Introduction
With the increasing incidence, osteosarcoma (OS) has
become a major threat of human health. OS occurs mostly in
childhood and adolescence. Approximately 80% of OS affect the
growth of the femur and tibia (1).
Previous studies have indicated that malignant transformation of OS
cells is associated with genetic and environmental factors. The
physiological dysfunction of OS cells is mainly caused by changes
in oncogene and tumor suppressor activity (2). Moreover, OS cells are highly
metastatic, including blood flow and lung metastasis. This high
degree of metastasis also leads to worse cure rate and prognosis in
patients with OS (3). Despite the
rapid development of modern diagnostic techniques, the complex
pathological mechanisms of OS remain obstacles to patient survival
(4). Therefore, further exploration
of the pathogenesis of OS and the development of new treatments are
of great significance for patients with OS.
MicroRNAs (miRNAs) are a class of evolutionarily
conserved non-coding small RNAs that function to regulate gene
expression at the translational level. The diversity and breadth of
miRNAs have been revealed in many diseases and cancers in recent
years, including OS. Zhou et al found the diagnostic effect
of miR-139-5p as an indicator in OS (5). Additionally, miR-199b-5p was
upregulated and promoted malignant progression of OS (6). Inversely, downregulation of miR-144 was
found in OS. miR-144 overexpression inhibited tumor growth and
metastasis in OS (7). Although many
miRNAs have been found in OS, the dysregulation of miR-744 has not
been investigated in OS. Moreover, the regulatory mechanism of
miR-744 varies depending on the type of cancer. For instance,
miR-744 was downregulated in glioblastoma and inhibited its
aggressive behavior (8). On the
contrary, miR-744 expression was increased in pancreatic cancer,
which increased its tumorigenicity (9). Furthermore, miR-744 was a potential
prognostic marker in patients with hepatocellular carcinoma
(10). However, the molecular
mechanism of miR-744 remains largely unknown in the pathological
process of OS.
As a member of LATS tumor suppressor family, large
tumor suppressor kinase 2 (LATS2) has been widely investigated in
human cancers. For example, LATS2 was overexpressed in
nasopharyngeal carcinoma and predicted poor prognosis (11). Functionally, LATS2, a putative tumor
suppressor, inhibited G1/S transition (12). Furthermore, LATS2 was found to induce
apoptosis by suppressing Bcl-2 expression (13). Dai et al found that LATS1/2
could inhibit f-actin binding, cell migration, and angiogenesis
(14). The interaction between
miRNAs and LATS2 had been investigated in some cancers. Lee et
al reported that miR-373 post-transcriptionally regulated LATS2
and stimulated proliferation in human esophageal cancer (15). Moreover, miR-103 promoted metastasis
and epithelial-mesenchymal transition (EMT) of hepatocellular
carcinoma by inhibiting LATS2 (16).
However, to the best of our knowledge, the relationship between
miR-744 and LATS2 has not been previously investigated.
Therefore, the regulatory mechanism of miR-744/LATS2
axis was elucidated in our study. Furthermore, the effect of
miR-744 on Wnt/β-catenin pathway and EMT was also investigated in
OS. This investigation will help us understand the pathogenesis of
OS.
Materials and methods
Experimental sample
Forty-one OS patients in Weifang People's Hospital
(Weifang, China) took part in this study. Informed consents of all
OS patients were acquired before the experiment. OS tissues and
normal tissues were acquired from these patients, who had not
received any treatment except for surgery. Permission for this
research was acquired from the Institutional Ethics Committee of
Weifang People's Hospital.
Cell culture and transfection
Human normal osteoblast hFOB1.19 cells
(ATCC® CRL-11372™) and MG-63 OS cell line
(ATCC® CRL-1427™) were selected in this study. The
growth conditions included 5% CO2, 37°C and culture
solution (90% RPMI-1640 + 10% FBS). Next, miR-744 mimics,
inhibitor, LATS2 siRNA and vector (RiboBio) were transfected into
MG-63 cells, respectively, using Lipofectamine 2000.
RT-qPCR
Total RNA extraction was performed by TRIzol reagent
(Sigma-Aldrich; Merck KGaA). The cDNA solution was synthesized
using First-Strand cDNA Synthesis kit (cat no. K1611; Promega
Corporation). The temperature conditions of the reverse
transcription were as follows: 37°C for 15 min and 85°C for 5 sec.
We performed RT-qPCR assay using miScript SYBR®-Green
PCR kit (cat. no. /ID: 218073; Qiagen, Inc.) based on the
manufacturer's instructions. The thermocycling parameters were as
follows: 95°C for 3 min and 40 cycles of 95°C for 15 sec followed
by 58°C for 30 sec. The 2−∆∆Cq method (17) was applied to measure miR-744 or LATS2
expression levels using internal reference U6 or GAPDH. The
following primers were used: miR-744 forward,
5′-ACACTCCAGCTGGGTGCGGGGCTAGGGCTAAC-3′ and reverse,
5′-CTCAACTGGTGTCGTGGA-3′; LATS2 forward,
5′-ATGAGCTCCACTCTGCTCAATGTCACGG-3′ and reverse,
5′-GCAAGCTTCTCTACCAAGAATGAAAGAGCAT-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
GAPDH forward, 5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse,
5′-GAGATGGTGATGGGATTTC-3′.
