Introduction
Breast cancer is the second most common cancer
worldwide and the most frequent cancer among women (1). One in 8 women in Europe will develop
breast cancer before the age of 85 (2). The highest prevalence is found in
northern and western European countries, suggesting a relation to
environmental factors (3,4). Metastatic breast cancer is the second
leading cause of cancer-related fatality in women: It has a
five-year relative survival rate of 23%, compared with 99% for
non-metastatic breast cancer (5).
Breast tumours are classified according to the Elston-Ellis
modification of the Scarff-Bloom-Richardson (SBR) grading system
(also known as the Nottingham grading system) (6). This system grades tumours according to
their differentiation, from well differentiated (grade 1), to
moderately differentiated (grade 2) or poorly differentiated (grade
3). This histological scale is used as a prognostic predictor in
patients with breast cancer, as tumour grade has a positive
correlation with metastasis and risk of recurrence.
Regulation of signal transduction and stress
response is critical to maintain cellular homeostasis. Cell signals
in response to stress can result in growth arrest and elimination
of damaged or premalignant cells. During cancer development,
signalling pathways are often impaired due to dysregulation of gene
expression or aberrant signal transduction, resulting in the
hallmarks of cancer (7–13). Gene expression is largely regulated
by epigenetic changes, including DNA methylation and histone
modification (14). More than 50% of
human cancers harbour mutations in enzymes involved in chromatin
organization (15). One of the
biggest challenges in cancer research is to understand how defects
arise during disease progression. This is likely to become
increasingly important to detect and validate biomarkers for tumour
gradings and subtypes that may help guide treatment decisions
(16).
One gene that has recently been found to be involved
in breast and ovarian cancer and lymph node metastasis in cervical
cancer is the Kelch domain-containing protein 7B (KLHDC7B) gene
(13,17–19). It
has been identified as hypermethylated and upregulated in breast
cancer (17), and, when associated
with alternative splicing events, may be involved in the
development and progression of cervical squamous cell carcinoma
(CSCC) (20).
The KLHDC7B gene (Hs.137007) (21) comprises a single exon, located on
human chromosome 22q13.33 (22,23). In
50 paired samples of breast cancer tissue and adjacent normal
tissue, the methylation level of the 14 CpG sites at the promoter
region of the gene was higher in cancerous tissue (72–93%) than in
normal tissue (31–83%) (17). A
clear relationship between high methylation levels and upregulated
expression was also observed in cultured breast cell lines. For
instance, MCF-7 (90–100%) and MDA-MB-468 (100%) cancer cell lines
had higher methylation at the 14 CpGs and higher gene expression
than BT549 (20–90%) and 184B5 (10–100%) cell lines (17).
Numerous reports have described an association
between hypermethylation of individual genes and clinical prognosis
for various types of cancer, and individual methylation markers
have previously been linked to breast cancer metastasis (24). DNA methylation is generally
associated with gene downregulation. However, some genes, including
survivin (25), the glycoprotein
hormone alpha-subunit (26), and
KLHDC7B, have been found to be upregulated when CpG sites are
hypermethylated (17).
The potential reversibility of epigenetic status
offers exciting opportunities for cancer treatments, and targeting
methylation represents the third wave of anticancer drug
development (24). DNA
methyltransferases currently represent one of the major drug
targets, and new drugs are expected to be added in the near future
(24,27,28).
The KLHDC7B gene encodes a 594-amino-acid protein
product that contains a Kelch domain in the C-terminal half
(29). The Kelch domain is a common
motif that forms a 4-stranded anti-parallel β-propeller.
Kelch-repeat β-propellers interact with a variety of other proteins
(30,31). Besides the presence of the Kelch
domain and a verified expressed sequence tag (EST), no other
information is available to determine the function of KLHDC7B
(22). Kelch motif-containing
proteins are involved in diverse biological processes, such as
signal transduction, building cell structures, regulating
transcription, metabolism and, notably, in stress responses
(22,32,33).
Mutations of Kelch proteins have been associated with cancer:
Examples include KLHL6 in lymphocytic leukaemia, KEAP1 in pulmonary
papillary adenocarcinoma, KLHL20 in prostate cancer, and KLHL37
(ENC1) in brain tumours (34). A
recent study showed that apoptosis of MCF-7 decreased and
proliferation increased when KLHDC7B was upregulated, and when
KLHDC7B was downregulated, the opposite occurred, indicating its
oncogenic properties (19). However,
the encoded protein and its role in these cancers remain largely
unknown.
Materials and methods
Tumour samples
Breast cancer specimens (n=26) and adjacent healthy
tissue specimens (n=17) were obtained from female patients with
breast cancer at Vall d'Hebron Hospital (Barcelona, Spain). The
study was approved by the Clinical Research Ethics Committee at
Vall d'Hebron Hospital [PR(AG)309/2016], and written informed
consent was obtained from patients prior to sample collection.
Tissues were extracted during 2009 and mRNA was extracted from 2015
to 2017. The selection criteria allowed different tumour types
(papillary, ductal, lobular, mucinous, tubular and ductal) and
grades, including metastatic and non-metastatic tumours.
