Introduction
The endocannabinoid system, including the seven
transmembrane cannabinoid receptors 1 and 2 (CB1 and CB2) and their
endogenous ligands, is involved in a wide variety of
pathophysiological situations such as inflammatory and oncogenic
processes (1–3). Endogenous and exogenous cannabinoids
are known to exert anti-inflammatory and anti-tumor effects
(1,4). Recently, we reported that hypoxic
stress modifies the endocannabinoid system to promote malignancy in
glioblastoma cells through downregulation of the CB1 receptor and
upregulation of cyclooxygenase-2 (COX2) (5), the enzyme responsible for metabolizing
endocannabinoids (4). This result
indicates that hypoxic stress aggravates cancer by inhibiting the
endocannabinoid system.
Emotional and physical stressors typically induce
the release of adrenal corticosteroid hormones such as
corticosterone and cortisol in rodents and human, respectively
(6). Corticosteroid hormones inhibit
immune responses by downregulating the activity of nuclear
factor-κB (NF-κB) signaling (7) and
the production of several cytokines, such as tumor necrosis
factor-α or interleukin (7).
Interestingly, emotional and physical stresses have been reported
to impair the endocannabinoid system concomitant with an increase
in serum corticosteroids (8).
In the present study, we examined the effects of
corticosterone, which is one of the corticosteroids (6), on the endocannabinoid system in
malignant glioblastoma cells. We found that corticosterone inhibits
the expression of the genes encoding CB1 and attenuates cannabinoid
receptor agonist (WIN 55,212-2)-induced decrease in glioblastoma
cell viability. Our findings reveal the mechanisms by which stress
hormones inhibit the endocannabinoid system through downregulation
of the CB1 receptor, thus reducing the beneficial anti-tumor
effects of endocannabinoids and consequently promoting the
malignant phenotype of glioblastoma.
Materials and methods
Chemicals
Corticosterone (CORT), WIN 55,212-2, and Dulbecco's
modified Eagle's medium were obtained from Wako Pure Chemical
Industries (Osaka, Japan). Fetal bovine serum was obtained from
Invitrogen; Thermo Fisher Scientific, Inc., (Waltham, MA, USA).
Anti-cannabinoid receptor 1 (CB1) rabbit antibody (ab137410) and
anti-glucocorticoid receptor (GR) (EPR4595) rabbit antibody (cat.
no. ab109022) were obtained from Abcam (Tokyo, Japan). Anti-GAPDH
rabbit antibody (cat. no. 2118) and horseradish
peroxidase-conjugated anti-rabbit IgG (cat. no. 7074) were
purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA).
Anti-COX2 antibody (cat. no. 160106) was obtained from Cayman
Chemical Company (Ann Arbor, MI, USA).
Cell culture
U-87 MG malignant glioblastoma cells (glioblastoma
of unknown origin; ATCC® HTB-14™) were provided by Prof.
Nakata (Kanazawa University, Kanazawa, Japan). U-87 MG cells were
maintained in Dulbecco's modified Eagle's medium containing 10%
fetal bovine serum at 37°C and 5% CO2.
Cell viability assay
Cell viability was analyzed using the Cell Counting
Kit-8 (Wako Pure Chemical Industries) as previously described
(5,9). U-87 MG cells were seeded into 96-well
plates at a density of 1×102 cells/well. After a 24-h
incubation period, the cells were stimulated with several
concentrations of CORT. On day 2 after stimulation, the cells were
incubated with WST-8 (10 µl of WST-8 in 100 µl of medium) for 3 h
at 37°C. The absorbance of the colored formazan product, generated
by mitochondrial dehydrogenases in metabolically active cells, was
recorded at 450 nm. Cell viability was expressed as a ratio of the
absorbance in treated wells relative to that in untreated control
wells.
Western blotting analysis
Western blotting was performed as described
previously (10). Briefly, proteins
were extracted from cells, and protein concentrations were
determined by a protein assay. Equal amounts of protein (30 µg)
were separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The resolved proteins were transferred onto
polyvinylidene fluoride membranes, which were incubated with
primary antibodies (1:1,000), followed by incubation with
horseradish peroxidase-linked secondary antibodies (1:2,000).
