Introduction
Prostate cancer (PCa) is the most commonly diagnosed
malignancy and the second most lethal cancer in men. In 2016, it
was estimated that 180,890 new cases of PCa were diagnosed, leading
to 26,120 deaths worldwide (1).
MicroRNAs (miRNAs/miRs) are endogenous non-coding
RNAs, 18–22 nucleotides in length, which bind to the
3′-untranslated region (3′UTR) of target genes and regulate
translational repression and/or mRNA degradation (2). miRNAs serve important roles in various
biological and pathological processes, including cell development,
infection, immunity and carcinogenesis (3,4).
Numerous studies have demonstrated that miRNAs may function as
oncogenes and/or tumor suppressors (5,6), and
that their abnormal expression levels are closely associated with
the development and progression of various types of cancer
(7).
miR-381 serves as a tumor suppressor in several
types of cancer, including PCa (8,9), colon
cancer (10), hepatocellular
carcinoma (11), lung adenocarcinoma
(12), gastric cancer (13) and breast cancer (14). It has been reported that miR-381
inhibits cell proliferation, migration and invasion in epithelial
ovarian cancer (15); however, Tang
et al (16) demonstrated that
miR-381 increases the proliferation of glioma cells in vitro
and in vivo, and that this action is associated with
decreased activation of extracellular signal-regulated kinase and
protein kinase B signaling. Moreover, miR-381 sensitizes renal
cancer cells to the cytotoxic action of 5-fluorouracil via
inhibition of WEE1 G2 checkpoint kinase expression levels and
upregulation of cell division cycle 2 in 786-O cells (17). Collectively, these data suggested
that miR-381 serves a complex role in tumorigenesis; however, the
underlying mechanism by which miR-381 contributes to PCa
tumorigenesis has not been fully elucidated.
The present study aimed to investigate the
expression pattern and potential biological function of miR-381 in
PCa cells. The results may provide novel insights into the
molecular mechanism underlying prostate tumorigenesis and may
reveal a potential therapeutic strategy for PCa.
Materials and methods
Cell culture
The PCa cell line LNCaP and the normal prostate
epithelial cell line PrEC were purchased from the American Type
Culture Collection (Manassas, VA, USA). Cells were cultured in
Dulbecco's modified Eagle medium (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at
37°C in a humidified atmosphere containing 5% CO2.
RNA isolation and
reverse-transcription quantitative polymerase chain reaction
(RT-qPCR)
Total RNA was extracted from the cell lines using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.).
cDNA was synthesized and amplified using the Transcript
First-strand cDNA Synthesis SuperMix (Beijing TransGen Biotech Co.,
Ltd., Beijing, China) in accordance with the manufacturer's
protocol. PCR was performed using a TaqMan Universal PCR Master Mix
kit (Applied Biosystems; Thermo Fisher Scientific, Inc.) in an
Applied Biosystems 7500 system (Thermo Fisher Scientific, Inc.).
Thermocycling conditions were as follows: 95°C for 10 min, followed
by 40 cycles of 95°C for 15 sec, and 60°C for 1 min. The small
nuclear RNA U6 was used as an internal control. The primers used
for RT-PCR were as follows: miR-381: 5′-AGTCTATACAAGGGCAAGCTCTC-3′
(forward primer) and 5′-ATCCATGACAGATCCCTACCG-3′ (reverse primer);
U6snRNA: 5′-CGCAAGGAUGACACGCAAAUUCGUGAAGCGUUCCAUAUUUUU-3′. Gene
expression was normalized to U6 and fold changes were estimated
using the relative quantification (2−ΔΔCq) method
(18). All reactions were performed
in triplicate.
Cell transfection
Hsa-miR-381 mimic (Sense:
5′-UAUACAAGGGCAAGCUCUCUGUTT-3′; anti-sense:
5′-ACAGAGAGCUUGCCCUUGUCGCTT-3′), miR-381 antisense
(5′-ACAGAGAGCUUGCCCUUGUAUA-3′) and miR-381 negative control
(5′-AUCAUCUAUACUGUAAGUAC-3′) oligonucleotides were chemically
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). For
transient transfection, 1×105 LNCaP cells were seeded in
6-well plates, cultured to 70–80% confluence and transfected with
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
final concentration of miR-381 mimic, miR-381 antisense
(anti-miR-381) or miR-381 scramble (miR-NC) in the transfection
system was 100 nM. At 24 h following transfection, the cells were
used for the subsequent experiments.
Cell proliferation
LNCaP cells proliferation was evaluated using the
MTT assay. A total of 5×103 LNCaP cells were seeded into
each well of a 96-well plate and incubated for 24, 48 or 72 h.
Subsequently, 20 µl 0.5 mg/ml MTT solution (dissolved in DMSO) was
added to each well. Following removal of the medium, the absorbance
was measured using a spectrophotometer at a wavelength of 570 nm
(Thermo Fisher Scientific, Inc.).
