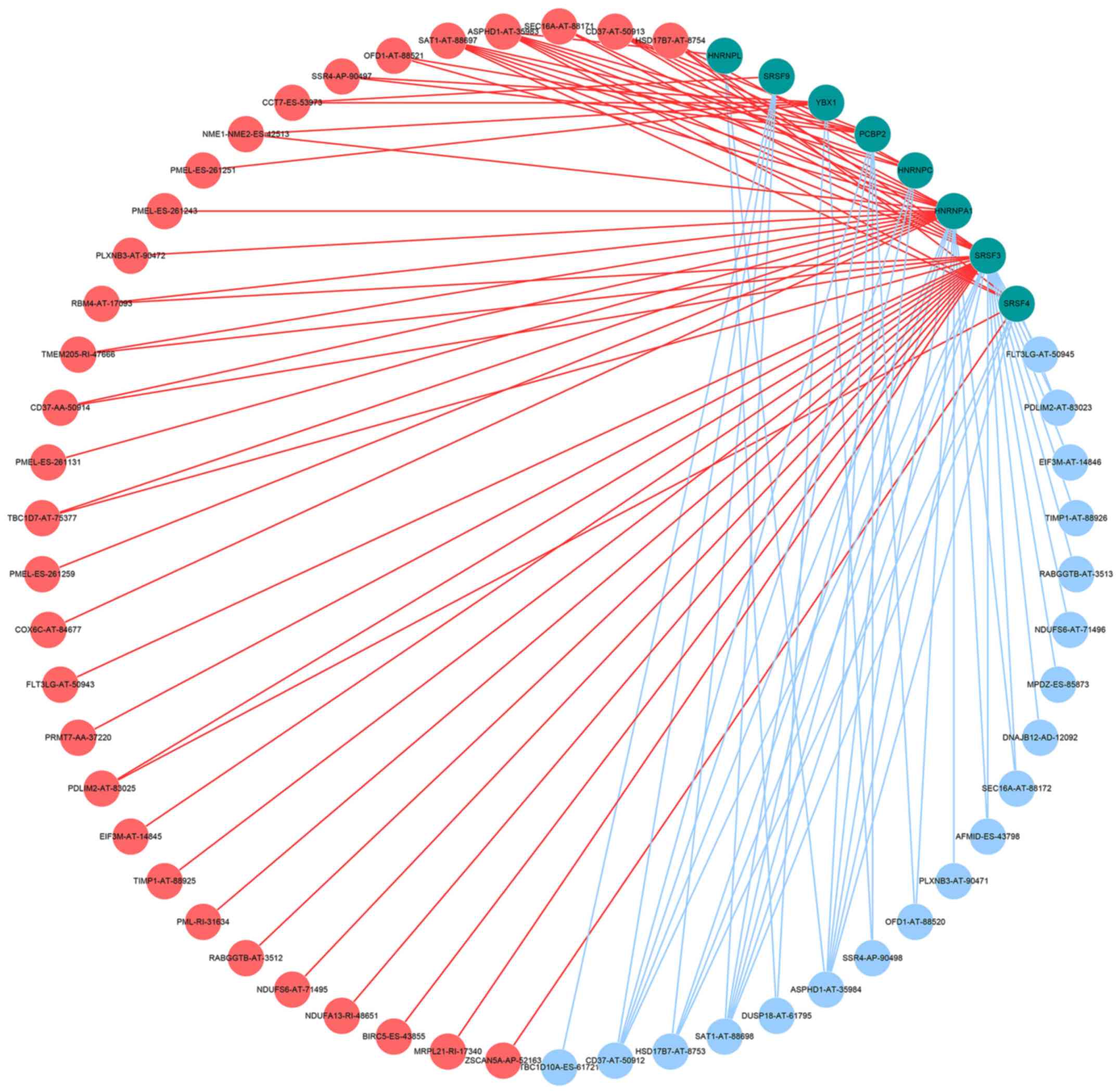
Alternative splicing correlation
network in melanoma
A total of eight splicing factors, heterogeneous
nuclear ribonucleoprotein L, serine and arginine rich splicing
factor 9, Y-box binding protein 1, poly(rC) binding protein 2,
heterogeneous nuclear ribonucleoprotein C (C1/C2), heterogeneous
nuclear ribonucleoprotein A1, serine and arginine rich splicing
factor 3 and serine and arginine rich splicing factor 4, were
identified. As demonstrated in Fig.