Western blot analysis
Transfected MG-63 cells were dissociated using RIPA
lysis buffer. Next, 10% SDS-PAGE was used to separate 25 µg
protein. Protein concentration was calculated using bicinchoninic
acid (BCA). Protein samples were transferred into a polyvinylidene
difluoride (PVDF) membrane (Thermo Fisher Scientifc, Inc.). Then,
the membranes were blocked with 5% non-fat milk for 1 h at room
temperature. Protein samples were incubated with vimentin (rabbit
polyclonal antibody; dilution, 1:1,000; cat. no. ab137321; Abcam),
N-cadherin (rabbit polyclonal antibody; dilution, 1:1,000; cat. no.
ab18203; Abcam), E-cadherin (rabbit monoclonal antibody; dilution,
1:1,000; cat. no. ab1416; Abcam), LATS2 (rabbit polyclonal
antibody; dilution, 1:1,000; cat. no. ab110780; Abcam), β-catenin
(rabbit polyclonal antibody; dilution, 1:1,000; cat. no. ab6302;
Abcam) and GAPDH (rabbit monoclonal primary antibody; dilution,
1:1,000; cat. no. ab181602; Abcam) overnight at 4°C. Secondary
mouse anti-rabbit (cat. no. 3678) or goat anti-mouse (cat. no.
58802) antibodies (dilution, 1:2,000) conjugated with horseradish
peroxidase (Cell Signaling Technologies, Inc.) were added to
incubate protein samples for 1 h at room temperature. Finally, ECL
(ECL, Pierce) was used to measure protein expression levels.
MTT assay
The MG-63 cell suspension was cultured in RPMI-1640
medium containing 10% FBS. Then, 96-well plates were seeded at a
density of 4,000 cells per well. After 24, 48, 72 and 96 h of
incubation at 37°C, 20 µl of MTT solution (5 mg/ml in PBS) was
added to each well. Incubation was continued for 4 h at 37°C and
the culture was terminated. The supernatant was discarded, 150 µl
of DMSO was added to each well. The mixture was shaken for 10 min
to allow the crystals to fully melt. Then, they were dissolved in
100 µl of dimethyl sulfoxide [CAS: (67-68-5), Sangon Biotech].
Finally, at a wavelength of 490 nm the light absorption value of
each well was measured on a microplate reader (ELx800; Bio-Tek
Instruments) to plot a cell growth curve.
Transwell assay
First, 60 µl of diluted Matrigel (3.9 µg/µl) was
