Introduction
Lung cancer is among the most common malignancies in
both men and women worldwide (1).
The majority of patients with lung cancer are diagnosed when the
disease reaches the advanced stages, resulting in a 5-year survival
rate of only 3–7% (2). The poor
prognosis makes lung cancer one of the leading causes of
cancer-associated mortality, with 1.8 million estimated novel cases
and 1.6 million estimated mortalities per year in the United States
(3). The majority of patients
present with a locally metastatic condition due to the high
metastatic potential of lung cancer cells (4). In addition, oncogenic drivers, such as
mutations of epidermal growth factor receptor (EGFR), Ki-ras2
Kirsten rat sarcoma viral oncogene homolog, human epidermal growth
factor receptor 2 and/or ROS proto-oncogene 1 have also contributed
to the development and progression of lung cancer, which means that
personalized and genotype-directed therapy, as well as novel
immunotherapies (programmed death-1 or programmed death-ligand 1),
have revolutionized the management of lung cancer (5,6). It is
for this reason that an increasing amount of researchers have been
focusing on identifying novel molecular regulators of lung
cancer.
Long non-coding RNAs (lncRNAs) are a class of RNAs
that consist of >200 nucleotides and lack protein-coding
abilities (7,8). Previous high-throughput transcriptome
analyses have revealed that >90% of the genes are transcribed
into non-coding RNAs, including lncRNAs, which are predicted to
regulate chromatin or function as genetic regulators, depending on
their location relative to the nucleus (9,10). To
date, >3,000 lncRNAs have been identified, 1% of which only have
had their roles identified (9).
lncRNAs are suggested to be classified into five categories
depending on their origin (11) and
have three subtypes based on genomic locations (12), namely intergenic lncRNAs, intronic
lncRNAs and antisense lncRNAs.
During recent decades, lncRNAs have been revealed to
serve crucial roles in various molecular genetics and cellular
processes (13), such as chromosomal
dosage compensation, maintenance of chromatin structure, splicing,
cellular differentiation, cell cycle and tumorigenesis (14). For instance, lncRNA sex determining
region Y-box 2 overlapping transcript was reported to regulate cell
proliferation and identified as a poor survival indictor in human
lung cancer (15). LncRNA SPRY4-IT1
was identified as an emerging factor in tumorigenesis of
osteosarcoma (16).
Overexpressed in colorectal cancer (OECC) is a newly
identified lncRNA, which originates from chromosome 8q24 and has
been revealed as being highly expressed in human colorectal
carcinoma (CRC) (17). However, its
detailed role and molecular mechanism in other types of tumor
remained largely unknown. The aim of the present study was to
identify the effects of OECC on cell proliferation and cell
metastasis in human lung cancer and sought to uncover the
underlying molecular mechanisms. To the best of our knowledge, the
present study is the first to investigate the role of OECC in lung
cancer, which may provide novel clues for the clinical treatment of
lung cancer.
Materials and methods
Human samples
The present study was approved by the Ethical
Committee of Guangzhou Medical University (Guangzhou, China) A
total of 50 patients with lung cancer (male/female, 34:16; age
range, 59±9 years) were recruited between January 2016 and December
2016 at the First Affiliated Hospital of Guangzhou Medical
University and included in the present study. Patients who had
received chemotherapy or radiotherapy treatments prior to surgery
were excluded from the study. The tumor tissues and adjacent
non-tumorous tissues of the patients were dissected during surgery
and immediately frozen in liquid nitrogen. Signed informed consent
was provided by all patients.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from human samples and
cultured cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) in a volume of 1 ml for each well in
6-well plates.RNAs were quantified using a Nanodrop™ 2000
instrument (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. A total of 1 µg RNA was transcribed into
cDNA using reverse transcriptase (Takara Biotechnology Co., Ltd.)
with the following protocols: 37°C for 15 min and 85°C for 5 sec.
Then, qPCR was performed in an ABI 7900 machine (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with hot start Taq DNA
polymerase and SYBR® Green (Takara Biotechnology Co.,
Ltd.). The procedure was as follows: Initial denaturation at 95°C
for 5 min, followed by 45 repeats of a three-step cycling program
consisting of 10 sec at 95°C (denaturation), 10 sec at 60°C (primer
annealing) and 10 sec at 72°C (elongation), and a final extension
step for 10 min at 72°C. The primers used were: OECC forward,
5′-AACCGTAGGAGCACATCACAG-3′ and reverse,
5′-CCGTGGTTTCAGTTGCCCTA-3′; phosphoinositide 3-kinase (PI3K)
forward, 5′-GTCCTATTGTCGTGCATGTGG-3′ and reverse,
5′-TGGGTTCTCCCAATTCAACC-3′; protein kinase B (Akt) forward,
5′-TTCTATGGCGCTGAGATTGTGT-3′ and reverse,
5′-GCCGTAGTCATTGTCCTCCAG-3′; mammalian target of rapamycin (mTOR)
forward, 5′-ATGCTTGGAACCGGACCTG-3′ and reverse,
5′-TCTTGACTCATCTCTCGGAGTT-3′; and GAPDH forward,
5′-GTGGACATCCGCAAAGAC-3′ and reverse, 5′-AAAGGGTGTAACGCAACTA-3′.
GAPDH was included as an internal control. The 2−ΔΔCq
method was used to calculate the relative expression normalized to
GAPDH (18).