Cell apoptosis
The induction of apoptosis was measured using flow
cytometry. The LNCaP cells were harvested, washed twice with
ice-cold PBS, and then resuspended in 1× binding buffer
(Invitrogen; Thermo Fisher Scientific, Inc.) at a concentration of
6×105 cells/ml. 100 µl of the solution (6×104 cells) was then
transferred to a 5 ml culture tube. The cells were then incubated
with 5 µl Annexin V-fluorescein isothiocyanate (BD Biosciences, San
Jose, CA, USA) and 5 µl of 1 mg/ml propidium iodide (PI) at room
temperature in the dark for 15 min. Following the incubation
period, 400 µl of 1X binding buffer was added to each sample and
the samples were analyzed using a FACSCalibur flow cytometer (BD
Biosciences, San Jose, CA, USA). The results were obtained by
analyzing the data with FlowJo software (version 7.6.1; FlowJo,
LLC, Ashland, OR, USA). All assays were repeated three times.
Transwell assay
The migratory and invasive abilities of LNCaP cells
were assayed using Transwell chamber (Corning Inc., Corning, NY,
USA). For the migration assays, 1×105 cells were plated in the top
chamber containing a non-coated membrane. The cells were plated in
the serum-free medium and medium (Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS was used as a
chemoattractant in the lower chamber. For invasion assays, the
filters were precoated with Matrigel (BD Biosciences) in the upper
chamber prior to cell seeding. Subsequently, 2×104 cells were
seeded into the upper chamber and the lower chamber was filled with
culture medium supplemented with 10% FBS. The cells were incubated
at 37°C in a tissue culture incubator with 5% CO2. After
16 h, cells that had migrated and invaded to the lower membrane
surface were fixed with 4% paraformaldehyde (Beijing Solarbio,
Science & Technology Co., Ltd., Beijing, China) for 10 min and
stained with 0.1% crystal violet for 10 min at room temperature.
Migratory/invasive cells in five random fields were counted using a
light microscope (Olympus Soft Imaging Solutions GmbH, Münster,
Germany). Three independent experiments were conducted for each
assay.
Bioinformatics analysis
Potential target genes of miR-381 were predicted and
analyzed using TargetScan 6.2 (www.targetscan.org) and miRanda (http://www.microrna.org/microrna/home.do) (19,20).
Western blot analysis
LNCaP cells were harvested and lysed in
radioimmunoprecipitation assay buffer (150 mM NaCl, 50 mM Tris-HCl,
1% NP-40; pH 7.5). The Bradford assay (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was used to determine protein concentration.
Equal amounts (50 µg) of protein were separated via SDS-PAGE on an
8% gel and transferred onto a nitrocellulose membrane (EMD
Millipore, Billerica, MA, USA). After blocking with 10% non-fat dry
milk in Tris-buffered saline for 1 h at room temperature, the
membranes were incubated overnight with anti-androgen receptor (AR)
antibodies (cat. no. sc-7305; dilution, 1:500; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at 4°C, followed by
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibodies (cat. no. 7074; dilution, 1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) for 1 h at room temperature.
The signals were detected using the SuperSignal West Pico
Chemiluminescent Substrate kit (Pierce; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Protein levels were
normalized to total GAPDH, using a monoclonal anti-GAPDH antibody
(cat. no. sc-293335; dilution, 1:1,000; Santa Cruz Biotechnology,
Inc.).
Dual-luciferase reporter assay
The 3′-untranslated region (3′-UTR) of the AR that
was predicted to interact with miR-381 was amplified and inserted
into pMIR-Report Luciferase vector (Ambion; Thermo Fisher
Scientific, Inc.). Mutations within potential miR-381-binding sites
were generated by nucleotide replacement. The 20 ng reporter
vectors, 1 ng pRL-TK Renilla plasmid (Promega Corporation,
Madison, WI, USA) and 20 nM miR-381 mimic or negative control were
transiently co-transfected into 1×106 cells using
Lipofectamine® 2000. After 24 h, the activities of
firefly luciferase and Renilla luciferase in the cell
lysates were measured with the Dual-Luciferase reporter assays
(Promega Corporation, Madison, WI, USA), and values for cells with
reporter genes containing the wild-type AR 3′-UTR were set equally
to 1. Each experiment was repeated three times.
Co-transfection of cells with miR-381
and AR
AR DNA sequences were amplified from LNCaP cell cDNA
by PCR, then subcloned into the pcDNA3.1+ vector (Invitrogen;
Thermo Fisher Scientific, Inc.) and verified by DNA sequencing
(Shanghai GenePharma Co., Ltd., Shanghai, China). Primers for the
amplification were as following: Forward:
5′-AAGCTTTACTCCTCTGCAGTGCCTTG-3′; reverse:
5′-GGATCCACTGGGCCATATGAGGATCA-3′. miR-381 mimics/mimic control (50
nM) and the pc-AR plasmids (0.25 µg) were co-transfected into
cultured 1×105 LNCaP cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) and incubated for 48
h. After 48 h, the cells were used for the subsequent
experiments.
