Introduction
T-cell acute lymphoblastic leukemia (T-ALL), a
highly aggressive hematological malignancy, carries an increased
risk of treatment failure and relapse compared with B-cell acute
lymphoblastic leukemia (B-ALL) (1).
The majority of studies investigating natural killer (NK) cell
recognition in ALL have focused on B-ALL (2,3), while a
limited number of studies have investigated the role of NK cells in
T-ALL. Thus, there is a requirement for studies surrounding the
mechanism of immune surveillance escape in T-ALL, with the aim to
improve the success of therapeutic strategies.
NK cells are important innate immune surveyors that
are critical for the removal of leukemia blasts. It has been
reported that following bone marrow transplantation, NK cells are
able to effectively destroy leukemia blasts in patients with acute
myeloid leukemia (AML) (4). However,
the NK response to treatments for T-ALL is limited (5,6). Factors
that affect immune surveillance by NK cells are characterized as
immune escape strategies used by leukemia blasts, including the
downregulation of NK cell cytotoxicity and activation receptors,
upregulation of inhibitory NK cell receptors and secretion of
soluble NK-inhibitory factors (7,8). In ALL,
it has been reported that T-ALL blasts downregulate cell-surface
ligands that activate NK cell receptors, facilitating immune
surveillance escape (5,6). NK cells exist in distinct functional
subsets that are characterized by their surface antigen profiles.
Further investigation of the NK cell maturation phenotype in T-ALL
is required, as to date, the effects of maturation arrest in T-ALL
have not been evaluated.
The murine NK cell maturation process consists of
four stages: CD27−CD11b− →
CD27+CD11b− →
CD27+CD11b+ →
CD27−CD11b+ (9); Murine CD11b+ mNK subsets can
be further categorized into CD27+ and CD27−
populations. CD11b+CD27+ NK cells produce a
greater range of cytokines and exhibit increased cytotoxicity
compared with their CD27− counterparts (10). Human mature NK subsets are
characterized as CD56brightCD16dim or
CD56dimCD16bright (11).
CD56brightCD16dim NK cells are the precursors
to CD56dimCD16bright NK cells, which
predominantly reside in secondary lymphoid tissue, produce a wider
range of cytokines, including interferon γ (IFNγ), tumor necrosis
factor-β, interleukin (IL)-10, IL-13 and granulocyte-macrophage
colony-stimulating factor, but are less cytotoxic. By contrast,
CD56dimCD16bright NK cells, enriched in the
peripheral blood, produce fewer cytokines but are more cytotoxic.
As a result, CD56brightCD16dim NK cells are
the primary IFNγ-producing subset (12–14). The
majority of NK-mediated anti-leukemia activity is the result of
IFNγ production, which is partially attributed to the upregulation
of natural killer receptor group 2, member D (NKG2D) (15), and which in turn activates NK cells
via the DAP10 and/or DAP12 signaling pathways (16).
MicroRNAs (miRs) serve an important role in
regulating the development and functions of NK cells (17,18).
However, the importance of miRs in regulating NK cells in T-ALL has
not been investigated. Due to >50% of human T-ALL exhibiting
activating neurogenic locus notch homolog protein 1 (Notch1)
mutations (19), the present study
demonstrated the effects of miR-29b on the dysregulation of NK cell
phenotype and function in Notch1-T-ALL mice. The selective
downregulation of CD27+CD11b+NK cell subset
was revealed, in addition to a reduction in IFN-γ secretion and
NKG2D expression on NK cells. Furthermore, the upregulation of
miR-29b expression level was observed, which may have regulatory
effects on NK cell development.
Materials and methods
Animals
Wild type (WT) C57BL/6, C57BL/6 Rag2−/−
and C57BL/6 miR29ab1−/−mice were bred in pathogen-free
conditions at the animal center affiliated to the First Bethune
Hospital (Changchun, China). Following experimentation, the animals
were sacrificed by cervical dislocation; the thumb and index finger
were placed on either side of the neck at the base of the skull.
With the other hand, the base of the tail was quickly pulled,
causing separation of the cervical vertebrae from the skull. Death
was verified by the lack of continuous spontaneous breathing or
blinking reflex.
Notch1-T-ALL mice and NK cell
infusion
NK cells were prepared as previously described
(20). Notch1-T-ALL blasts were
prepared and injected into mice as previously described (21). The Notch1-T-ALL blasts used in the
present study contained >95% leukemia (GFP+) cells. C57BL/6 mice
or C57BL/6 Rag2−/− mice received 2×105
Notch1-T-ALL blasts. In the NK cell transfusion experiment,
2×107 WT NK cells or miR-29b knock-out (KO) NK cells
were injected into C57BL/6 Rag2−/− mice.
Patients
Fresh peripheral blood samples with CD7+
T-ALL blasts were obtained from 8 newly diagnosed T-ALL patients (5
males and 3 females), aged 20–56 years old, in the Hematology
department of the First Bethune Hospital between March 2017 and
September 2017. Fresh peripheral blood of the age-and sex-matched
healthy control group was obtained from volunteers at the hospital
in the same time period.
Cell isolation
Fresh peripheral blood was layered onto Ficoll (cat.
no. 171440–02; GE Healthcare) and centrifuged at 400 × g for 10 min
at room temperature. NK cells, CD3+ T cells and
CD7+ T-ALL blasts were collected from the single-cell
suspension and sorted for immunophenotypic analysis or quantitative
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis as described below. The NK cell subset was
enriched using the human NK cell isolation kit (cat. no. 17955;
STEMCELL Technologies), and CD3+ T cells were enriched
using the human CD3+ T-cell isolation kit (cat. no.
17951; STEMCELL Technologies) according to the manufacturer's
recommendations. Human T-ALL blasts were enriched by positive
selection from the CD3− population, using anti-CD7 (cat.
no. 239299; Abcam), anti-CD45 (cat. no. 40763; Abcam) and anti-CD34
(cat. no. 157304; Abcam) antibodies. The procedure was carried out
according to the manufacturer's recommendations of the fluorescence
activated cell sorter (FACS) Aria™ III (BD Biosciences).
CD56brightCD16dim or
CD56dimCD16bright NK cell subsets were also
isolated according to these recommendations. Mouse mononuclear
cells from the bone marrow, spleen and peripheral blood were
collected according to a previous study (21). NK1.1+CD3− cells
were enriched using the murine NK cell isolation kit (cat. no.
17855; Stem Cell Technologies) and CD3+ T cells were
enriched with the mouse T cell isolation kit (cat. no. 19851; Stem
Cell Technologies) as per the manufacturer's protocols. GFP+ murine
Notch1-T-ALL blasts were isolated using the FACS Aria™ III sorter
(BD Biosciences). All sorted cells were identified to be >99%
pure using the FACS Canto™ II flow cytometer (BD Biosciences).
Antibodies and flow cytometry
Mononuclear cells from humans or mice were prepared
as above. Antibodies purchased from BD Biosciences included: mouse
anti-mouse NK1.1-APC (cat. no. 550627), rat anti-mouse CD3-PE (cat.
no. 565643), rat anti-mouse CD11b-V450 (cat. no. 560455), mouse
anti-human CD56-Alexa Fluor® (cat. no. 557919), mouse
anti-human CD3-PerCP (cat. no. 347,344), mouse anti-human
CD16-PE-cy7-A (cat. no. 557744), mouse anti-human CD34-PE-CF594
(cat. no. 550761), mouse anti-human-CD45-BV650 (cat. no. 563717)
and mouse anti-human-CD7-APC (cat. no. 561604).