RT-qPCR
To evaluate the expression of human glucocorticoid
receptor (GR: NR3C1), human COX2 (PTGS2), and human
CB1 (CNR1) and human β-actin (ACTB) mRNAs in the
cells, Real time PCR was performed as follows. Briefly, RNA was
extracted from the cells, and cDNA was generated using reverse
transcriptase ReverTra Ace® (Toyobo, Tokyo, Japan). We
performed PCR-based, subtype-specific gene amplification with
QuantiTect SYBR Green PCR kit (Qiagen, Hilden, Germany) using the
sets of primers as follows: 5′-GGCTAGACACCCATTTTCACA-3′ and
5′-ACAGAAAGGGCTACTACAGCT-3′ for GR (NR3C1),
5′-CAGCAAATCCTTGCTGTTCC-3′ and 5′-GTGAAGTGCTGGGCAAAGAAT-3′ for
COX-2 (PTGS2), 5′-CAGGCCTTCCTACCACTTCAT-3′ and
5′-ACCCCACCCAGTTTGAACAGA-3′ for CB1 (CNR1), and
5′-ATGGTGGGCATGGGTCAGAA-3′ and 5′-CTGGGGTGTTGAAGGTCTCAA-3′ for
β-actin (ACTB). The quantified value of each sample was
normalized with that of the β-actin value of the same sample, which
was amplified simultaneously with the target gene.
Cannabinoid receptor agonist
treatment
The cells were treated with the cannabinoid receptor
agonist WIN 55,212-2 following 2-day exposure to corticosterone.
After 2 days of CORT treatment, cell viability was analyzed using
the Cell Counting Kit-8 (Wako Pure Chemical Industries) as
described above.
Statistical analysis
Data are presented as the mean ± standard error of
the mean (SEM) from at least three independent experiments. The
Stat View statistical package (version 5.0 Japanese version,
Hulinks, Japan) was used for statistical analysis. Student's t-test
was used to examine the differences between the two groups (with or
without CORT stimulation) in the real time PCR and western blot
analyses. A one-way analysis of variance (ANOVA) was used to
analyze the cell viability data. P<0.05 was considered to
indicate a statistically significant difference.
Results
Corticosterone does not affect cell
viability in U-87 MG cells
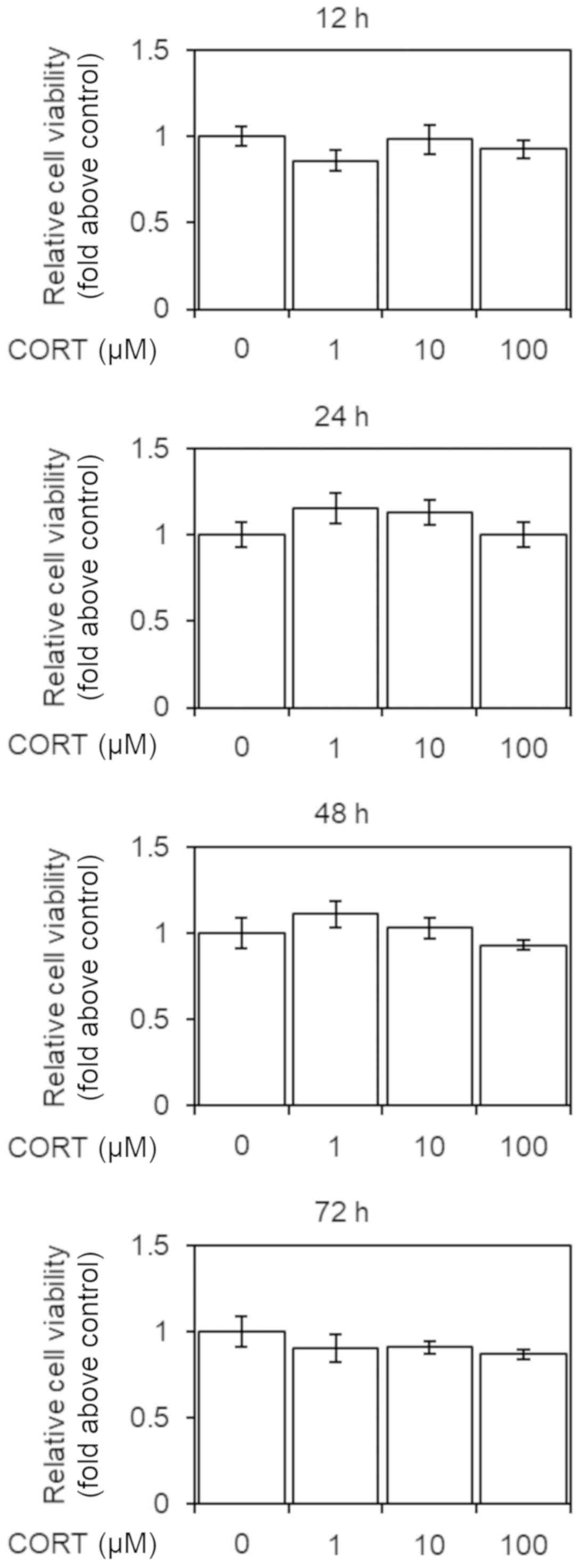
We investigated the effect of 12, 24, 48, or 72-h
treatment with CORT (1–100 µM) on cell viability in U-87 MG cells.
Cell viability was not significantly altered by CORT (Fig. 1).