Statistical analysis
Data are presented as the mean ± standard deviation
from at least three independent experiments. Statistical analysis
was performed by SPSS software (version 13.0; SPSS, Inc., Chicago,
IL, USA). Comparisons between two groups were performed using
Student's t-test. The differences among three or more groups were
compared by one-way analysis of variance followed by the least
significant difference post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Downregulation of miR-381 in PCa
cells
To examine the function of miR-381 in the malignant
transformation of prostate epithelial cells, its expression levels
were analyzed in the PCa cell line LNCaP and the normal human
prostate epithelial cell line PrEC. The expression levels of
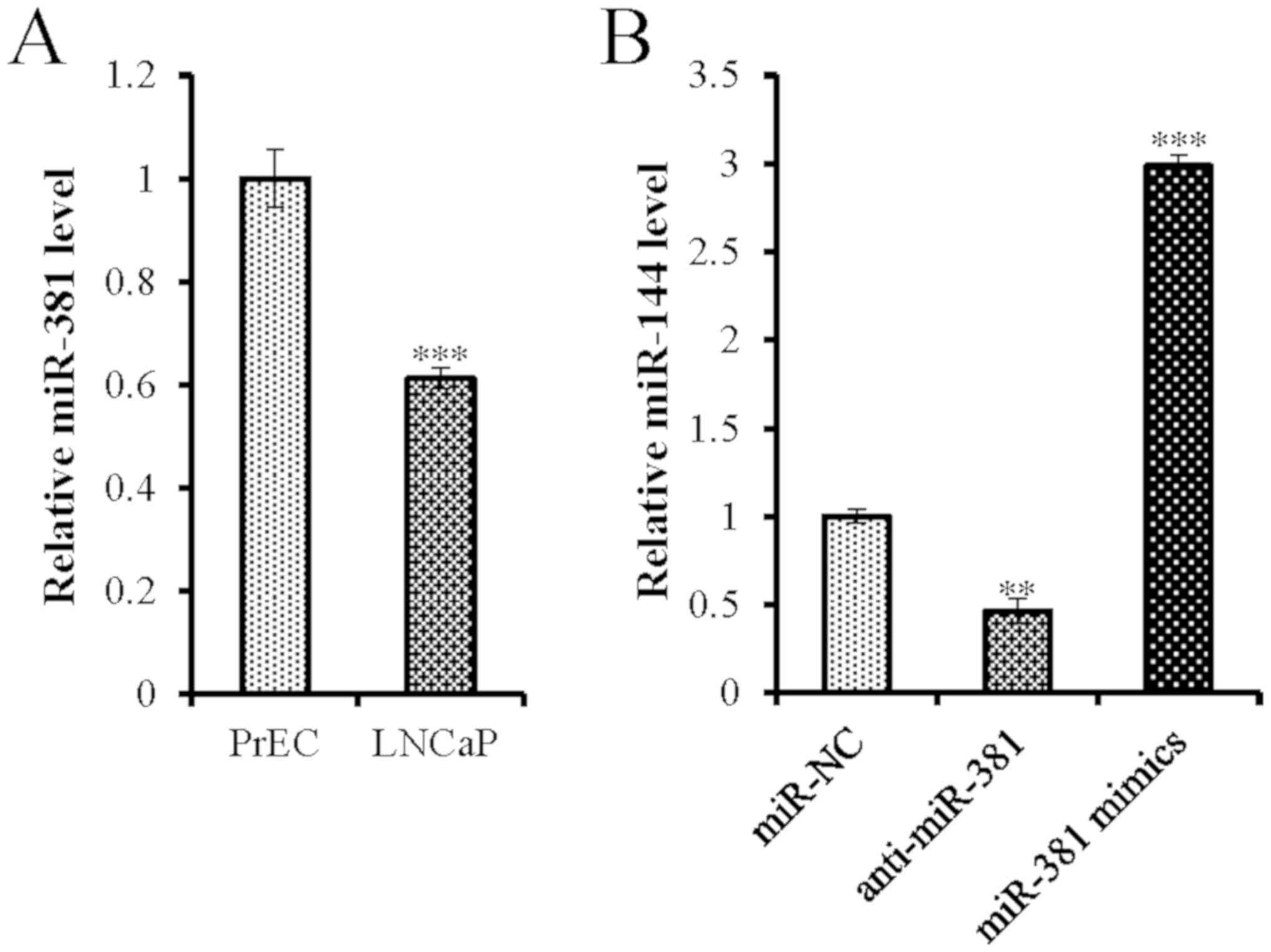
miR-381 in LNCaP cells were ~38.83% lower than those detected in
the PrEC cells (Fig. 1A), indicating
that miR-381 was downregulated in PCa cells.
miR-381 suppresses the proliferation
of PCa cells
To clarify the role of miR-381 in PCa progression,
miR-381 antisense oligonucleotides and mimics were transfected into
LNCaP cells. The cells were further assessed in terms of their
proliferative and apoptotic activities. Successful transfection of
miR-381 antisense oligonucleotides and mimics was confirmed by
RT-qPCR (Fig. 1B). Overexpression of
miR-381 significantly inhibited cell proliferation (Fig. 2A) and increased apoptosis of LNCaP
cells (Fig. 2B and C), whereas the
opposite results were observed when miR-381 was silenced.
