Introduction
Ferroptosis is a novel form of regulated cell death
(RCD) that was described in 2012 (1). Ferroptosis involves the accumulation of
lipid peroxidation products and reactive oxygen species (ROS)
derived from iron metabolism. Ferroptosis is morphologically,
biochemically and genetically distinct from other pathways for RCD,
including necroptosis, apoptosis and autophagy (2). Amino acid, iron and lipid metabolism
are involved in the process of ferroptosis. Glutamate and glutamine
are pivotal regulators of ferroptosis (3). During normal physiological function,
extracellular glutamate induces ferroptosis. Lipid metabolism may
affect the sensitivity of cells to ferroptosis. Polyunsaturated
fatty acids are susceptible to lipid peroxidation and are required
for ferroptosis (4). Iron is
involved in the accumulation of lipid peroxides and ferroptosis.
Iron import, export, storage and turnover influence the sensitivity
of cells to ferroptosis (5). Several
molecules, including voltage dependent anion channel (VDAC) 2/3,
glutathione peroxidase 4 (GPX4), heat shock protein β-1, nuclear
factor E2-related factor 2 (NRF2), NADPH oxidase, the tumor
suppressor p53 (TP53) and solute carrier family 7 member 1
(SLC7A1), regulate ferroptosis through the direct or indirect
targets of iron metabolism as well as lipid peroxidation (2).
Previous studies demonstrated that ferroptosis
participated in diverse diseases, including neuropathy (1,6),
ischemia reperfusion injury (3)
acute kidney failure (6–8) and cancer (3). Sorafenib was revealed to induce
ferroptosis in hepatocellular carcinoma (HCC) and several studies
have subsequently investigated the mechanism of ferroptosis in HCC
(9–11). The digestive system comprises several
organs and the incidence of cancer in the digestive system is high.
The current review presents the role of ferroptosis in digestive
system cancer.
Ferroptosis in digestive system cancer
Ferroptosis was revealed to be involved in various
types of digestive system cancer, excluding esophageal and biliary
system cancer (9,12–15). The
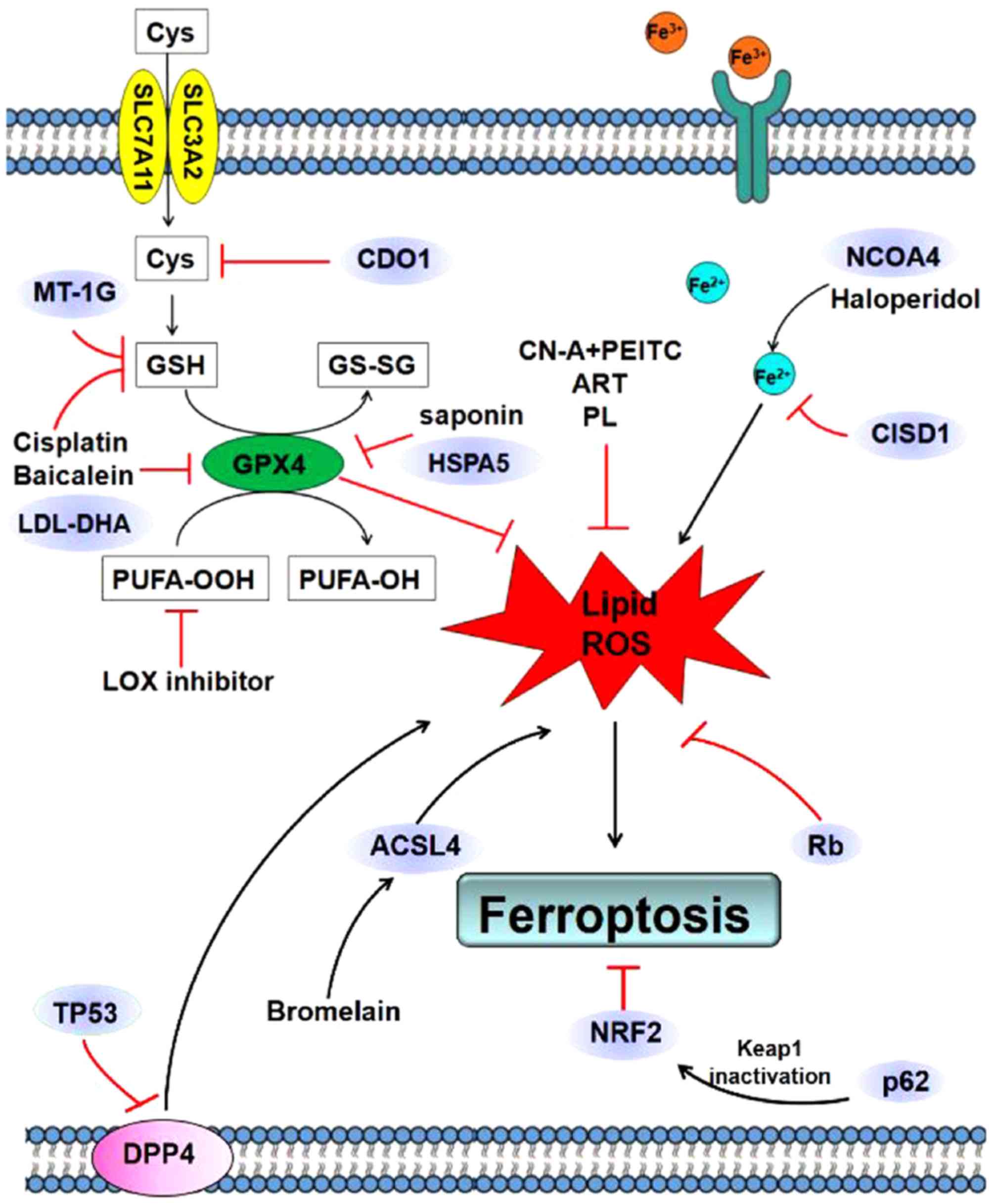
mechanisms of ferroptosis in digestive system cancer are presented
in Table I and Fig. 1.
 | Table I.Mechanisms of ferroptosis in
digestive system cancer. |
Table I.
Mechanisms of ferroptosis in
digestive system cancer.
| Author, year | Cancer type | Mechanism | (Refs.) |
|---|
| Hao et al,
2017 | Gastric cancer | Silencing cysteine
dioxygenase type 1 inhibited ferroptosis by restoring cellular GSH
and preventing ROS generation. | (12) |
| Wei et al,
2018 | Colon cancer | Suppressed the
expression of GPX4 and induced the accumulation of lipid
peroxidation products by activating TP53. | (21) |
| Xie et al,
2017 |
| TP53 limited
erastin-induced ferroptosis by blocking dipeptidyl peptidase 4
activity in a transcription-independent manner. | (20) |
| Guo et al,
2018 |
| Cisplatin induced
ferroptosis via GSH depletion and GPX4 inactivation. | (22) |
| Hong et al,
2017 |
| TP53-independent
PUMA axis is involved in ferroptosis in human colon cancer HCT116
