Introduction
Colon cancer is the third most frequently occurring
cancer in the United States, following prostate cancer in males (or
breast cancer in females) and lung cancer (1). In the Republic of Korea, colon cancer
is the third most commonly diagnosed type of cancer, following
thyroid and stomach cancer, and in 2014, the mortality rate of
patients with colon cancer was the fourth highest of all
cancer-associated deaths (2).
Therefore, the development of novel anti-colorectal cancer drugs is
urgently required. Numerous types of anti-cancer drug have been
developed, and a select few, including 5-fluorourical
(antimetabolite), oxaliplatin (a platinum-based antineoplastic),
irinotecan (a topoisomerase inhibitor) and bevacizumab (a
monoclonal antibody) are commercially available for the treatment
of colon cancer. These drugs are used in different combinations and
dosages depending on the cancer stage and condition of the patient
(3).
Anti-cancer drugs exert their effects by inducing
apoptosis in cancer cells. Apoptosis is a distinct form of
programmed cell death and apoptotic cells possess certain
characteristic features. Morphologically, apoptotic cells exhibit
shrinkage, pyknosis and the formation of apoptotic bodies (4–6).
Apoptosis incorporates a number of crucial biochemical steps. In
the cytosol, BH3-interacting domain death agonist (Bid) becomes
truncated and translocates to the mitochondria (7). In addition, the phosphorylation of
Bcl-2-associated agonist of cell death (Bad) induced by
phospho-protein kinase B (p-Akt) is interrupted (8). Upon apoptotic signal transduction,
cytochrome C is released from mitochondria, causing the activation
of pro-caspase-9 by forming an apoptosome with apoptotic
protease-activating factor 1. In turn, activated caspase-9 is able
to activate pro-caspase-3 by proteolytic cleavage (9). During apoptosis, activated caspases
promote the degradation or cleavage of diverse cytoskeletal and
nuclear proteins, including poly(ADP-ribose) polymerase (PARP)
(10).
Various MHY compounds that exert growth inhibitory
effects against different types of cancer have previously been
synthesized; these include the hydroxamic acid derivatives MHY218
and MHY219. MHY218 enhanced apoptotic cell death and exhibited
anti-cancer activity in gastric, colon, breast and ovarian cancer.
In the AGS human gastric cancer cell line, MHY218 inhibited
autophagy and enhanced apoptotic cell death (11–14).
MHY219, a novel histone deacetylase inhibitor, promoted the
upregulation of androgen receptor expression levels, and induced
apoptosis in human prostate cancer cells (15,16). The
oxirane derivatives MHY336, MHY407, MHY412 and MHY449 have also
been investigated. MHY336, a novel epoxypropoxy flavonoid, induced
apoptosis and cell cycle arrest in colon and prostate cancer cells
by inhibiting topoisomerase II (17,18).
Additionally, the novel carbazole derivative MHY407 produced
anti-cancerous effects in breast cancer cells by sensitizing the
cells to doxorubicin-, etoposide- and radiation-induced DNA damage
(19). The anthracene derivative
MHY412 induced apoptosis in doxorubicin-resistant breast cancer
cells by modulating cell cycle arrest and the downregulation of
P-glycoprotein expression (20).
Furthermore, MHY449, a novel
dihydrobenzofuro[4,5-b][1,8]naphthyridine-6-one derivative, exerted
potent anti-cancerous effects on prostate, gastric and lung cancer
cells. Furthermore, in human lung cancer cells, MHY449 induced cell
cycle arrest and apoptosis by inhibiting the activation of Akt
(21–24). Further studies have also reported
that benzylidene compounds exert anti-cancerous effects in various
types of cancer cells by inducing apoptosis (25–27).
Therefore in the present study, a novel benzylidene derivative,
(Z)-5-(3-ethoxy-4-hydroxybenzylidene)- 2-thioxothiazolidin-4-one
(MHY695) was synthesized and investigated for cytotoxic effects in
human colon cancer cells.
Materials and methods
Synthesis of MHY695
A suspension of 3-ethoxy-4-hydroxybenzaldehyde
(purity, 99%; 300 mg; 1.81 mmol; Sigma-Aldrich; Merck KGaA) and
rhodanine (purity, 97%; 2-thioxothiazolidin-4-one; 264.5 mg; 1.99
mmol; Sigma-Aldrich; Merck KGaA) in 4 ml ethanol was refluxed for 5
h in the presence of piperidine (purity, 99%; 0.05 ml; 0.54 mmol;
Sigma-Aldrich; Merck KGaA) (28).
After cooling to room temperature in the air, water (20 ml) was
added to the reaction mixture to induce precipitation. The
precipitate was filtered and washed using water (50 ml), ethyl
acetate (20 ml) and methylene chloride (20 ml) and air-dried to
yield MHY695 (purity, >98%; 124.9 mg; 24.6%) as an amorphous
orange solid. Melting point, 207.8–210.1°C; 1H-NMR (500
MHz, DMSO-d6, δ in ppm): 13.70 (brs, 1H, NH),
10.02 (s, 1H, OH), 7.55 (s, 1H, vinylic H), 7.12 (d, 1H, J
=2.0 Hz, 2′-H), 7.06 (dd; 1H; J=2.0, 8.5 Hz; 6′-H), 6.93 (d,
1H, J=8.5 Hz, 5′-H), 4.07 (q, 2H, J=7.0 Hz,
3′-OCH2), 1.35 (t, 3 H, J=7.5 Hz,
CH2CH3); 13C-NMR (100 MHz,
DMSO-d6, δ in ppm): 196.1 (C2), 170.1 (C4), 150.9
(C3′), 147.9 (C4′), 133.5 (benzylic C), 125.8 (C6′), 125.0 (C1),
121.7 (C5), 117.1 (C5′), 116.1 (C2′), 64.6 (3′-OCH2),
15.3 (CH2CH3); LRMS (ESI-) m/z 280
(M-H)−; HRMS (ESI) m/z
C12H11NO3S2
(M-H)− calculated mass: 280.0108; observed mass:
280.0122.