added to the upper chamber for cell invasion. Cell migration assay
was performed without Matrigel. After 30 min, MG-63 cell suspension
(5×103 cells/well) was added to the Transwell upper
chamber, and 500 µl of RPMI-1640 medium (10% FBS) was added to the
24-well plate in the lower chamber. After routine incubation for 24
h, the cells on the lower surface of the membrane were fixed with
4% paraformaldehyde [CAS: (30525-89-4), Sangon Biotech] and stained
with 1% crystal violet (Solarbio) for 5 min. Observation and
photographing were performed using a light microscope (Olympus
Corporation).
Luciferase reporter assay
Luciferase vector pcDNA3.1 (Promega Corporation)
with Wt-LATS2-3′UTR or Mut-LATS2-3′UTR and miR-744 mimics were
transfected into MG-63 OS cell line (ATCC). Next, the transfected
cells were incubated in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) at room temperature for 20 min. After 48 h, the
medium was discarded and washed with PBS. Finally, luciferase
activity was assessed using dual luciferase assay system (Promega
Corporation). This experiment was repeated 3 times.
Statistical analysis
Data were analyzed by SPSS 18.0 (SPSS, Inc.) or
GraphPad Prism 6 (GraphPad Software, Inc.), and shown as mean ± SD.
Chi-square test, one-way ANOVA with Tukey's multiple comparison
tests and univariate Kaplan-Meier method with log-rank test were
used to analyze differences between the groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-744 expression is increased in
OS
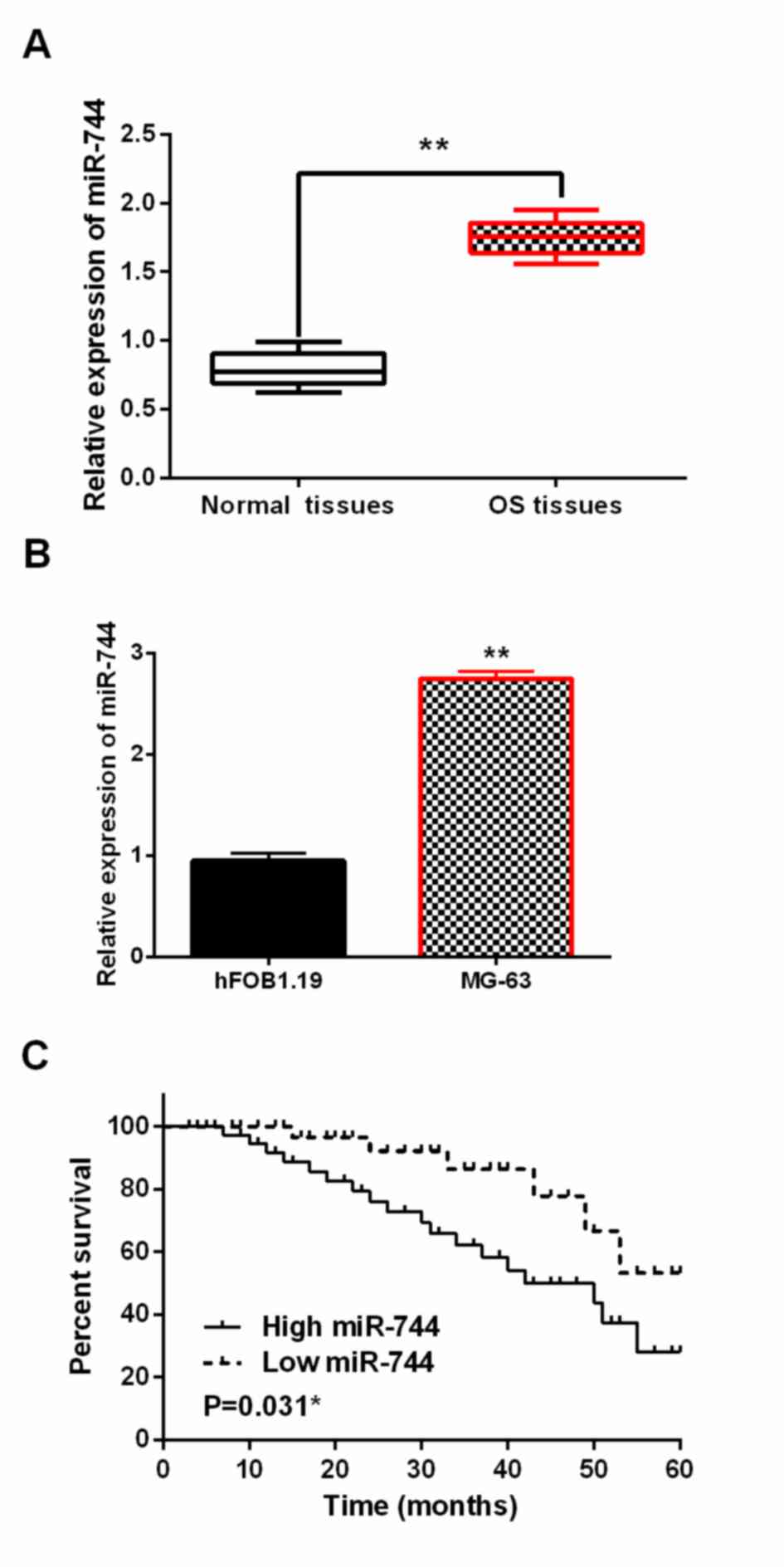
The alteration of miR-744 expression was assessed in
OS by RT-qPCR. We found that miR-744 expression was higher in OS
tissues than in normal tissues (P<0.01, Fig. 1A). Moreover, miR-744 was also
upregulated in MG-63 cells compared to hFOB1.19 cells (P<0.01,
Fig. 1B). In addition, dysregulation
of miR-744 affected the prognosis of OS patients. Upregulation of
miR-744 was closely associated with worse prognosis in OS patients
(P<0.05, Fig. 1C). Besides,
abnormal miR-744 expression was associated with distant metastasis
and clinical stage (P<0.05, Table
I). These findings implied that miR-744 might influence OS