Tumours were classified according to the
Elston-Ellis modification of the Scarff-Bloom-Richardson (SBR)
grading system (Nottingham grading system) (6) as well differentiated (grade 1, n=5),
moderately differentiated (grade 2, n=10) or poorly differentiated
(grade 3, n=11).
Histology and
immunohistochemistry
Immunohistochemical staining were performed on
five-micron-thick sections from formalin fixed and paraffin
embedded (FFPE) tissues, on the Ventana Benchmark XT Automated IHC
Stainer, using the Ventana ultraView Universal DAB Detection kit
(760–500). After deparaffinization with Ventana EZ Prep solution
(950–102), antigen retrieval was performed using Ventana Tris-based
buffer solution CC1 pH 8 (950–124). Endogenous peroxidase was
blocked with 3% hydrogen peroxide. After rinsing using Reaction
Buffer (950–300), slides were incubated at 37°C with each primary
antibody (Ventana Medical Systems Inc, Tucson, AZ, USA; EEUU):
Ki67-20 min (rabbit monoclonal antibody, 790-4286), p53-44 min
(mouse monoclonal, 800-2912), HER2/neu-28 min (rabbit monoclonal,
790-2991), ER-40 min (rabbit monoclonal, 790-4324) and PR-16 min
(rabbit monoclonal, 790-2223). Following incubation with HRP
Multimer secondary antibody, primary antibodies-horseradish
peroxidase-labelled antibody complex were visualized using
diaminobenzidine tetrahydrochloride chromogen. Slides were then
counterstained for 8 min with haematoxylin (760–2021), for 4 min
with bluing reagent (760–2037), dehydrated and mounted. Appropriate
positive and negative controls were included within the study
sections.
Cell culture and reagents
MCF-10A, MCF-7, MDA-MB-231 and MDA-MB-468 breast
cancer cell lines were purchased from the American Type Culture
Collection (ATCC, Manassas, VA, USA) and authenticated by DNA
profiling using short tandem repeat (STR) (GenePrint® 10
System, Promega, Fitchburg, WI, USA) at Genomics Core Facility,
Instituto de Investigaciones Biomédicas ‘Alberto Sols’ CSIC-UAM
(Madrid, Spain) (35). Mycoplasma
PCR analysis detected no genetic material. Cells were maintained at
37°C in a 5% CO2 humidified incubator (AutoFlow UN-5510,
Nuaire, Plymouth, MN, USA). MCF-7, MDA-MB-231 and MDA-MB-468 cells
were cultured in Dulbecco's modified Eagle's medium (DMEM) (Life
Technologies, Carlsbad, CA, USA) supplemented with 10%
heat-inactivated fetal bovine serum (FBS; Biowest, Nuaillé, France)
and antibiotics (penicillin, streptomycin; Gibco-ThermoFisher
Scientific, Waltham, MA, USA). MCF-10A medium was additionally
supplemented with 20 ng/ml EGF (cat. no: E9644; Sigma, St. Louis,
MO, USA), 0.5 µg/ml hydrocortisone, 100 ng/ml cholera toxin (cat.
no: C9903; Sigma) and 10 µg/ml insulin (cat. no: I9278; Sigma).
Cells were trypsinized and passaged using TrypLE reagent
(ThermoFisher Scientific).
MCF-10A is a non-tumour breast cell line, which is
hormone-receptor [oestrogen-receptor (ER) and progesterone-receptor
(PR)] negative, HER2 negative and p53 wildtype (36–38)
(Table I). The three breast cancer
cell lines derive from breast adenocarcinomas (Table I). MCF-7 is classified as luminal A
molecular subtype, hormone-receptor positive, HER2 negative and p53
wildtype. Luminal A cancers are low-grade, tend to grow slowly and
have the best prognosis. MDA-MB-231 and MDA-MB-468 are
triple-negative/basal-like breast cancer cell lines,
hormone-receptor negative, HER2 negative and p53 mutated. Although
both the MDA-MB cell lines are triple-negative, they show
significant differences: MDA-MB-468 is classified as type A,
showing a core basal-like morphology, and MDA-MB-231 is classified
as type B, being the least differentiated, highly invasive and
having the worst prognosis (39,40).
 | Table I.Cell line characterization: Oestrogen
receptor, progesterone receptor and p53 status, tumour subtype,
origin and morphology. |
Table I.