cells. | (19) |
| Park et al,
2018 |
| Bromelain increased
erastin-induced ferroptosis by increasing acyl-CoA synthetase long
chain family member 4 in Kras mutant human colorectal carcinoma
cells. | (23) |
| Kasukabe et
al, 2016 | Pancreatic
cancer | Cotylenin A and
phenethyl isothiocyanate induced ferroptosis by generating
ROS. | (26) |
| Zhu et al,
2017 |
| HSPA5 inhibited
ferroptosis via the HSPA5-GPX4 signaling pathway. | (14) |
| Eling et al,
2015 |
| Artesunate induced
ferroptosis through ROS generation and lipid peroxidation. | (27) |
| Xie et al,
2016 |
| Baicalein inhibited
erastin-induced ferroptosis by preventing GSH depletion and GPX4
degradation. | (18) |
| Yamaguchi et
al, 2018 |
| Piperlongumine
induced ferroptosis through ROS generation. | (15) |
| Hou et al,
2016 |
| Knockdown of ATG5
and ATG7 limited erastin-induced ferroptosis and overexpression of
NCOA4 increased ferritin degradation and promoted ferroptosis. | (30) |
| Shintoku et
al, 2017 |
| Lipoyxgenase
inhibitors prevented pancreatic cell ferroptosis and arachidonate
15-lipoxygenase-activating compounds accelerated ferroptosis. | (29) |
| Hong et al,
2017 |
| TP53-independent
PUMA axis is involved in ferroptosis. | (19) |
| Zhang et al,
2018 | HCC | Increased
expression of ELAV like RNA binding protein 1 through inhibition of
the ubiquitin-proteasome pathway in ferroptosis. | (38) |
| Sun et al,
2016 |
| p62 expression
prevented NRF2 degradation and enhanced NRF2 nuclear accumulation
through inactivation of kelch like ECH associated protein 1 in HCC
cells treated with a ferroptosis inducer. | (10) |
| Yuan et al,
2016 |
| CDGSH iron sulfur
domain 1 inhibited ferroptosis by protecting against mitochondrial
lipid peroxidation. | (32) |
| Bai et al,
2017 |
| Haloperidol
increased the cellular levels of Fe2+ and lipid
peroxidation and promoted ferroptosis. | (9) |
| Louadre et
al, 2013 |
| Iron chelator
deferoxamine inhibited ferroptosis by preventing oxidative stress
in HCC cells. | (31) |
| Ou et al,
2017 |
| LDL-DHA
nanoparticles induced ferroptotic cell death by blocking GPX4. | (34) |
| Houessinon et
al, 2016 |
| MT1 constituted a
biomarker of altered redox metabolism and ferroptosis in HCC cells
exposed to sorafenib. | (44) |
| Sun et al,
2016 |
| Inhibition of MT-1G
enhanced the anticancer activity of sorafenib in vitro and
in tumor xenograft models. | (45) |
| Dehart et
al, 2018 |
| Erastin and
erastin-like compounds caused HCC ferroptosis by opening voltage
gated anion channels. | (41) |
| Sauzay et
al, 2018 |
| Sorafenib inhibited
the initiation of translation and the mechanistic target of
rapamycin kinase signaling cascade. | (11) |
| Louadre et
al, 2015 |
| Reduced levels of
etinoblastoma protein increased ferroptosis induction upon exposure
to sorafenib. | (42) |
Gastric cancer
Studies investigating ferroptosis in gastric cancer
are lacking. Human cysteine dioxygenase 1 (CDO1) converts cysteine
to taurine (16); therefore, an
increase in CDO1 activity is expected to decrease cellular cysteine