Cell culture and transfection
Normal lung epithelial cell line BEAS-2B and human
lung cancer cell lines H1975 and SPC-A-1 were purchased from the
American Type Culture Collection (ATCC). Other lung cancer cell
lines A549, 95D and H-125 were from the Cell Bank of the Chinese
Academy of Sciences. All the cell lines were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) supplied with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.) at 37°C with 5% CO2. The
culture medium was replaced once every 2 days, unless otherwise
stated. Short hairpin (sh)RNA against OECC (shOECC) were designed
by Shanghai GenePharma Co., Ltd. and a negative control shRNA
(shNC) was included as a control. Akt-expressing plasmid (pLNCX1 HA
Akt1) was purchased from Addgene, Inc. (cat. no. 15990). A total of
1×105 cells were transfected with 2 µg plasmid using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h at 37°C according to the manufacturer's
protocol.
Colony formation assay
A549 and 95D cells were seeded in 12-well plates in
DMEM in triplicate (100 cells/well) and transfected with specific
shRNA against OECC. Subsequently, the plates were incubated in a
37°C incubator for 14 days and the colonies that contained >50
cells were fixed with pre-iced methanol for 10 min at room
temperature and stained with crystal violet (1%) for 5 min at room
temperature. Colonies were counted under a light microscope (Nikon
Corporation) at a magnification of ×200.
5-Ethynyl-2′-deoxyuridine (EdU) cell
proliferation assay
A549 and 95D cells were seeded into 24-well plates
and transfected with or without shOECC. At 48 h post-transfection,
medium was replaced with complete DMEM supplemented with 50 µM EdU
(Thermo Fisher Scientific, Inc.) and further incubated at 37°C for
2 h. Following washing with ice-cold PBS twice, the cells were
fixed with 4% polyoxymethylene containing 0.5% Triton™ X-100
(Sigma-Aldrich; Merck KGaA) for 5 min at room temperature and then
stained with Apollo dye (Thermo Fisher Scientific, Inc.) for 30 min
at 37°C. Following staining with DAPI (1:1,000; 10 min at room
temperature), cells were imaged with a light microscope (Nikon
Corporation) at a magnification of ×200.
Cell viability assay
A549 and 95D cells were seeded into chamber slides
and transfected with shOECC and cultured in a 37°C incubator for 48
h with co-incubation of G418 (Sigma-Aldrich; Merck KGaA). Cell
viability was assessed using the Cell Counting Kit-8 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Relative proliferative rate was assessed by determining the mean
and standard deviations of five randomly selected image fields. To
evaluate the overall survival rate, a total of 1×103
A549 and 95D cells were seeded into 96-well plates and transfected
with shRNAs in triplicate. Following selection for 6 days, overall
survival was examined with 0.5 mg/ml thiazolyl blue tetrazolium
blue reagent (Sigma-Aldrich; Merck KGaA) and calculated at 590 nm
using a Tecan microplate reader (Tecan Group, Ltd.).
Transwell and Matrigel assay
A549 and 95D were seeded into 6-well plates and
transfected with shRNAs against OECC for 72 h. Cells were washed,
trypsinized and collected by centrifuge (1,000 × g for 5 min at
4°C. Approximately 1×105 cells were seeded into the
upper chamber in DMEM without FBS. A volume of 600 µl complete DMEM
(supplied with 10% FBS) were added into the lower chamber. The
chambers were incubated for another 24 h, fixed with ice-cold
methanol for 5 min at room temperature and stained with crystal
violet (1%) for 5 min at room temperature. The upper chambers were
scraped with a cotton swab and the lower chamber was photographed
under a light microscope (Nikon Corporation) with five randomly
selected image fields. For the cell invasion assay, the membrane
was pre-coated with Matrigel (Corning Inc., Corning, NY, USA) for 6
h at 37°C.
RNA sequence analysis
A549 cells were transfected with or without shOECC
for 72 h and the total RNAs were extracted for RNA-sequencing (seq)
in triplicate. RNA-seq was performed and analyzed by AnNuo Co.
Different signaling pathways were investigated and the various
genes (P<0.05) were classified into corresponding signaling
pathways.
Wound-healing assay
A total of 1×105 A549 and 95D cells were
seeded into 6-well plates and co-incubated with the same amount of
shRNAs (shNC or shOECC) for 72 h. Wound-healing assays were
performed by creating identical wound areas with 10 µl pipette tips
for anchorage-dependent A549 and 95D cells. Cells were washed with
PBS three times and replaced with fresh, serum-free medium
immediately. Images of the cells were captured once the scratch was
made (0 h). After 24 h of proliferation, cells were also observed
and images were captured under light microscope (Nikon Corporation)
at a magnification of ×200 for each group.