Corticosterone did not affect the
levels of GR mRNA and protein in U-87 MG cells
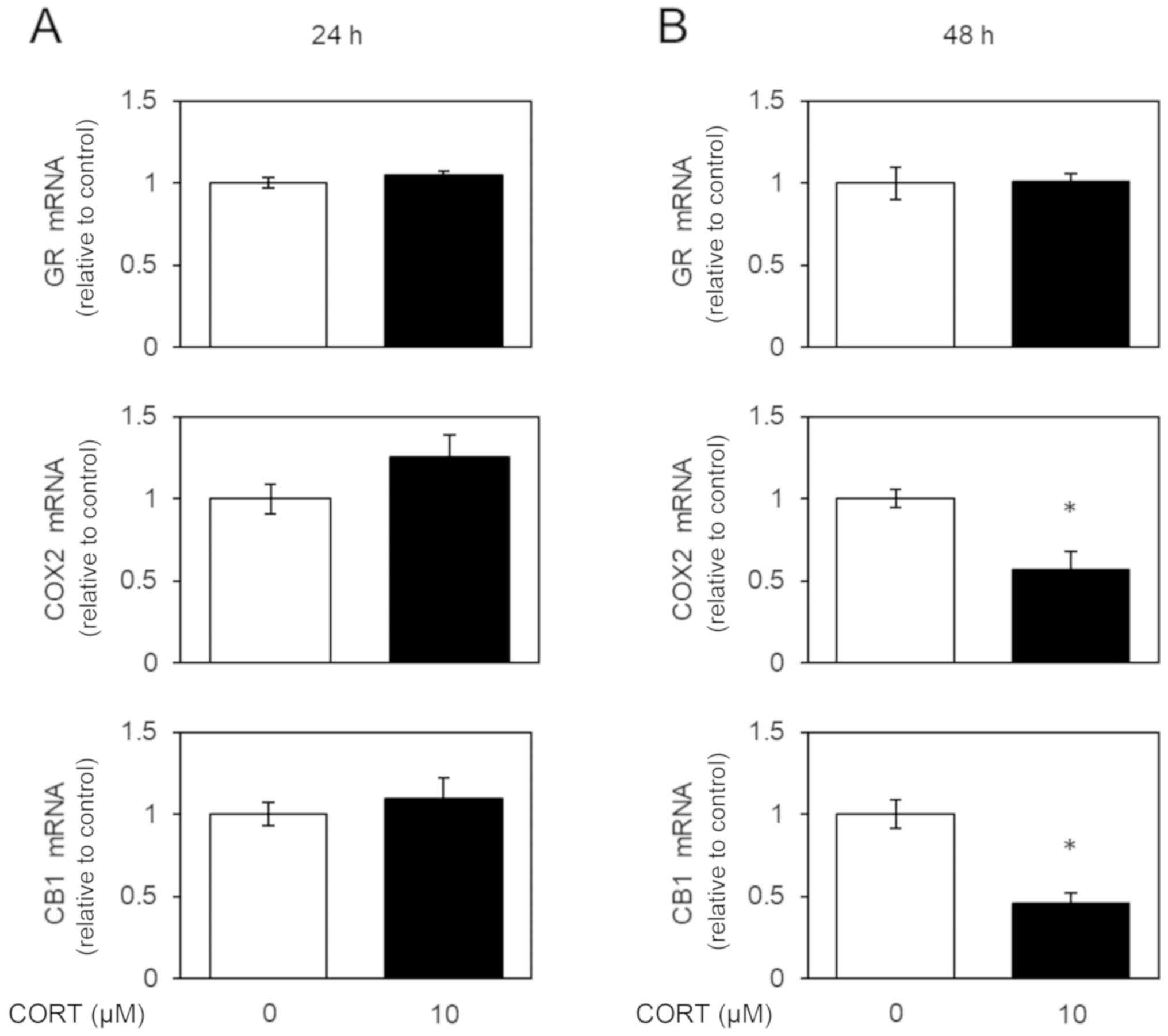
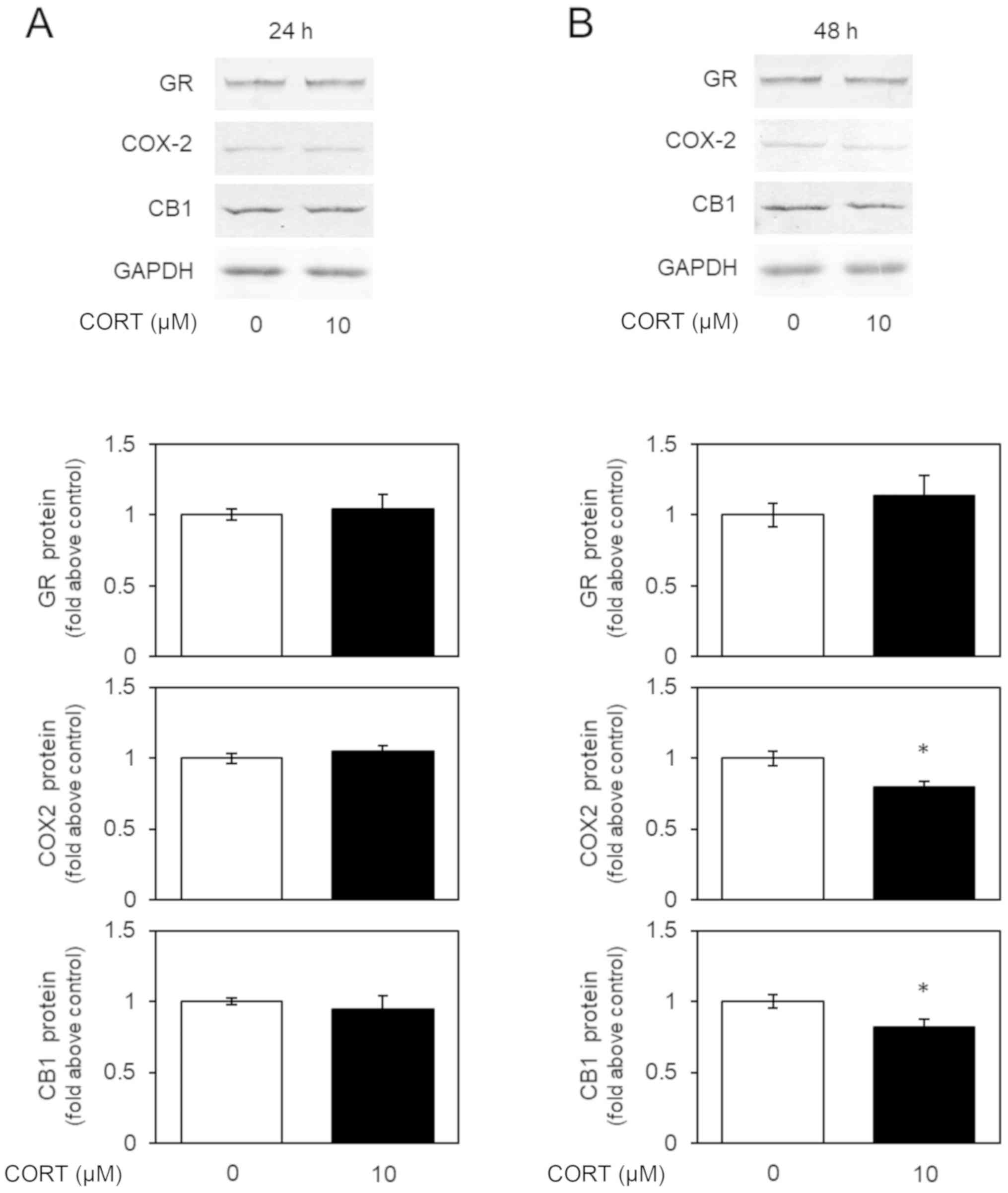
Next, we examined the expression of glucocorticoid
receptor (GR) mRNA and protein in U-87 MG cells by real-time PCR
and western blotting, respectively. GR is one of main receptors for
CORT (6). GR was spontaneously
detectable and the levels of GR mRNA and protein were did not
changed by CORT stimulation in U-87 MG cells (Figs. 2 and 3). CORT may act transduction through GR
receptor in U-87 MG cells.
Corticosterone decreases the levels of
COX2 mRNA and protein in U-87 MG cells
We examined the expression of COX2 mRNA and protein
in U-87 MG cells by real-time PCR and western blotting,
respectively. Previously, COX2 expression was shown to be regulated
by NF-κB signaling (7). COX2 was
spontaneously detectable in U-87 MG cells (5), while 48-h, but not 24-h, treatment with
CORT decreased the expression of mRNA encoding COX2 (Fig. 2) and COX2 protein (Fig. 3), indicating the inhibition of NF-κB
signaling by CORT.
Corticosterone decreases the levels of
CB1 mRNA and protein in U-87 MG cells
We examined the expression of CB1 mRNA and protein
in U-87 MG cells by real-time PCR and western blotting,
respectively. The cannabinoid receptors facilitate the transduction
of extracellular signals to the cytoplasm (2). CB1 was spontaneously detectable in U-87
MG cells (5), while 48-h, but not
24-h, treatment with CORT decreased the expression of mRNA encoding
CB1 (Fig. 2) and CB1 protein
(Fig. 3). These results indicate
that CORT regulates the cannabinoid system by modulating CB1
expression.