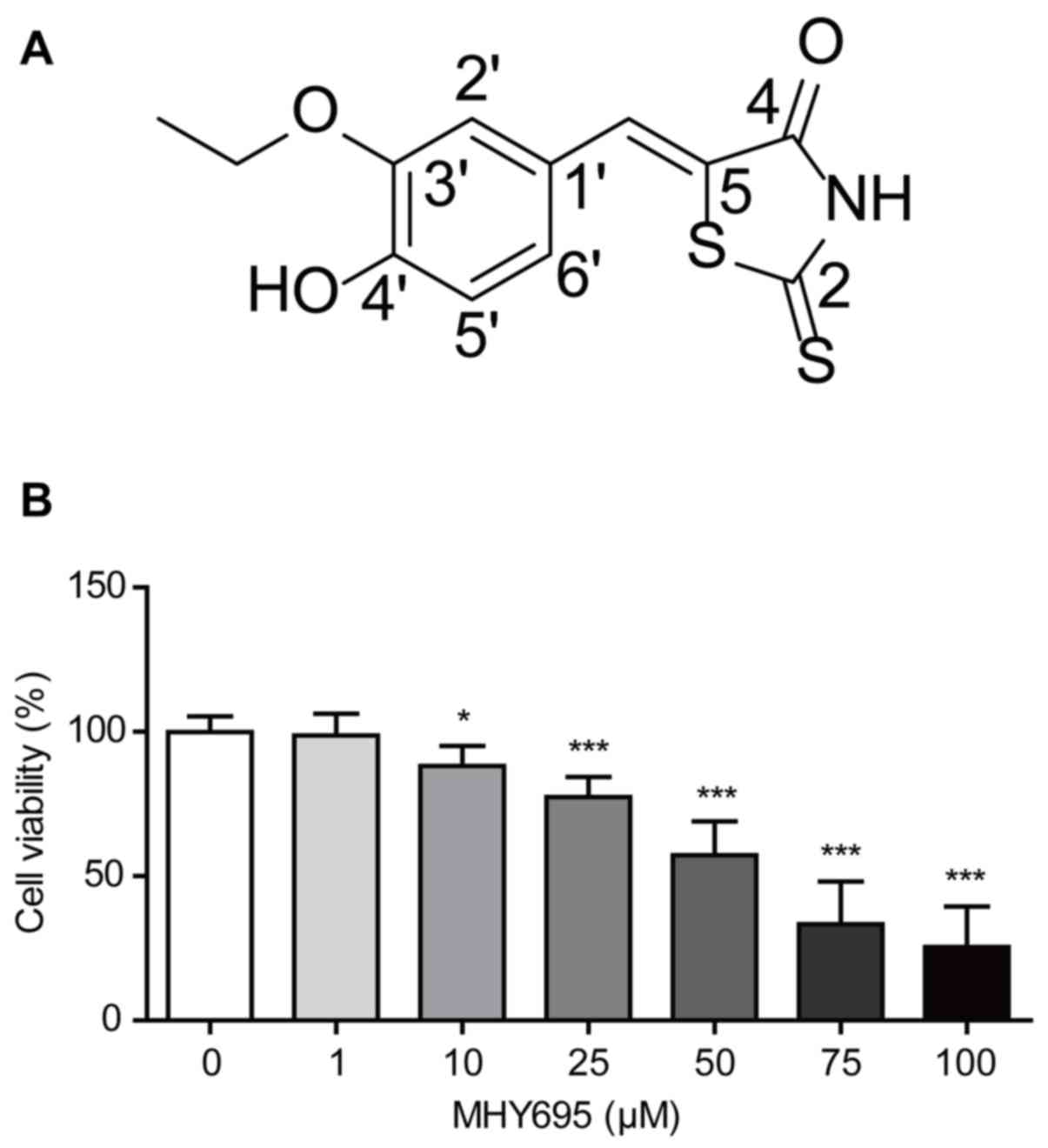
To produce a stock solution, MHY695 was dissolved in
DMSO to a concentration of 100 mM (Fig.
1A). The solution was serially diluted in RPMI-1640 or
DMEM/High glucose medium (HyClone; GE Healthcare Life Sciences) as
required; the final DMSO concentration was <0.1%.
Cell culture
NCM460, a normal human colonic epithelial cell line,
was obtained from INCELL Corporation LLC. The human colon cancer
cell line HCT116 [p53 wild-type (wt)] was obtained from the Korea
Cell Line Bank, and p53-null HCT116 cells were kindly provided by
Dr Young-Chae Chang of the Department of Cell Biology, Catholic
University of Daegu School of Medicine (Daegu, Republic of Korea).
Human colon cancer DLD-1, Caco-2 and HT-29 cells were purchased
from the American Type Tissue Culture Collection. The HCT116 and
DLD-1 cells were cultured in RPMI-1640 medium supplemented with 10%
fetal bovine serum (HyClone; GE Healthcare Life Sciences) and 1%
penicillin/streptomycin (HyClone; GE Healthcare Life Sciences) at
37°C, 5% CO2. NCM460, Caco-2 and HT-29 cells were
cultured in DMEM/High glucose medium (HyClone; GE Healthcare Life
Sciences) supplemented in the same manner as the RPMI-1640.
Cell viability assay
An MTT assay was performed to investigate the
effects of MHY695 on the viabilities of NCM460, Caco-2, DLD-1,
HT-29 and HCT116 cells. Thiazolyl blue tetrazolium bromide
(Sigma-Aldrich; Merck KGaA) was dissolved in PBS to a stock
concentration of 5 mg/ml, and further diluted with RPMI-1640 or
DMEM/High glucose medium to a working concentration of 0.5 mg/ml.
Cells were cultured in a 96-well cell culture plate at a density of
5×103 cells/well for 24 h prior to treatment. The
vehicle control (0.05% DMSO in culture medium) or MHY695 samples
were treated with 25 or 50 µM in sextuplicate for 24 h. Thiazolyl
blue tetrazolium bromide (0.5 mg/ml) was subsequently added, and
the cells were incubated in the dark for 2 h. The formazan crystals
were dissolved in DMSO and absorbance was measured at 540 nm using
a microplate spectrophotometer (Thermo Fisher Scientific, Inc.).
Each experiment was performed three times independently. The half
maximal inhibitory concentration (IC50) values were
obtained using GraphPad Prism 5 (GraphPad Software).
Cytochrome C and DAPI staining
HCT116 cells were cultured in 4-well chamber slides
at a density of 5.0×104 cells/well for 24 h, and treated
with the vehicle or MHY695 (25 or 50 µM) for 24 h at 37°C. The
cells were fixed with 10% neutral-buffered formalin solution
(Sigma-Aldrich; Merck KGaA) for 1 h at room temperature. The fixed
cells were incubated in blocking/permeabilization solution,
containing 0.1% triton-X-100 in protein block serum-free reagent
(Dako; Agilent Technologies, Inc.) for 1 h at room temperature. The
cells were subsequently incubated overnight with cytochrome C
primary antibody (1:100; cat. no. sc-7159; Santa Cruz
Biotechnology) at 4°C. A goat anti-rabbit IgG, fluorescein
isothiocyanate (FITC)-conjugated secondary antibody (1:200; cat.
no. A120-101F; Bethyl Laboratories, Inc.) was added and incubated
for a further 3 h in the dark at room temperature. Vectashield
mounting medium with DAPI (Vector Laboratories, Inc.) was used for
mounting, and the cells were washed with PBS between each step.