progression.
 | Table I.Relationship between miR-744
expression and the clinicopathological characteristics of OS
patients. |
Table I.
Relationship between miR-744
expression and the clinicopathological characteristics of OS
patients.
|
|
| miR-744 |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Cases | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.75 |
| ≥60 | 25 | 18 | 7 |
|
|
<60 | 16 | 14 | 2 |
|
| Sex |
|
|
| 0.52 |
| Male | 24 | 19 | 5 |
|
|
Female | 17 | 13 | 4 |
|
| Tumor size |
|
|
| 0.12 |
| <5
cm | 20 | 13 | 7 |
|
| ≥5
cm | 21 | 19 | 2 |
|
| Clinical stage |
|
|
| 0.02a |
|
I–II | 32 | 25 | 7 |
|
|
III–IV | 9 | 7 | 2 |
|
| Distant
metastasis |
|
|
| 0.03a |
| No | 35 | 29 | 6 |
|
|
Yes | 6 | 3 | 3 |
|
miR-744 promotes cell viability and
metastasis in OS
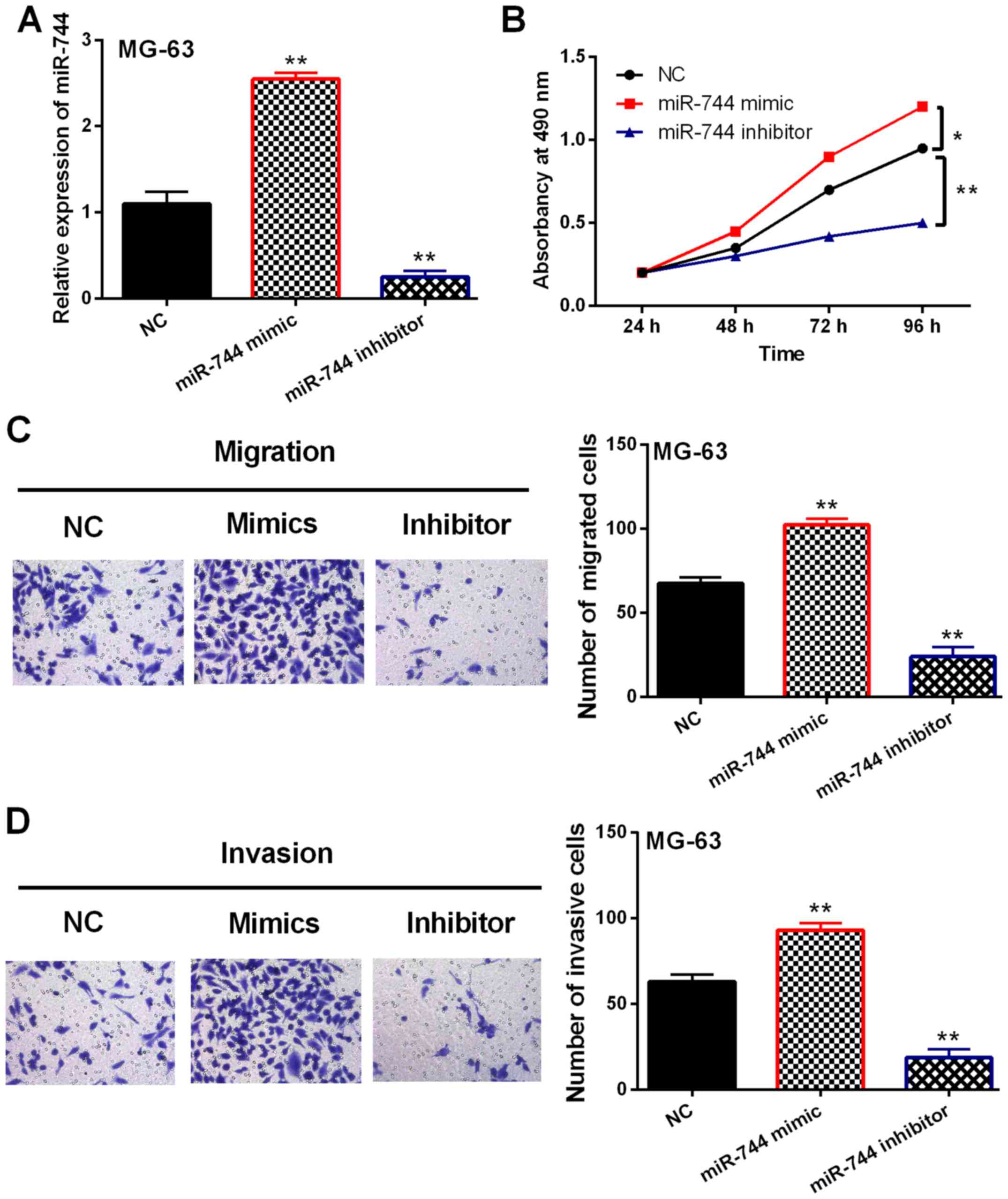
In order to confirm whether miR-744 was involved in
OS development, gain-loss experiment of miR-744 was performed in
MG-63 cells. After transfection, miR-744 expression was enhanced by
its mimics and reduced by its inhibitor (P<0.01, Fig. 2A). Next, miR-744 mimic overexpression
was found to promote cell proliferation in OS. Correspondingly,
knockdown of miR-744 inhibitor restrained MG-63 cell proliferation
(P<0.05 or 0.01, Fig. 2B).
Consistently, cell migration was also promoted by miR-744 mimic
overexpression and inhibited by downregulation of miR-744 inhibitor
(P<0.01, Fig. 2C). Similarly,
miR-744 mimics promoted MG-63 cell invasion. miR-744 inhibitor
restrained cell invasion in OS cells (P<0.01, Fig. 2D). Hence, miR-744 played a
carcinogenic role in OS development.
miR-744 induces Wnt/β-catenin pathway
and EMT in OS
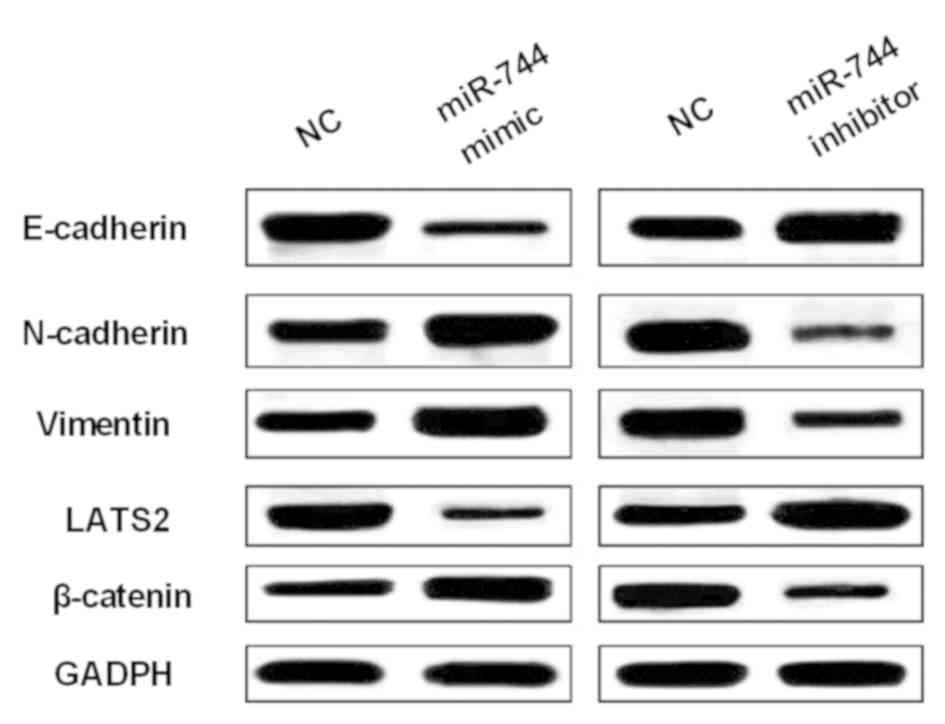
Next, how miR-744 regulated Wnt/β-catenin pathway
and EMT was explored to confirm its effect on OS cell viability and
metastasis. The expression levels of EMT makers (E-cadherin,
N-cadherin and vimentin) were measured in MG-63 cells with miR-744
mimics or inhibitor. We found that miR-744 mimics promoted
N-cadherin and vimentin expression levels and restrained E-cadherin
expression in MG-63 cell (Fig. 3).
