Introduction
Ovarian cancer (OC) is the eighth most common type
of cancer and the eighth leading cause of cancer-associated
mortality among women worldwide. A total of 295,414 women worldwide
were reported to be diagnosed with OC in 2018; and 184,799 females
succumbed from this gynecologic malignancy (1). The combination of surgery with
chemotherapeutic agents based on platinum salts and taxanes is the
main treatment approach for patients with advanced-stage OC. A
total of 65–80% of patients with OC respond to chemotherapy
initially; however, the majority of patients experience cancer
recurrence and become resistant to chemotherapy (2). In addition, the 5-year survival of
patients with advanced-stage OC is only 27% (3). Therefore, novel agents for OC treatment
are urgently required.
Chaetocin is a natural product isolated from
Chaetomium fungi, and possesses antibiotic properties and a
thiodioxopiperazine structure (4,5).
Chaetocin has been reported to exhibit anticancer activity against
various cancer cell lines, including hepatocellular carcinoma,
glioma, myeloma, non-small cell lung cancer and leukemia cells
(6–14). Isham et al (7) revealed that chaetocin exerts its
antimyeloma activity by affecting oxidative stress. Additionally,
chaetocin was reported to exhibit antihepatoma activity by
dysregulating the splicing of hypoxia-inducible factor 1α pre-mRNA
(11); however, the pharmacological
effects and detailed mechanism of chaetocin against OC remain
unclear. In the present study, the pharmacological effects of
chaetocin on OC and the underlying mechanism were investigated.
Materials and methods
Reagents
Chaetocin and N-acetyl-L-cysteine (NAC) were
obtained from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
z-VAD-fmk was purchased from Selleck Chemicals (Houston, TX, USA).
Antibodies against poly (ADP-ribose) polymerase (PARP; 1:1,000;
cat. no. 9532), caspase-3 (1:1,000; cat. no. 9662) and
cleaved-caspase-3 (1:1,000; cat. no. 9661) were obtained from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Anti-β-actin
(1:10,000; cat. no. 60008-1-Ig), and anti-mouse immunoglobulin G
(1:5,000; cat. no. SA00001-1) and anti-rabbit immunoglobulin G
(1:5,000; cat. no. SA00001-2) horseradish peroxidase-conjugated
secondary antibodies were purchased from ProteinTech Group, Inc.
(Chicago, IL, USA).
Cell culture
SKOV-3 (cat. no. ATCC HTB-77) and OVCAR-3 (cat. no.
ATCC HTB-161) cells were obtained from the American Type Culture
Collection (Manassas, VA, USA). KGN (cat. no. BNCC337610), A2780
(cat. no. BNCC341157) and IOSE80 (cat. no. BNCC340318) cells were
obtained from BeNa Culture Collection (Beijing, China). All cell
lines were cultured in RPMI-1640 (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), supplemented with 10% fetal bovine serum (Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 10 mg/ml
streptomycin (Thermo Fisher Scientific, Inc.) in a humidified
atmosphere with 5% CO2 at 37°C.
Cell viability assay
Cell viability was analyzed via a Cell Counting
kit-8 (CCK8) assay (Nanjing KeyGen Biotech Co., Ltd., Nanjing,
China). The OC cells were seeded in 96-well microplates
(1×104 cells/well) and incubated at 37°C overnight.
Following incubation with chaetocin (0.05, 0.1, 0.25, 0.5, 0.75, 1
and 2 µM) at 37°C for 24 h, 20 µl of CCK8 reagent was added into
each well and incubated for another 4 h. The absorbance was
measured using a multimode reader at 450 nm.
Colony formation assay
A total of 500 cells/well were seeded in a 6-well
plate and incubated at 37°C overnight. The following day, the cells
were treated with 2.5 and 5 nM chaetocin and incubated for 9 days.
The cells were then washed with PBS and fixed in ice-cold methanol
for 10 min following treatment with chaetocin. The cells were then
stained with crystal violet solution at room temperature for 10 min
and washed with water. Images of the colonies were captured using
an Epson Perfection V370 Photo scanner (Epson America, Inc., Long
Beach, CA, USA).
Cell cycle analysis
The cells were treated with chaetocin (0.5 and 1 µM)
for 12 h. Following treatment, the cells were collected and fixed
with 66% ice-cold ethanol at 4°C overnight, and then stained with
500 µl propidium iodide (PI; BD Biosciences, Franklin Lakes, NJ,
USA) at room temperature for 15 min in the dark. The cell cycle
distribution was analyzed by flow cytometry (λex=488 nm,
λem=630 nm, 10000 events analyzed).