Therefore, these results suggested that overexpression of miR-381
may inhibit PCa cell proliferation and promote apoptosis in
vitro.
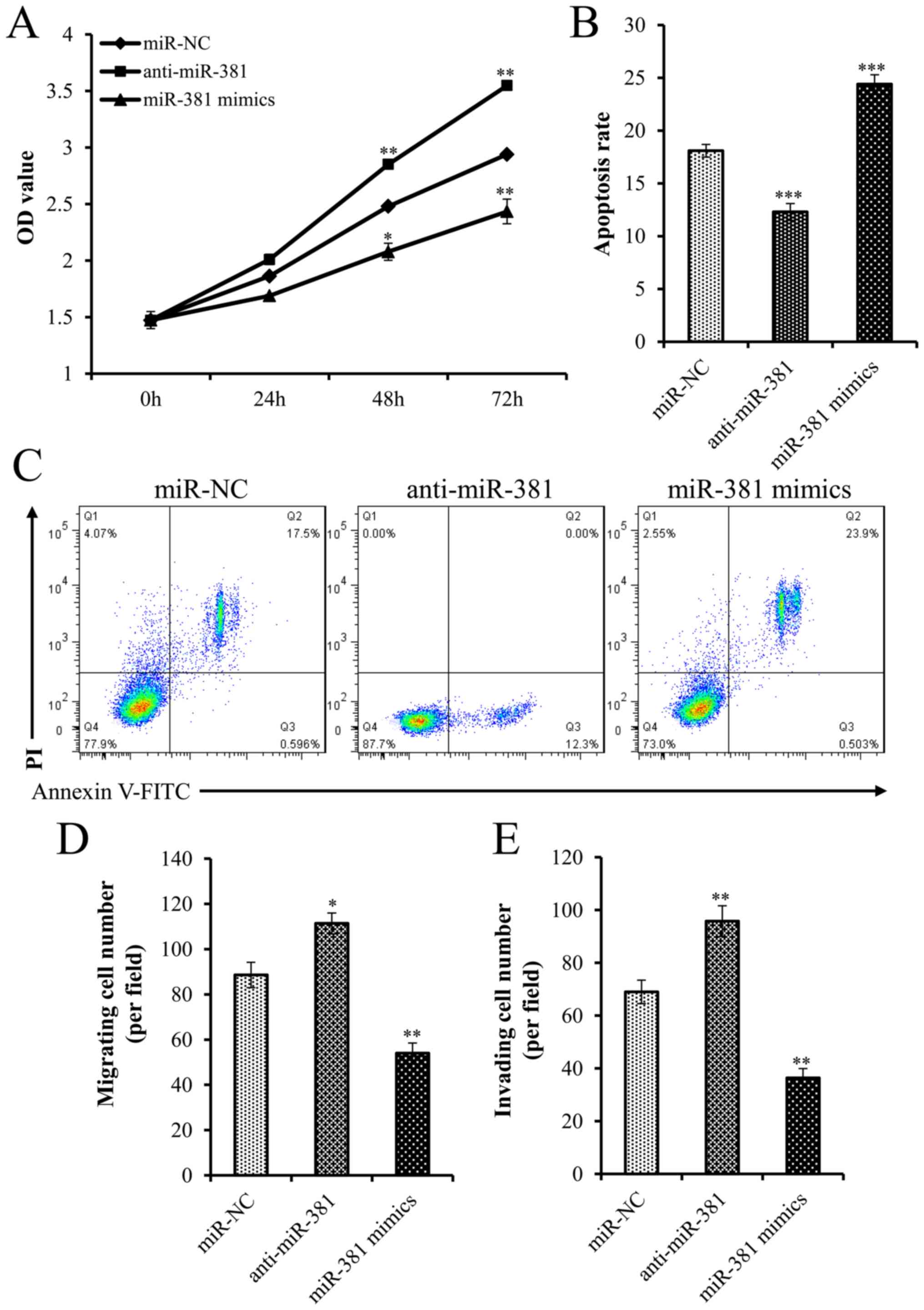 | Figure 2.miR-381 suppressed the proliferation,
migration and invasion of PCa cells. (A) MTT assay was performed to
examine LNCaP cell proliferation. (B and C) Flow cytometry was
performed to detect the apoptotic rate of LNCaP cells. (D)
Migration assay of LNCaP cells transfected with miR-NC,
anti-miR-381 or miR-381 mimics. (E) Invasion assay of LNCaP cells
transfected with miR-NC, anti-miR-381 or miR-381 mimics.