levels. Cysteine is required for glutathione (GSH) synthesis and
increased CDO1 activity may decrease the synthesis of GSH and
promote ferroptosis (16). Hao et
al (12) revealed that that
silencing of CDO1 in gastric cancer cells reduced the
erastin-induced ferroptosis by restoring cellular GSH, which
prevented ROS generation and lipid peroxidation. MYB proto-oncogene
transcription factor, which interacts with the CDO1 promoter as a
transcription factor, regulated CDO1 and GPX4 expression during
ferroptosis (12). Further
investigation is required to identify the role and underlying
mechanism of ferroptosis in gastric cancer.
Colon cancer
Previous studies investigating the role of
ferroptosis in colon cancer have largely focused on TP53 (13,17–19).
TP53 is an evolutionarily conserved protein involved in the
regulation of cell proliferation, death and differentiation and
metabolism. TP53 may initiate cell-cycle arrest and induce
apoptosis in response to stress stimuli by means of the
transcriptional and transcription-independent processes (17). Additionally, the induction of
ferroptosis may contribute to the oncosuppressive activities of
TP53 (13). Furthermore, TP53 was
revealed to promote ferroptosis by means of a
transcription-dependent process (20). Xie et al (20) revealed that TP53 constrained
erastin-stimulated ferroptosis through the inhibition of
dipeptidyl-peptidase-4 (DPP4) function in a
transcription-independent manner in colorectal cancer. Silencing of
TP53 facilitated plasma membrane-associated DPP4-dependent lipid
peroxidation, and thus prevented nuclear accumulation of DPP4. As a
result, SLC7A11 expression was increased which resulted in
ferroptosis (20). Furthermore, the
TP53-independent p53 upregulated modulator of apoptosis axis was
implicated in ferroptosis induction in human colon cancer HCT116
cells (19).
The role of TP53 in ferroptosis may provide a
therapeutic target for colon cancer. Albiziabioside A, a saponin
derivative, suppressed the expression of GPX4, and together with
the accumulation of lipid peroxidation products, resulted in
ferroptosis by TP53 activation (21). Cisplatin induced ferroptosis by
depleting GSH and inactivating GPX4 in A549 and HCT116 cells,
suggesting that ferroptosis may be a novel target in cancer therapy
(22). Bromelain exhibited
anticancer effects in Kras mutant human colorectal carcinoma cells
by upregulating the expression of acyl-CoA synthetase long chain
family member 4, which acted as a ferroptosis target (23).
Defective ribosome biogenesis was reported in colon
cancer lacking TP53 (24).
5-fluorouracil induced ribosomal protein L3 (RPL3) activation in
TP53-/- colon cancer cells by inhibiting the expression of
cystathionine-β-synthase (24),
which may subsequently influence the cysteine level. Additionally,
RPL3 decreased the expression of cysteine/glutamate antiporter and
induced ferroptosis in TP53-mutated lung cancer cells (25). Ribosomal protein-dependent pathways
are associated with ferroptosis and may be potential targets of
anticancer drugs.