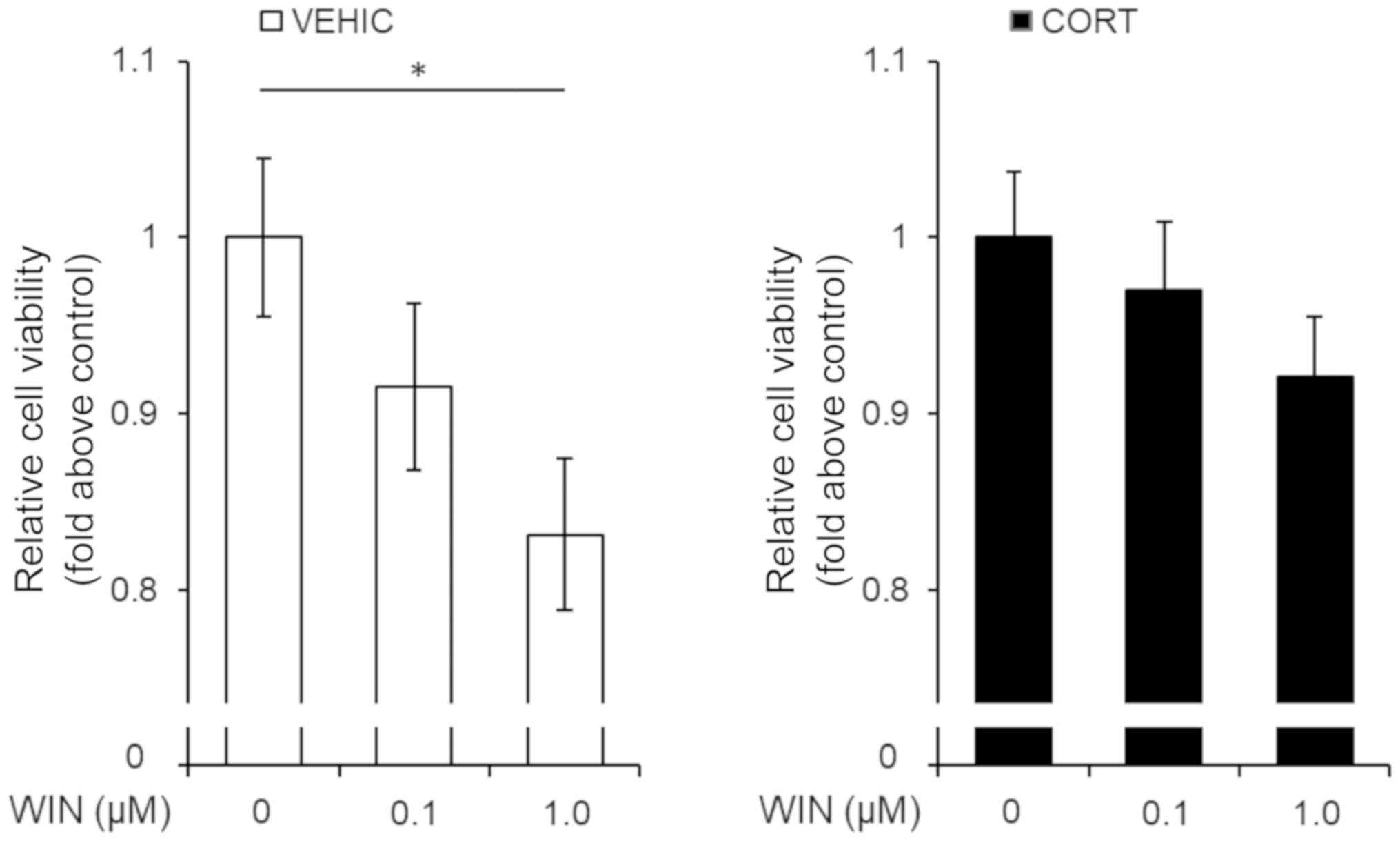
Corticosterone attenuates the
cannabinoid receptor agonist-induced decrease in cell viability in
U-87 MG cells
Recently, cannabinoid receptor agonists were shown
to mimic the effects of endogenous cannabinoids and cause cell
death in glioblastoma (5,11,12). The
observed corticosterone-induced downregulation of cannabinoid
receptor expression; thus, we investigated the effects of 48 h
treatment with CORT on cannabinoid receptor agonist-induced
decrease in glioblastoma cell viability. We treated the cells with
WIN 55,212-2 (0.1 or 1.0 µM) after 48 h treatment with CORT. We
analyzed cell viability 48 h after treatment with WIN 55,212-2. WIN
55,212-2 treatment significantly decreased the viability of
vehicle-treated cells in a dose-dependent manner (Fig. 4). However, the doses of WIN 55,212-2
(0.1 or 1.0 µM) failed to decrease cell viability in the CORT
treatment group (Fig. 4). These
results suggest that CORT confers tolerance to cannabinoid receptor
agonist-induced cellular toxicity in glioblastoma.