Analysis of apoptosis
The chaetocin-induced apoptosis of OVCAR-3 cells was
analyzed using an Annexin V-fluorescein isothiocyanate (FITC)/PI
staining kit (Nanjing KeyGen Biotech Co., Ltd.). The cells were
treated with various concentrations of chaetocin (0.5, 1 and 2 µM)
for 24 h and then washed with PBS. Subsequently, the cells were
resuspended in a solution containing 500 µl binding buffer, 5 µl
annexin V-FITC and 5 µl PI for 15 min at 37°C in the dark. The
apoptotic cells were then analyzed by flow cytometry (for annexin
V-FITC, λex=488 nm, λem=530 nm; for PI,
λex=488 nm, λem=630 nm; 10000 events
analyzed). Annexin V+ and Annexin
V+/PI+ cells were determined as apoptotic
cells.
Determination of intracellular
reactive oxygen species (ROS)
ROS generation was measured using a ROS assay kit
(Beyotime Institute of Biotechnology, Shanghai, China). The cells
were loaded with 2,7-dichlorofluorescin diacetate (10 µM) and
incubated for 20 min at 37°C in the dark. Following this, the cells
were treated with chaetocin for 1 h and washed with PBS. The
fluorescence was measured by flow cytometry (λex=488 nm,
λem=530 nm, 10 000 events analyzed).
Western blot analysis
The cells were treated with chaetocin and proteins
were extracted using radioimmunoprecipitation assay buffer (Cell
Signaling Technology, Inc.) containing protease and phosphatase
inhibitors (Nanjing KeyGen Biotech Co., Ltd.). The concentration of
proteins was determined with a BCA protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.); 50 µg total cellular protein were
separated by 10% SDS-PAGE (PARP, caspase-3 and β-actin) or 15%
SDS-PAGE (cleaved-caspase-3) and transferred onto polyvinylidene
difluoride membranes. The membranes were then blocked in 5% non-fat
dry milk and incubated with primary antibodies overnight at 4°C.
The following day, the membranes were incubated with the anti-mouse
immunoglobulin G or anti-rabbit immunoglobulin G horseradish