Pancreatic cancer
Pancreatic cancer is one of the most lethal
types of digestive malignancies, and current chemotherapeutic drugs
are often ineffective. Previous studies suggested that certain
natural plant extracts possess a potential therapeutic effect in
pancreatic cancer cells by inducing ferroptosis (15,18,26,27). The
combination of cotylenin A (CN-A) and phenethyl isothiocyanate
synergistically induced pancreatic cancer cell death by the
generation of ROS, which drives ferroptosis (26). In addition, artesunate (ART) induced
ferroptosis by ROS generation and lipid peroxidation in
KRAS-activated human pancreatic ductal adenocarcinoma (PDAC) and
the AT-induced ferroptosis was inhibited by ferrostatin-1. Notably,
ART did not exert an effect on non-neoplastic human pancreatic
ductal epithelial cells (27). This
suggests that ART may be a candidate for pancreatic cancer therapy
by inducing ferroptosis. Piperlongumine (PL) induces human
pancreatic cancer cell line death by substantially increasing the
intracellular ROS level. This effect was inhibited by ferroptosis
inhibitors and iron chelators but not apoptosis or necrosis
inhibitors, suggesting that PL induced cell death by ferroptosis
(15). Furthermore, the
aforementioned study revealed that triple combined therapy with PL,
CN-A and sulfasalazine had a high efficacy in pancreatic cancer
in vitro (15). Baicalein
inhibited erastin-induced ferroptosis by preventing GSH depletion
and GPX4 degradation in pancreatic cancer cells (18).
Previous studies have investigated the mechanism of
ferroptosis in pancreatic cancer cells (14,28,29).
Heat shock protein family A (Hsp70) member 5 (HSPA5) negatively
regulated ferroptosis in human PDAC cells through the HSPA5-GPX4
signaling pathway, which mediated ferroptosis resistance (14). The inhibition of the HSPA5-GPX4
signaling pathway improved gemcitabine sensitivity through the
disinhibition of ferroptosis in vitro and in vivo
(14). Nuclear receptor coactivator
4 (NCOA4) was revealed as the cargo receptor mediating ferroptosis
(28). Silencing of NCOA4 inhibited
ferritin degradation and suppressed ferroptosis while the
overexpression of NCOA4 augmented ferritin degradation and promoted
ferroptosis in human pancreatic cells (30). Lipoxygenase inhibitors prevented
pancreatic cell ferroptosis induced by erastin as well as the
ferroptosis inhibitor RSL3. In addition, treatment of the human
pancreatic cancer cell line Panc-1 with arachidonate
15-lipoxygenase-activating compounds accelerated ferroptosis
(29). The TP53-independent PUMA
axis was implicated in ferroptosis in human pancreatic cancer cells
as well as the previously mentioned colon cancer cells (19).
HCC
The majority of previous studies associated with
ferroptosis involved the investigation of HCC (10,31,32).
Sorafenib, a multikinase inhibitor, is used for the treatment of
advanced HCC and is currently being investigated as a ferroptosis
inducer (2,9). Mechanistically, sorafenib induces
ferroptosis as a form of regulated cell death in HCC. Sorafenib
inhibits the initiation of translation mediated by the rapamycin
kinase signaling pathway which is considered to constitutes an
important role in the initiation of ferroptosis in HCC (11). Iron metabolism is identified as a
vital mediator of ferroptosis and the iron chelator deferoxamine
prevents sorafenib from inducing oxidative stress in HCC cells
(31). CDGSH iron sulfur domain 1
(CISD1) is an iron-possessing external mitochondrial membrane
protein that modulates mitochondrial iron uptake and respiration
(33). A previous study revealed
that CISD1 decreased ferroptotic cell death in HCC cells by
regulating iron metabolism and preventing mitochondrial damage in
ferroptosis (32). In addition to
iron metabolism regulating ferroptosis, lipid metabolism may serve
an important role in the regulation of ferroptosis (31). Low-density
lipoproteins-docosahexaenoic acid nanoparticles stimulated
ferroptosis by promoting lipid peroxidation, depleting glutathione
and inactivating the lipid antioxidant GPX4 in a rat hepatoma
model, HCC cell lines and human HCC tumor xenografts in mice
(34).