Fluorescence was visualized using a confocal microscope at ×480
magnification (Olympus).
FITC Annexin V staining
HCT116 cells were cultured in a 6-well culture plate
at a density of 1×106 cells/well for 24 h, and treated
with the vehicle or MHY695 (25 or 50 µM) for 24 h at 37°C. The
treated cells were collected by centrifugation at 380 × g for 3 min
at room temperature and stained using a FITC Annexin V Apoptosis
Detection Kit I (BD Biosciences) per the manufacturer's protocol.
The stained cells were counted using a flow cytometer (BD
Biosciences) within 1 h, and the data were analyzed using C Flow
Plus software (version 1.0.227.4; BD Biosciences).
Western blot analysis
HCT116 cells were cultured in a 6-well culture plate
at a density of 1×106 cells/well for 24 h, and treated
with the vehicle or MHY695 (25 or 50 µM) for a further 24 h at
37°C. The cells were harvested with cell scrapers and lysed in RIPA
assay buffer (Elpis BioMed). The total protein was quantified using
the Pierce® BCA Protein Assay Kit (Thermo Fisher
Scientific, Inc.) and separated using SDS-PAGE on a 10 or 15% gel.
The proteins were transferred to PVDF membranes (Merck KGaA) and
incubated in 5% bovine serum albumin, fraction V (cat. no. 160069;
MP Biomedicals, LLC) or 5% non-fat dry milk (BD Difco™) for 1 h.
The membranes were subsequently incubated overnight at 4°C with the
appropriate primary antibodies, diluted in blocking buffer. All of
the primary antibodies were purchased from Cell Signaling
Technology, with the exception of β-actin (Sigma-Aldrich; Merck
KGaA). The catalog numbers and the dilution factors of the primary
antibodies were as follows: p-Akt (Ser 473; 1:1,000; cat. no.
#9275), Akt (1:2,000; cat. no. #9272), p-Bad (Ser136; 1:500; cat.
no. #4366), Bad (1:1,000; cat. no. #9239), induced myeloid
leukaemia cell differentiation protein (Mcl-1; 1:1,000; cat. no.
#5453), Bid (1:1,000; cat. no. #2002), caspase-9 (1:1,000; cat. no.
#9508), cleaved caspase-9 (Asp330; 1:500; cat. no. #9501),
caspase-3 (1:1,000; cat. no. #9662), cleaved caspase-3 (1:500; cat.
no. #9661), cleaved PARP (Asp214; 1:1,000; cat. no. #9541),
Bcl-2-binding component 3 (PUMA;1:1,000; cat. no. #12450), Bcl-xL
(1:1,000; cat. no. #2764), Apoptosis regulator BAX (Bax; 1:1,000;
cat. no. #5023), Bcl-2 homologous antagonist/killer (Bak; 1:1,000;
cat. no. #12105) and β-actin (1:10,000; cat. no. #A5316). The
membranes were washed using TBS-T buffer (10 mM Tris, 150 mM NaCl,
pH 8.0, 0.2% Tween-20) after each step. The secondary antibodies
were also diluted in blocking buffer and added for 2 h at room
temperature. HRP conjugated goat anti-rabbit IgG polyclonal
antibody (1:5,000; cat. no. ADI-SAB-300-J) and HRP conjugated goat
anti-mouse IgG polyclonal antibody (1:20,000; cat. no.
ADI-SAB-100-J) were purchased from Enzo Life Sciences Inc.. The
blots were visualized using an enhanced chemiluminescence solution
(Advansta Inc.) and the ChemiDoc™ Touch Gel Imaging System (Bio-Rad
Laboratories, Inc.).
Mouse xenograft model
A total of 20 female BALB/c nude mice (four weeks of
age) were obtained from OrientBio, Inc. The mice were housed under
a 12 h light/dark cycle and fed rodent chow (Samtako Bio Korea) and
tap water ad libitum. After a 1-week acclimation period, the
mice were subcutaneously inoculated with HCT116 cells
(5×106 resuspended in 100 µl PBS) in each flank. After 1
week, vehicle solution or MHY695 (2 mg/kg diluted in PBS) was
subcutaneously injected near the inoculated tumor site once a day.
No animal exhibited signs of toxicity following the administration
of MHY695. All inoculations were performed under anesthesia with
isoflurane (Hana Pharm Co., Ltd) using the Small Animal
O2 Single Flow Anesthesia System for Rat, Mouse (LMS
Co., Ltd). The concentration of isoflurane was 3% for induction and
2% for maintenance, with 1 l/min oxygen. Inoculations were
performed when the mice didn't respond to physical stimuli when
under anesthesia. The diameter of each tumor was measured daily and
tumor volume (mm3) was calculated as (long
diameter)2 × (short diameter) × 0.5. The mice were
euthanized using carbon dioxide (CO2; with a flow rate
20% per min) and the tumor was dissected following the 12th
injection of vehicle solution or MHY695. All animal research and
protocols were reviewed and approved by the Pusan National
University Institutional Animal Care and Use Committee
(PNU-IACUC).
In vivo fluorescence imaging
GFP vectors, kindly provided by Dr Bum-Joon Park
(Department of Molecular Biology, College of Natural Science, Pusan
National University, Busan, Republic of Korea), were transfected
into HCT116 colon cancer cells using the Lipofectamine®
3000 transfection kit (Invitrogen; Thermo Fisher Scientific, Inc.)
and Opti-MEM medium (Thermo Fisher Scientific). A total of 4 female
BALB/c nude mice (four weeks of age) were obtained from OrientBio,
Inc., and acclimated for one week. The GFP-tagged HCT116 cells
(1.5×107 cells resuspended in 100 µl PBS) were
inoculated subcutaneously into the flanks of each mouse. The
vehicle or MHY695 (5 mg/kg diluted in PBS) was administered
subcutaneously once a day near each tumor. Tumor fluorescence was
observed daily using a fluorescence-labeled Organism Bioimaging
Instrument (NeoScience Co., Ltd.) After the 4th injection of
vehicle or MHY695, the mice were sacrificed using CO2
inhalation and the tumors were dissected for analysis.