Discussion
Emotional and physical stress have been suggested to
affect the development and progression of malignancies (13). Interestingly, emotional and physical
stressors release corticosteroids and, in turn, impair the
endocannabinoid system (8). We
previously showed that hypoxia aggravates cancer by inhibiting the
endocannabinoid system in malignant glioblastoma cells (5). Thus, dysfunction of the endocannabinoid
system by stress-released corticosteroids may lead to the
development and progression of malignancies. However, the effects
of corticosteroids on endocannabinoid system-dependent malignancy
are not fully understood. Endocannabinoids have been shown to
influence diverse pathological processes, including malignancy
(2,14,15).
Endogenous and exogenous cannabinoids have anti-cancer effects;
exogenous cannabinoids and many synthetic agonists for cannabinoid
receptors, which mimic the effects of endocannabinoids, have been
proposed as potential anti-cancer agents (14,16–20).
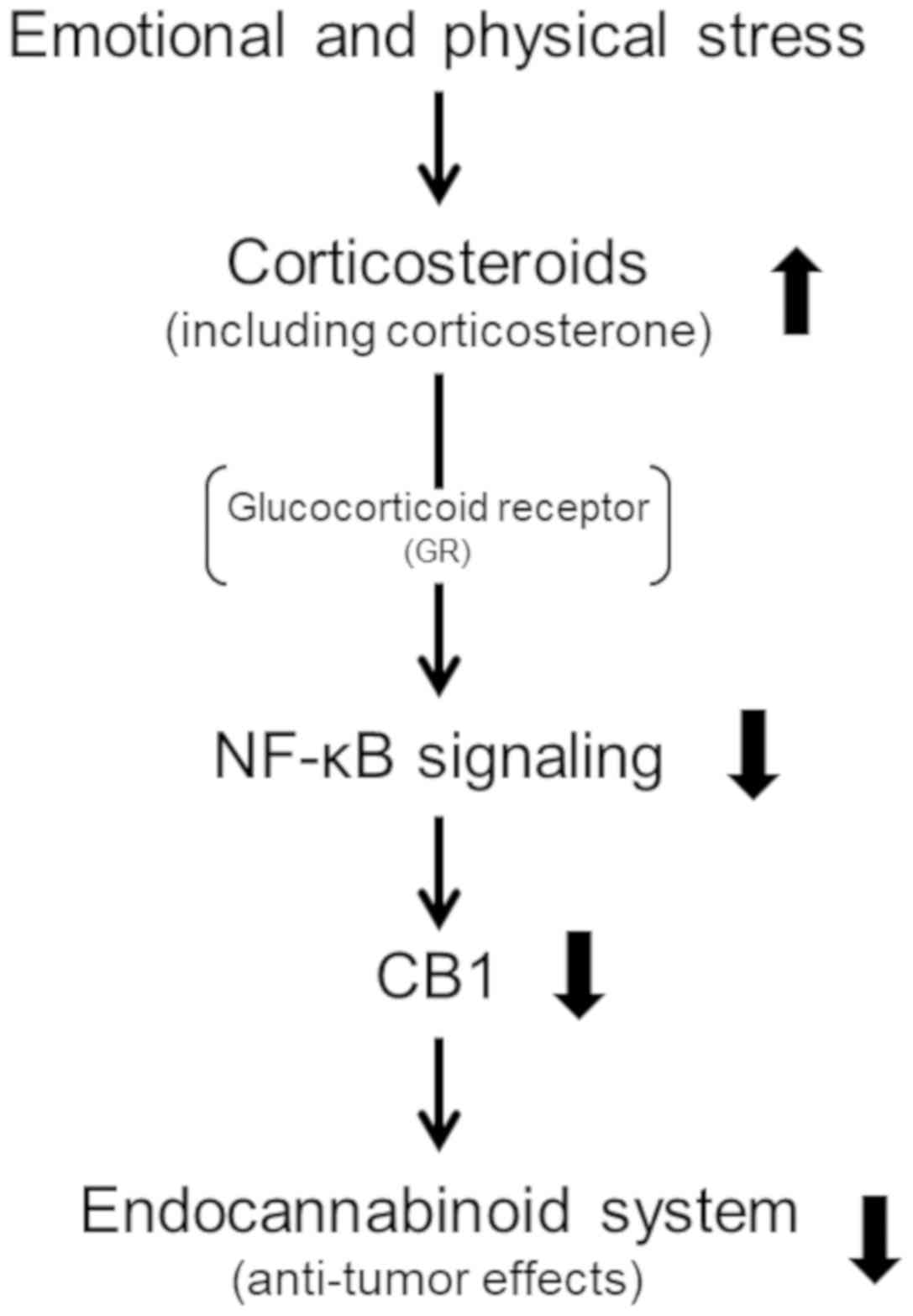
Here, we found that corticosteroids downregulate both mRNA and
protein of CB1 and attenuate cannabinoid receptor agonist-induced
decrease in U-87 MG cell viability. Our results suggest that
stress-released corticosteroids promote the progression of
glioblastoma by inhibiting the endocannabinoid system (Fig. 5).