Anti-mouse/rat/human CD27-PECy7 (cat. no. 124215) was purchased
from BioLegend, Inc.; rat anti-mouse IFNγ-APC (cat. no. 554413) and
isotype control mouse IgG2a (cat. no. 551414) were obtained from BD
Pharmingen (BD Biosciences). Notch1-T-ALL cells were identified as
GFP+.
Analysis of intracellular IFNγ
expression
Intracellular production of IFNγ was analyzed using
rat anti-mouse IFNγ-APC, as previously reported (16). Briefly, freshly isolated splenocytes
from Notch1-T-ALL mice and control C57BL/6 mice were labeled with
anti-NK1.1 and anti-CD3 antibodies for 6 h; followed by fixation,
permeabilization, and labeling with anti-IFNγ antibodies. Cells
were subsequently evaluated using the FACS Canto™ II flow cytometer
(BD Biosciences).
Cell lines and cytotoxicity
assays
The EL4 cell line (C57BL/6 origin) was obtained from
the laboratory of the Translational Medicine Institution in Jilin
University, and was thawed from a frozen stock. EL4 cells were
cultured in RPMI 1640 (cat. no. 11875085; Thermo Fisher Scientific,
Inc.) supplemented with 100 U/ml penicillin (cat. no. 15140163;
Thermo Fisher Scientific, Inc.), 10% fetal bovine serum (cat. no.
10437028; Thermo Fisher Scientific, Inc.), 100 µg/ml streptomycin
(cat. no. 15070063; Thermo Fisher Scientific, Inc.) and 50 µmol/l
2-mercaptoethanol (cat. no. 21985023; Thermo Fisher Scientific,
Inc.), at 37°C with 5% CO2. EL4 and Notch1-T-ALL blasts
were cultured with NK cells in standard 4-h chromium release assay
as reported previously (22).
RT-qPCR
Total RNA was isolated from NK cells, Notch1-T-ALL
blasts and T-ALL blasts using the RNAqueous™ Total RNA Isolation
kit (cat. no. AM1912; Invitrogen; Thermo Fisher Scientific, Inc.,),
followed by reverse transcription using the High-Capacity cDNA
Reverse Transcription kit (cat. no. 4368814; Applied Biosystems;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocols. qPCR was performed using the SYBR® Green
master mix (cat. no. 4334973; Applied Biosystems; Thermo Fisher
Scientific, Inc.) on a QuantStudio™ 6 Flex Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The primers
for murine and human IFNγ, NKG2D and GAPDH are listed in Table I. miR-29b expression was detected
using the murine and human microRNA-29b TaqMan™ microRNA assay
(cat. no. 4427975 (000413); Thermo Fisher Scientific, Inc.), the
murine U6 control TaqMan™ microRNA assay (cat. no. 4427975
(001973); Thermo Fisher Scientific, Inc.), and the human RNU6B
control TaqMan™ microRNA assay (cat. no. 4427975 (001093); Thermo
Fisher Scientific, Inc.). The thermocycling conditions were as
follows: 95°C for 2 min; 40 cycles of 95°C for 15 sec, 60°C for 15
sec and 68°C for 30 sec. Gene expression was normalized to an
internal control and the relative mRNA expression level of each
gene was determined using the as 2−ΔΔCq method (17).
 | Table I.Primers for reverse
transcription-quantitative PCR. |
Table I.
Primers for reverse
transcription-quantitative PCR.
| Name | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Human NKG2D |
TTGATGGGGTGGATTCGTGG |
TCTCGGTTGGCAGTGTTACC |
| Human IFNγ, |
AGCTCTGCATCGTTTTGGGT |
CGCTTCCCTGTTTTAGCTGC |
| Mouse NKG2D, |
AGCCAGCAAAGTGGGATACT |
CTGGGACTTCCTTGTTGCAC |
| Mouse IFNγ, |
ACGGCACAGTCATTGAAAGC |
CGAATCAGCAGCGACTCCTT |
| Human GAPDH |
GAAGGTGAAGGTCGGAGT |
CATGGGTGGAATCATATTGGAA |
| Mouse GAPDH |
GGCAAATTCAACGGCACAGT |
TAGGGCCTCTCTTGCTCAGT |
Statistical analysis
The unpaired Student's t-test was used to statically
analyze the differences between two independent groups. Multiple
comparisons between groups were performed using one-way ANOVA
followed by the S-N-K method. The flow cytometry results were
analyzed using FlowJo 10.0.7 software (FlowJo LLC), and the RT-qPCR
data was analyzed using SPSS 12.0 (SPSS, Inc.). Survival rates were
analyzed using Kaplan-Meier analysis and the log-rank test of Prism
5 (GraphPad Software, Inc.). Data are presented as the mean ±
standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
Alterations to miR-29b expression
level in NK cells and leukemia blasts in T-ALL
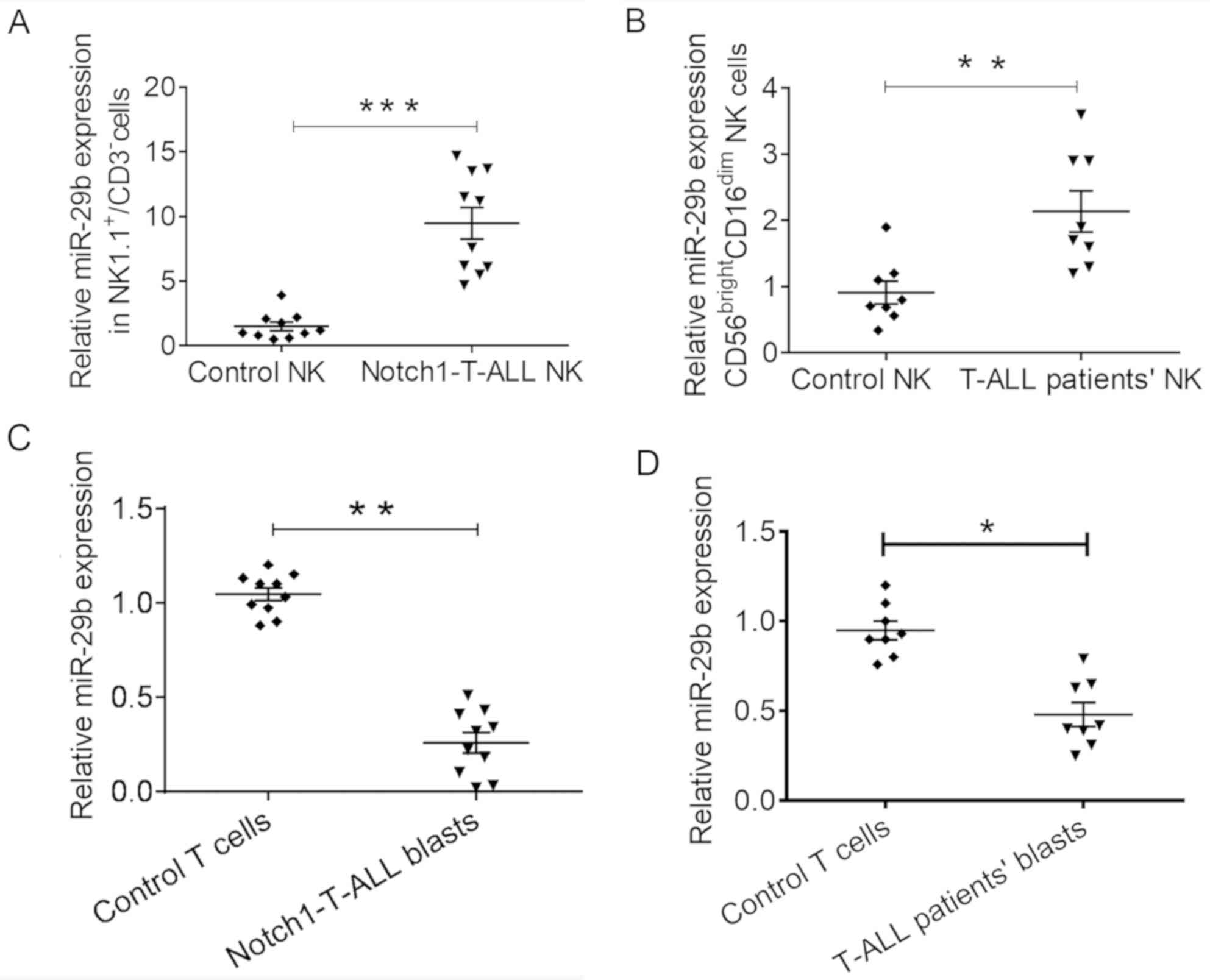
An increase in the expression levels of miR-29b in
NK cells has previously been associated with an aggressive disease
phenotype (21,23). Specifically, a recent study reported
that increased levels of miR-29b expression influenced the
differentiation and function of NK cells in AML (17). To determine whether miR-29b
dysregulation also occurred in T-ALL, the expression levels of
miR-29b were determined in mouse NK cells 21 days after
Notch1-T-ALL blast administration. There was a significant increase
in the miR-29b expression level of NK cells from the spleen of
Notch1-T-ALL mice, compared with that of the control mice
(9.47±3.83 vs. 1.50±1.03; P=0.0005; Fig.