8, these splicing factors, represented by green dots, were
markedly associated with the OS time of the patients in the
survival analysis. The expression levels and PSI values of these
eight splicing factors identified from the most significant
alternative splicing events were used to conduct the correlation
analysis in the melanoma cohort based on the significant
correlation results (P<0.05). As presented in Fig. 8, 50 survival-associated alternative
splicing events were identified in the splicing correlation
network, including 31 adverse alternative splicing events (red
dots) and 19 favorable alternative splicing events (blue dots). The
most favorable prognostic alternative splicing events (blue dots)
were positively correlated (blue line) with the expression of the
eight splicing factors (green dots), while the majority of the
adverse prognosis alternative splicing events (red dots) were
negatively correlated (red line) with the expression of the eight
splicing factors.
Discussion
Previous studies have reported several aberrant
alternative splicing events in melanoma (28–31).
Evidence has demonstrated that alternative splicing serves an
important role in the occurrence and development of melanoma. For
example, jumonji domain containing 6, arginine demethylase and
lysine hydroxylase (JMJD6) regulates alternative splicing of p21
(RAC1) activated kinase 1, which is a key component of
mitogen-activated protein kinase signaling; therefore, JMJD6 may
promote carcinogenesis of melanoma (29). Alternative splicing of CD44 mediated
by U2 small nuclear RNA auxiliary factor 2 promotes melanoma
metastasis (30). However, thorough
studies of alternative splicing may provide further information
regarding the oncogenes and tumor suppressor genes of melanoma.
Splicing factor 3b subunit 1 (SF3B1) is a key spliceosome component
that was mutated in melanoma (30),
chronic lymphocytic leukemia (32)
and pancreatic cancer (33).
Mutation of SF3B1 has been associated with good melanoma prognosis
(31). A splicesome of human
telomerase reverse transcriptase may control the activity of
telomerase in melanoma. In the present study, 2,348 alternative
splicing events were markedly associated with the OS time of
patients with melanoma. The results of the present study are
consistent with the findings reported in previous studies (29,30).
These previous studies provide information regarding alternative
splicing and splicing factors in melanoma, which may result in the
identification of potential biomarkers and the development of
therapeutic targets.
The development of high throughput sequencing
technology has provided information regarding the features of
alternative splicing (34,35). It is challenging to identify the
splicesomes associated with cancer that determine the splicing
patterns in the disease. Aberrant patterns of splicing leads to
unlimited proliferation of cancer (36). The motility and invasion of cancer
cells can be enhanced by serine and arginine-rich splicing factor
1, which modifies the expression of Ron-receptor tyrosine kinase
(37). It was identified that
vascular endothelial growth factor (VEGF) leads to angiogenesis in
cancer. VEGF-A165, a splice variant of VEGF, is highly angiogenic;
however, another splice variant of VEGF, VEGF-A165b, is
downregulated in several types of cancer, such as colorectal cancer
(38) and renal cell carcinoma
(39). VEGF-A165b is associated with
a poor prognosis of colorectal cancer (38). Although the role of splice variants
is not completely understood, evidence indicates that they are
strongly associated with specific types of cancer and exhibit
potential prognostic value.
In the present study, alternative splicing features
in 96 patients with melanoma were profiled using integrated
survival analysis. A total of 41,446 alternative splicing events
from 9,780 genes were detected and ~38% were ES events, followed by
AT and AP events. ROC curves and Cox regression models were used to
evaluate alternative splicing patterns as early biomarkers for
melanoma occurrence or prognosis. Among the seven types of
alternative splicing events, the AT events exhibited the highest
efficiency for differentiating good or poor outcomes in patients
with melanoma. The AUC in this model was 0.902, which is the
highest value for all seven types of alternative splicing events,
suggesting that it has the greatest potential for use as a
prognostic marker for melanoma.
In addition, the present analysis highlighted the
potential participation of splicing factors. The majority of the
favorable prognosis alternative splicing events were positively
correlated with the expression of splicing factors and the adverse
prognosis alternative splicing events were negatively correlated
with the expression of splicing factors. Expression of specific
splicing factors may help determine if favorable or adverse
alternative splicing events for prognosis require further
validation by functional experiments. The present study
investigated potential key splicing factors in melanoma and
provided a background for understanding the role of splicesomes in
melanoma.
In conclusion, the present study reported that
alternative splicing events, which are associated with survival,
may be used to generate a prognostic model for risk stratification
in melanoma that could be applied in clinical practice.
Furthermore, the present study provided novel insight into how
pivotal splicing factors regulate aberrant alternative splicing.
From the alternative splicing events and splicing factors that are
associated with survival, several potential biomarkers or
therapeutic targets were identified that require validation in
future studies. Finally, a mechanism for deciphering alternative
splicing events in tumorigenesis in melanoma was proposed.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets analyzed during the study are available
from the corresponding author upon reasonable request.