Corticosterone is considered an adrenal
corticosteroid and functions as both a glucocorticoid and
mineralocorticoid (6).
Glucocorticoids are known to have anti-inflammatory and
immunosuppressive potentials through inhibition of NF-κB activities
(7,21,22).
NF-κB signaling is involved in controlling the expression of COX2
(7). We showed that corticosterone
decreases the levels of COX2 mRNA and protein (Figs. 2 and 3) in this study, indicating that
corticosterone might downregulate NF-κB signaling. Interestingly,
inhibition of NF-κB signaling induced downregulation of CB1
receptor expression (23). Thus,
inhibition of NF-κB signaling by corticosterone may downregulate
the expression of the CB1 receptor (Figs. 2 and 3).
Nakatani et al (24), showed that corticosterone inhibited
the proliferation of C6 glioma cells. However, in our present
study, corticosterone did not inhibit the cell viability in U-87 MG
cells. Since U-87 MG cells are glioblastoma, a poorly
differentiated type of glioma, highly malignant and exhibits
aggressive invasive growth in comparison with glioma,
corticosterone might not be able to inhibit the cell viability in
U-87 MG cells.
Corticosterone alone downregulates the expression of
CB1 (Figs. 2 and 3), but does not affect cell viability in
12, 24, 48, and 72 h after stimulation (Fig. 1). These results suggest that CB1
might not be committed to cell viability directly in U-87 cells.
However, as shown in our previous study that hypoxia decreased the
expression of CB1 in U-87 MG cells, the cannabinoid receptor
agonist WIN 55,212-2 decreased glioblastoma cell viability, with
the extent depending on the expression level of the CB1 receptor
(5). In the present study, we showed
that corticosterone downregulated the expression of CB1 and
suppressed WIN 55,212-2-induced decrease in U-87 MG cell viability.
Endocannabinoids in the brain (25)
have the potential to prevent brain tumor development by activating
cannabinoid receptors. Thus, our results suggest that
corticosterone may counteract the beneficial anti-tumor effects of
endocannabinoids by downregulating cannabinoid receptor expression
in the brain.
The present study demonstrated that corticosterone
downregulates CB1 receptors and suppresses cannabinoid receptor
agonist-induced decrease in U-87 MG cell viability in vitro.
These results suggest that corticosterone promotes brain tumor
progression by inhibiting endocannabinoid receptor signaling.
Acknowledgements
Not applicable.
Funding
The present study was supported by Grants-in-Aid for
Science and Culture (grant nos. 26430013, 16H005513 and 17H01963)
from the Ministry of Education, Culture, Sports, Science, and
Technology of Japan.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
NS designed the study and prepared the manuscript.
NS, HI, and TS conducted the experiments. NS, YU, HN and AY
analyzed and interpreted the data. NS, HI, and TS obtained funding.
AY and TS edited the manuscript. All authors read and approved the
final version of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of
interest.
Glossary
Abbreviations
Abbreviations:
|
CB
|
cannabinoid receptor
|
|
NF-κB
|
nuclear factor-κB
|
|
COX2
|
cyclooxygenase-2
|
|
CORT
|
corticosterone
|
References
|
1
|
Gui H, Tong Q, Qu W, Mao CM and Dai SM:
The endocannabinoid system and its therapeutic implications in
rheumatoid arthritis. Int Immunopharmacol. 26:86–91. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kano M, Ohno-Shosaku T, Hashimotodani Y,
Uchigashima M and Watanabe M: Endocannabinoid-mediated control of
synaptic transmission. Physiol Rev. 89:309–380. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ligresti A, De Petrocellis L and Di Marzo
V: From phytocannabinoids to cannabinoid receptors and
endocannabinoids: Pleiotropic physiological and pathological roles
through complex pharmacology. Physiol Rev. 96:1593–1659. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cabral GA and Griffin-Thomas L:
Cannabinoids as therapeutic agents for ablating neuroinflammatory
disease. Endocr Metab Immune Disord Drug Targets. 8:159–172. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sugimoto N, Ishibashi H, Nakamura H,
Yachie A and Ohno-Shosaku T: Hypoxia-induced inhibition of the
endocannabinoid system in glioblastoma cells. Oncol Rep.