1A).
Human mature NK cells were characterized as
CD56brightCD16dim or CD56dim
CD16bright. The CD56dimCD16bright
NK subset is more common in the peripheral blood, and exhibits
higher cytotoxicity, but a lower level of cytokine production
compared with the CD56highCD16dim NK subset
(12). It was determined that a
similar increased level of miR-29b expression level occurred in NK
cells from the peripheral blood of patients with T-ALL; the miR-29b
expression in the CD56bright CD16dim NK cell
subset of untreated patients with newly diagnosed T-ALL was
compared with that of the healthy age-matched control group. The
results revealed a significantly increased level of miR-29b
expression in the CD56bright CD16dim NK cell
subset from T-ALL patients, compared with that of the healthy
control group (2.13±0.87 vs. 0.91±0.48; P=0.0039; Fig. 1B). These results suggested that NK
cells express increased levels of miR-29b in both murine
Notch1-T-ALL mice and T-ALL patients.
Previous studies have indicated the occurrence of
miRNA exchange between cells in chronic leukemia (24,25). In
addition, increased miR-29b expression levels in NK cells are
associated with decreased expression in AML blasts (17). The present study investigated whether
a similar reduction in miR-29b expression level occurred in T-ALL
blasts. 21 days after the administration of Notch1-T-ALL blasts
into mice, the miR-29b expression level in splenic blasts had
decreased, compared with that in the splenic WT CD3+ T
cells of mice without blast transfer (0.25±0.17 vs. 1.04±0.11;
P=0.0026; Fig. 1C). Furthermore, it
was demonstrated that the miR-29b expression level decreased in
Notch1-T-ALL blasts from the peripheral blood of patients with
T-ALL, compared with that in T cells from the peripheral blood of
healthy individuals (0.48±0.18 vs. 0.94±0.18; P=0.0315; Fig. 1D). These data indicated that miR-29b
expression was dysregulated in both T-ALL mice and patients.
Opposing trends in miR-29b expression levels were observed between
NK cells and leukemic blasts in the T-ALL microenvironment.
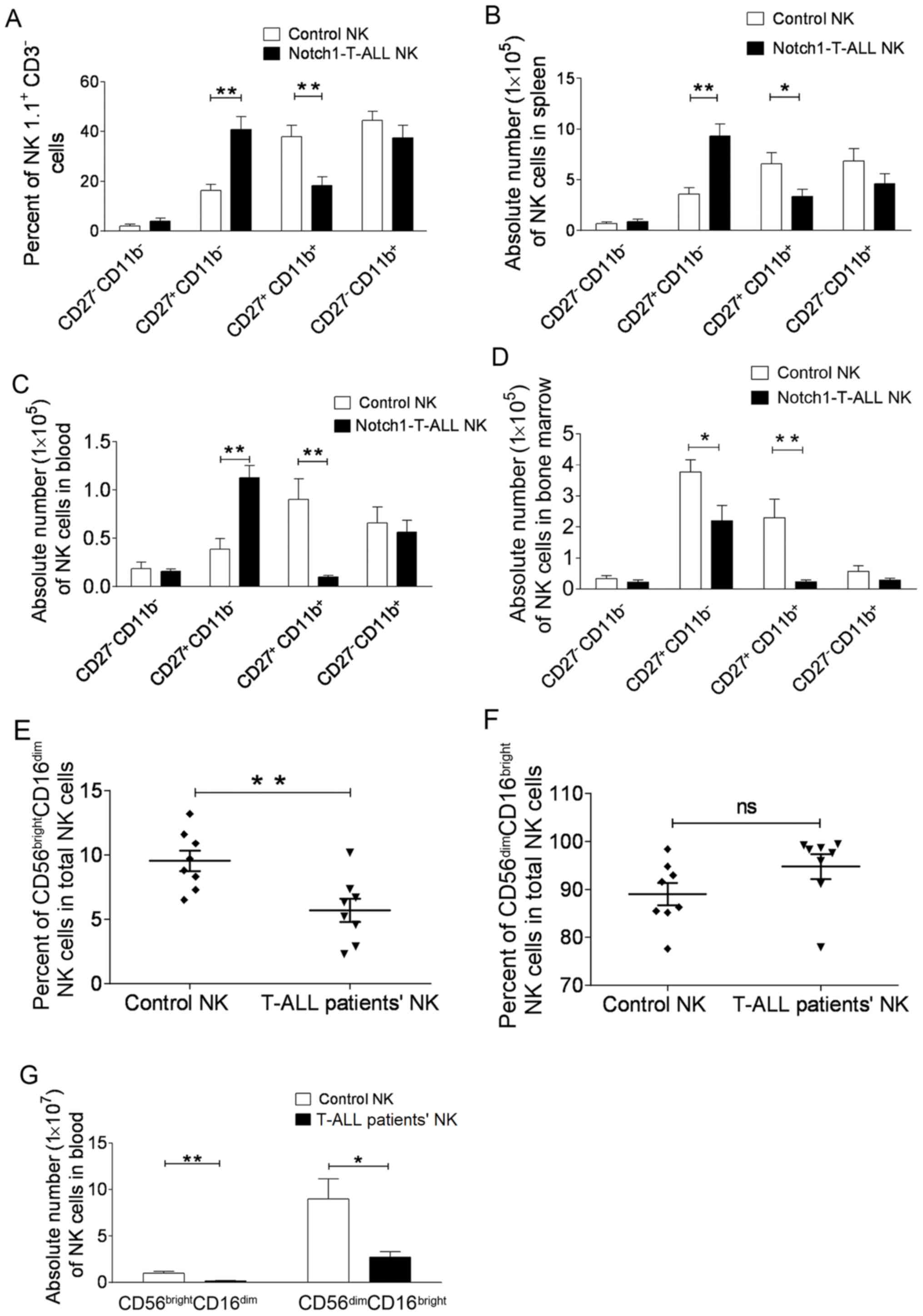
NK cell subset alterations in
Notch1-T-ALL mice and patients with T-ALL
A previous study demonstrated that the absolute
number of CD27+CD11b+ NK cells increased in
the peripheral blood, spleen and bone marrow of AML mice, which was
associated with an increase in NK cell miR-29b expression level
(17). Therefore, the present study
investigated NK cell subsets in Notch1-T-ALL mice. A significant
reduction in the CD27+CD11b+ subset in the
spleens of Notch1-T-ALL mice was observed compared with that in
control WT C57BL/6 mice (37.94±10.07% vs. 18.34±7.65%; P=0.0085;
Fig. 2A). There was no significant
change in the numbers of the most mature
(CD27−CD11b+) or the most immature
(CD27−CD11b−) NK cells (44.46±8.09% vs.