Statistical analysis
GraphPad Prism software (version 5.0; GraphPad
Software, Inc.) was used for statistical analysis. Independent
Student's t-test was used for comparisons between groups, whereas
differences between tumor and adjacent normal control samples were
analyzed using a paired Student's t-test. For comparisons among
multiple groups (≥3 groups), one-way analysis of variance was
applied, followed by a least significance difference post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
LncRNA OECC is overexpressed in human
lung cancer in vivo and in vitro
In total, 50 patients with clinical lung cancer were
involved in the present study. Tumor tissues and adjacent
non-cancerous tissues were collected from the patients for RT-qPCR
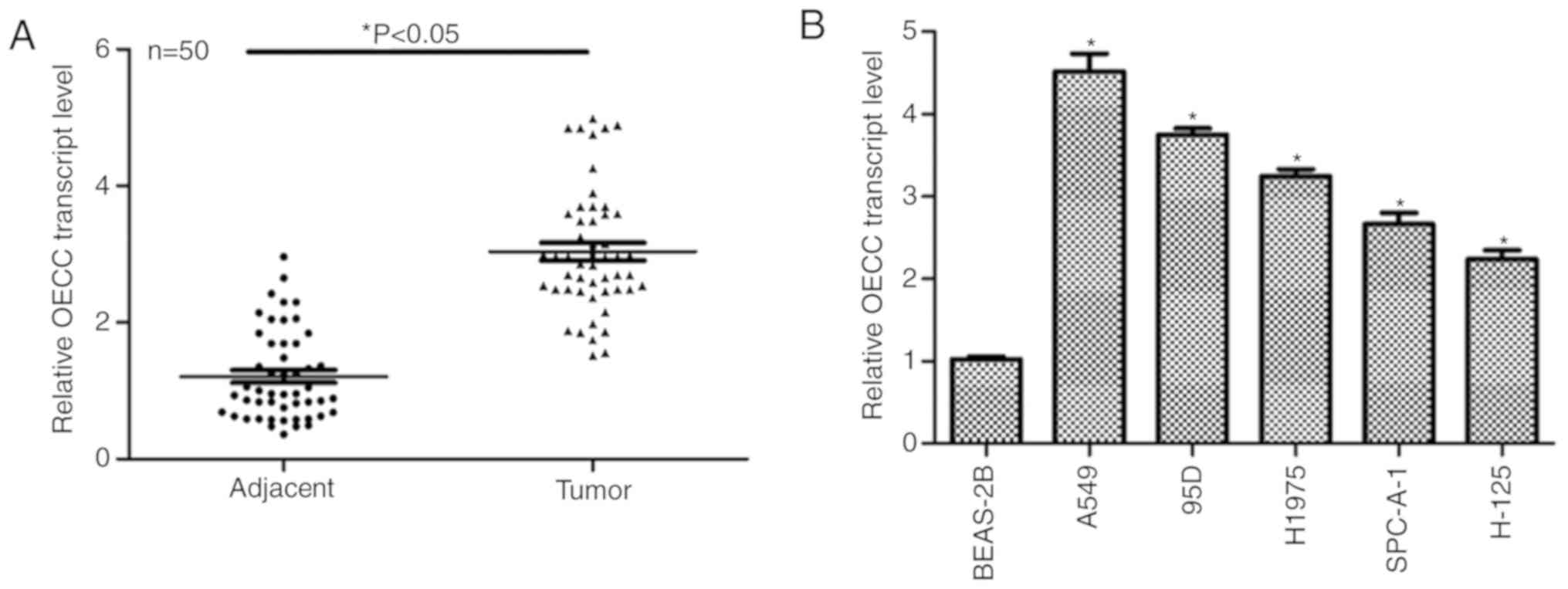
analysis. As presented in Fig. 1A,
the relative OECC transcript level was markedly increased in tumor
tissues compared with in their adjacent non-cancerous counterparts.
The expression of OECC was then detected in vitro. As
presented in Fig. 1B, the transcript
level of OECC was notably upregulated in all the lung cancer cell
lines as compared with in the normal lung epithelial cell line
BEAS-2B. Notably, A549 and 95D cells exhibited the highest OECC
expression of all the lung cancer cell lines. Thus, these two cell
lines were chosen for the subsequent functional assays. These data
suggested that the transcript level of OECC was upregulated in
human lung cancer in vivo and in vitro.
Knockdown of OECC inhibits cell
proliferation in A549 and 95D cells
Next, the expression of OECC was knocked down by
specific shOECC in order to investigate the detailed roles of OECC
in human lung cancer. As presented in Fig. 2A, three shRNAs against OECC were
designed and transfected into A549 and 95D cells and only the first
shRNA was effective, thus it was chosen for the subsequent analysis
and renamed as shOECC. Colony formation assays revealed >150
colonies in the A549 control cells compared with only 75 colonies
in A549 cells following transfection with shOECC. Likewise,
transfection of shOECC resulted in the decrease of almost 50% of
95D cells (Fig. 2B). The results
from the EdU assays, presented in Fig.
2C and D, demonstrated that depletion of OECC in A549 cells
resulted in a decrease in EdU positive cells, indicating the
effects that OECC suppression has on cell proliferation. Cell
viability assays were performed in A549 and 95D cells transfected
with shOECC for 5 consecutive days. No clear difference was
observed in the first 3 days among the three groups of A549 and 95D
cells; however, the rate of cell proliferation was suppressed by
almost 20% on the fourth day and 25% on the fifth day in A549 cells
(Fig. 2E). Similarly, the cell
viability of 95D cells was also inhibited by the knockdown of OECC
on the fourth and fifth days (Fig.
2F). These data collectively suggested that depletion of OECC
in A549 and 95D cells inhibited cell proliferation in
vitro.