Statistical analysis
Statistical analysis was conducted using GraphPad
Prism 5 (GraphPad Software, Inc.). The data were analyzed using
one-way analysis of variance followed by Tukey's test for multiple
comparisons. The Student's t-test was performed for the comparison
of two groups. The results are expressed as the mean ± standard
deviation, and P<0.05 was considered to indicate a statistically
significant difference.
Results
MHY695 decreases cell viability of
HCT116 human colon cancer cells
To investigate the effects of MHY695 on the
viability of colon cancer cells, MTT assays were performed using
the HCT116 human colon cancer cell line. HCT116 cells were treated
with various concentrations of MHY695 for 24 h, and cell viability
was determined using an MTT assay. As shown in Fig. 1B, MHY695 significantly reduced the
viability of HCT116 cells in a concentration-dependent manner. The
percentages of viable cells were 98.8, 88.2, 77.4, 57.3, 33.4 and
25.5% following treatment with 1, 10, 25, 50, 75 and 100 µM MHY695,
respectively.
MHY695 induces apoptotic cell death in
HCT116 human colon cancer cells
To determine whether MHY695-induced cytotoxicity was
the result of apoptotic cell death, nuclear morphological
alterations were observed using DAPI staining. Confocal microscopic
analysis of HCT116 cells treated with MHY695 (25 or 50 µM) for 24 h
showed cellular shrinkage, nuclear condensation and the formation
of apoptotic bodies. Moreover, cytochrome C release from the
mitochondria into the cytosol was also observed in MHY695-treated
HCT116 cells (Fig. 2A). To further
investigate MHY695-induced apoptotic cell death, flow cytometric
analysis of HCT116 cells treated with MHY695 was conducted after
Annexin V/PI staining. As shown in Fig.
2B, the number of apoptotic cells increased in a
concentration-dependent manner following MHY695 treatment. The
percentages of early and late apoptotic cells (quadrants IV and I,
respectively) were 15.8, 25.3 and 33.5% following treatment with
MHY695 at 0, 25 and 50 µM, respectively. In addition, the
percentages of MHY695-induced cytotoxicity appeared to be higher in
the MTT assays (Fig. 1B) compared
with the Annexin V/PI assays (Fig.
2B), suggesting that MHY695 may also induce cell cycle
arrest.
MHY695 modulates the expression of
apoptosis-related proteins
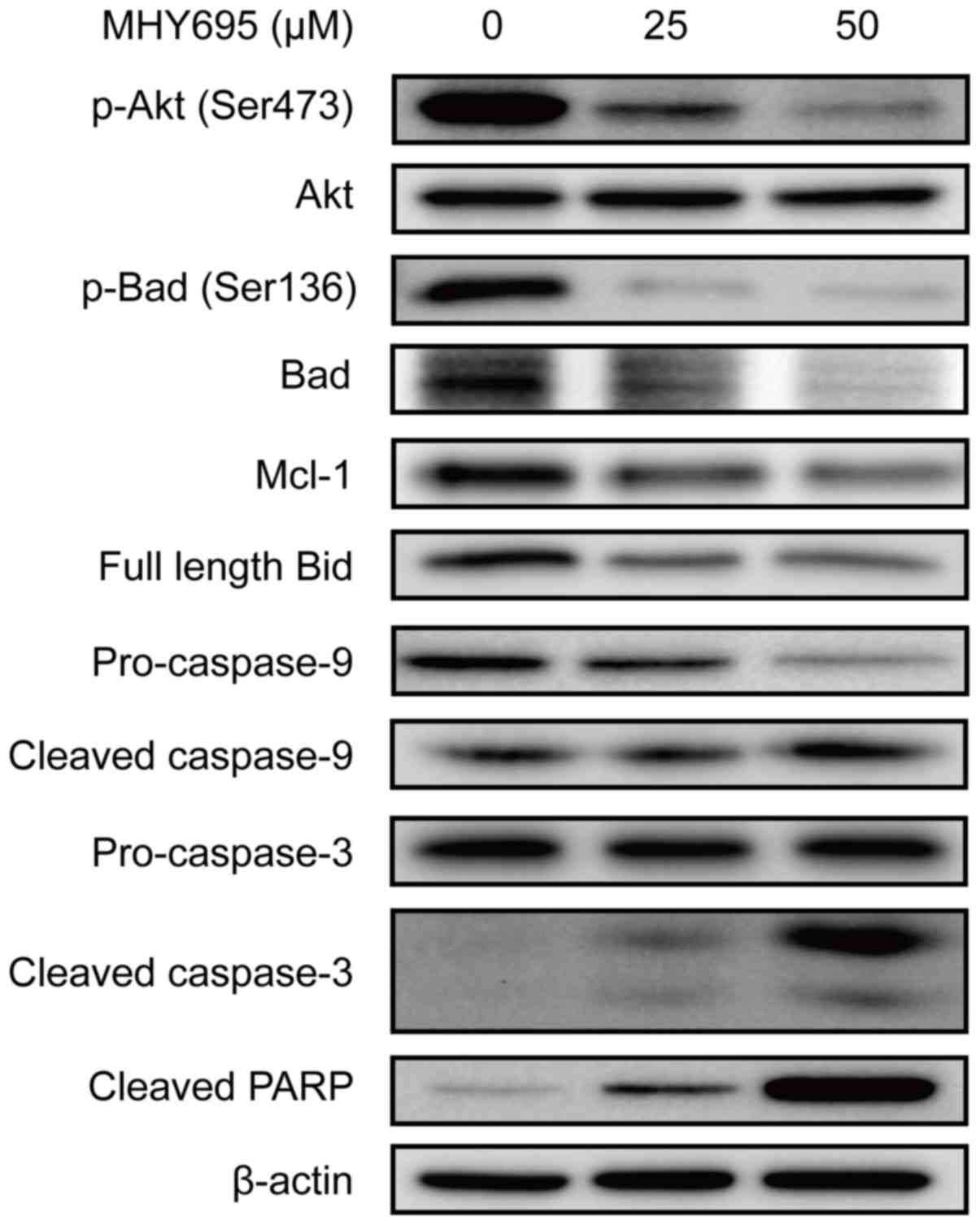
To elucidate the apoptotic signal transduction
induced by MHY695 in HCT116 cells, the expression levels of
apoptosis-associated proteins were analyzed by western blot
analysis. As shown in Fig. 3, Akt
and Bad phosphorylation was inhibited by MHY695 treatment at 25 and
50 µM for 24 h. Mcl-1 expression was also inhibited by MHY695. The
expression of full-length Bid was also decreased, suggesting that
Bid truncation was increased by MHY695. As a result, caspase-9,
caspase-3 and PARP cleavage was increased in a
concentration-dependent manner.