*P<0.05, **P<0.01, ***P<0.001 compared with miR-NC. FITC,
fluorescein isothiocyanate; miR, microRNA; NC, negative control;
OD, optical density; PI, propidium iodide. |
miR-381 inhibits migration and
invasion of PCa cells
The role of miR-381 in PCa cell migration and
invasion was investigated. miR-381 oligonucleotides and mimics were
transfected into PCa cells. Overexpression of miR-381 inhibited the
migratory and invasive activities of PCa cells compared with the
corresponding activities of the cells in the control group
(Fig. 2D and E). In addition, with
decreasing expression of miR-381, the migration and invasion of PCa
cells was attenuated (Fig. 2D and
E).
miR-381 directly targets AR
The aforementioned results suggested that miR-381
may act as a tumor suppressor in PCa. Consequently, the putative
target genes of miR-381 were investigated using TargetScan and
miRanda. Excitingly, the results revealed that there were
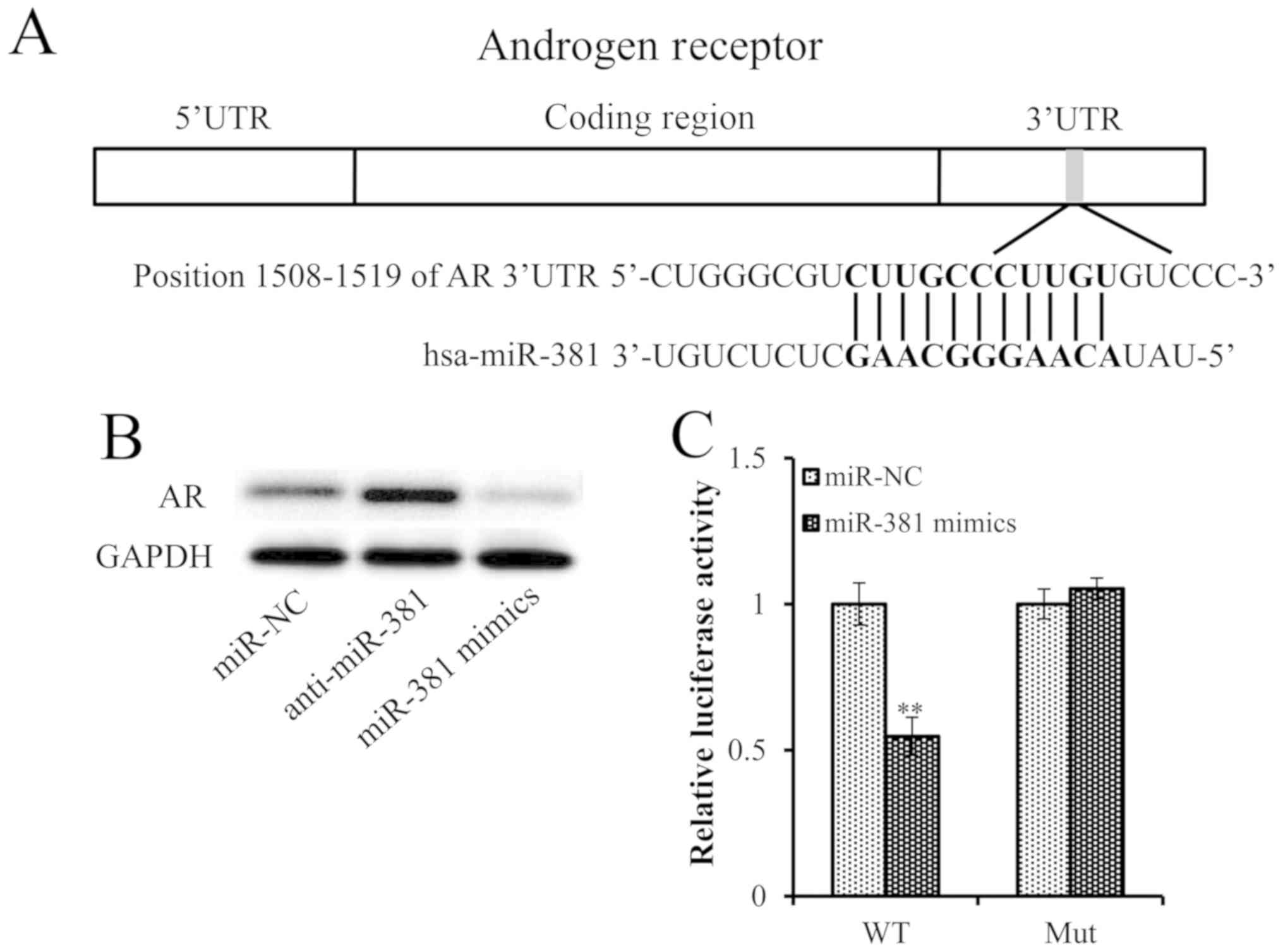
consequential pairs between miR-381 and 3′UTR of AR. A sequence
analysis revealed that the putative targeting site of miR-381 was
located at nt 1508–1519 of the AR 3′UTR (Fig. 3A), indicating that AR may be a
regulatory target of miR-381 and was selected for subsequent
investigation (Fig. 3A).