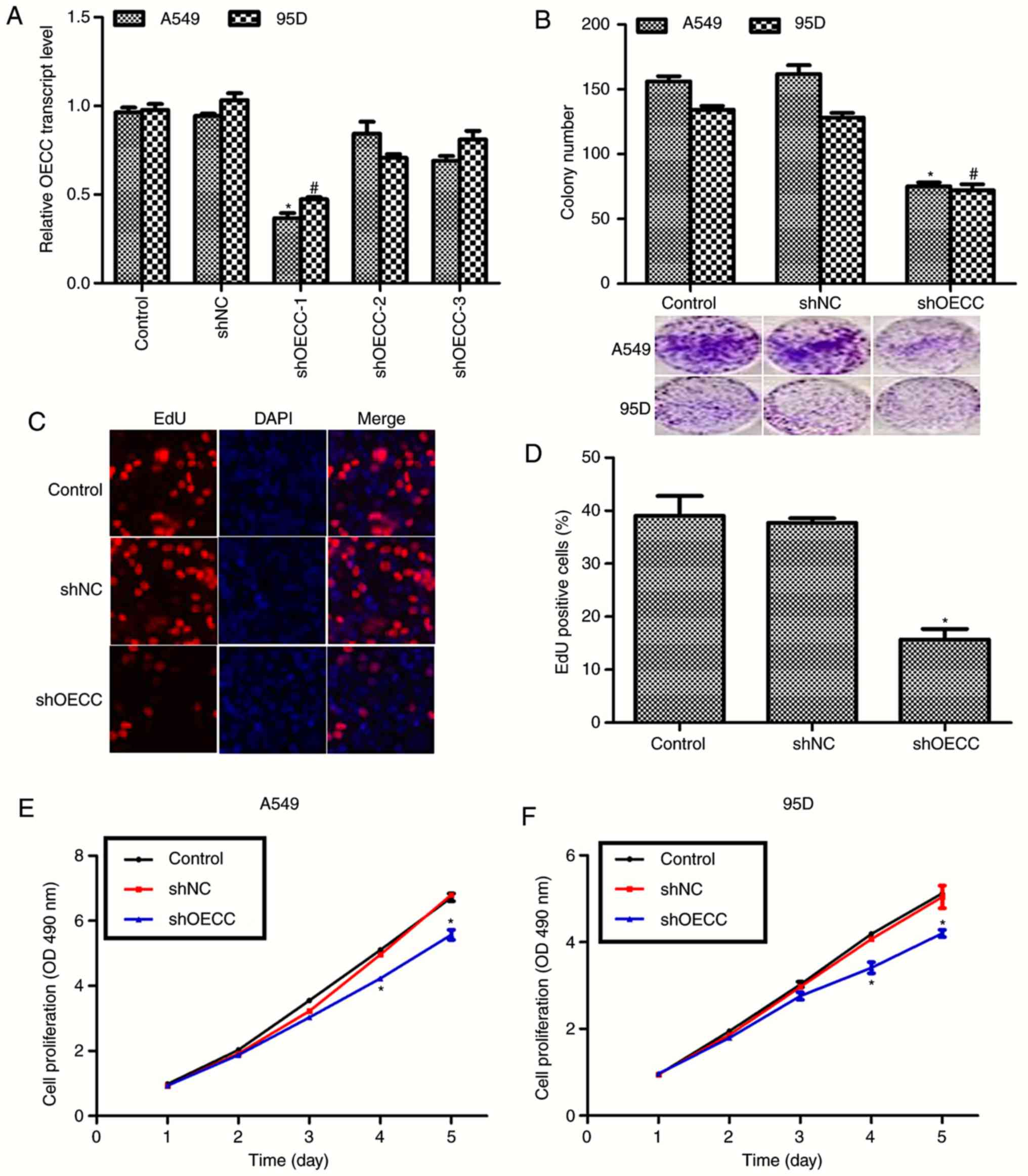 | Figure 2.Knockdown of OECC inhibits cell
proliferation in A549 and 95D cells. (A) RT-qPCR analysis was
performed in A549 and 95D cells transfected with shRNA against
OECC. (B) Colony formation assays were performed in A549 and 95D
cells transfected with shRNA against OECC. The images included were
the representative photos of colony formation assays. *P<0.05,
vs. control in A549 cells. #P<0.05, vs. control in
95D cells. (C) Representative images of EdU assays in A549 cells,
(magnification, ×200). (D) Quantification of EdU assays in A549
cells transfected with shOECC. (E) Cell proliferation assays were
performed in A549 cells over 5 consecutive days when the cells were
treated with shOECC. (F) Cell proliferation assays were performed
in 95D cells over 5 consecutive days when the cells were treated
with shOECC. *P<0.05, vs. control. OECC, overexpressed in
colorectal cancer; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; shRNA, short hairpin RNA; EdU,
5-ethynyl-2′-deoxyuridine; shOECC, short hairpin RNA against OECC;
OD, optical density; shNC, negative control short hairpin RNA. |
Knockdown of OECC in A549 and 95D
cells suppresses cell metastasis in vitro
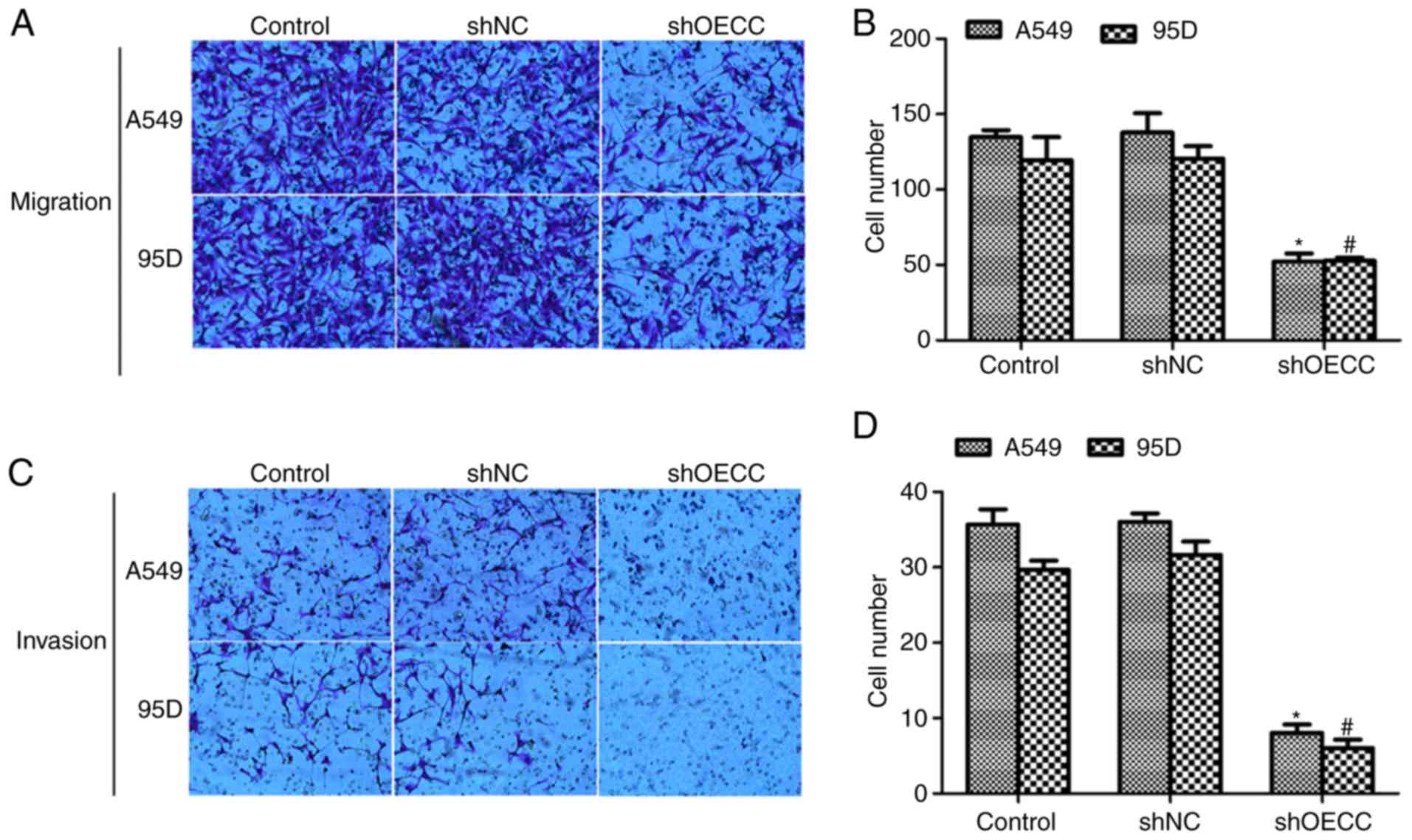
As presented in Fig. 3A
and B, transfection of shOECC into A549 and 95D cells delayed
cell migration through the membrane. In the control group, ~35 A549
cells and ~30 95D cells invaded the membrane whereas only ~10 A549
cells and ~8 9 5D cells were observed on the lower surface of the