MHY695 suppresses tumor growth in a
mouse xenograft model
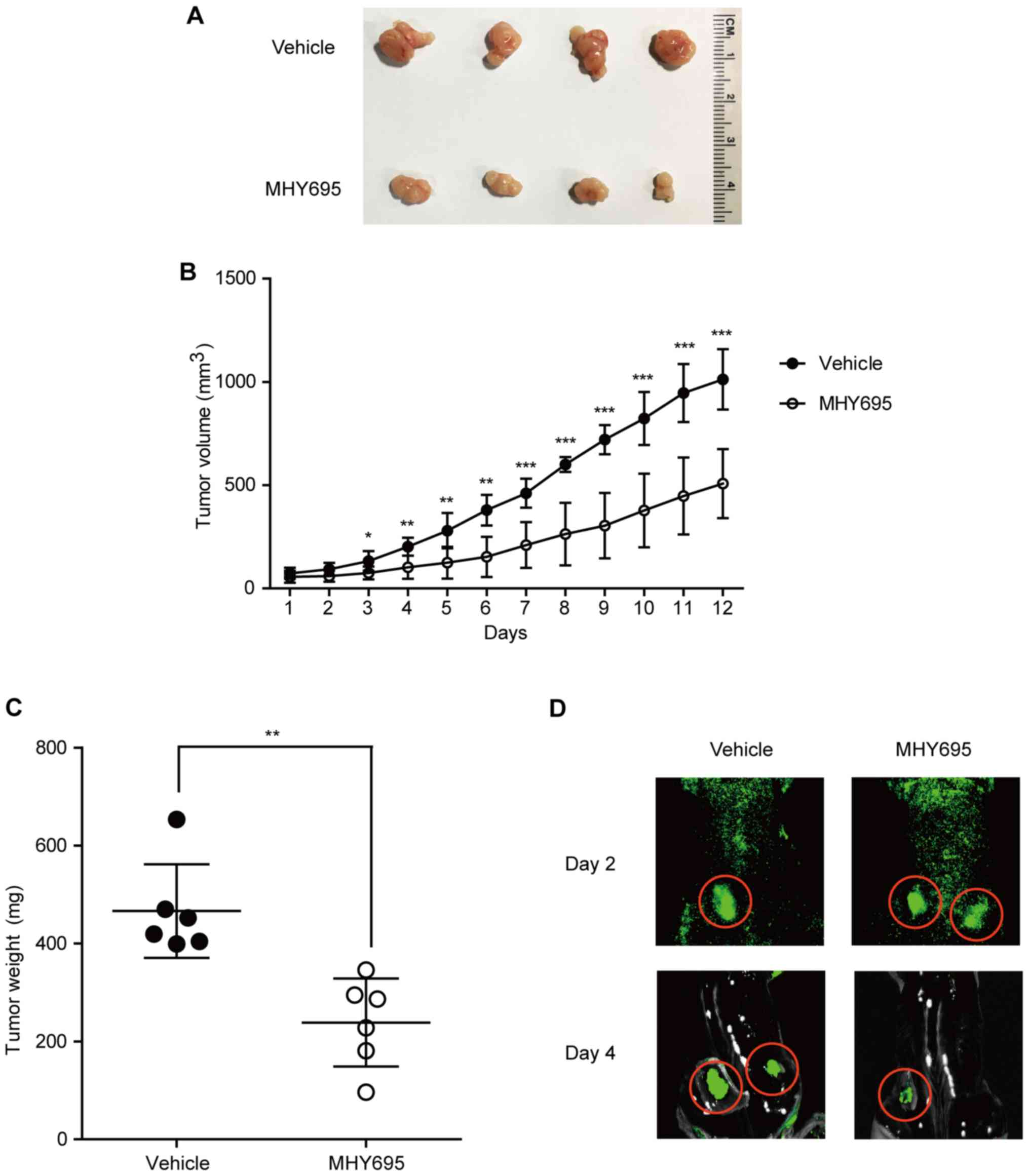
To investigate the anti-cancerous effects of MHY695
in vivo, a mouse xenograft model was employed. HCT116 cells
were implanted into the left and right flanks of BALB/c nude mice;
1 week after cancer cell inoculation, the vehicle or MHY695 (2
mg/kg) were injected subcutaneously next to each tumor on a daily
basis. As shown in Fig. 4A and B,
MHY695 significantly suppressed tumor growth compared with vehicle.
Although tumor volume was increased in both the vehicle and MHY695
groups, the growth rate of the MHY695-treated group was
significantly lower than that of the vehicle group. As illustrated
in Fig. 4C, tumor weight was also
lower in the MHY695-treated group compared with the vehicle-treated
group. Furthermore, xenograft tumors were visualized using an in
vivo fluorescence imaging system, which revealed that four days
after treatment, 5 mg/kg MHY695 resulted in reduced tumor growth
compared with the vehicle (Fig.
4D).
Cytotoxic effect of MHY695 is
independent of p53
To further investigate the cytotoxic effects of
MHY695, a normal colon epithelial cell line and additional human
colon cancer cell lines with different genetic features were
employed. MTT assays were performed in NCM460, Caco-2, DLD-1, HT-29
and HCT116 cells. Fig. 5A
illustrates that concentrations of MHY695 ≤50 µM had no cytotoxic
effect on NCM460 or HT-29 cells, while 50 µM was able to reduce the
viability of the other human colon cancer cell lines. Specifically,
MHY695 showed significant inhibitory effects towards HCT116 cells
at a concentration of 10 µM. In addition, the lowest
IC50 value among the five cell lines was observed in
HCT116 cells (Table I). Moreover,
the expression levels of apoptosis-associated proteins were
analyzed using western blotting. Fig.
5B illustrates that in DLD-1 cells, Akt phosphorylation
(Ser473) was inhibited, but PARP cleavage was increased by 25 or 50
µM MHY695 after 24 h. However, Akt phosphorylation and PARP
cleavage were not altered by MHY695 treatment in Caco-2, HT-29 and
NCM460 cells, suggesting that these effects are cell
type-specific.