Overexpression of miR-381 decreased AR expression levels, whereas
inhibition of miR-381 increased the protein expression levels of AR
(Fig. 3B), suggesting that the AR
was a target gene of miR-381. To further confirm that the AR is a
direct target of miR-381, a dual-luciferase reporter assay was
performed. Overexpression of miR-381 significantly suppressed the
luciferase activity of wild-type AR 3′UTR. This effect was fully
reversed when the potential miR-381 binding site was mutated
(Fig. 3C).
AR overexpression partially reverses
the tumor-suppressive effects of miR-381
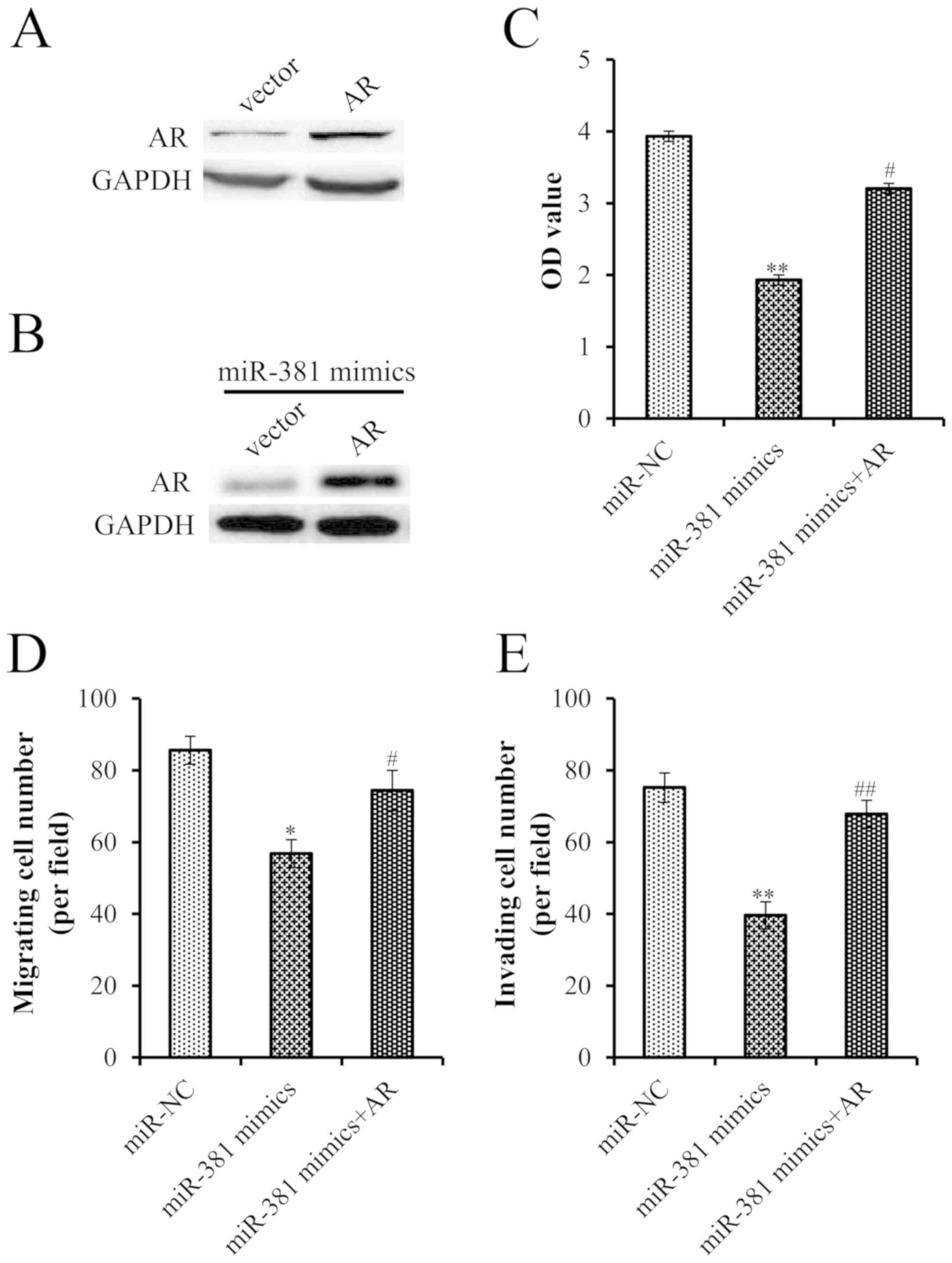
To further verify the interaction between miR-381
and the AR, this study investigated whether overexpression of AR
rescued PCa cells from the inhibitory effects of miR-381.
Successful transfection of an AR plasmid was confirmed by western
blot analysis (Fig. 4A).
Furthermore, transfection with the AR plasmid reversed the effects
of miR-381 overexpression on AR expression (Fig. 4B). Furthermore, overexpression of AR
significantly attenuated the tumor suppressive effects of miR-381,
highlighting the importance of the AR for the action of miR-381 in
cell proliferation, migration and invasion (Fig. 4C-E).