NRF2 is implicated in several types of cancer,
including HCC (35–37). RF2 is involved in ferroptosis in HCC
cells. When HCC cells were exposed to a ferroptosis inducer, p62
expression decreased NRF2 degradation and increased NRF2 nuclear
accumulation by inactivating Kelchlike ECH-associated protein 1
(KEAP1). Knockdown of p62 in HCC cells promoted the anticancer
function of erastin and sorafenib by inducing ferroptosis (10). Activating the p62-KEAP1-NRF2
signaling pathway may thus protect against ferroptosis in HCC cells
(10). Furthermore, NRF2 determined
the therapeutic efficacy of ferroptosis-targeted therapies in HCC
cells (10).
Zhang et al (38) revealed that ELAV-like RNA binding
protein 1 (ELAVL1) serves an important role in the regulation of
ferroptosis in liver fibrosis and hepatocarcinoma. When erastin was
exposed to ferroptosis-inducing compounds, ELAVL1 protein
expression was significantly increased due to inhibition of the
ubiquitin-proteasome pathway (38).
The σ 1 receptor (S1R) is present in the mitochondrial membrane and
endoplasmic reticulum and modulates oxidative stress (39,40).
Haloperidol, an S1R antagonist, significantly increased the
cellular concentration of Fe2+ as well as lipid
peroxidation, and accordingly promoted erastin-and
sorafenib-induced ferroptosis (9).
Furthermore, erastin, coupled with other erastin-like lead
compounds, induced HCC ferroptosis through the opening of VDACs,
which increased the membrane potential, mitochondrial ROS and
oxidative stress-induced cell death (41).
Louandre et al (42) revealed that HCC cells with decreased
expression of retinoblastoma protein (Rb) manifested a 2-to 3-fold
increase in ferroptotic cell death induced by sorafenib compared
with controls, highlighting the function of Rb in determining the
susceptibility of HCC cells to sorafenib as well as regulating
ferroptosis. However, clinically-applicable biomarkers reflecting
the susceptibility of cancer cells to sorafenib are still in
shortage (43). Microarray analysis
and subsequent experiments revealed that metallothionein (MT)-1 may
be a biomarker of modified redox metabolism as well as ferroptosis
in HCC cells exposed to sorafenib (44). Furthermore, MT-1G may serve as a
potential therapeutic target for overcoming sorafenib resistance in
human HCC cells (45). Additionally,
knockdown of MT-1G by RNA interference augmented glutathione
depletion and lipid peroxidation, which promoted sorafenib-induced
ferroptosis (45).
The association between ferroptosis and
other forms of regulated cell death
While ferroptosis is distinct from other forms of
regulated cell death, including apoptosis, necrosis and autophagy,
it is connected with other types of cell deaths (19,30).
Apoptosis
ART and erastin, which are common ferroptosis
inducers, enhance tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL)-induced apoptosis in human pancreatic cancer PANC-1
and BxPC-3 cells as well as human colon cancer HCT116 cells
(19). An increased activation of
caspase 8/9/3, regarded as the hallmark of apoptosis, was observed
in the aforementioned study. In vitro, the ferroptotic
agents stimulated endoplasmic reticulum (ER) stress in human colon
cells (46). Furthermore, they
increased the activation of the death receptor 5 via the TRAIL and
initiated apoptosis (46).
Autophagy
A recent study revealed that autophagy was conducive
to ferroptosis by degrading ferritin in cancer cells. Knockdown of
autophagy-related (ATG) 5 and ATG7 limited erastin-stimulated
ferroptosis by reducing the intracellular ferrous iron
concentration and lipid peroxidation (30). ELAVL1 is a positive regulator of
ferroptosis by inhibiting the ubiquitin-proteasome pathway.
Upregulated ELAVL1 expression revealed increased autophagosome
production coupled with autophagic flux. ELAVL1 promoted autophagy
by binding to the adenine uracil-rich region in the 3′-untranslated
region of beclin 1 (BECN1) mRNA, resulting in BECN1 overexpression
(38).
ER stress
Previous studies have revealed that inhibiting
cystine-glutamate exchange with the aid of ferroptotic agents
activates an ER stress feedback loop and upregulates the
glutathione-specific γ glutamylcyclotransferase 1 gene (47,48).
Hong et al (14) demonstrated
that the unfolded protein response was detected in ART-treated
colon cancer cells, suggesting that the ferroptotic agent induced
ER stress. Furthermore, the ER stress markers HSPA5 and DNA damage
inducible transcript 3 were detected in ferroptotic agent-treated
HCT116 cells, suggesting that ferroptotic agents induce ER stress
(14,19). Additionally, ART-induced ER stress
was inhibited by iron chelators and lipid peroxidation inhibitors,
including ferrostatin-1 and lipoxstatin-1.