Authors' contributions
GC and XHH designed the study. FCM, RQH, PL, JCZ, JM
and HY conducted the experiments and analyzed the data. FM wrote
the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cui L, Li Y, Lv X, Li J, Wang X, Lei Z and
Li X: Expression of MicroRNA-301a and its functional roles in
malignant melanoma. Cell Physiol Biochem. 40:230–244. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mahgoub T, Eustace AJ, Collins DM, Walsh
N, O'Donovan N and Crown J: Kinase inhibitor screening identifies
CDK4 as a potential therapeutic target for melanoma. Int J Oncol.
47:900–908. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Finn L, Markovic SN and Joseph RW: Therapy
for metastatic melanoma: The past, present, and future. BMC Med.
10:232012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carpinteiro A, Beckmann N, Seitz A,
Hessler G, Wilker B, Soddemann M, Helfrich I, Edelmann B, Gulbins E
and Becker KA: Role of acid sphingomyelinase-induced signaling in
melanoma cells for hematogenous tumor metastasis. Cell Physiol
Biochem. 38:1–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pagani E, Ruffini F, Antonini Cappellini
GC, Scoppola A, Fortes C, Marchetti P, Graziani G, D'Atri S and
Lacal PM: Placenta growth factor and neuropilin-1 collaborate in
promoting melanoma aggressiveness. Int J Oncol. 48:1581–1589. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Houghton AN and Polsky D: Focus on
melanoma. Cancer Cell. 2:275–278. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Caporali S, Alvino E, Lacal PM, Levati L,
Giurato G, Memoli D, Caprini E, Antonini Cappellini GC and D'Atri
S: Targeting the PI3K/AKT/mTOR pathway overcomes the stimulating
effect of dabrafenib on the invasive behavior of melanoma cells
with acquired resistance to the BRAF inhibitor. Int J Oncol.
49:1164–1174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
David CJ, Chen M, Assanah M, Canoll P and
Manley JL: HnRNP proteins controlled by c-Myc deregulate pyruvate
kinase mRNA splicing in cancer. Nature. 463:364–368. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Shao J, Zhu L, Zhao R, Xing J,
Wang J, Guo X, Tu S, Han B and Yu K: Molecular profiling identifies
prognostic markers of stage IA lung adenocarcinoma. Oncotarget.
8:74846–74855. 2017.PubMed/NCBI
|
|
11
|
Alaterre E, Raimbault S, Goldschmidt H,
Bouhya S, Requirand G, Robert N, Boireau S, Seckinger, Hose D,
Klein B and Moreaux J: CD24, CD27, CD36 and CD302 gene expression
for outcome prediction in patients with multiple myeloma.
Oncotarget. 8:98931–98944. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kelemen O, Convertini P, Zhang Z, Wen Y,
Shen M, Falaleeva M and Stamm S: Function of alternative splicing.
Gene. 344:1–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iqbal SM, Andavan GS and Lemmens-Gruber R:
Differential modulation of fast inactivation in cardiac sodium
channel splice variants by Fyn tyrosine kinase. Cell Physiol
Biochem. 37:825–837. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Avin BA, Umbricht CB and Zeiger MA: Human
telomerase reverse transcriptase regulation by DNA methylation,
transcription factor binding and alternative splicing (Review). Int
J Oncol. 49:2199–2205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bonomi S, Gallo S, Catillo M, Pignataro D,
Biamonti G and Ghigna C: Oncogenic alternative splicing switches:
Role in cancer progression and prospects for therapy. Int J Cell
Biol. 2013:9620382013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fackenthal JD and Godley LA: Aberrant RNA
splicing and its functional consequences in cancer cells. Dis Model
Mech. 1:37–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu Q, Gao J and Li Z: Identification of a
novel alternative splicing transcript variant of the suppressor of
fused: Relationship with lymph node metastasis in pancreatic ductal
adenocarcinoma. Int J Oncol. 49:2611–2619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Izaguirre DI, Zhu W, Hai T, Cheung HC,
Krahe R and Cote GJ: PTBP1-dependent regulation of USP5 alternative
RNA splicing plays a role in glioblastoma tumorigenesis. Mol
Carcinog. 51:895–906. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dhir A and Buratti E: Alternative
splicing: Role of pseudoexons in human disease and potential
therapeutic strategies. FEBS J. 277:841–855. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karni R, de Stanchina E, Lowe SW, Sinha R,
Mu D and Krainer AR: The gene encoding the splicing factor SF2/ASF
is a proto-oncogene. Nat Struct Mol Biol. 14:185–193. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Sun N, Lu Z, Sun S, Huang J, Chen Z
and He J: Prognostic alternative mRNA splicing signature in
non-small cell lung cancer. Cancer Lett. 393:40–51. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu J, Chen Z and Yong L: Systematic
profiling of alternative splicing signature reveals prognostic
predictor for ovarian cancer. Gynecol Oncol. 148:368–374. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Suo C, Hrydziuszko O, Lee D, Pramana S,
Saputra D, Joshi H, Calza S and Pawitan Y: Integration of somatic
mutation, expression and functional data reveals potential driver
genes predictive of breast cancer survival. Bioinformatics.