peroxidase-conjugated secondary antibodies at room temperature for
1 h, and target proteins were detected using enhanced
chemiluminescence. The gray values were analyzed using ImageJ
software (version 1.52; National Institutes of Health, Bethesda,
MD, USA).
Statistical analysis
All experiments were performed in triplicate, and
data are presented as the mean ± standard deviation. Statistical
analysis was conducted via one-way analysis of variance followed by
Tukeys post hoc test using GraphPad Prism software (version 6;
GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Chaetocin significantly reduces OC
cell viability
The present study determined the effects of
chaetocin on OC cell viability. Various OC cell lines were treated
with chaetocin (0.05, 0.1, 0.25, 0.5, 0.75, 1 and 2 µM) for 24 h,
and the cell viability was then detected via a CCK8 assay. As
presented in Fig. 1A-D, chaetocin
significantly inhibited the viability of SKOV-3, OVCAR-3, KGN and
A2780 cells in a dose-dependent manner, with half-maximal
inhibitory concentrations of 0.30, 60.66, 81.86 and 100.60 nM,
respectively. In addition, the half-maximal inhibitory
concentration of chaetocin in the IOSE80 normal ovarian surface
epithelium cell line was 206.10 nM, (Fig. 1E), which was higher than the
half-maximal inhibitory concentration of chaetocin in the OC cells
(Fig. 1F).
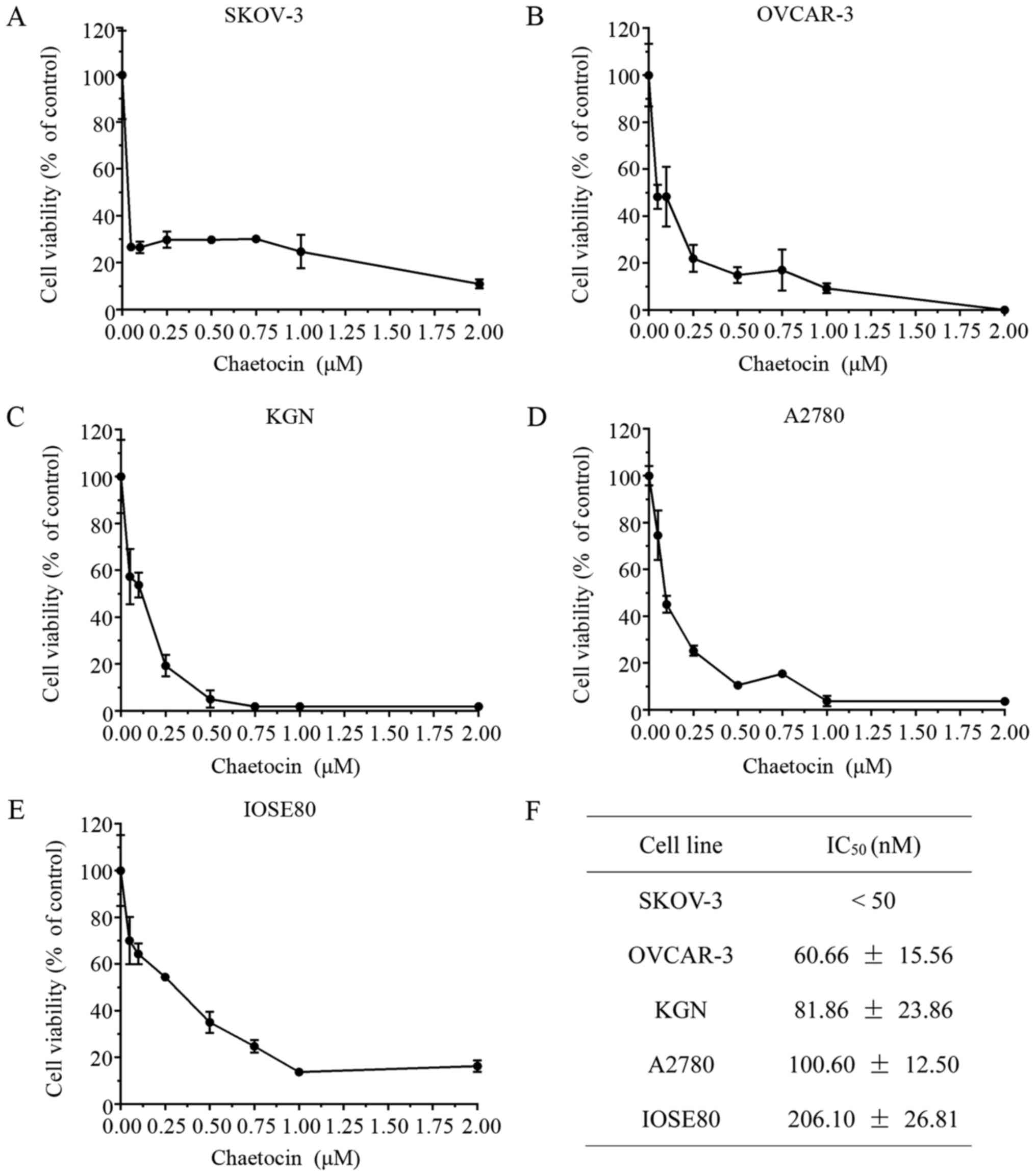 | Figure 1.Chaetocin significantly reduces OC
cell viability. (A) SKOV-3, (B) OVCAR-3, (C) KGN, (D) A2780 and (E)
IOSE80 cells were treated with various concentrations of chaetocin
(0.05, 0.1, 0.25, 0.5, 0.75, 1 and 2 µM) for 24 h; cell viability
was detected via a Cell Counting kit-8 assay. (F) IC50
values of chaetocin for each OC cell line and the normal ovarian
surface epithelium cell line. OC, ovarian cancer; IC50,
half-maximal inhibitory concentration. |
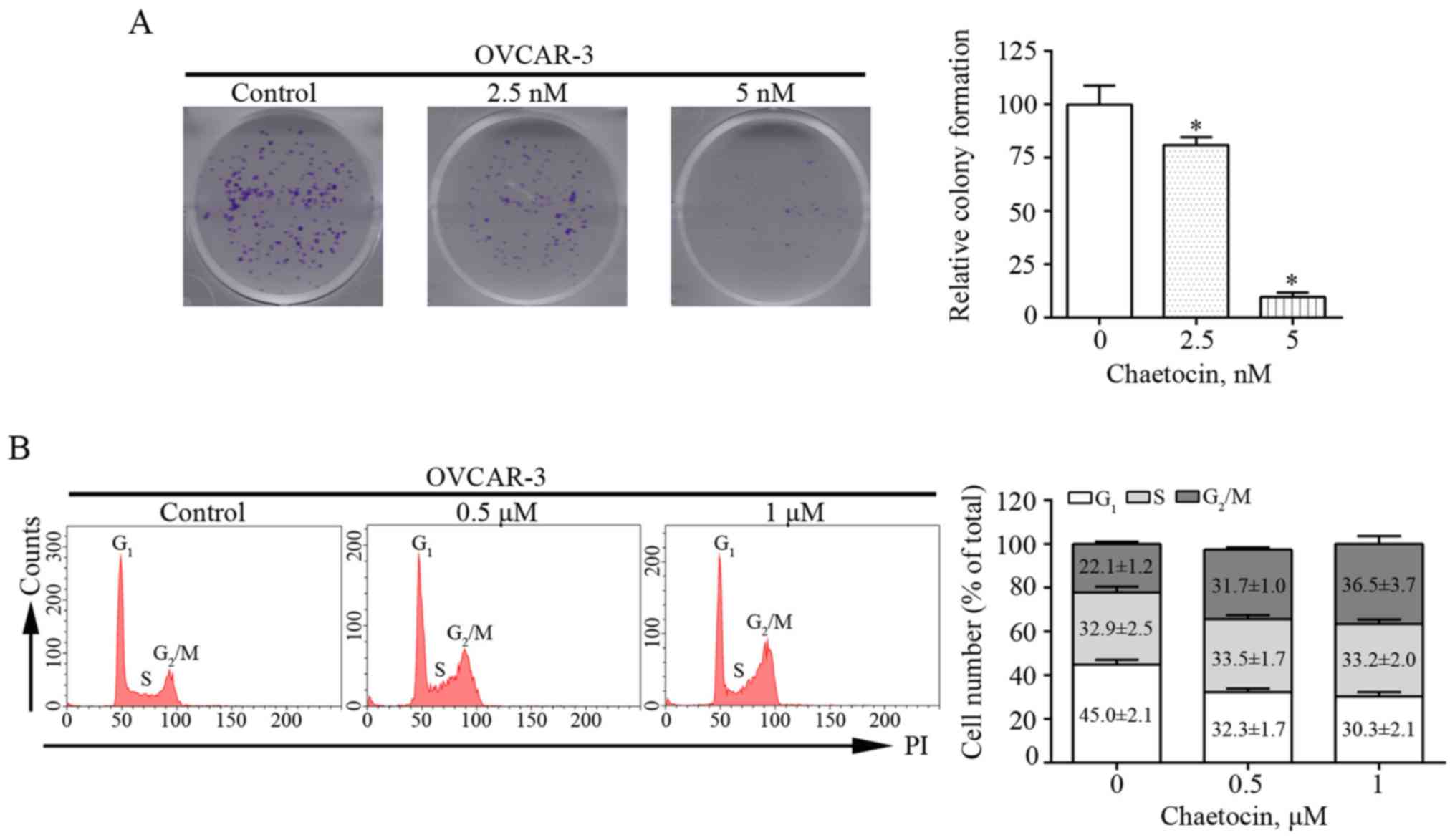