Conclusions
The digestive system consists of a number of organs
and cancer of the digestive system has a relatively high morbidity
and mortality rates. Furthermore, patients with digestive system
cancer have a poor quality of life. Ferroptosis, which is a novel
form of regulated cell death, is implicated in several types of
digestive system cancer, excluding esophageal and biliary system
cancer. As described in the present review, iron, lipid and amino
acid metabolism are involved in ferroptosis. Molecules involved in
ferroptosis in digestive system cancer, include TP53, Rb, NRF2,
p62, CDO1, MT-1G, NCOA4, CISD1 and HSPA5. Several drugs, such as
cisplatin, baicalein, haloperidol, ART, PL, bromelain and saponin,
induce cancer cell death by ferroptosis in the digestive system and
exert therapeutic effects. Nevertheless, the signaling pathways and
major transcriptional regulators of ferroptosis in digestive system
cancer remain unknown. Further studies are required to establish
the roles of ferroptosis in metastasis, energy metabolism,
autophagy and drug resistance. Furthermore, the elucidation of
molecular pathways underlying ferroptosis in digestive system
cancer may provide novel therapeutic targets and improve the
prognosis of patients with digestive system cancer.
Acknowledgements
Not applicable.
Funding
The present review was supported by the National
Natural Science Foundation of China (grant no. 81600509) and the
Youth Incubation foundation of Tianjin Medical University General
Hospital (grant no. ZYYFY2016020).
Availability of data and materials
Not applicable.
Authors' contributions
YS and RL conceived and designed the review. YS and
HY drafted the manuscript. KJ and BMW critically revised the
article for intellectual content. All the authors approved the
final version of manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
Not applicable.
References
|
1
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao M, Monian P, Quadri N, Ramasamy R and
Jiang X: Glutaminolysis and transferrin regulate ferroptosis. Mol
Cell. 59:298–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang WS and Stockwell BR: Ferroptosis:
Death by lipid peroxidation. Trends Cell Biol. 26:165–176. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stockwell BR, Friedmann Angeli JP, Bayir
H, Bush AI, Conrad M, Dixon SJ, Fulda S, Gascón S, Hatzios SK,
Kagan VE, et al: Ferroptosis: A regulated cell death nexus linking
metabolism, redox biology and disease. Cell. 171:273–285. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Skouta R, Dixon SJ, Wang J, Dunn DE, Orman
M, Shimada K, Rosenberg PA, Lo DC, Weinberg JM, Linkermann A and
Stockwell BR: Ferrostatins inhibit oxidative lipid damage and cell
death in diverse disease models. J Am Chem Soc. 136:4551–4556.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Friedmann Angeli JP, Schneider M, Proneth
B, Tyurina YY, Tyurin VA, Hammond VJ, Herbach N, Aichler M, Walch
A, Eggenhofer E, et al: Inactivation of the ferroptosis regulator
Gpx4 triggers acute renal failure in mice. Nat Cell Biol.
16:1180–1191. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Linkermann A, Skouta R, Himmerkus N, Mulay
SR, Dewitz C, De Zen F, Prokai A, Zuchtriegel G, Krombach F, Welz
PS, et al: Synchronized renal tubular cell death involves
ferroptosis. Proc Natl Acad Sci USA. 111:16836–16841. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bai T, Wang S, Zhao Y, Zhu R, Wang W and
Sun Y: Haloperidol, a sigma receptor 1 antagonist, promotes
ferroptosis in hepatocellular carcinoma cells. Biochem Biophys Res
Commun. 491:919–925. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun X, Ou Z, Chen R, Niu X, Chen D, Kang R
and Tang D: Activation of the p62-Keap1-NRF2 pathway protects
against ferroptosis in hepatocellular carcinoma cells. Hepatology.
63:173–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sauzay C, Louandre C, Bodeau S, Anglade F,
Godin C, Saidak Z, Fontaine JX, Usureau C, Martin N, Molinie R, et
al: Protein biosynthesis, a target of sorafenib, interferes with
the unfolded protein response (UPR) and ferroptosis in
hepatocellular carcinoma cells. Oncotarget. 9:8400–8414. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hao S, Yu J, He W, Huang Q, Zhao Y, Liang
B, Zhang S, Wen Z, Dong S, Rao J, et al: Cysteine dioxygenase 1
mediates erastin-induced ferroptosis in human gastric cancer cells.