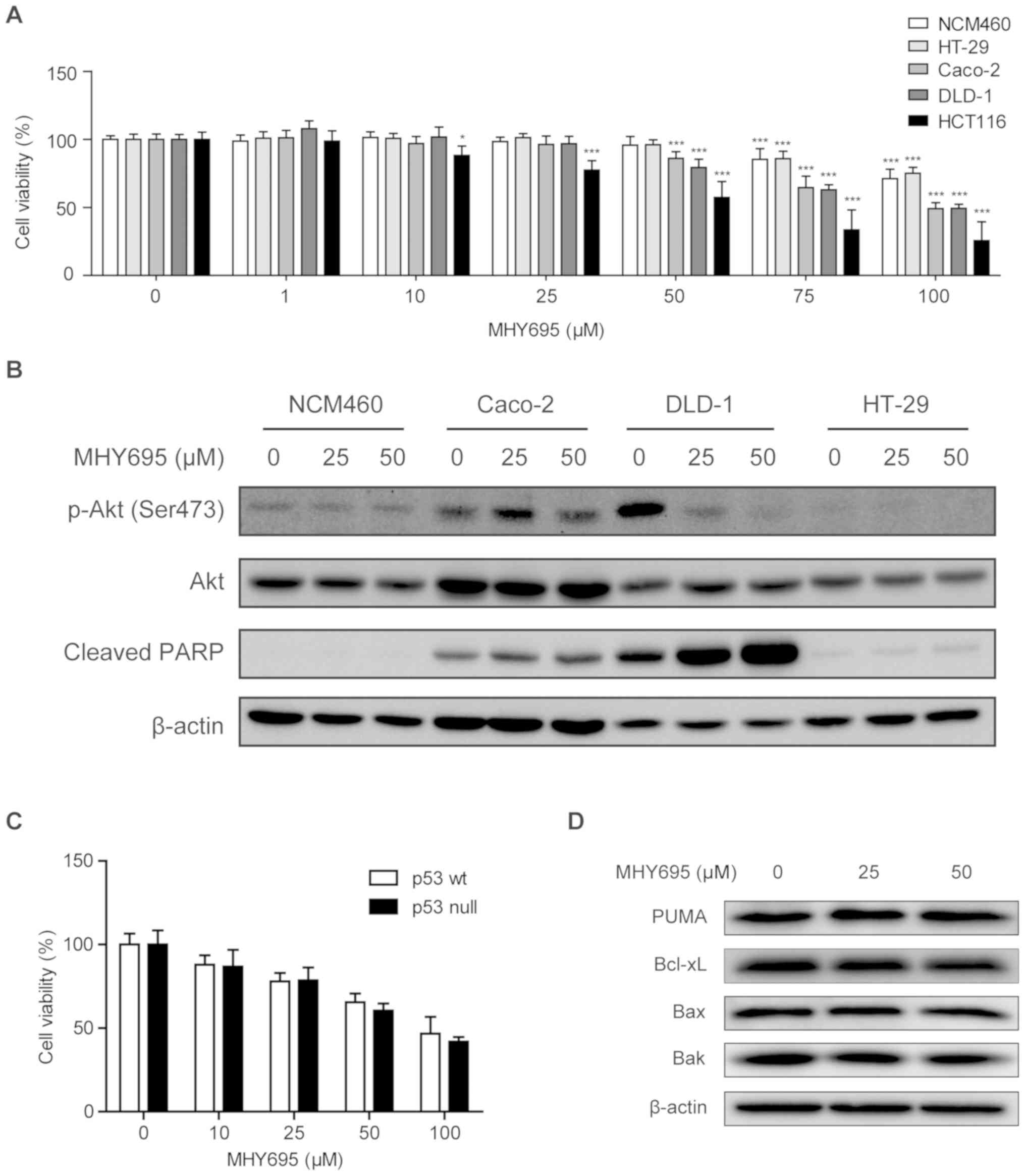 | Figure 5.Cytotoxic effects of MHY695 are
independent of p53. (A) NCM460, HT-29, Caco-2, DLD-1 and HCT116
cells were treated with various concentrations of MHY695.
*P<0.05, **P<0.01 and ***P<0.001 vs. the vehicle-treated
group. (B) Expression levels of phospho-Akt and cleaved PARP were
investigated in NCM460, Caco-2, DLD-1 and HT-29 cells by western
blot analysis. Cells were treated with vehicle or MHY695 (25 or 50
µM) for 24 h. (C) Cell viability of p53 wt and p53 null HCT116
cells were investigated by MTT assay. p53 wt or p53 null HCT116
cells were treated with vehicle or MHY695 at the specified
concentrations for 24 h. (D) Downstream proteins of the
p53-associated apoptosis pathway were investigated by western blot
analysis. p53 wt HCT116 cells were treated with vehicle or MHY695
at the specified concentrations for 24 h. MHY695,
(Z)-5-(3-ethoxy-4-hydroxybenzylidene)-2-thioxothiazolidin-4-one;
wt, wild-type; p-Akt, phospho-protein kinase B; PARP,
poly(ADP-ribose) polymerase; PUMA, Bcl-2-binding component 3; Bax,
Apoptosis regulator BAX; Bak, Bcl-2 homologous
antagonist/killer. |
 | Table I.IC50 of MHY695 in human
colon cancer cells. |
Table I.
IC50 of MHY695 in human
colon cancer cells.
| Cell line | IC50
(µM) |
|---|
| HT-29 | 268.7 |
| NCM460 | 236.8 |
| Caco-2 | 110.2 |
| DLD-1 | 104.8 |
| HCT116 | 47.9 |
It is well known that the p53 gene is mutated or
deleted in the majority of cancer cell types. Therefore, the p53
status of the cells was also determined to elucidate the different
cytotoxic effects. As shown in Table
II, the p53 gene is mutated in Caco-2, DLD-1 and HT-29 cells,
but HCT116 cells express wild-type p53 (29). Based on these findings, the cytotoxic
effects of MHY695 on p53 wt and p53 null HCT116 cells were
investigated using an MTT assay. As shown in Fig. 5C, the cytotoxic effect of MHY695 did
not differ between p53 wt and p53 null HCT116 cells. Moreover, the
expression levels of PUMA, Bcl-xL, Bax and Bak, downstream proteins
of the p53-related apoptosis pathway, were not altered by MHY695
treatment, suggesting that MHY695-mediated cytotoxicity was
independent of p53 (Fig. 5D).
| Table II.Genetic features of human colon
cancer cells. |
Table II.
Genetic features of human colon
cancer cells.
| Cell line | KRAS | BRAF | PIK3CA | PTEN | TP53 |
|---|
| HT-29 | wt | V600E | P449T | wt | R273H |
| Caco-2 | wt | wt | wt | wt | E204X |
| DLD-1 | G13D | wt | E545K, D549N | wt | S241H |
| HCT116 | G13D | wt | H1047R | wt | wt |
Discussion
In the present study, MHY695 was demonstrated to
induce cytotoxicity and apoptosis in HCT116 colon cancer cells.