Discussion
It is well known that a number of miRNAs are
involved in prostate tumorigenesis (21,22). In
the present study, miR-381 was significantly downregulated in PCa
cells compared with in normal prostate epithelial cells.
Furthermore, overexpression of miR-381 suppressed the
proliferation, migration and invasion of PCa cells, suggesting that
miR-381 may serve a role as a potential tumor suppressor in
PCa.
miR-381 has attracted attention for its role in
carcinogenesis and cancer treatment. Several studies have
demonstrated that miR-381 functions as a tumor suppressor, and is
able to suppress tumor growth and progression in colon cancer,
ovarian cancer and renal carcinoma (10,15,23).
Zhang et al (11)
demonstrated that miR-381 may serve as a novel tumor suppressor
that decreases hepatocellular carcinoma growth and invasion by
targeting liver receptor homolog-1. Xue et al (14) suggested that miR-381 serves a major
role in breast cancer growth, epithelial-mesenchymal transition
(EMT) and metastasis by targeting C-X-C chemokine receptor 4. In
addition, the restoration of miR-381 significantly inhibits the
invasion and migration of colorectal cancer cells and the EMT
(24). Conversely, loss of miR-381
expression predicts poor prognosis in lung adenocarcinoma (12). Furthermore, miR-381 is aberrantly
downregulated in human myelogenous leukemia (K562/ADM) cells,
whereas overexpression of miR-381 is correlated with reduced
expression of the multidrug resistance protein 1 and increased drug
uptake by cells (25). These
findings suggest that miR-381 may target different genes in various
cell types, contributing to different biological processes.
Notably, it has been reported that the expression levels of miR-381
are reduced in PCa (8,9). Nevertheless, the biological function
and the underlying mechanism of miR-381 in PCa remain unclear. The
current study identified miR-381 as a potential tumor suppressor in
PCa cells.
In order to elucidate the molecular mechanisms
involved in miR-381-mediated inhibition of cell proliferation,
migration and invasion, the AR was selected for further analysis as
it was identified as one the predicted target genes of miR-381. The
AR is a steroid hormone-activated transcription factor that belongs
to the steroid/nuclear receptor superfamily (26). This receptor controls the growth and
differentiation of the normal prostate gland, and serves an
important role in the development and progression of
castration-resistant PCa (27).
Although androgens are the primary ligand for the AR, additional
factors such as interleukin-6, transcriptional intermediary factor
2 may contribute to its abnormal activation in their absence
(28,29). The AR regulates downstream target
genes and alters various cell functions, including cell
proliferation, cell cycle progression and apoptosis (30–32). It
has been reported that the AR serves a central role in the
development and progression of androgen-independent PCa via
multiple mechanisms (33).
Therefore, it is essential to examine the effective inhibition of
the expression and function of the AR, so as to prevent the
development of PCa. In the present study, overexpression of miR-381
reduced the expression levels of AR. Bioinformatics analysis
revealed binding sites for the miR-381 seed sequence in the 3′UTR
of AR. Deletion mutagenesis and a dual-luciferase reporter gene
assay demonstrated that the predicted miR-381 target sites in the
3′UTR of AR were functional. Furthermore, the present study
indicated that miR-381 effectively decreased the protein expression
levels of AR, whereas inhibition of miR-381 revealed opposite
results. Finally, overexpression of AR partially reversed
miR-381-induced inhibition of PCa cell proliferation, migration and
invasion. Future studies are required to explore other target genes
of miR-381 that may be involved in PCa.
In conclusion, the present study highlighted the
roles of miR-381 in terms of its ability to suppress growth,
migration and invasion of PCa cells by directly targeting the AR.
These findings suggested that miR-381 may act as a tumor suppressor
in PCa, and may serve as a novel therapeutic target.
Acknowledgements
The authors would like to thank Dr Huizhi Zhang
(Ningbo Diagnostic Pathology Center) for assistance with the
Transwell assay.
Funding
The present work was supported by the Hospital
Foundation of the HwaMei hospital, University of Chinese Academy of
Sciences (Ningbo No. 2 Hospital) (grant no. 2018HMKY36).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XR, TTG and HXS conceived the study and revised the
manuscript. XR, HFP and SLS wrote the main text and prepared all of
the figures. XR performed all the experiments. HXS provided funding
support. All authors reviewed the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He Y, Zhang Q, Jiang R and Li Y: Role and
clinical significance of miRNA-381 in prostate cancer. Int J Clin
Exp Med. 10:2173–2180. 2017.
|
|
9
|
Formosa A, Markert EK, Lena AM, Italiano
D, Finazzi-Agro′ E, Levine AJ, B rnardini S, Garabadgiu AV, Melino
G and Candi E: MicroRNAs, miR-154, miR-299-5p, miR-376a, miR-376c,
miR-377, miR-381, miR-487b, miR-485-3p, miR-495 and miR-654-3p,
mapped to the 14q32.31 locus, regulate proliferation, apoptosis,
migration and invasion in metastatic prostate cancer cells.