Neoplasia. 19:1022–1032. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jennis M, Kung CP, Basu S, Budina-Kolomets
A, Leu JI, Khaku S, Scott JP, Cai KQ, Campbell MR, Porter DK, et
al: An african-specific polymorphism in the TP53 gene impairs p53
tumor suppressor function in a mouse model. Genes Dev. 30:918–930.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu S, Zhang Q, Sun X, Zeh HJ III, Lotze
MT, Kang R and Tang D: HSPA5 regulates ferroptotic cell death in
cancer cells. Cancer Res. 77:2064–2077. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamaguchi Y, Kasukabe T and Kumakura S:
Piperlongumine rapidly induces the death of human pancreatic cancer
cells mainly through the induction of ferroptosis. Int J Oncol.
52:1011–1022. 2018.PubMed/NCBI
|
|
16
|
Stipanuk MH, Ueki I, Dominy JE Jr, Simmons
CR and Hirschberger LL: Cysteine dioxygenase: A robust system for
regulation of cellular cysteine levels. Amino acids. 37:55–63.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Khoo KH, Verma CS and Lane DP: Drugging
the p53 pathway: Understanding the route to clinical efficacy. Nat
Rev Drug Discov. 13:217–236. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie Y, Song X, Sun X, Huang J, Zhong M,
Lotze MT, Zeh HJ Rd, Kang R and Tang D: Identification of baicalein
as a ferroptosis inhibitor by natural product library screening.
Biochem Biophys Res Commun. 473:775–780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hong SH, Lee DH, Lee YS, Jo MJ, Jeong YA,
Kwon WT, Choudry HA, Bartlett DL and Lee YJ: Molecular crosstalk
between ferroptosis and apoptosis: Emerging role of ER
stress-induced p53-independent PUMA expression. Oncotarget.
8:115164–115178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie Y, Zhu S, Song X, Sun X, Fan Y, Liu J,
Zhong M, Yuan H, Zhang L, Billiar TR, et al: The tumor suppressor
p53 limits ferroptosis by blocking DPP4 activity. Cell Rep.
20:1692–1704. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wei G, Sun J, Hou Z, Luan W, Wang S, Cui
S, Cheng M and Liu Y: Novel antitumor compound optimized from
natural saponin albiziabioside a induced caspase-dependent
apoptosis and ferroptosis as a p53 activator through the
mitochondrial pathway. Eur J Med Chem. 157:759–772. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo J, Xu B, Han Q, Zhou H, Xia Y, Gong C,
Dai X, Li Z and Wu G: Ferroptosis: A novel anti-tumor action for
cisplatin. Cancer Res Treat. 50:445–460. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park S, Oh J, Kim M and Jin EJ: Bromelain
effectively suppresses Kras-mutant colorectal cancer by stimulating
ferroptosis. Anim Cells Syst (Seoul). 22:334–340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pagliara V, Saide A, Mitidieri E,
d'Emmanuele di Villa Bianca R, Sorrentino R, Russo G and Russo A:
5-FU targets rpL3 to induce mitochondrial apoptosis via
cystathionine-β-synthase in colon cancer cells lacking p53.
Oncotarget. 7:50333–50348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Russo A, Saide A, Smaldone S, Faraonio R
and Russo G: Role of uL3 in multidrug resistance in p53-mutated
lung cancer cells. Int J Mol Sci. 18(pii): E5472017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kasukabe T, Honma Y, Okabe-Kado J, Higuchi
Y, Kato N and Kumakura S: Combined treatment with cotylenin A and
phenethyl isothiocyanate induces strong antitumor activity mainly
through the induction of ferroptotic cell death in human pancreatic
cancer cells. Oncol Rep. 36:968–976. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Eling N, Reuter L, Hazin J, Hamacher-Brady
A and Brady NR: Identification of artesunate as a specific
activator of ferroptosis in pancreatic cancer cells. Oncoscience.