When used to treat various colon cancer cell lines, including
Caco-2, DLD-1, HT-29 and HCT116 cells, MHY695 showed the greatest
potency towards HCT116 cells. In an attempt to establish the reason
for this finding, the mutation status of the p53 gene was
investigated. The tumor suppressor protein p53 is a transcription
factor that serves a pivotal role in apoptosis and cell cycle
arrest. When p53 is mutated, cancer cells exhibit resistance to
chemotherapy. According to a previous study, p53 is mutated in
Caco-2 (E204X), DLD-1 (S241F) and HT-29 (R273H) cells, but not in
HCT116 cells (29). Thus, the
effects of MHY695 were compared between p53 wt and null HCT116
cells. MHY695 showed equivalent cytotoxicity in both cell types.
Also, PUMA, Bcl-xL, Bax and Bak, which are downstream proteins of
p53, were not affected by MHY695 (30,31).
Collectively, these findings suggested that p53 was not associated
with the cytotoxic effect of MHY695. The reasoning for the
selectivity of MHY695 towards different types of cancer cell
remains a future challenge.
When the results of MTT assays (Fig. 1B) were compared with those of Annexin
V/PI staining (Fig. 2B), MHY695
appeared to exhibit lesser cytotoxicity during Annexin V/PI
analysis. This result suggests that MHY695 may also promote cell
cycle inhibition in HCT116 cells. Therefore, future studies using
FACS analysis and/or immunoblot analysis of cell cycle proteins are
warranted to determine whether MHY695 induces cell cycle arrest in
other colon cancer cell lines.
The expression levels of numerous
apoptosis-associated proteins were investigated using western blot
analysis. Among them, Bax and Bak, which oligomerize in the
mitochondrial membrane and form pores to facilitate the release of
cytochrome C, were not affected by MHY695 treatment. Generally
speaking, pro-apoptotic proteins such as Bax and Bak are
upregulated in apoptotic cells (32). However, MHY695 significantly
inhibited the phosphorylation of Akt and Bad. Also, caspase-3 and
PARP cleavage, reliable markers of apoptosis, were markedly
apparent. According to Datta et al (8) reduced phosphorylation of Akt precedes
Bad-induced cell death, hence it was concluded that MHY695 may
induce apoptosis by influencing the phosphoinositide 3-kinase
(PI3K)/Akt signaling pathway. Furthermore, as the PI3K/Akt pathway
is primarily activated by growth factors, such as epidermal growth
factor and fibroblast growth factor, it was suggested that MHY695
may target those growth factor receptors, the specifics of which
present a future research objective (33,34).
In addition to in vitro studies, the in
vivo anti-cancer effects of MHY695 we investigated using a
mouse xenograft model. Subcutaneous injection of MHY695 around the
tumor significantly inhibited tumor growth compared with the
vehicle. However, intraperitoneal injection of MHY695 did not
inhibit the growth of tumors located in the flanks (data not
shown). Therefore, the route of administration and optimal in
vivo dose may be another focus of future research. Taken
together, these findings suggested that MHY695 may constitute a
candidate for the development of anti-cancer drugs against colon
cancer.
Acknowledgements
Not applicable.
Funding
This work was supported by a 2-year research grant
from Pusan National University (EI).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GH conducted and analyzed the in vitro cell
based and animal experiments, and wrote the manuscript. DK, CP and
HRM synthesized the chemical reagents. SJK, JC and YL conducted
in vitro experiments. JWY, YJ, JL, NDK and HYC substantially
contributed to the design of the study, interpreted the data, and
wrote the manuscript. HRM and EI analyzed the data, conceived,
supervised and directed the study, and wrote the manuscript. All
the authors approved the final version of this manuscript and
agreed to be accountable for all aspects of the work in ensuring
that questions related to the accuracy or integrity of any part of
the work are appropriately investigated and revised.
Ethics approval and consent to
participate
All animal research and protocols were reviewed and
approved by the Pusan National University Institutional Animal Care
and Use Committee (PNU-IACUC).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jung KW, Won YJ, Oh CM, Kong HJ, Lee DH,
Lee KH and Community of Population-Based Regional Cancer
Registries: Cancer Statistics in Korea: Incidence, mortality,
survival, and prevalence in 2014. Cancer Res Treat. 49:292–305.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Benson AB III, Venook AP, Cederquist L,
Chan E, Chen YJ, Cooper HS, Deming D, Engstrom PF, Enzinger PC,
Fichera A, Grem JL, et al: Colon cancer, version 1.2017, NCCN
clinical practice guidelines in oncology. J Natl Compr Canc Netw.
15:370–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Levin S, Bucci TJ, Cohen SM, Fix AS,
Hardisty JF, LeGrand EK, Maronpot RR and Trump BF: The nomenclature
of cell death: Recommendations of an ad hoc Committee of the
society of toxicologic pathologists. Toxicol Pathol. 27:484–490.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li H, Zhu H, Xu CJ and Yuan J: Cleavage of
BID by caspase-8 mediates the mitochondrial damage in the Fas
pathway of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Datta SR, Dudek H, Tao X, Masters S, Fu H,
Gotoh Y and Greenberg ME: Akt phosphorylation of BAD couples
survival signals to the cell-intrinsic death machinery. Cell.
91:231–241. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai J, Yang J and Jones DP: Mitochondrial
control of apoptosis: the role of cytochrome c. Biochim Biophys
Acta. 1366:139–149. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Slee EA, Adrain C and Martin SJ:
Executioner caspase-3, −6, and −7 perform distinct, non-redundant
roles during the demolition phase of apoptosis. J Biol Chem.
276:7320–7326. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choi PR, Kang YJ, Sung B, Kim JH, Moon HR,
Chung HY, Kim SE, Park MI, Park SJ and Kim ND: MHY218-induced
apoptotic cell death is enhanced by the inhibition of autophagy in
AGS human gastric cancer cells. Int J Oncol. 47:563–572. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeon HS, Ahn MY, Park JH, Kim TH, Chun P,
Kim WH, Kim J, Moon HR, Jung JH and Kim HS: Anticancer effects of
the MHY218 novel hydroxamic acid-derived histone deacetylase
inhibitor in human ovarian cancer cells. Int J Oncol. 37:419–428.