Oncogene. 33:5173–5182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang Y, Zhao Q, Fan L, Zhang Z, Tan B,
Liu Y and Li Y: Down-regulation of MicroRNA-381 promotes cell
proliferation and invasion in colon cancer through up-regulation of
LRH-1. Biomed Pharmacother. 75:137–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Q, Zhao S, Pang X and Chi B:
MicroRNA-381 suppresses cell growth and invasion by targeting the
liver receptor homolog-1 in hepatocellular carcinoma. Oncol Rep.
35:1831–1840. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rothschild SI, Tschan MP, Jaggi R, Fey MF,
Gugger M and Gautschi O: MicroRNA-381 represses ID1 and is
deregulated in lung adenocarcinoma. J Thorac Oncol. 7:1069–1077.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cao Q, Liu F, Ji K, Liu N, He Y, Zhang W
and Wang L: MicroRNA-381 inhibits the metastasis of gastric cancer
by targeting TMEM16A expression. J Exp Clin Cancer Res. 36:292017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xue Y, Xu W, Zhao W, Wang W, Zhang D and
Wu P: miR-381 inhibited breast cancer cells proliferation,
epithelial-to-mesenchymal transition and metastasis by targeting
CXCR4. Biomed Pharmacother. 86:426–433. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xia B, Li H, Yang S, Liu T and Lou G:
miR-381 inhibits epithelial ovarian cancer malignancy via YY1
suppression. Tumor Biol. 37:9157–9167. 2016. View Article : Google Scholar
|
|
16
|
Tang H, Liu X, Wang Z, She X, Zeng X, Deng
M, Liao Q, Guo X, Wang R, Li X, et al: Interaction of hsa-miR-381
and glioma suppressor LRRC4 is involved in glioma growth. Brain
Res. 1390:21–32. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen B, Duan L, Yin G, Tan J and Jiang X:
miR-381, a novel intrinsic WEE1 inhibitor, sensitizes renal cancer
cells to 5-FU by up-regulation of Cdc2 activities in 786-O. J
Chemother. 25:229–238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in drosophila. Genome Biol.
5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Porkka KP, Pfeiffer MJ, Waltering KK,
Vessella RL, Tammela TL and Visakorpi T: MicroRNA expression
profiling in prostate cancer. Cancer Res. 67:6130–6135. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ozen M, Creighton C, Ozdemir M and Ittmann
M: Widespread deregulation of microRNA expression in human prostate
cancer. Oncogene. 27:1788–1793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen B and Liu B: miRNA-381 inhibits the
invasion of renal carcinoma and the underlying mechanisms. Zhong
Nan Da Xue Xue Bao. Yi Xue Ban. 40:1053–1059. 2015.(In
Chinese).
|
|
24
|
He X, Wei Y, Wang Y, Liu L, Wang W and Li
N: miR-381 functions as a tumor suppressor in colorectal cancer by
targeting Twist1. Onco Targets Ther. 9:1231–1239. 2016.PubMed/NCBI
|
|
25
|
Xu Y, Ohms SJ, Li Z, Wang Q, Gong G, Hu Y,
Mao Z, Shannon MF and Fan JY: Changes in the expression of miR-381
and miR-495 are inversely associated with the expression of the
MDR1 gene and development of multi-drug resistance. PLoS One.
8:e820622013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heinlein CA and Chang C: Androgen receptor
(AR) coregulators: An overview. Endocr Rev. 23:175–200. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nadiminty N, Tummala R, Lou W, Zhu Y,
Zhang J, Chen X, eVere White RW, Kung HJ, Evans CP and Gao AC:
MicroRNA let-7c suppresses androgen receptor expression and
activity via regulation of Myc expression in prostate cancer cells.
J Biol Chem. 287:1527–1537. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koochekpour S: Androgen receptor signaling
and mutations in prostate cancer. Asian J Androl. 12:639–657. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marques RB, Dits NF, Erkens-Schulze S, van
IJcken WF, van Weerden WM and Jenster G: Modulation of androgen
receptor signaling in hormonal therapy-resistant prostate cancer
cell lines. PLoS One. 6:e231442011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dehm SM and Tindall DJ: Molecular
regulation of androgen action in prostate cancer. J Cell Biochem.
99:333–344. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin B, Wang J, Hong X, Yan X, Hwang D, Cho
JH, Yi D, Utleg AG, Fang X, Schones DE, et al: Integrated
expression profiling and ChIP-seq analyses of the growth inhibition
response program of the androgen receptor. PLoS One. 4:e65892009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ngan S, Stronach E, Photiou A, Waxman J,
Ali S and Buluwela L: Microarray coupled to quantitative RT-PCR
analysis of androgen-regulated genes in human LNCaP prostate cancer
cells. Oncogene. 28:2051–2063. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heinlein CA and Chang C: Androgen receptor
in prostate cancer. Endocr Rev. 25:276–308. 2004. View Article : Google Scholar : PubMed/NCBI
|