Chaetocin inhibits the proliferation
of OC cells by inducing G2/M phase arrest
Colony formation assays were performed in the
present study to determine the effects of chaetocin on the
proliferation of OVCAR-3 cells. Chaetocin treatment at various
concentrations resulted in fewer and smaller colonies of OVCAR-3
cells after 9 days (Fig. 2A). This
suggested that chaetocin effectively inhibited the proliferation of
OVCAR-3 cells. In addition, whether the reduced proliferation of
OVCAR-3 cells induced by chaetocin was associated with alterations
in the cell cycle was investigated. The distribution of the cell
cycle was analyzed by flow cytometry. Compared with that in the
control group, the percentage of cells in the G2/M phase
increased from 22.1% to 31.7 and 36.5% in OVCAR-3 cells treated
with 0.5 and 1 µM chaetocin for 12 h, respectively (Fig. 2B). These results indicated that
chaetocin inhibited the proliferation of OC cells by arresting the
cell cycle at the G2/M phase.
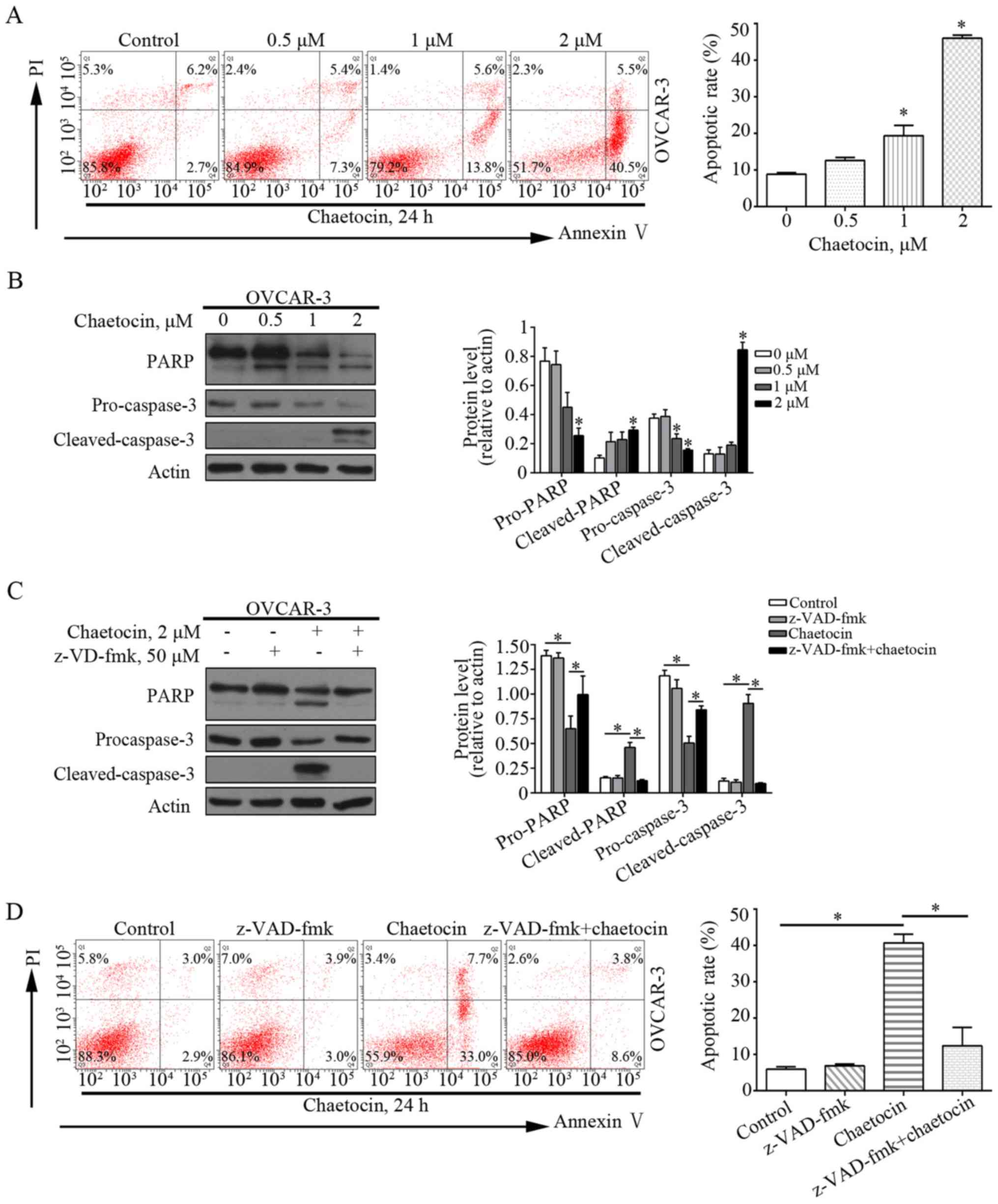
Chaetocin induces the
caspase-dependent apoptosis of OC cells
Flow cytometry was performed to analyze whether
chaetocin induced the apoptosis of OVCAR-3 cells. As presented in
Fig. 3A, chaetocin significantly
increased the percentage of apoptotic OVCAR-3 cells, from 8.9% in
the control group to 19.4 and 46.0% in the 1 and 2 µM chaetocin
treatment groups, respectively. This indicated that chaetocin
induced apoptotic cell death in OC cells. In addition,
apoptosis-associated proteins, including caspase-3 and PARP, were
affected by chaetocin. Chaetocin decreased the expression levels of
caspase-3 and PARP, but increased those of the cleaved forms of
caspase-3 and PARP in OVCAR-3 cells (Fig. 3B). This further suggested that
apoptosis was induced by chaetocin, which may occur via the
caspase-dependent pathway. The pan-caspase inhibitor z-VAD-fmk was
then used to validate whether the chaetocin-induced GC cell