Correspondingly, miR-744 inhibitor enhanced E-cadherin expression
and blocked N-cadherin and vimentin expression levels in OS cells
(Fig. 3). In addition, we examined
β-catenin expression as a regulator in Wnt/β-catenin pathway in
MG-63 cells with miR-744 mimics or inhibitor. The expression of
β-catenin was promoted by miR-744 mimics and suppressed by miR-744
inhibitor (Fig. 3). These findings
indicated that miR-744 promoted OS development through activating
Wnt/β-catenin pathway and EMT.
miR-744 negatively regulates LATS2
expression in OS
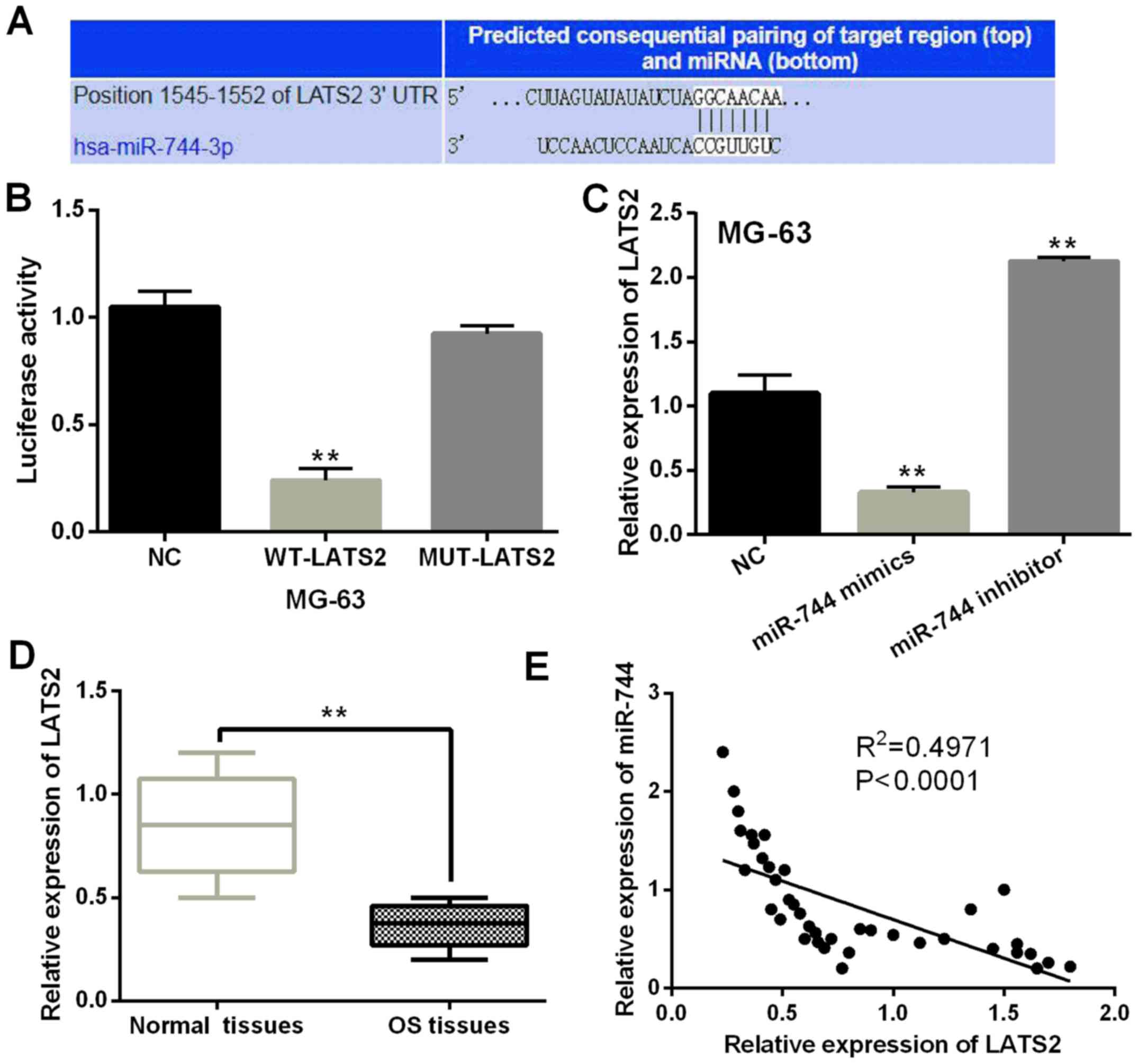
TargetScan database (http://www.targetscan.org/, Fig. 4A) showed that there were binding
sites between LATS2 and miR-744 and the prediction was confirmed by
luciferase reporter assay. miR-744 mimics lessened luciferase
activities of wild LATS2, but had no effect on mutant LATS2
(P<0.01, Fig. 4B). LATS2
expression affected by miR-744 mimics or inhibitor was detected in
MG-63 cells. LATS2 expression was found to be blocked by miR-744
mimics and increased by miR-744 inhibitor (P<0.01, Fig. 4C). Downregulation of LATS2 was found
in OS tissues compared to normal tissues (P<0.01, Fig. 4D). Furthermore, miR-744 was found to
have a negative association with LATS2 expression levels in OS
tissues (P<0.0001, R2=0.4971; Fig. 4E). Based on these results, miR-744
directly targeted LATS2 and blocked its expression in OS.