membrane (Fig. 3C and D). These data
suggested that depletion of OECC inhibited cell metastasis in human
lung cancer cells in vitro.
Knockdown of OECC in A549 cells
regulates the PI3K/Akt/mTOR signaling pathway
To elucidate the detailed regulatory molecular
mechanism of OECC in cell proliferation and cell metastasis, a
RNA-seq analysis was performed. Of all the pathways observed to be
altered, the PI3K/Akt/mTOR signaling pathway was the most marked.
As presented in Fig. 4, knockdown of
OECC in A549 cells decreased the mRNA levels of PI3K,
phosphoinositide-dependent kinase-1, Akt, 5′-AMP-activated protein
kinase and endothelial nitric synthase etc., and increased
expression of genes such as tumor protein 53, neurofibromin 1 and
regulator of cullins-1, which are all part of and/or involved in
crosstalk with the PI3K/Akt/mTOR signaling pathway. These results
suggested that OECC expression was closely associated with the
PI3K/Akt/mTOR signaling pathway.
OECC regulates cell proliferation and
cell metastasis through the PI3K/Akt/mTOR signaling pathway in
human lung cancer
In the present study, the role of the PI3K/Akt/mTOR
signaling pathway in the function of OECC in lung cancer was
investigated. To this end, Akt-expressing plasmid was
co-transfected with shOECC into A549 cells. As presented in
Fig. 5A, the mRNA levels of OECC,
PI3K, Akt and mTOR were significantly decreased when OECC was
knocked down in A549 cells, which was consistent with the result of
RNA-seq analysis (Fig. 4).