apoptosis depended on the caspase pathway. Of note, when OVCAR-3
cells were co-treated with chaetocin and z-VAD-fmk, the
chaetocin-induced activation of the caspase pathway and resultant
apoptosis were reduced (Fig. 3C and
D). Collectively, these results suggested that chaetocin
induced the caspase-dependent apoptosis of OC cells.
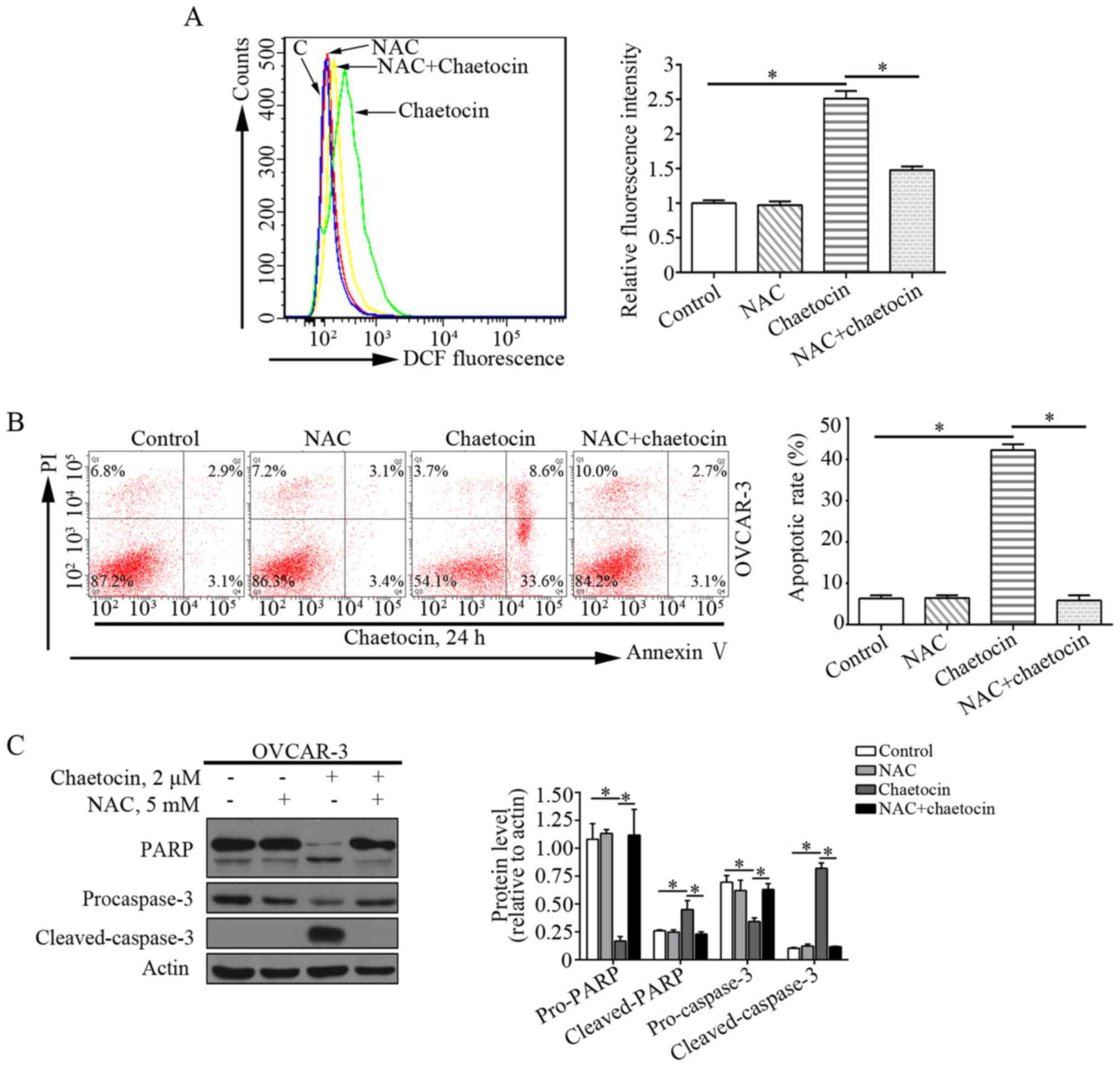
Chaetocin increases ROS accumulation
in OC cells
ROS serve an essential role in cell proliferation
and apoptosis. Previous studies have reported that chaetocin exerts
the antimyeloma activity by affecting oxidative stress (7,15). In
addition, the effects of chaetocin on the levels of ROS in OVCAR-3
cells were determined. Compared with the control group, the levels
of ROS were markedly increased in OVCAR-3 cells treated with
chaetocin. This suggested that chaetocin induced ROS accumulation
in OVCAR-3 cells (Fig. 4A).
ROS production is crucial for the
chaetocin-induced caspase-dependent apoptosis of OC cells
The present study investigated whether ROS
production mediated the anticancer activity of chaetocin against
OVCAR-3 cells. The ROS scavenger NAC was used to inhibit the
excessive ROS accumulation induced by chaetocin (Fig. 4A). When the cells were co-treated
with chaetocin and NAC, the effects of chaetocin on the apoptosis
of OVCAR-3 cells were notably abrogated (Fig. 4B). In addition, the cleavage of
caspase-3 induced by chaetocin in OVCAR-3 cells was reversed
(Fig. 4C). These data indicated that
chaetocin-induced ROS accumulation may be associated with the
caspase-dependent apoptosis of OVCAR-3 cells.
Discussion
OC is one of the most common types of gynecological
cancer (1,16). Carboplatin or cisplatin combined with
paclitaxel is the standard treatment for patients with
advanced-stage OC, however, the therapeutic effects are limited as
the majority of patients experience cancer recurrence and become