38.52±11.23%, P=0.2915; 2.06±1.55% vs. 3.96±2.90%, P=0.0593,
respectively; Fig. 2A). However, the
CD27+CD11b−subset increased significantly in
Notch1-T-ALL mice (16.36±5.46% vs. 40.85±11.32%; P=0.0024; Fig. 2A).
Furthermore, the absolute number of the NK cell
subsets in the spleen, peripheral blood and bone marrow of
Notch1-T-ALL mice was evaluated and compared with control mice. The
data revealed that the CD27+CD11b+ subset was
significantly reduced in the spleen (6.58±2.38×105/l vs.
3.36±1.59×105/l; P=0.0036; Fig. 2B), peripheral blood
(0.90±0.47×105/l vs. 0.11±0.05×105/l;
P=0.0061; Fig. 2C) and bone marrow
(2.29±1.34×105/l vs. 0.23±0.14×105/l;
P=0.0089; Fig. 2D). Also,
CD27+ CD11b−NK cells were increased in the
spleen (3.59±1.36×105/l vs. 9.30±2.68×105/l;
P=0.0029; Fig. 2B) and peripheral
blood (0.38±0.24×105/l vs. 1.12±0.28×105/l;
=0.0022; Fig. 2C) of Notch1-T-ALL
mice, compared with those of the control mice. However, this cell
subset was decreased in the bone marrow of Notch1-T-ALL mice,
compared with that in the control mice (3.77±0.88×105/l
vs. 2.19±0.99×105/l; P=0.0372; Fig. 2D). This may be due to the presence of
T-ALL blasts in the bone marrow of Notch1-T-ALL mice, which may
result in reduced hemocyte production compared with that of the
control group. These data illustrated the developmental retardation
of the CD27+CD11b−NK cell subset in
Notch1-T-ALL mice.
In the peripheral blood, 90% of NK cells are
characterized as CD56dim CD16bright, and only
10% as CD56brightCD16dim (11). NK cells were identified in the
peripheral blood of eight T-ALL patients at diagnosis, and in
age-matched healthy volunteers. The percentage of
CD56brightCD16dim NK cells in the total NK
cell population was decreased in patients with T-ALL, compared with
that in the control group (5.69±2.53% vs. 9.54±2.26%; P=0.0065;
Fig. 2E). In addition, a
corresponding elevation in the percentage of
CD56dimCD16bright NK cells from T-ALL
patients was detected, compared with that of the healthy controls
(89.04±6.62% vs. 94.78±7.32%; P=0.1228; Fig. 2F). As predicted, the absolute number
of CD56brightCD16dim NK cells was reduced in
patients with T-ALL, compared with the healthy control group
(0.15±0.07×107/l vs. 0.99±0.45×107/l;
P=0.0034; Fig. 2G). Again, this may
be as a result of T-ALL blasts in the peripheral blood of T-ALL
patients. Furthermore, it was observed that the absolute number of
CD56 dim CD16bright NK cells was decreased in
T-ALL patients when compared with that in the healthy control group
(8.98±4.78×107/l vs. 2.69±1.34×107/l;
P=0.0245; Fig. 2G). These data
suggested a developmental blockage in
CD56dimCD16bright NK cell subsets in patients
with T-ALL.
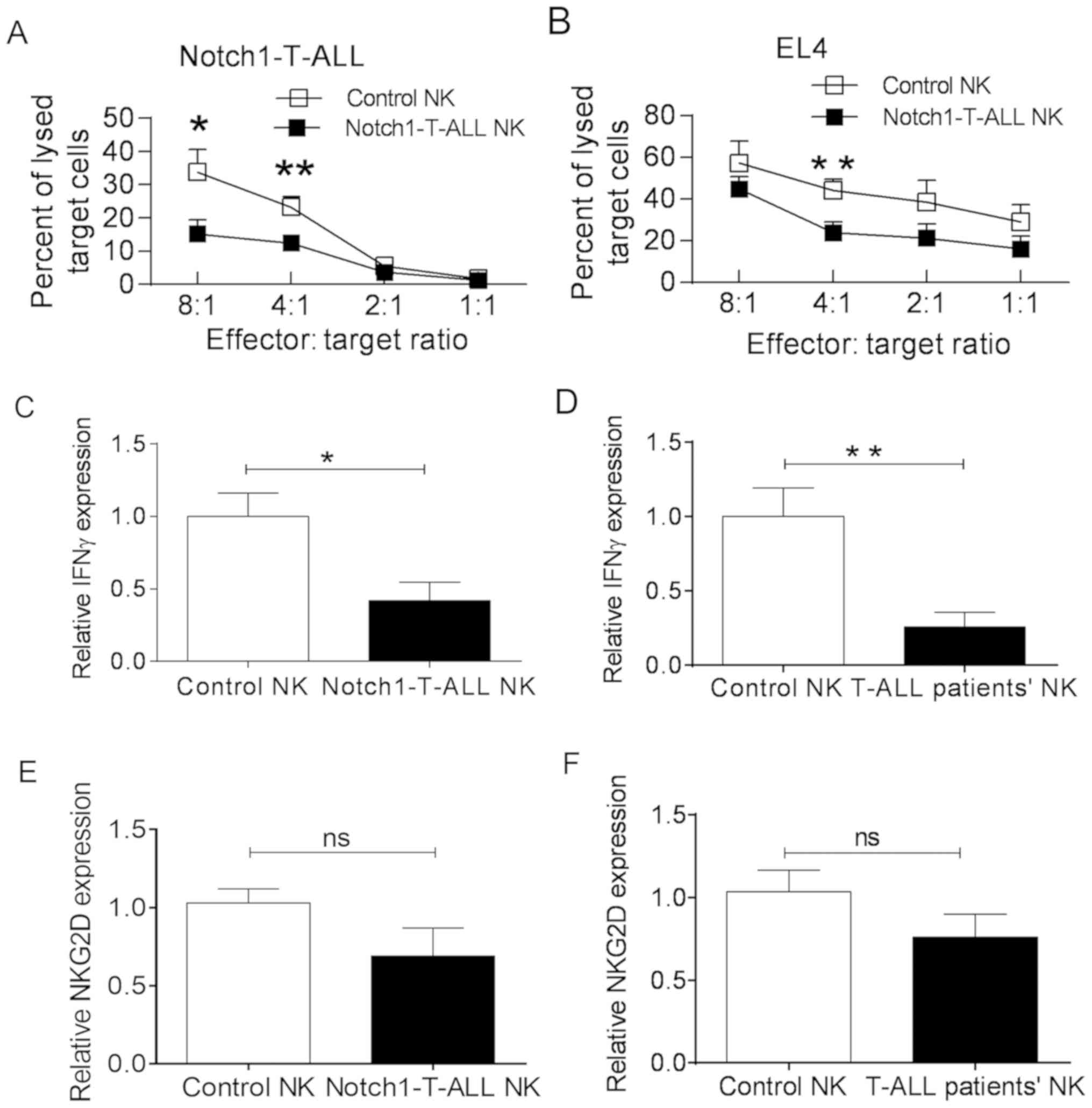
Alterations to NK cell function in
T-ALL
NK cells serve a critical role in the destruction
leukemia blasts. The cytotoxicity of splenic NK cells (from
Notch1-T-ALL mice or control WT C57BL/6 mice) towards Notch1-T-ALL
blasts or the murine T cell lymphoma EL4 cell line was assessed by
effector-target ratio of the number of cells at 8:1, 4:1, 2:1, 1:1,
respectively. The cytotoxic capacity of NK cells from Notch1-T-ALL
mice was markedly decreased, compared with that of NK cells from WT
C57BL/6 mice at an 8:1 (15.17±4.12% vs. 33.73±6.94%; P=0.0163) and
4:1 (12.43±2.42% vs. 23.20±3.19%; P=0.0095) ratio, respectively
(Fig. 3A). This trend was also
observed for EL4 cells, it was statistically significantly at a
ratio of 4:1 (23.80±5.21% vs. 44.17±5.33%; P=0.0091; Fig. 3B). To demonstrate whether a reduction
in IFNγ secretion from NK cells was one of the influencing factors
for NK cell cytotoxicity, the relative expression levels of IFNγ in
NK cells from both Notch1-T-ALL mice and WT C57BL/6 mice was
evaluated. The results revealed that the relative level of IFNγ
expression was significantly decreased in NK cells from
Notch1-T-ALL mice compared with that of the WT C57BL/6 mice
(0.41±0.22 vs. 1.03±0.27; P=0.0468; Fig.