2010.PubMed/NCBI
|
|
13
|
Kim MK, Kang YJ, Kim DH, Hossain MA, Jang
JY, Lee SH, Yoon JH, Chun P, Moon HR, Kim HS, et al: A novel
hydroxamic acid derivative, MHY218, induces apoptosis and cell
cycle arrest through downregulation of NF-κB in HCT116 human colon
cancer cells. Int J Oncol. 44:256–264. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park JH, Ahn MY, Kim TH, Yoon S, Kang KW,
Lee J, Moon HR, Jung JH, Chung HY and Kim HS: A new synthetic HDAC
inhibitor, MHY218, induces apoptosis or autophagy-related cell
death in tamoxifen-resistant MCF-7 breast cancer cells. Invest New
Drugs. 30:1887–1898. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
De U, Kundu S, Patra N, Ahn MY, Ahn JH,
Son JY, Yoon JH, Moon HR, Lee BM and Kim HS: A new histone
deacetylase inhibitor, MHY219, inhibits the migration of human
prostate cancer cells via HDAC1. Biomol Ther (Seoul). 23:434–441.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Patra N, De U, Kim TH, Lee YJ, Ahn MY, Kim
ND, Yoon JH, Choi WS, Moon HR, Lee BM and Kim HS: A novel histone
deacetylase (HDAC) inhibitor MHY219 induces apoptosis via
up-regulation of androgen receptor expression in human prostate
cancer cells. Biomed Pharmacother. 67:407–415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee SH, Kang YJ, Kim DH, Sung B, Kang JA,
Chun P, Yoon JH, Moon HR, Kim HS, Chung HY and Kim ND: A novel
oxiranylchromenone derivative, MHY336, induces apoptosis and cell
cycle arrest via a p53- and p21-dependent pathway in HCT116 human
colon cancer cells. Int J Oncol. 44:943–949. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Patra N, De U, Kang JA, Kim JM, Ahn MY,
Lee J, Jung JH, Chung HY, Moon HR and Kim HS: A novel epoxypropoxy
flavonoid derivative and topoisomerase II inhibitor, MHY336,
induces apoptosis in prostate cancer cells. Eur J Pharmacol.
658:98–107. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yoon S, Kim JH, Lee YJ, Ahn MY, Choi G,
Kim WK, Yang Z, Lee HJ, Moon HR and Kim HS: A novel carbazole
derivative, MHY407, sensitizes cancer cells to doxorubicin-,
etoposide-, and radiation treatment via DNA damage. Eur J
Pharmacol. 697:24–31. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
De U, Chun P, Choi WS, Lee BM, Kim ND,
Moon HR, Jung JH and Kim HS: A novel anthracene derivative, MHY412,
induces apoptosis in doxorubicin-resistant MCF-7/Adr human breast
cancer cells through cell cycle arrest and downregulation of
P-glycoprotein expression. Int J Oncol. 44:167–176. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hwang HJ, Kang YJ, Hossain MA, Kim DH,
Jang JY, Lee SH, Yoon JH, Moon HR, Kim HS, Chung HY and Kim ND:
Novel dihydrobenzofuro[4,5-b][1,8]naphthyridin-6-one derivative,
MHY-449, induces apoptosis and cell cycle arrest in HCT116 human
colon cancer cells. Int J Oncol. 41:2057–2064. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim SH, Kang YJ, Sung B, Kim DH, Lim HS,
Kim HR, Kim SJ, Yoon JH, Moon HR, Chung HY and Kim ND: MHY-449, a
novel dihydrobenzofuro[4,5-b][1,8]naphthyridin-6-one derivative,
mediates oxidative stress-induced apoptosis in AGS human gastric
cancer cells. Oncol Rep. 34:288–294. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee SH, Kang YJ, Sung B, Kim DH, Lim HS,
Kim HR, Kim SJ, Yoon JH, Moon HR, Chung HY and Kim ND: MHY-449, a
novel dihydrobenzofuro[4,5-b][1,8] naphthyridin-6-one derivative,
induces apoptotic cell death through modulation of Akt/FoxO1 and
ERK signaling in PC3 human prostate cancer cells. Int J Oncol.
44:905–911. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lim HS, Kang YJ, Sung B, Kim SH, Kim MJ,
Kim HR, Kim SJ, Choi YH, Moon HR, Chung HY and Kim ND: Novel
dihydrobenzofuro[4,5-b][1,8]naphthyridin-6-one derivative, MHY-449,
induces cell cycle arrest and apoptosis via the downregulation of
Akt in human lung cancer cells. Oncol Rep. 34:2431–2438. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jada SR, Matthews C, Saad MS, Hamzah AS,
Lajis NH, Stevens MF and Stanslas J: Benzylidene derivatives of
andrographolide inhibit growth of breast and colon cancer cells in
vitro by inducing G(1) arrest and apoptosis. Br J Pharmacol.
155:641–654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zuliani V, Carmi C, Rivara M, Fantini M,
Lodola A, Vacondio F, Bordi F, Plazzi PV, Cavazzoni A, Galetti M,
et al: 5-Benzylidene-hydantoins: Synthesis and antiproliferative
activity on A549 lung cancer cell line. Eur J Med Chem.
44:3471–3479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang X, Wang W, Qin JJ, Wang MH, Sharma H,
Buolamwini JK, Wang H and Zhang R: JKA97, a novel benzylidene
analog of harmine, exerts anti-cancer effects by inducing G1
arrest, apoptosis, and p53-independent up-regulation of p21. PLoS
One. 7:e343032012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pinson JA, Schmidt-Kittler O, Zhu J,
Jennings IG, Kinzler KW, Vogelstein B, Chalmers DK and Thompson PE:
Thiazolidinedione-based PI3Kα inhibitors: An analysis of
biochemical and virtual screening methods. Chem Med Chem.
6:514–522. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ahmed D, Eide PW, Eilertsen IA, Danielsen
SA, Eknæs M, Hektoen M, Lind GE and Lothe RA: Epigenetic and
genetic features of 24 colon cancer cell lines. Oncogenesis.
2:e712013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakano K and Vousden KH: PUMA, a novel
proapoptotic gene, is induced by p53. Mol Cell. 7:683–694. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ashkenazi A: Directing cancer cells to
self-destruct with pro-apoptotic receptor agonists. Nat Rev Drug
Discov. 7:1001–1012. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wei MC, Zong WX, Cheng EH, Lindsten T,
Panoutsakopoulou V, Ross AJ, Roth KA, MacGregor GR, Thompson CB and
Korsmeyer SJ: Proapoptotic BAX and BAK: A requisite gateway to
mitochondrial dysfunction and death. Science. 292:727–730. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, McCullough KD, Franke TF and
Holbrook NJ: Epidermal growth factor receptor-dependent Akt
activation by oxidative stress enhances cell survival. J Biol Chem.
275:14624–14631. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu Y, Lu H and Zhang J, Chen J, Chai Z and
Zhang J: Essential role of AKT in tumor cells addicted to FGFR.
Anticancer Drugs. 25:183–188. 2014. View Article : Google Scholar : PubMed/NCBI
|