resistant to these anticancer agents (2,3,17). Therefore, the development of novel
effective agents against OC is required. In the present study, the
therapeutic effects of chaetocin, a natural product isolated from
Chaetomium fungi, were investigated in OC cells. Chaetocin
induced cell cycle arrest of the OC cells in the G2/M
phase and inhibited proliferation. Additionally, chaetocin caused
the caspase-dependent apoptosis of OC cells. The levels of ROS were
increased in OC cells following chaetocin treatment, which led to
chaetocin-induced caspase-dependent apoptosis. The results of the
present study suggest that chaetocin may be a potential therapeutic
agent for the treatment of OC.
Apoptosis is a process of programmed cell death;
several natural products have been reported to induce cancer cell
death via apoptosis (18,19). There are two major pro-apoptotic
pathways: The death receptor-mediated extrinsic pathway and the
mitochondrial-mediated intrinsic pathway, in which the caspase
family of proteins serve a pivotal role in the process of
programmed cell death. The extrinsic pathway activates caspase-8,
while the intrinsic pathway activates caspase-9. These pathways
activate caspase-3, which cleaves its substrate PARP and induces
apoptosis (20,21). Han et al (15), reported that chaetocin exhibited
pro-apoptotic effects on human melanoma cells via the intrinsic
mitochondrial pathway. In acute myeloid leukemia cells, chaetocin
induced apoptosis via the death receptor-mediated extrinsic pathway
and mitochondrial-mediated intrinsic pathway (12). Similar to these studies, the results
of the present study revealed that chaetocin induced the apoptosis
of OC cells. Additionally, the caspase pathway was activated by
chaetocin in OC cells; the pan-caspase inhibitor z-VAD-fmk reversed
the apoptotic effects of chaetocin, suggesting that chaetocin
induces the caspase-dependent apoptosis of OC cells.