3C). Additionally, the expression levels of IFNγ in NK cells
from patients with T-ALL were significantly decreased when compared
with those of the healthy controls (0.25±0.22 vs. 1.02±0.42;
P=0.0086; Fig. 3D).
The cytotoxic functions of NK cells are regulated by
the expression of activating and inhibitory receptors on the cell
surface. Interactions between the activating receptor NKG2D and its
ligands promote IFNγ production in NK cells (26). Therefore, alterations to NKG2D
expression in NK cells were investigated, which may be associated
with the reduction in IFNγ production and NK cell cytotoxicity. As
predicted, a reduction in the NKG2D expression level was detected
in the NK cells of Notch1-T-ALL mice, compared with that of the WT
C57BL/6 mice (0.7569±0.31 vs. 1.03±0.20; P=0.1036; Fig. 3E); a similar reduction was observed
in T-ALL patients (0.75±0.31 vs. 1.03±0.29; P=0.1882; Fig. 3F). These results suggested that NKG2D
expression levels decrease in the leukemia microenvironment of
T-ALL, which may be one of factors to influence the reduction of
IFNγ production.
ALL progression is inhibited in
miR-29b KO NK cells
It has previously been demonstrated that miR-29b
deletion in NK cells reverses dysregulation in the NK cell subsets
and modulates AML progression (17).
Subsequently, the impact of miR-29b KO NK cells on the progression
of Notch1-T-ALL was assessed. A total of 2×105
Notch1-T-ALL blasts were infused into three groups of C57BL/6
Rag2−/− recipient mice. In addition, 2×107 WT
NK cells or miR-29b KO NK cells were adoptively infused into two
groups of recipient mice. After 3 weeks, the percentage of
circulating Notch1-T-ALL blasts was evaluated. The number of
circulating Notch1-T-ALL blasts in mice with WT NK cells was
reduced, compared with mice without NK cells (44.69±6.81% vs.
52.03±7.85%; P=0.0885; Fig. 4A).
Notably, the number of Notch1-T-ALL blasts decreased significantly
in mice with miR-29b KO NK cells, compared with those without
(36.09±7.60% vs. 52.03±7.85%; P=0.0023; Fig. 4A). Also, improved survival time was
observed in mice with WT NK cells compared with those without NK
cells. Specifically, a significant improvement was observed in mice
receiving miR-29b KO NK cells (P=0.0230), compared with mice
without NK cell infusion. These results suggested that miR-29b KO
NK cells served a crucial role in controlling the progression of
T-ALL.
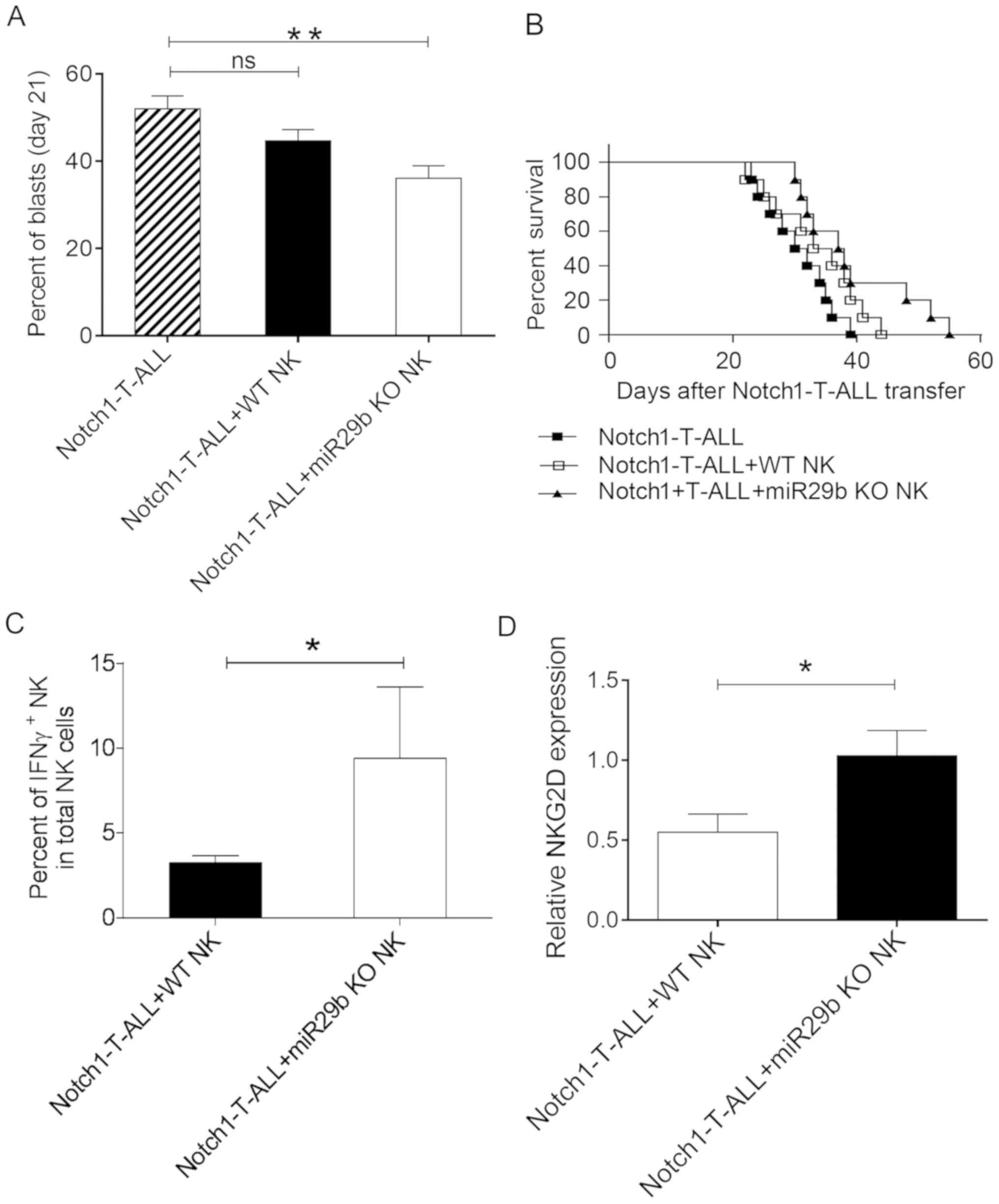 | Figure 4.miR29b KO NK cells decrease ALL
progression. Three groups of C57BL/6 Rag2−/− mice
received either 2×105 Notch1-T-ALL blasts without NK
cells; 2×105 Notch1-T-ALL blasts and 2×107 WT
NK cells from WT C57BL/6 mice; or 2×105 Notch1-T-ALL
blasts and 2×107 miR-29b KO cells from C57BL/6
miR29ab1−/−mice. (A) Percentages of circulating
Notch1-T-ALL blasts were evaluated using flow cytometry. (B)
Survival rate was compared between three groups. Notch1-T-ALL, ■;
Notch1-T-ALL+WT NK cells, □; and Notch1-T-ALL+miR-29b KO NK cells,
▲. (C) Percentage of IFNγ+ NK cells in the
NK1.1+CD3−NK cell population from
Notch1-T-ALL mice transfused with WT NK cells or miR-29b KO NK
cells. (D) Relative NKG2D expression levels in NK cells from
Notch1-T-ALL mice transfused with WT NK cells or miR-29b KO NK
cells. A: n=10 mice per group; B: n=10 per group (Kaplan-Meier
curve and log-rank test); C and D: n=10 mice per group. The data
are presented as the mean ± SD. *P<0.05 and **P<0.01, as
determined using one-way ANOVA or the unpaired Student's t-test.
miR, microRNA; KO, knock-out; T-ALL, T-cell acute lymphoblastic
leukemia; NK, natural killer; WT, wild type; Notch1, neurogenic
locus notch homolog protein 1; IFN, interferon; NKG2D, natural
killer receptor group 2, member D; ns, not significant. |
Following the deletion of miR-29b in Notch1-T-ALL
mice, NK cell function was restored and survival time subsequently
improved. The percentage of IFNγ+ NK cells and relative
NKG2D expression levels in the NK1.1+CD3−NK