miR-744 accelerates OS progression
through targeting LATS2
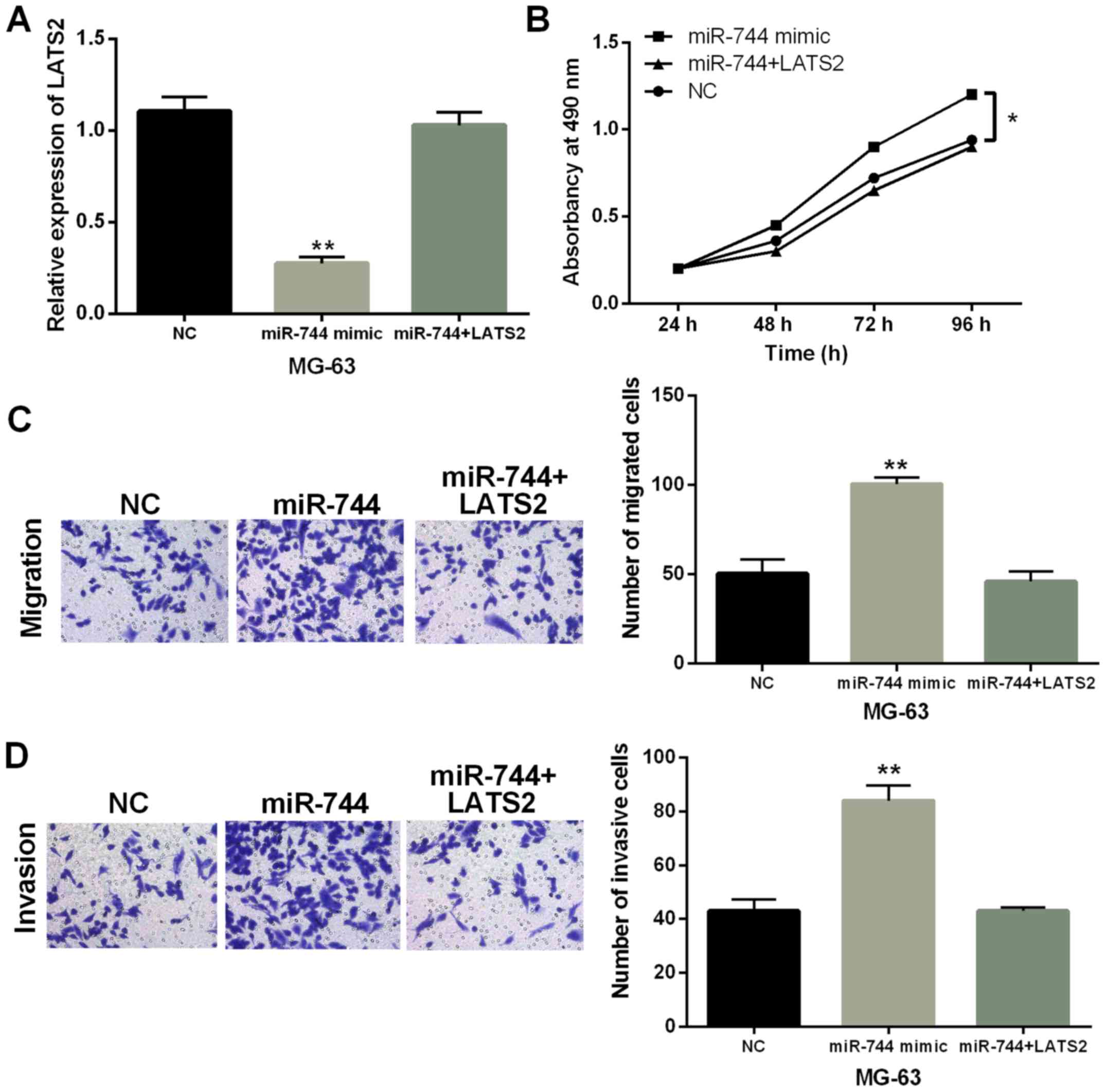
Finally, MG-63 cells with miR-744 mimics were
transfected with LATS2 vector to explore their interaction in OS.
We found that miR-744 mimics mediated reduction of LATS2 expression
which was recovered by LATS2 vector (Fig. 5A). Functionally, promotion of cell
proliferation induced by miR-744 overexpression was impaired by
upregulation of LATS2 (P<0.01, Fig.
5B). Similarly, promotion of cell migration and invasion
regulated by miR-744 was also weakened by LATS2 upregulation
(P<0.01, Fig. 5C and D).
Combining these results, miR-744 was considered to accelerate OS
progression through targeting LATS2.
Discussion
As important regulators, miRNAs have been found to
take part in the regulation of OS progress. Similarly to our
results, miR-214 expression was increased in OS and promoted its
malignant behavior (18). Li et
al demonstrated that miR-374a activated Wnt/β-catenin signaling
to promote OS development (19).
Here, upregulation of miR-744 was also measured in OS. Furthermore,
this dysregulation was associated with worse clinical features and
prognosis of OS patients. Functionally, promotion of OS cell
proliferation, migration and invasion mediated by miR-744 was
identified. Moreover, miR-744 activated Wnt/β-catenin pathway and
EMT to exhibit carcinogenesis in OS. Besides, we found that miR-744
accelerated OS progression through targeting LATS2. These findings
implied that miR-744 acted as an oncogene in OS.