38:3702–3708. 2017.PubMed/NCBI
|
|
6
|
Joëls M: Corticosteroids and the brain. J
Endocrinol. 238:R121–R130. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brattsand R and Linden M: Cytokine
modulation by glucocorticoids: Mechanisms and actions in cellular
studies. Aliment Pharmacol Ther. 10 (Suppl 2):S81–S92. 1996.
View Article : Google Scholar
|
|
8
|
Wamsteeker JI, Kuzmiski JB and Bains JS:
Repeated stress impairs endocannabinoid signaling in the
paraventricular nucleus of the hypothalamus. J Neurosci.
30:11188–11196. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leu H, Sugimoto N, Shimizu M, Toma T, Wada
T, Ohta K and Yachie A: Tumor necrosis factor-α modifies the
effects of Shiga toxin on glial cells. Int Immunopharmacol.
38:139–143. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sugimoto N, Miwa S, Nakamura H, Tsuchiya H
and Yachie A: Protein kinase A and Epac activation by cAMP
regulates the expression of glial fibrillary acidic protein in
glial cells. Arch Biol Sci. 68:795–801. 2016. View Article : Google Scholar
|
|
11
|
Solinas G and Becattini B: JNK at the
crossroad of obesity, insulin resistance, and cell stress response.
Mol Metab. 6:174–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ortega A, Rangel-López E, Hidalgo-Miranda
A, Morales A, Ruiz-García E, Meneses-García A, Herrera-Gómez A,
Aguilar-Ponce JL, González-Herrera IG, Guevara-Salazar P, et al: On
the effects of CP 55–940 and other cannabinoid receptor agonists in
C6 and U373 cell lines. Toxicol In Vitro. 29:1941–1951. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moreno-Smith M, Lutgendorf SK and Sood AK:
Impact of stress on cancer metastasis. Future Oncol. 6:1863–1881.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guindon J and Hohmann AG: The
endocannabinoid system and cancer: Therapeutic implication. Br J
Pharmacol. 163:1447–1463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klein TW: Cannabinoid-based drugs as
anti-inflammatory therapeutics. Nat Rev Immunol. 5:400–411. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Echigo R, Sugimoto N, Yachie A and
Ohno-Shosaku T: Cannabinoids inhibit peptidoglycan-induced
phosphorylation of NF-κB and cell growth in U87MG human malignant
glioma cells. Oncol Rep. 28:1176–1180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gurley SN, Abidi AH, Allison P, Guan P,
Duntsch C, Robertson JH, Kosanke SD, Keir ST, Bigner DD, Elberger
AJ and Moore BM II: Mechanism of anti-glioma activity and in vivo
efficacy of the cannabinoid ligand KM-233. J Neurooncol.
110:163–177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guzmán M: Cannabinoids: Potential
anticancer agents. Nat Rev Cancer. 3:745–755. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salazar M, Carracedo A, Salanueva IJ,
Hernández-Tiedra S, Lorente M, Egia A, Vázquez P, Blázquez C,
Torres S, García S, et al: Cannabinoid action induces
autophagy-mediated cell death through stimulation of ER stress in
human glioma cells. J Clin Invest. 119:1359–1372. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sarfaraz S, Adhami VM, Syed DN, Afaq F and
Mukhtar H: Cannabinoids for cancer treatment: Progress and promise.
Cancer Res. 68:339–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jang BC, Lim KJ, Suh MH, Park JG and Suh
SI: Dexamethasone suppresses interleukin-1beta-induced human
beta-defensin 2 mRNA expression: involvement of p38 MAPK, JNK,
MKP-1, and NF-kappaB transcriptional factor in A549 cells. FEMS
Immunol Med Microbiol. 51:171–184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chantong B, Kratschmar DV, Nashev LG,
Balazs Z and Odermatt A: Mineralocorticoid and glucocorticoid
receptors differentially regulate NF-kappaB activity and
pro-inflammatory cytokine production in murine BV-2 microglial
cells. J Neuroinflammation. 9:2602012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang SS, Chen DZ, Wu H, Chen RC, Du SJ,
Dong JJ, Liang G, Xu LM, Wang XD, Yang YP, et al: Cannabinoid
receptors are involved in the protective effect of a novel curcumin
derivative C66 against CCl4-induced liver fibrosis. Eur J
Pharmacol. 779:22–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakatani Y, Amano T and Takeda H:
Corticosterone inhibits the proliferation of C6 glioma cells via
the translocation of unphosphorylated glucocorticoid receptor. Biol
Pharm Bull. 39:1121–1129. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zoerner AA, Gutzki FM, Batkai S, May M,
Rakers C, Engeli S, Jordan J and Tsikas D: Quantification of
endocannabinoids in biological systems by chromatography and mass
spectrometry: A comprehensive review from an analytical and
biological perspective. Biochim Biophys Acta. 1811:706–723. 2011.
View Article : Google Scholar : PubMed/NCBI
|