cell population was determined. The results demonstrated that both
of these factors were significantly increased in Notch1-T-ALL mice
infused with miR-29b KO NK cells, compared with those infused with
WT NK cells [(9.39±4.22 vs. 3.23±1.38; P=0.0143; Fig. 4C) and (1.02±0.44 vs. 0.55±0.32;
P=0.0282; Fig. 4D) respectively].
Therefore, the mechanism of miR-29b KO NK cells in regulating T-ALL
progression may involve IFNγ production and/or NKG2D expression in
the NK cells of Notch1-T-ALL mice.
Discussion
NK cell-based immunotherapeutic strategies have
shown great promise in patients with AML and consist in infusions
of alloreactive NK cells with KIR ligand mismatch that protect
against AML relapse (4). Similar
types of treatments have been less successful for patients with
ALL, which may be due to the increased resistance of T-ALL blasts
to NK cell cytotoxicity, as they less frequently express ligands
for activating NK cell receptors (6,27).
Additionally, the available research on T-ALL blast resistance to
NK cells is fragmentary. The present study revealed an increase in
the miR-29b expression levels of NK cells, with a reduction in
miR-29b expression in T-ALL blasts. Furthermore, the NK phenotype
was determined, which revealed a selective downregulation of
CD27+CD11b+ NK cell subset. In addition, NK
cell alterations included a reduction in IFNγ production, decreased
NKG2D expression levels and impaired cytotoxicity in T-ALL, which
was associated with increased miR-29b expression levels in NK
cells.
miR-29 family members have been shown to be
downregulated in AML, as well as in chronic lymphocytic leukemia
(23). Furthermore, a reduced
expression level of miR-29b in AML blasts has been shown to
correlate with disease progression (23,24). As
a result, the overexpression of miR-29b enhanced the apoptosis of
AML blasts. Previous studies have identified that miR-29b
interferes with the development of CD4+ T cells by
regulating T-box-related TBX21 (T-bet) and eomesodermin (Eomes)
expression (28,29). Furthermore, in NK cells from T-bet or
Eomes KO mice, the resulting inhibition of terminal differentiation
was associated with a lack of CD11b+ NK cells (30). In the AML microenvironment, increased
miR-29b expression in NK cells altered NK differentiation by
regulating T-bet and Eomes (17).
However, to the best of our knowledge, miR-29b expression in NK
cells was yet to be evaluated in the T-ALL microenvironment.
In the present study, miR-29b expression in NK cells
was initially determined in both Notch1-T-ALL mice and T-ALL
patients. The results revealed that in Notch1-T-ALL mice (Fig. 1A) and T-ALL patients (Fig. 1B), miR-29b expression of NK cells was
upregulated, but miR-29b expression of T-ALL blasts (Fig. 1C and D) was downregulated. Previous
studies have reported that elevated levels of miR-29b in NK cells
may be associated with the exchange of miR-29b released from
chronic leukemia blasts (25,31).
Exosomes and vesicles are involved in the transfer of miR-29b
between cells, as well as leukemia progression and immune
suppression (25,32). Further studies that focus on the
mechanisms of miR-29b expression regulation are required.
A blockage in NK cell differentiation was also
observed in both Notch1-T-ALL mice and patients with T-ALL. In the
mouse model, the percentage and absolute number of
CD27+CD11b+ NK cells decreased in mice with
leukemia, compared with the control group. Comparatively, a
decreased percentage and absolute number of
CD56brightCD16dim NK cells was also observed
in patients with T-ALL, compared with the healthy control group.
This result was consistent with a previous study which reported
that NK cell development is arrested in AML (17); this study demonstrated that the
increased level of miR-29b expression in NK cells downregulated
T-bet and Eomes in AML mice, which resulted in the accumulation of
CD27+CD11b−NK cells and a reduction in
CD27+CD11b+ NK cells.