Consistent with our results, high expression of
miR-744 was observed in nasopharyngeal carcinoma, which predicts
poor prognosis (20). Moreover,
similar results were also identified in pancreatic cancer (21). In addition, Li et al showed
that miR-744 enhanced motility of SiHa cervical cancer cells
(22), which agreed with our
findings. At the same time, oncogenic miR-744 was found to promote
prostate cancer growth (23). In our
study, we also found miR-744 mediated promotion of cell viability
in OS. It was reported that miR-744 promoted prostate cancer
progression through aberrantly activating Wnt/β-catenin signaling
(24). Similarly, Wnt/β-catenin
pathway was also activated by miR-744 in OS, but the activation of
EMT induced by miR-744 has not been investigated in previous
studies. Previous studies have indicated that miR-744 carries out
its functions by regulating the expression levels of its targets,
such as KLLN and EEF1A2 (25,26).
Therefore, we investigated the interaction of LATS2 and miR-744 in
this study. miR-744 was found to directly target LATS2 and block
its expression in OS.
As a target gene, the expression of LATS2 is
affected by other miRNAs in different cancers, such as miR-93 and
miR-372 (27,28). As a tumor suppressor, dysregulation
of LATS2 has been assessed in many malignant tumors. Downregulation
of LATS2 was detected in colorectal cancer and esophageal squamous
cell carcinoma (29,30). In the current study, LATS2 was also
downregulated in OS. Furthermore, LATS2 had a negative association
with miR-744 expression in OS tissues, which was consistent with
previous studies. Functionally, miR-25 promoted ovarian cancer
proliferation and motility by targeting LATS2 (31). Moreover, miR-650 promoted metastasis
and EMT of hepatocellular carcinoma by inhibiting LATS2 expression
(32). Here, miR-744 was also
considered to facilitate cell proliferation and metastasis in OS
through targeting LATS2.
Collectively, miR-744 expression was increased in
OS, which predicted poor clinical features and prognosis. Moreover,
miR-744 accelerated cell viability and metastasis in OS through
downregulating LATS2. Furthermore, miR-744 activated Wnt/β-catenin
pathway and EMT to facilitate OS progression. Therefore, our study
provides a new therapeutic target for OS patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS contributed to the study design, data acquisition
and analysis and drafted the manuscript. ML performed RT-qPCR and
western blot analysis. SL and YS were responsible for MTT,
Transwell and dual-luciferase reporter assays. QW contributed to
analysis of observation indexes. All the authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Institutional Ethics
Committee of Weifang People's Hospital (Weifang, China). Patients
who participated in this research, had complete clinical data.
Signed informed consents were obtained from the patients or the
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Clark JC, Dass CR and Choong PF: A review
of clinical and molecular prognostic factors in osteosarcoma. J
Cancer Res Clin Oncol. 134:281–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Geller DS and Gorlick R: Osteosarcoma: A
review of diagnosis, management, and treatment strategies. Clin Adv
Hematol Oncol. 8:705–718. 2010.PubMed/NCBI
|
|
3
|
Cortini M, Avnet S and Baldini N:
Mesenchymal stroma: Role in osteosarcoma progression. Cancer Lett.
405:90–99. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsiambas E, Fotiades PP, Sioka C,
Kotrotsios D, Gkika E, Fotopoulos A, Mastronikolis SN, Armata IE,
Giotakis E and Ragos V: Novel molecular and metabolic aspects in
osteosarcoma. J BUON. 22:1595–1598. 2017.PubMed/NCBI
|
|
5
|
Zhou L, Ma X, Yue J, Chen T, Wang XY, Wang
ZW, Pan J and Lin Y: The diagnostic effect of serum miR-139-5p as
an indicator in osteosarcoma. Cancer Biomark. 23:561–567. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Z, Zhao G, Zhang Y, Ma Y, Ding Y and
Xu N: MiR-199b-5p promotes malignant progression of osteosarcoma by
regulating HER2. J BUON. 23:1816–1824. 2018.PubMed/NCBI
|
|
7
|
Liu JL, Li J, Xu JJ, Xiao F, Cui PL, Qiao
ZG, Chen XD, Tao WD and Zhang XL: MiR-144 inhibits tumor growth and
metastasis in osteosarcoma via dual-suppressing RhoA/ROCK1
signaling pathway. Mol Pharmacol. 95:451–461. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Deng Y, Li Y, Fang Q, Luo H and Zhu G:
microRNA-744 is downregulated in glioblastoma and inhibits the
aggressive behaviors by directly targeting NOB1. Am J Cancer Res.
8:2238–2253. 2018.PubMed/NCBI
|
|
9
|
Zhou W, Li Y, Gou S, Xiong J, Wu H, Wang
C, Yan H and Liu T: MiR-744 increases tumorigenicity of pancreatic
cancer by activating Wnt/β-catenin pathway. Oncotarget.
6:37557–37569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan YL, Bai ZG, Zou WL, Ma XM, Wang TT,
Guo W, Liu J, Li JS, Jie-Yin, Zang YJ, et al: miR-744 is a
potential prognostic marker in patients with hepatocellular
carcinoma. Clin Res Hepatol Gastroenterol. 39:359–365. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Hu CF, Chen J, Yan LX, Zeng YX
and Shao JY: LATS2 is de-methylated and overexpressed in
nasopharyngeal carcinoma and predicts poor prognosis. BMC Cancer.