Furthermore, co-incubation of Akt-expressing plasmid increased the
Akt and mTOR mRNA levels, but not OECC and PI3K, which hinted that
OECC was the upstream regulator of PI3K/Akt/mTOR signaling. The
activated forms of PI3K and Akt were also investigated, namely
p-PI3K and p-Akt. Fig. 5B indicates
that knockdown of OECC in A549 cells decreased the protein levels
of p-PI3K and p-Akt, whereas in contrast, co-overexpression of Akt
increased the expression of p-PI3K and p-Akt. Furthermore,
knockdown of OECC with specific shRNA inhibited the cells' ability
to heal the wound, and in contrast, co-overexpression of Akt
increased cell migration abilities in the A549 cells (Fig. 5C and E) and the 95D cells (Fig. 5D and E). Subsequently, depletion of
OECC inhibited the formation of colonies in A549 and 95D cells, but
co-treatment with Akt-expressing plasmids reversed these effects
(Fig. 5F). These data suggested that
OECC regulated cell proliferation and cell metastasis through the
PI3K/Akt/mTOR signaling pathway.
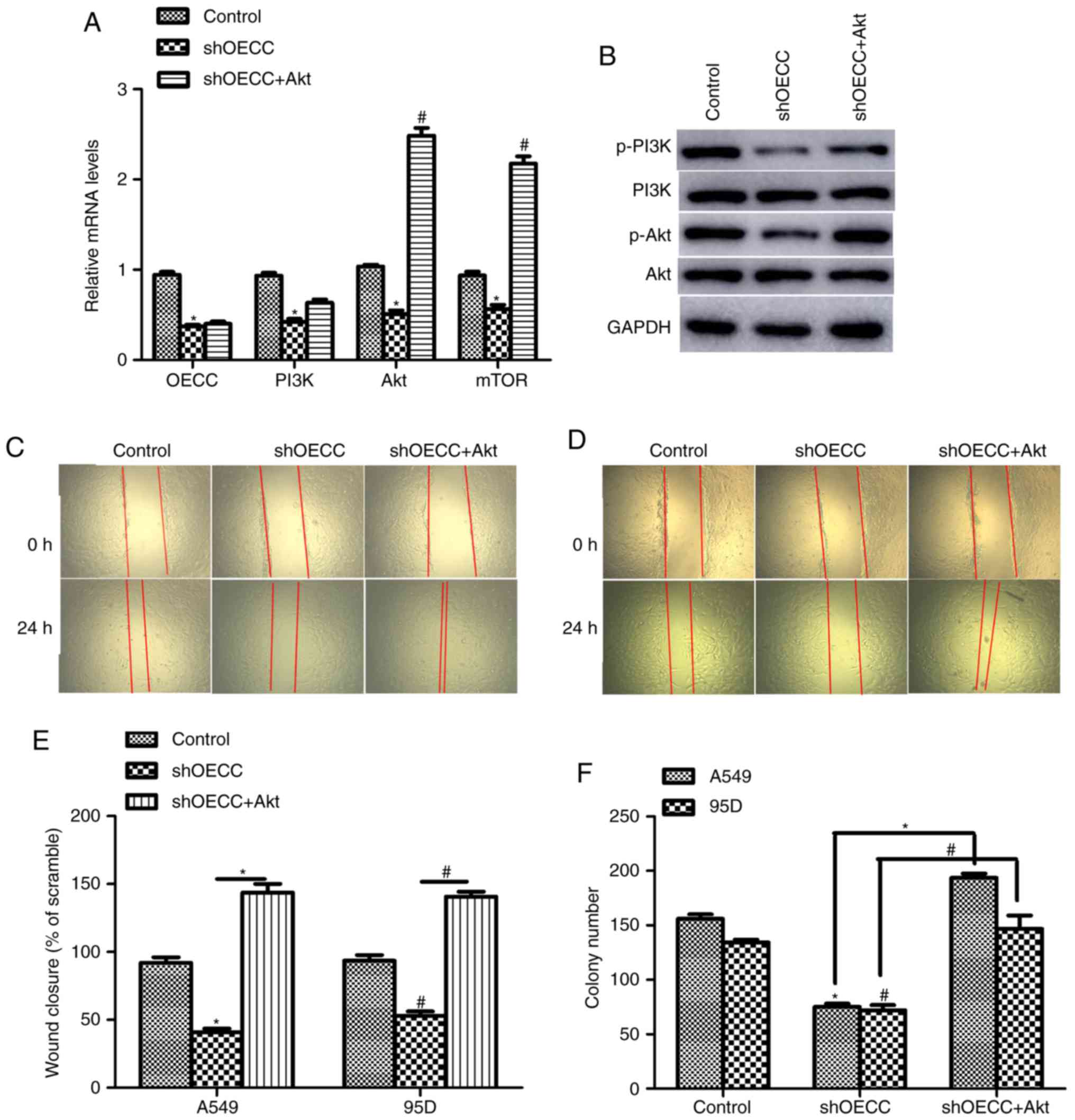 | Figure 5.OECC regulates cell proliferation and
cell metastasis through the PI3K/Akt/mTOR signaling pathway in
human lung cancer. (A) The relative mRNA levels of OECC, PI3K, Akt
and mTOR in A549 cells transfected with shOECC in the presence or
absence of Akt-expressing plasmid. *P<0.05, shOECC vs. control.