Furthermore, a decrease in the percentage and
absolute number of the CD56brightCD16dim NK
cell subset was demonstrated in untreated T-ALL patients, compared
with the control group. This suggested that T-ALL blasts
selectively eliminate this NK subset and block NK cell maturation,
which results in NK cell immune surveillance failure. A previous
report indicated that the absolute number of CD56bright
NK cells increased at the complete remission stage of leukemia
(33). Furthermore, Mundy-Bosse
et al (17) demonstrated that
the depletion of miR-29b restored the intermediate
CD27+CD11b+ NK cell population in AML. In the
present study, although the impact of adoptively transferred
miR-29b KO NK cells on the restoration of NK cells subsets was not
investigated, the impact of these cells on NK cell function in the
Notch1-T-ALL microenvironment was determined.
In the present study, NK cell cytotoxicity towards
Notch1-T-ALL cells and EL4 murine lymphoma cells was significantly
decreased in Notch1-T-ALL mice, compared with WT C57BL/6 mice. NK
cells secrete cytokines that kill target cells and influence the
host immune response (13,34). As a prototypical NK cell cytokine,
IFNγ activates antigen-presenting cells to induce MHC-I expression
(35), and inhibits the
proliferation of malignant cells (36). As the primary producer of IFNγ, two
signals are involved in IFNγ production by CD56bright NK
cells; NKG2D, a critical NK-activating receptor expressed on all
cells from the NK cell lineage (37), is involved in IFNγ production and NK
cell activation (11,38), and is also fundamentally involved in
the NK cell-mediated antitumor and antiviral immune responses
(15,39). The upregulation of NKG2D promotes
IFNγ secretion via co-stimulatory signaling of NK cells (40). When NKG2D is blocked, fewer NK cells
secrete IFNγ (41). In the present
study, relative NKG2D expression on NK cells was significantly
decreased in both Notch1-T-ALL mice and patients with T-ALL,
implying that the cytotoxicity of NK cells was decreased in the
lymphoblastic leukemia microenvironment. Accordingly, IFNγ
secretion by NK cells significantly decreased in both Notch1-T-ALL
mice and patients with T-ALL. The reduction in IFNγ secretion was
in line with a previous study, which reported that the reduction in
intracellular IFNγ production by NK cells was associated with an
elevation in miR-29b expression in the AML microenvironment
(17). It was further hypothesized
that the increase in miR-29b expression level in the present study
may have been associated with a reduction in IFNγ production. When
Notch1-T-ALL mice received miR-29b KO NK cells, the partial
restoration of NK cell IFNγ production and NKG2D expression were
observed. Furthermore, adoptive transfusion of miR-29b KO NK cells
into Notch1-T-ALL mice promoted ALL progression and improved
survival time. These results identified the elevation of miR-29b
expression level as one of the contributing factors to decreased NK
cell function in the T-ALL microenvironment. Further research is
required to determine how miR-29b affects NK cell IFNγ production
and NKG2D expression in T-ALL.
To conclude, the present study identified previously
unknown NK cell defects that are associated with miR-29b
dysregulation in the T-ALL microenvironment. Alterations in NK cell
maturation and function resulted in decreased NK cytotoxicity in
T-ALL, suggesting that miR-29b is involved in NK cell development
arrest and functional defects, which is used by leukemia cells to
evade immune surveillance in T-ALL. Further research is required to
reveal the mechanism driving this phenomenon, and solutions that
restore NK cell maturation and function to decrease T-ALL
relapse.
Acknowledgements
The authors would like to thank Dr Mengmeng Liu and
Dr Fuming Zhang for their technical support.
Funding
The present study was supported by the Norman
Bethune Program of Jilin University (grant no. 2012224) and the
National Natural Science Foundation of China (grant nos. 81770149
and 81100350).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Author's contributions
YPY designed the study, provided funding and was a
major contributor in writing the manuscript. FYJ, ZHD, YT and LXW
performed the examinations. FYJ analyzed and interpreted the
patient data, and ZHD curated the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Each patient provided written informed consent and
the study was approved by the Ethical Review Board of the First
Bethune Hospital of Jilin University (protocol no. 2017-004). The
research was performed according to the Guidance on the Operation
of the Animals (Scientific Procedures) Act 1986, as well as the
Care and Use of Laboratory Animals of the National Institutes of
Health.
Patient consent for publication
Patients consented to publication of the
article.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Onciu M: Acute lymphoblastic leukemia.
Hematol Oncol Clin North Am. 23:655–674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rouce RH, Shaim H, Sekine T, Weber G,
Ballard B, Ku S, Barese C, Murali V, Wu MF, Liu H, et al: The
TGF-β/SMAD pathway is an important mechanism for NK cell immune
evasion in childhood B-acute lymphoblastic leukemia. Leukemia.
30:800–811. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oelsner S, Wagner J, Friede ME, Pfirrmann
V, Genßler S, Rettinger E, Buchholz CJ, Pfeifer H, Schubert R,
Ottmann OG, et al: Chimeric antigen receptor-engineered
cytokine-induced killer cells overcome treatment resistance of
pre-B-cell acute lymphoblastic leukemia and enhance survival. Int J
Cancer. 139:1799–1809. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miller JS, Soignier Y,
Panoskaltsis-Mortari A, McNearney SA, Yun GH, Fautsch SK, McKenna
D, Le C, Defor TE, Burns LJ, et al: Successful adoptive transfer
and in vivo expansion of human haploidentical NK cells in
patients with cancer. Blood. 105:3051–3057. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pende D, Spaggiari GM, Marcenaro S,
Martini S, Rivera P, Capobianco A, Falco M, Lanino E, Pierri I,
Zambello R, et al: Analysis of the receptor-ligand interactions in
the natural killer-mediated lysis of freshly isolated myeloid or
lymphoblastic leukemias: Evidence for the involvement of the
Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood.
105:2066–2073. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Romanski A, Bug G, Becker S, Kampfmann M,
Seifried E, Hoelzer D, Ottmann OG and Tonn T: Mechanisms of
resistance to natural killer cell-mediated cytotoxicity in acute
lymphoblastic leukemia. Exp Hematol. 33:344–352. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sanchez-Correa B, Morgado S, Gayoso I,
Bergua JM, Casado JG, Arcos MJ, Bengochea ML, Duran E, Solana R and
Tarazona R: Human NK cells in acute myeloid leukaemia patients:
Analysis of NK cell-activating cellactivating receptors and their
ligands. Cancer Immunol Immunother. 60:1195–1205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lion E, Willemen Y, Berneman ZN, Van
Tendeloo VF and Smits EL: Natural killer cell immune escape in
acute myeloid leukemia. Leukemia. 26:2019–2026. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chiossone L, Chaix J, Fuseri N, Roth C,
Vivier E and Walzer T: Maturation of mouse NK cells is a 4-stage
developmental program. Blood. 113:5488–5496. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hayakawa Y and Smyth MJ: CD27 dissects
mature NK cells into two subsets with distinct responsiveness and
migratory capacity. J Immunol. 176:1517–1524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cooper MA, Fehniger TA, Turner SC, Chen
KS, Ghaheri BA, Ghayur T, Carson WE and Caligiuri MA: Human natural
killer cells: A unique innate immunoregulatory role for the CD56
(bright) subset. Blood. 97:3146–3151. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu J, Freud AG and Caligiuri MA: Location
and cellular stages of natural killer cell development. Trends
Immunol. 34:573–582. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Caligiuri MA: Human natural killer cells.