10:5382010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Pei J, Xia H, Ke H, Wang H and Tao
W: Lats2, a putative tumor suppressor, inhibits G1/S transition.
Oncogene. 22:4398–4405. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ke H, Pei J, Ni Z, Xia H, Qi H, Woods T,
Kelekar A and Tao W: Putative tumor suppressor Lats2 induces
apoptosis through downregulation of Bcl-2 and Bcl-x(L). Exp Cell
Res. 298:329–338. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai X, She P, Chi F, Feng Y, Liu H, Jin D,
Zhao Y, Guo X, Jiang D, Guan KL, et al: Phosphorylation of
angiomotin by Lats1/2 kinases inhibits F-actin binding, cell
migration, and angiogenesis. J Biol Chem. 288:34041–34051. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee KH, Goan YG, Hsiao M, Lee CH, Jian SH,
Lin JT, Chen YL and Lu PJ: MicroRNA-373 (miR-373)
post-transcriptionally regulates large tumor suppressor, homolog 2
(LATS2) and stimulates proliferation in human esophageal cancer.
Exp Cell Res. 315:2529–2538. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han LL, Yin XR and Zhang SQ: miR-103
promotes the metastasis and EMT of hepatocellular carcinoma by
directly inhibiting LATS2. Int J Oncol. 53:2433–2444.
2018.PubMed/NCBI
|
|
17
|
Livak KJ and Scmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cai H, Miao M and Wang Z: miR-214-3p
promotes the proliferation, migration and invasion of osteosarcoma
cells by targeting CADM1. Oncol Lett. 16:2620–2628. 2018.PubMed/NCBI
|
|
19
|
Li W, Meng Z, Zou T, Wang G, Su Y, Yao S
and Sun X: MiR-374a activates Wnt/β-catenin signaling to promote
osteosarcoma cell migration by targeting WIF-1. Pathol Oncol Res.
Dec 6–2018.(Epub ahead of print). View Article : Google Scholar
|
|
20
|
Yu Q, Zhang F, Du Z and Xiang Y:
Up-regulation of serum miR-744 predicts poor prognosis in patients
with nasopharyngeal carcinoma. Int J Clin Exp Med. 8:13296–13302.
2015.PubMed/NCBI
|
|
21
|
Miyamae M, Komatsu S, Ichikawa D,
Kawaguchi T, Hirajima S, Okajima W, Ohashi T, Imamura T, Konishi H,
Shiozaki A, et al: Plasma microRNA profiles: Identification of
miR-744 as a novel diagnostic and prognostic biomarker in
pancreatic cancer. Br J Cancer. 113:1467–1476. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li C, Jia L, Yu Y and Jin L: Lactic acid
induced microRNA-744 enhances motility of SiHa cervical cancer
cells through targeting ARHGAP5. Chem Biol Interact. 298:86–95.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang M and Li H, Zhang Y and Li H:
Oncogenic miR-744 promotes prostate cancer growth through direct
targeting of LKB1. Oncol Lett. 17:2257–2265. 2019.PubMed/NCBI
|
|
24
|
Guan H, Liu C, Fang F, Huang Y, Tao T,
Ling Z, You Z, Han X, Chen S, Xu B, et al: MicroRNA-744 promotes
prostate cancer progression through aberrantly activating
Wnt/β-catenin signaling. Oncotarget. 8:14693–14707. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang C, Zong J, Li Y, Wang X, Du W and Li
L: MiR-744-3p regulates keratinocyte proliferation and
differentiation via targeting KLLN in psoriasis. Exp Dermatol.
28:283–291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vislovukh A, Kratassiouk G, Porto E,
Gralievska N, Beldiman C, Pinna G, El'skaya A, Harel-Bellan A,
Negrutskii B and Groisman I: Proto-oncogenic isoform A2 of
eukaryotic translation elongation factor eEF1 is a target of
miR-663 and miR-744. Br J Cancer. 108:2304–2311. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fang L, Du WW, Yang W, Rutnam ZJ, Peng C,
Li H, O'Malley YQ, Askeland RW, Sugg S, Liu M, et al: MiR-93
enhances angiogenesis and metastasis by targeting LATS2. Cell
Cycle. 11:4352–4365. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cho WJ, Shin JM, Kim JS, Lee MR, Hong KS,
Lee JH, Koo KH, Park JW and Kim KS: miR-372 regulates cell cycle
and apoptosis of ags human gastric cancer cell line through direct
regulation of LATS2. Mol Cells. 28:521–527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng YB, Xiao K, Xiao GC, Tong SL, Ding
Y, Wang QS, Li SB and Hao ZN: MicroRNA-103 promotes tumor growth
and metastasis in colorectal cancer by directly targeting LATS2.
Oncol Lett. 12:2194–2200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao Y, Yi J, Zhang K, Bai F, Feng B, Wang
R, Chu X, Chen L and Song H: Downregulation of MiR-31 stimulates
expression of LATS2 via the hippo pathway and promotes
epithelial-mesenchymal transition in esophageal squamous cell
carcinoma. J Exp Clin Cancer Res. 36:1612017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Feng S, Pan W, Jin Y and Zheng J: MiR-25
promotes ovarian cancer proliferation and motility by targeting
LATS2. Tumour Biol. 35:12339–12344. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han LL, Yin XR and Zhang SQ: miR-650
promotes the metastasis and epithelial-mesenchymal transition of
hepatocellular carcinoma by directly inhibiting LATS2 expression.
Cell Physiol Biochem. 51:1179–1192. 2018. View Article : Google Scholar : PubMed/NCBI
|