#P<0.05, shOECC+Akt vs. shOECC. (B) Western blot
assays were performed in A549 cells treated with shOECC. (C)
Representative images of wound-healing assays in A549 cells. (D)
Representative images of wound-healing assays in 95D cells. (E)
Quantification assay of wound-healing assay in A549 and 95D cells
transfected with shOECC in the presence or absence of
Akt-expressing plasmid. *P<0.05, shOECC vs. control.
#P<0.05, shOECC+Akt vs. shOECC. (F) Colony formation
assays were used in both A549 and 95D cells transfected with shOECC
in the presence or absence of Akt-expressing plasmid. *P<0.05,
vs. control or as indicated in A549 cells. #P<0.05,
vs. control or as indicated in 95D cells. PI3K, phosphoinositide
3-kinase; Akt, protein kinase-B; mTOR, mammalian target of
rapamycin; p, phosphorylated; shOECC, short hairpin RNA against
OECC; shNC, negative control short hairpin RNA. |
Discussion
Lung cancer remains a marked threat to human health.
The overall survival rate continues to be low despite efforts to
make improvements over the last few decades (19). Recently, studies have revealed that
aberrant transcript levels of lncRNA is involved in the
tumorigenesis and tumor progression of human lung cancer (19–21). Of
note, there are multiple genes located on chromosome q824 that
increase the risk of developing lung cancer, such as CCAT1
(20–22), which promoted cell metastasis via
epithelial-to-mesenchymal transition in lung adenocarcinoma
(20) and the Wnt signaling in
non-small cell lung cancer (21).
The present study also indicated that the novel lncRNA OECC,
located on chromosome band q824, upregulated cell proliferation and
metastasis in human lung cancer. However, the limitations of the
present study are that the subtype of lung cancer tissues was not
determined, and the correlation between OECC expression and EGFR
inhibitors was not investigated; these issues will form the basis
of future studies.
LncRNA OECC was first identified by Huang et
al (17) in human CRC, where
OECC was revealed to be overexpressed in human CRC tissues and
cultured CRC cells. Attenuation of OECC inhibited CRC cell
proliferation and metastasis, and increased cell apoptosis, since
OECC was demonstrated to be a direct target of microRNA-143-3p,
leading to the downregulation of its target genes, including
nuclear factor-κB and p38 mitogen-activated protein kinase
signaling pathways (17). In the
present study, OECC was also demonstrated to be upregulated in lung
cancer and associated with cell proliferation and metastasis.
Furthermore, with the aid of the RNA-seq technique, it was
identified that OECC regulated lung cancer progression through
PI3K/Akt/mTOR signaling. Thousands of genes were identified to be
altered when A549 cells were transfected with shOECC; the present
study classified these genes into associated pathways and revealed
that the genes in the PI3K/Akt/mTOR signaling pathway were the most
marked and, thus, this pathway was investigated further. Comparing
with the aforementioned study (17),
depletion of OECC in CRC and lung cancer resulted in similar
phenotypes; however, the underlying molecular mechanism was
markedly different.
PI3K/Akt/mTOR signaling is an intracellular pathway
that regulates the cell cycle, making it directly associated with
cell proliferation, cellular quiescence, cancer and longevity
(23). Activation of PI3K
phosphorylates and activates Akt, resulting in its translocation
onto the plasma membrane and activate multiple downstream effects,
including the inhibition of p27 (24) and activation of cAMP response
element-binding protein and mTOR (25). The present study revealed that the
downregulation of OECC decreased the mRNA levels of PI3K, Akt and
mTOR, and inhibited signaling of the whole pathway. Thus,
PI3K/Akt/mTOR signaling was activated when OECC was depleted in
A549 and 95D cells. Since a PI3K-expressing plasmid could not be
commercially purchased, Akt was overexpressed as an alternative,
using its expression plasmid to activate this signaling. It was
revealed that overexpression of Akt significantly blunted the
suppression effects of OECC on cell proliferation and cell
metastasis, which reinforced the conclusion that OECC regulated
lung cancer progression through the PI3K/Akt/mTOR signaling
pathway. However, the detailed molecular mechanism underlying how
OECC regulates the PI3K/Akt/mTOR signaling pathway remains unknown.
The PI3K regulatory subunit has two major isoforms, p85α and P85β,
which has been identified as being regulated by a newly identified
lncRNA, AK023948 (26), which has
been demonstrated to interact with multiple DNAs, RNAs and
proteins, which may provide hints for investigating the regulatory
details of the PI3K/Akt/mTOR signaling by OECC. Indeed, the present
study is just a preliminary investigation of the role of OECC in
human lung cancer and, thus, further mechanistic studies should be
performed in order to fully corroborate the role of OECC in human
lung cancer.
In conclusion, the present study revealed that the
recently discovered oncogenic lncRNA, OECC, is also involved in
human lung cancer. Depletion of lncRNA OECC suppressed cell
proliferation and metastasis in A549 and 95D cells. These results
may provide novel clues for the treatment of lung cancer in the
clinical setting.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YZ and BZ performed the experiments. YM and HZ
helped analyzing data and revised the manuscript. WH designed the
project and prepared the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Committee of Guangzhou Medical University (Guangzhou, China).