ROS, comprising oxygen radicals and non-radicals,
are a group of chemical molecules with high reactivity and
important roles in cellular proliferation, differentiation and
death (22–24). Excessive ROS accumulation can result
in cell damage and death; cancer cells exhibit increased ROS levels
compared with their normal counterparts (25–28).
Therefore, developing ROS-targeting agents may be a potential
strategy for the treatment of various types of cancer. In addition,
numerous natural products have been reported to have notable
anticancer activity by inducing ROS production. For example,
piperlongumine, a natural alkaloid isolated from the fruit of long
pepper, was observed to exhibit notable cytotoxic activity in
numerous types of cancer by elevating ROS levels (29,30). It
has also been reported that chaetocin increases ROS levels and
induces apoptosis in acute myeloid leukemia, myeloma and
intrahepatic cholangiocarcinoma cells (7,12,31). In
the present study, elevated levels of ROS were detected in OVCAR-3
cells following chaetocin treatment. Of note, when
chaetocin-induced excessive ROS production was inhibited by NAC,
the chaetocin-induced apoptosis of OC cells was markedly abrogated.
Additionally, the activation of caspases induced by chaetocin in OC
cells was reversed. Collectively, these results indicated that
chaetocin increased the production of ROS, which may be associated
with the caspase-dependent apoptosis of OVCAR-3 cells induced by
chaetocin.
In conclusion, the findings of the present study
indicated that chaetocin suppressed the proliferation and induced
the caspase-dependent apoptosis of OC cells via the excessive
production of ROS. To the best of our knowledge, the present study
is the first to investigate the detailed pharmacological effects of
chaetocin on OC cells and the underlying mechanism. Therefore,
chaetocin may be considered as a potential therapeutic agent in
treating patients with OC; however, the effects of chaetocin in
vivo and the molecular mechanism by which chaetocin affects OC
cells require further investigation.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by
grants from the National Natural Science Foundation of China (grant
no. 81702399), Guangdong Province Medical Scientific Research
Foundation (grant nos. C2018053 and C2017034) and the National
Medical Science and Technology Foundation (grant no.
W2016CWGD05).
Availability of data and materials
The datasets used in the present study are available
from the corresponding author upon reasonable request.
Authors contributions
SH and ZH made substantial contributions to the
conception of the present study and designed the experiments. ZL
and WY performed the experiments and drafted the manuscript. LH and
LW participated in data collection and analysis. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
OC
|
ovarian cancer
|
|
ROS
|
reactive oxygen species
|
|
NAC
|
N-acetyl-L-cysteine
|
|
PI
|
propidium iodide
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gamarra-Luques CD, Hapon MB, Goyeneche AA
and Telleria CM: Resistance to cisplatin and paclitaxel does not
affect the sensitivity of human ovarian cancer cells to
antiprogestin-induced cytotoxicity. J Ovarian Res. 7:452014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cho KR and Shih IeM: Ovarian cancer. Annu
Rev Pathol. 4:287–313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sekita S, Yoshihira K, Natori S, Udagawa
S, Muroi T, Sugiyama Y, Kurata H and Umeda M: Mycotoxin production
by Chaetomium spp. and related fungi. Can J Microbiol.
27:766–772. 1981. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hauser D, Weber HP and Sigg HP: Isolation
and configuration of Chaetocin. Helv Chim Acta. 53:1061–1073.
1970.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu X, Guo S, Liu X and Su L: Chaetocin
induces endoplasmic reticulum stress response and leads to death
receptor 5-dependent apoptosis in human non-small cell lung cancer
cells. Apoptosis. 20:1499–1507. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Isham CR, Tibodeau JD, Jin W, Xu R, Timm
MM and Bible KC: Chaetocin: A promising new antimyeloma agent with
in vitro and in vivo activity mediated via imposition of oxidative
stress. Blood. 109:2579–2588. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lai YS, Chen JY, Tsai HJ, Chen TY and Hung
WC: The SUV39H1 inhibitor chaetocin induces differentiation and
shows synergistic cytotoxicity with other epigenetic drugs in acute
myeloid leukemia cells. Blood Cancer J. 5:e3132015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Isham CR, Tibodeau JD, Bossou AR, Merchan
JR and Bible KC: The anticancer effects of chaetocin are
independent of programmed cell death and hypoxia, and are
associated with inhibition of endothelial cell proliferation. Br J
Cancer. 106:314–323. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dixit D, Ghildiyal R, Anto NP and Sen E:
Chaetocin-induced ROS-mediated apoptosis involves ATM-YAP1 axis and
JNK-dependent inhibition of glucose metabolism. Cell Death Dis.