2:517–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mancias JD, Wang X, Gygi SP, Harper JW and
Kimmelman AC: Quantitative proteomics identifies NCOA4 as the cargo
receptor mediating ferritinophagy. Nature. 509:105–109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shintoku R, Takigawa Y, Yamada K, Kubota
C, Yoshimoto Y, Takeuchi T, Koshiishi I and Torii S:
Lipoxygenase-mediated generation of lipid peroxides enhances
ferroptosis induced by erastin and RSL3. Cancer Sci. 108:2187–2194.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hou W, Xie Y, Song X, Sun X, Lotze MT, Zeh
HJ III, Kang R and Tang D: Autophagy promotes ferroptosis by
degradation of ferritin. Autophagy. 12:1425–1428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Louandre C, Ezzoukhry Z, Godin C, Barbare
JC, Mazière JC, Chauffert B and Galmiche A: Iron-dependent cell
death of hepatocellular carcinoma cells exposed to sorafenib. Int J
Cancer. 133:1732–1742. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yuan H, Li X, Zhang X, Kang R and Tang D:
CISD1 inhibits ferroptosis by protection against mitochondrial
lipid peroxidation. Biochem Biophys Res Commun. 478:838–844. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geldenhuys WJ, Leeper TC and Carroll RT:
mitoNEET as a novel drug target for mitochondrial dysfunction. Drug
Discov Today. 19:1601–1606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ou W, Mulik RS, Anwar A, McDonald JG, He X
and Corbin IR: Low-density lipoprotein docosahexaenoic acid
nanoparticles induce ferroptotic cell death in hepatocellular
carcinoma. Free Radic Biol Med. 112:597–607. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sporn MB and Liby KT: NRF2 and cancer: The
good, the bad and the importance of context. Nat Rev Cancer.
12:564–571. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jaramillo MC and Zhang DD: The emerging
role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev.
27:2179–2191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ren D, Villeneuve NF, Jiang T, Wu T, Lau
A, Toppin HA and Zhang DD: Brusatol enhances the efficacy of
chemotherapy by inhibiting the Nrf2-mediated defense mechanism.
Proc Natl Acad Sci USA. 108:1433–1438. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Z, Yao Z, Wang L, Ding H, Shao J,
Chen A, Zhang F and Zheng S: Activation of ferritinophagy is
required for the RNA-binding protein ELAVL1/HuR to regulate
ferroptosis in hepatic stellate cells. Autophagy. 14:2083–2103.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hayashi T and Su TP: Sigma-1 receptor
chaperones at the ER-mitochondrion interface regulate Ca(2+)
signaling and cell survival. Cell. 131:596–610. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kourrich S, Hayashi T, Chuang JY, Tsai SY,
Su TP and Bonci A: Dynamic interaction between sigma-1 receptor and
Kv1.2 shapes neuronal and behavioral responses to cocaine. Cell.
152:236–247. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dehart DN, Fang D, Heslop K, Li L,
Lemasters JJ and Maldonado EN: Opening of voltage dependent anion
channels promotes reactive oxygen species generation, mitochondrial
dysfunction and cell death in cancer cells. Biochem Pharmacol.
148:155–162. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Louandre C, Marcq I, Bouhlal H, Lachaier
E, Godin C, Saidak Z, François C, Chatelain D, Debuysscher V,
Barbare JC, et al: The retinoblastoma (Rb) protein regulates
ferroptosis induced by sorafenib in human hepatocellular carcinoma
cells. Cancer Lett. 356:971–977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cao W, Hou FF and Nie J: AOPPs and the
progression of kidney disease. Kidney Int Suppl (2011). 4:102–106.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Houessinon A, François C, Sauzay C,
Louandre C, Mongelard G, Godin C, Bodeau S, Takahashi S, Saidak Z,
Gutierrez L, et al: Metallothionein-1 as a biomarker of altered
redox metabolism in hepatocellular carcinoma cells exposed to
sorafenib. Mol Cancer. 15:382016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun X, Niu X, Chen R, He W, Chen D, Kang R
and Tang D: Metallothionein-1G facilitates sorafenib resistance
through inhibition of ferroptosis. Hepatology. 64:488–500. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lee YS, Lee DH, Jeong SY, Park SH, Oh SC,
Park YS, Yu J, Choudry HA, Bartlett DL and Lee YJ:
Ferroptosis-inducing agents enhance TRAIL-induced apoptosis through
upregulation of death receptor 5. J Cell Biochem. 120:928–939.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dixon SJ, Patel DN, Welsch M, Skouta R,
Lee ED, Hayano M, Thomas AG, Gleason CE, Tatonetti NP, Slusher BS
and Stockwell BR: Pharmacological inhibition of cystine-glutamate
exchange induces endoplasmic reticulum stress and ferroptosis.
Elife. 3:e025232014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Dai Z, Huang Y, Sadee W and Blower P:
Chemoinformatics analysis identifies cytotoxic compounds
susceptible to chemoresistance mediated by glutathione and
cystine/glutamate transport system xc. J Med Chem. 50:1896–1906.
2007. View Article : Google Scholar : PubMed/NCBI
|