Signed informed consent was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang T, Nelson RA, Bogardus A and Grannis
FW Jr: Five-year lung cancer survival: Which advanced stage
nonsmall cell lung cancer patients attain long-term survival?
Cancer. 116:1518–1525. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tracey E, Roder D, Zorbas H, Villanueva E,
Jelfs P and Bishop J: Survival and degree of spread for female
breast cancers in New South Wales from 1980 to 2003: Implications
for cancer control. Cancer Causes Control. 19:1121–1130. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levy A and Doyen J: Metformin for
non-small cell lung cancer patients: Opportunities and pitfalls.
Crit Rev Oncol Hematol. 125:41–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DeSantis CE, Siegel RL, Sauer AG, Miller
KD, Fedewa SA, Alcaraz KI and Jemal A: Cancer statistics for
African Americans, 2016: Progress and opportunities in reducing
racial disparities. CA Cancer J Clin. 66:290–308. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Herbst RS, Baas P, Kim DW, Felip E,
Pérez-Gracia JL, Han JY, Molina J, Kim JH, Arvis CD, Ahn MJ, et al:
Pembrolizumab versus docetaxel for previously treated,
PD-L1-positive, advanced non-small-cell lung cancer (KEYNOTE-010):
A randomised controlled trial. Lancet. 387:1540–50. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Solomon BJ, Mok T, Kim DW, Wu YL, Nakagawa
K, Mekhail T, Felip E, Cappuzzo F, Paolini J, Usari T, et al:
First-line crizotinib versus chemotherapy in ALK-positive lung
cancer. N Engl J Med. 371:2167–2177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chu C, Spitale RC and Chang HY:
Technologies to probe functions and mechanisms of long noncoding
RNAs. Nat Struct Mol Biol. 22:29–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hutchinson JN, Ensminger AW, Clemson CM,
Lynch CR, Lawrence JB and Chess A: A screen for nuclear transcripts
identifies two linked noncoding RNAs associated with SC35 splicing
domains. BMC Genomics. 8:392007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yunusov D, Anderson L, DaSilva LF, Wysocka
J, Ezashi T, Roberts RM and Verjovski-Almeida S: HIPSTR and
thousands of lncRNAs are heterogeneously expressed in human
embryos, primordial germ cells and stable cell lines. Sci Rep.
6:327532016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Clark MB and Mattick JS: Long noncoding
RNAs in cell biology. Semin Cell Dev Biol. 22:366–376. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen J, Wang R, Zhang K and Chen LB: Long
non-coding RNAs in non-small cell lung cancer as biomarkers and
therapeutic targets. J Cell Mol Med. 18:2425–2436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hou Z, Zhao W, Zhou J, Shen L, Zhan P, Xu
C, Chang C, Bi H, Zou J, Yao X, et al: A long noncoding RNA Sox2ot
regulates lung cancer cell proliferation and is a prognostic
indicator of poor survival. Int J Biochem Cell Biol. 53:380–388.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Z, Shen J, Chan MTV and Wu WKK: The
long non-coding RNA SPRY4-IT1: An emerging player in tumorigenesis
and osteosarcoma. Cell Prolif. 51:e124462018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang F, Wen C, Zhuansun Y, Huang L, Chen
W, Yang X and Liu H: A novel long noncoding RNA OECC promotes
colorectal cancer development and is negatively regulated by
miR-143-3p. Biochem Biophys Res Commun. 503:2949–2955. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mari-Alexandre J, Diaz-Lagares A, Villalba
M, Juan O, Crujeiras AB, Calvo A and Sandoval J: Translating cancer
epigenomics into the clinic: Focus on lung cancer. Transl Res.
189:76–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin H, Cheng W, Yan H and Zhang X:
Overexpression of the long noncoding RNA CCAT1 promotes metastasis
via epithelial-to-mesenchymal transition in lung adenocarcinoma.
Oncol Lett. 16:1809–1814. 2018.PubMed/NCBI
|
|
21
|
Xu C, Xiao G, Zhang B, Wang M, Wang J, Liu
D, Zhang J, Ren H and Sun X: CCAT1 stimulation of the symmetric
division of NSCLC stem cells through activation of the Wnt
signalling cascade. Gene Ther. 25:4–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu B, Zhang H, Wang Z, Zhang F, Wei H and
Li L: LncRNA CCAT1/miR-130a-3p axis increases cisplatin resistance
in non-small-cell lung cancer cell line by targeting SOX4. Cancer
Biol Ther. 18:974–983. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
King D, Yeomanson D and Bryant HE: PI3King
the lock: Targeting the PI3K/Akt/mTOR pathway as a novel
therapeutic strategy in neuroblastoma. J Pediatr Hematol Oncol.
37:245–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rafalski VA and Brunet A: Energy
metabolism in adult neural stem cell fate. Prog Neurobiol.
93:182–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Man HY, Wang Q, Lu WY, Ju W, Ahmadian G,
Liu L, D'Souza S, Wong TP, Taghibiglou C, Lu J, et al: Activation
of PI3-kinase is required for AMPA receptor insertion during LTP of
mEPSCs in cultured hippocampal neurons. Neuron. 38:611–624. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Koirala P, Huang J, Ho TT, Wu F, Ding X
and Mo YY: LncRNA AK023948 is a positive regulator of AKT. Nat
Commun. 8:144222017. View Article : Google Scholar : PubMed/NCBI
|