Blood. 112:461–469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chan A, Hong DL, Atzberger A, Kollnberger
S, Filer AD, Buckley CD, McMichael A, Enver T and Bowness P:
CD56bright human NK cells differentiate into CD56dim cells: Role of
contact with peripheral fibroblasts. J Immunol. 179:89–94. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jin F, Lin H, Gao S, Hu Z, Zuo S, Sun L,
Jin C, Li W and Yang Y: The anti-tumor role of NK cells in vivo
pre-activated and re-stimulated by interleukins in acute
lymphoblastic leukemia. Oncotarget. 7:79187–79202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu J, Song Y, Bakker AB, Bauer S, Spies T,
Lanier LL and Phillips JH: An activating immunoreceptor complex
formed by NKG2D and DAP10. Science. 285:730–732. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mundy-Bosse BL, Scoville SD, Chen L,
McConnell K, Mao HC, Ahmed EH, Zorko N, Harvey S, Cole J, Zhang X,
et al: MicroRNA-29b mediates altered innate immune development in
acute leukemia. J Clin Invest. 126:4404–4416. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leong JW, Sullivan RP and Fehniger TA:
MicroRNA management of NK-cell developmental and functional
programs. Eur J Immunol. 44:2862–2868. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weng AP, Ferrando AA, Lee W, Morris JP IV,
Silverman LB, Sanchez-Irizarry C, Blacklow SC, Look AT and Aster
JC: Activating mutations of NOTCH1 in human T cell acute
lymphoblastic leukemia. Science. 306:269–271. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kloss M, Decker P, Baltz KM, Baessler T,
Jung G, Rammensee HG, Steinle A, Krusch M and Salih HR: Interaction
of monocytes with NK cells upon Toll-like receptor-induced
expression of the NKG2D ligand MICA. J Immunol. 181:6711–6719.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y, Wang H, Yu H, Yeap BY, Liang T,
Wang G, Cheng T and Yang YG: IFN-γ promotes graft-versus-leukemia
effects without directly interacting with leukemia cells in mice
after allogeneic hematopoietic cell transplantation. Blood.
118:3721–3724. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lieberman NAP, DeGolier K, Haberthur K,
Chinn H, Moyes KW, Bouchlaka MN, Walker KL, Capitini CM and Crane
CA: An uncoupling of canonical phenotypic markers and functional
potency of ex vivo-expanded natural killer cells. Front Immunol.
9:1502018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garzon R, Heaphy CE, Havelange V, Fabbri
M, Volinia S, Tsao T, Zanesi N, Kornblau SM, Marcucci G, Calin GA,
et al: MicroRNA 29b functions in acute myeloid leukemia. Blood.
114:5331–5341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu S, Wu LC, Pang J, Santhanam R, Schwind
S, Wu YZ, Hickey CJ, Yu J, Becker H, Maharry K, et al:
Sp1/NFkappaB/HDAC/miR-29b regulatory network in KIT-driven myeloid
leukemia. Cancer Cell. 17:333–347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huan J, Hornick NI, Shurtleff MJ, Skinner
AM, Goloviznina NA, Roberts CT Jr and Kurre P: RNA trafficking by
acute myelogenous leukemia exosomes. Cancer Res. 73:918–929. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boukouaci W, Al-Daccak R, Dulphy N, Lauden
L, Amokrane K, Fortier C, Marzais F, Bennabi M, Peffault de Latour
R, Socie G, et al: Soluble MICA-NKG2D interaction upregulates IFN-γ
production by activated CD3-CD56+ NK cells: Potential impact on
chronic graft versus host disease. Hum Immunol. 74:1536–1541. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Boieri M, Ulvmoen A, Sudworth A, Lendrem
C, Collin M, Dickinson AM, Kveberg L and Inngjerdingen M: IL-12,
IL-15, and IL-18 pre-activated NK cells target resistant T cell
acute lymphoblastic leukemia and delay leukemia development in
vivo. Oncoimmunology. 6:e12744782017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Steiner DF, Thomas MF, Hu JK, Yang Z,
Babiarz JE, Allen CD, Matloubian M, Blelloch R and Ansel KM:
MicroRNA-29 regulates T-box transcription factors and interferon-γ
production in helper T cells. Immunity. 35:169–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Smith KM, Guerau-de-Arellano M, Costinean
S, Williams JL, Bottoni A, Mavrikis Cox G, Satoskar AR, Croce CM,
Racke MK, Lovett-Racke AE and Whitacre CC: miR-29ab1 deficiency
identifies a negative feedback loop controlling Th1 bias that is
dysregulated in multiple sclerosis. J Immunol. 189:1567–1576. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gordon SM, Chaix J, Rupp LJ, Wu J, Madera
S, Sun JC, Lindsten T and Reiner SL: The transcription factors
T-bet and Eomes control key checkpoints of natural killer cell
maturation. Immunity. 36:55–67. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yeh YY, Ozer HG, Lehman AM, Maddocks K, Yu
L, Johnson AJ and Byrd JC: Characterization of CLL exosomes reveals
a distinct microRNA signature and enhanced secretion by activation
of BCR signaling. Blood. 125:3297–3305. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Whiteside TL: Immune modulation of T-cell
and NK (natural killer) cell activities by TEXs (tumour-derived
exosomes). Biochem Soc Trans. 41:245–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dauguet N, Récher C, Demur C, Fournié JJ,
Poupot M and Poupot R: Pre-eminence and persistence of immature
natural killer cells in acute myeloid leukemia patients in first
complete remission. Am J Hematol. 86:209–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Smyth MJ, Cretney E, Kelly JM, Westwood
JA, Street SE, Yagita H, Takeda K, van Dommelen SL, Degli-Esposti
MA and Hayakawa Y: Activation of NK cell cytotoxicity. Mol Immunol.
42:501–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wallach D, Fellous M and Revel M:
Preferential effect of gamma interferon on the synthesis of HLA
antigens and their mRNAs in human cells. Nature. 299:833–836. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Maher SG, Romero-Weaver AL, Scarzello AJ
and Gamero AM: Interferon: Cellular executioner or white knight?
Curr Med Chem. 14:1279–1289. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jelenčić V, Lenartić M, Wensveen FM and
Polić B: NKG2D: A versatile player in the immune system. Immunol
Lett. 189:48–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Moretta A, Bottino C, Vitale M, Pende D,
Cantoni C, Mingari MC, Biassoni R and Moretta L: Activating
receptors and coreceptors involved in human natural killer
cell-mediated cytolysis. Annu Rev Immunol. 19:197–223. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mistry AR and O'Callaghan CA: Regulation
of ligands for the activating receptor NKG2D. Immunology.
121:439–447. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Girart MV, Fuertes MB, Domaica CI, Rossi
LE and Zwirner NW: Engagement of TLR3, TLR7, and NKG2D regulate
IFN-gamma secretion but not NKG2D-mediated cytotoxicity by human NK
cells stimulated with suboptimal doses of IL-12. J Immunol.
179:3472–3479. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim JY, Huh K, Lee KY, Yang JM and Kim TJ:
Nickel induces secretion of IFN-gamma by splenic natural killer
cells. Exp Mol Med. 41:288–295. 2009. View Article : Google Scholar : PubMed/NCBI
|