31:2607–2613. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pal S, Bi Y, Macyszyn L, Showe LC,
O'Rourke DM and Davuluri RV: Isoform-level gene signature improves
prognostic stratification and accurately classifies glioblastoma
subtypes. Nucleic Acids Res. 42:e642014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Robertson AG, Shih J, Yau C, Gibb EA, Oba
J, Mungall KL, Hess JM, Uzunangelov V, Walter V, Danilova L, et al:
Integrative analysis identifies four molecular and clinical subsets
in uveal melanoma. Cancer Cell. 32:204–220.e15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ryan MC, Cleland J, Kim R, Wong WC and
Weinstein JN: SpliceSeq: A resource for analysis and visualization
of RNA-Seq data on alternative splicing and its functional impacts.
Bioinformatics. 28:2385–2387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lex A, Gehlenborg N, Strobelt H, Vuillemot
R and Pfister H: UpSet: Visualization of intersecting sets. IEEE
Trans Vis Comput Graph. 20:1983–1992. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu G, Feng X and Stein L: A human
functional protein interaction network and its application to
cancer data analysis. Genome Biol. 11:R532010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu X, Si W, Liu X, He L, Ren J, Yang Z,
Yang J, Li W, Liu S, Pei F, et al: JMJD6 promotes melanoma
carcinogenesis through regulation of the alternative splicing of
PAK1, a key MAPK signaling component. Mol Cancer. 16:1752017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang P, Feng S, Liu G, Wang H, Fu A, Zhu
H, Ren Q, Wang B, Xu X, Bai H and Dong C: CD82 suppresses CD44
alternative splicing-dependent melanoma metastasis by mediating
U2AF2 ubiquitination and degradation. Oncogene. 35:5056–5069. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Furney SJ, Pedersen M, Gentien D, Dumont
AG, Rapinat A, Desjardins L, Turajlic S, Piperno-Neumann S, de la
Grange P, Roman-Roman S, et al: SF3B1 mutations are associated with
alternative splicing in uveal melanoma. Cancer Discov. 3:1122–1129.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Quesada V, Conde L, Villamor N, Ordóñez
GR, Jares P, Bassaganyas L, Ramsay AJ, Beà S, Pinyol M,
Martínez-Trillos A, et al: Exome sequencing identifies recurrent
mutations of the splicing factor SF3B1 gene in chronic lymphocytic
leukemia. Nat Genet. 44:47–52. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Biankin AV, Waddell N, Kassahn KS, Gingras
MC, Muthuswamy LB, Johns AL, Miller DK, Wilson PJ, Patch AM, Wu J,
et al: Pancreatic cancer genomes reveal aberrations in axon
guidance pathway genes. Nature. 491:399–405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan Q, Shai O, Lee LJ, Frey BJ and
Blencowe BJ: Deep surveying of alternative splicing complexity in
the human transcriptome by high-throughput sequencing. Nat Genet.
40:1413–1415. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baty F, Klingbiel D, Zappa F and Brutsche
M: High-throughput alternative splicing detection using dually
constrained correspondence analysis (DCCA). J Biomed Inform.
58:175–185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Oltean S and Bates DO: Hallmarks of
alternative splicing in cancer. Oncogene. 33:5311–5318. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Das S and Krainer AR: Emerging functions
of SRSF1, splicing factor and oncoprotein, in RNA metabolism and
cancer. Mol Cancer Res. 12:1195–1204. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Díaz R, Peña C, Silva J, Lorenzo Y, García
V, García JM, Sánchez A, Espinosa P, Yuste R, Bonilla F and
Domínguez G: p73 isoforms affect VEGF, VEGF165b and PEDF expression
in human colorectal tumours. VEGF165b downregulation as a marker
for poor prognosis. Int J Cancer. 123:1060–1067. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bates DO, Cui TG, Doughty JM, Winkler M,
Sugiono M, Shields JD, Peat D, Gillatt D and Harper SJ: VEGF165b,
an inhibitory splice variant of vascular endothelial growth factor,
is down-regulated in renal cell carcinoma. Cancer Res.
62:4123–4131. 2002.PubMed/NCBI
|