5:e12122014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee YM, Lim JH, Yoon H, Chun YS and Park
JW: Antihepatoma activity of chaetocin due to deregulated splicing
of hypoxia-inducible factor 1α pre-mRNA in mice and in vitro.
Hepatology. 53:171–180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chaib H, Nebbioso A, Prebet T, Castellano
R, Garbit S, Restouin A, Vey N, Altucci L and Collette Y:
Anti-leukemia activity of chaetocin via death receptor-dependent
apoptosis and dual modulation of the histone methyl-transferase
SUV39H1. Leukemia. 26:662–674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Truitt L, Hutchinson C, DeCoteau JF and
Geyer CR: Chaetocin antileukemia activity against chronic
myelogenous leukemia cells is potentiated by bone marrow stromal
factors and overcomes innate imatinib resistance. Oncogenesis.
3:e1222014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vo MC, Nguyen-Pham TN, Lee HJ, Jung SH,
Choi NR, Hoang MD, Kim HJ and Lee JJ: Chaetocin enhances dendritic
cell function via the induction of heat shock protein and cancer
testis antigens in myeloma cells. Oncotarget. 8:46047–46056. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han X, Han Y, Zheng Y, Sun Q, Ma T, Zhang
J and Xu L: Chaetocin induces apoptosis in human melanoma cells
through the generation of reactive oxygen species and the intrinsic
mitochondrial pathway, and exerts its anti-tumor activity in vivo.
PLoS One. 12:e1759502017.
|
|
16
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Coleman RL, Monk BJ, Sood AK and Herzog
TJ: Latest research and treatment of advanced-stage epithelial
ovarian cancer. Nat Rev Clin Oncol. 10:211–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cotter TG: Apoptosis and cancer: The
genesis of a research field. Nat Rev Cancer. 9:501–507. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ferrin G, Linares CI and Muntané J:
Mitochondrial drug targets in cell death and cancer. Curr Pharm
Des. 17:2002–2016. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong Y, Cao A, Shi J, Yin P, Wang L, Ji G,
Xie J and Wu D: Tangeretin, a citrus polymethoxyflavonoid, induces
apoptosis of human gastric cancer AGS cells through extrinsic and
intrinsic signaling pathways. Oncol Rep. 31:1788–1794. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roy J, Galano JM, Durand T, Le Guennec JY
and Lee JC: Physiological role of reactive oxygen species as
promoters of natural defenses. FASEB J. 31:3729–3745. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Holmström KM and Finkel T: Cellular
mechanisms and physiological consequences of redox-dependent
signalling. Nat Rev Mol Cell Biol. 15:411–421. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
D'Autréaux B and Toledano MB: ROS as
signalling molecules: Mechanisms that generate specificity in ROS
homeostasis. Nat Rev Mol Cell Biol. 8:813–824. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Reuter S, Gupta SC, Chaturvedi MM and
Aggarwal BB: Oxidative stress, inflammation, and cancer: How are
they linked? Free Radic Biol Med. 49:1603–1616. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Galadari S, Rahman A, Pallichankandy S and
Thayyullathil F: Reactive oxygen species and cancer paradox: To
promote or to suppress? Free Radic Biol Med. 104:144–164. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Weng MS, Chang JH, Hung WY, Yang YC and
Chien MH: The interplay of reactive oxygen species and the
epidermal growth factor receptor in tumor progression and drug
resistance. J Exp Clin Cancer Res. 37:612018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tafani M, Sansone L, Limana F, Arcangeli
T, De Santis E, Polese M, Fini M and Russo MA: The interplay of
reactive oxygen species, hypoxia, inflammation, and sirtuins in
cancer initiation and progression. Oxid Med Cell Longev.
2016:39071472016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zou P, Xia Y, Ji J, Chen W, Zhang J, Chen
X, Rajamanickam V, Chen G, Wang Z, Chen L, et al: Piperlongumine as
a direct TrxR1 inhibitor with suppressive activity against gastric
cancer. Cancer Lett. 375:114–126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Raj L, Ide T, Gurkar AU, Foley M, Schenone
M, Li X, Tolliday NJ, Golub TR, Carr SA, Shamji AF, et al:
Selective killing of cancer cells by a small molecule targeting the
stress response to ROS. Nature. 475:231–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He J, Chen X, Li B, Zhou W, Xiao J, He K,
Zhang J and Xiang G: Chaetocin induces cell cycle arrest and
apoptosis by regulating the ROS-mediated ASK-1/JNK signaling
pathways. Oncol Rep. 38:2489–2497. 2017. View Article : Google Scholar : PubMed/NCBI
|