Cell line characterization: Oestrogen
receptor, progesterone receptor and p53 status, tumour subtype,
origin and morphology.
| Cell line | ER | PR | HER2 | p53 | Subtype | Origin | Morphology |
|---|
| MCF-10A | − | − | − | Wild type | Non-tumour | Fibrocystic
disease | Epithelial |
| MCF-7 | + | + | − | Wild type | Luminal A | Adenocarcinoma | Most
differentiated; tight cell-cell junctions |
| MDA-MB-468 | − | − | − | Mutated | Triple negative
A | Adenocarcinoma | Core
basal-like |
| MDA-MB-231 | − | − | − | Mutated | Triple negative
B | Adenocarcinoma | Least
differentiated and highly invasive |
RNA extraction and quantification
Tissue samples were lysed using Tissue Lyser II
(Qiagen, Venlo, the Netherlands) and RNA was extracted by the
L'Hospital Universitari Vall d'Hebron Biobank (HUVH Biobank,
Barcelona, Spain), using QuickGene RNA tissue SII kit (RT-S2)
(Fujifilm, Neuss, Germany) in the automated nucleic acid extraction
system QuickGene 810 (Fujifilm), according to the manufacturer's
instructions. RNA from culture cell lines was extracted using the
PureLink™ RNA Mini Kit (ThermoFisher Scientific). Quantification
and assessment of RNA purity was performed using a NanoDrop ND2000
Spectrophotometer (ThermoFisher Scientific) and confirmed according
to the RIN (RNA integrity number) using an Agilent Bioanalyzer
(Agilent Technologies, Santa Clara, CA, USA).
RT-PCR
One microgram of total RNA was used to synthesize
cDNA using Maxima Reverse Transcriptase (ThermoFisher Scientific)
on a Veriti 96-well Thermal Cycler (Applied Biosystems,
ThermoFisher Scientific). RT-qPCR was performed according to the
manufacturer's instructions, on an Applied Biosystems ABI 7500 Fast
Real Time-PCR sequence detection system, using Taqman Technology
(ThermoFisher Scientific): TaqMan GeX Master Mix (4369016), KLHDC7B
probe (Hs00536653_s1) and HPOL probe (Hs00172187_m1) as
housekeeping. RNA from healthy tissue was used as a normalisation
control: FirstChoice® Human Breast Total RNA (AM6952,
AppliedBiosystems-Ambion-Thermo Fisher). Analysis of relative gene
expression data was conducted using RT-qPCR and the
2−ΔΔCq method (41).
The average KLHDC7B expression for grade 1 (G1)
tumours (n=5) was calculated, and mRNA expression of every tumour
was reported as relative to this average (value=1). The two G1
tumours with the highest KLHDC7B expression were used as cut-offs
for low expression (G1 tumours being those with the lowest KLHDC7B
expression overall) (Table II).
| Table II.Tumour features. |
Table II.
Tumour features.
| Patient ID | Tumour type (breast
carcinoma) | Grade | ER | PR | HER | p53 | Ki67 (% positive
cells) | Vascular
invasion | Metastases | Relative mRNA
expression (to G1 average=1) |
|---|
| B09-13526 | Papillary | 2 | +++ | +++ | − | − | 5–10 | − | No | 0.18 |
| B09-3889 | Ductal | 2 | ? | ? | ? | ? | ? | − | No | 0.43 |
| B09-24083 | Lobular | 1 | +++ | +++ | − | − | <5 | − | Yes | 0.58 |
| B09-16306 | Mucinous | 1 | +++ | +++ | − | − | <1 | − | No | 0.60 |
| B09-12616 |
Cribriform/tubular | 1 | +++ | +++ | − | ? | ? | − | No | 0.77 |
| B09-5308 | Ductal | 3 | ? | ? | ? | ? | ? | − | No | 0.77 |
| B09-18339 | Lobular | 2 | +++ | + | − | − | 15 | − | No | 0.87 |
| B09-11839 | Ductal | 2 | +++ | − | − | − | 5 | − | Yes | 0.91 |
| B09-18841 | Ductal | 3 | +++ | +++ | − | − | 5 | − | Yes | 1.03 |
| B09-16005 | Ductal | 3 | +++ | +++ | − | + | 60 | + | Yes | 1.09 |
| B09-988 | Lobular | 2 | ? | ? | ? | ? | ? | ? | Yes | 1.21 |
| B09-25265 | Lobular | 2 | +++ | +++ | − | − | 2 | − | Yes | 1.21 |
| B09-19643 | Lobular | 2 | ? | ? | ? | ? | ? | ? | No | 1.34 |
|
B09-24926a | Ductal | 1 | +++ | +++ | − | − | <5 | + | Yes | 1.48 |
| B09-24052 | Ductal | 3 | +++ | +++ | − | − | 20 | − | No | 1.52 |
| B09-22493 | Lobular | 2 | +++ | +++ | − | − | <5 | − | No | 1.63 |
|
B09-24372a | Ductal | 1 | +++ | +++ | − | − | 1 | − | No | 1.64 |
| B09-8511 | Ductal | 2 | +++ | +++ | − | − | 15 | − | Yes | 1.69 |
| B09-7717 | Ductal | 3 | +++ | ++ | − | + | 70 | − | ? | 1.79 |
| B09-11451 | Papillary | 3 | +++ | ++ | − | ? | ? | − | ? | 1.80 |
| B09-3173 | Ductal | 3 | − | − | − | + | >70 | + | Yes | 1.85 |
| B09-26055 | Ductal | 3 | +++ | + | − | − | 40 | − | Yes | 1.98 |
| B09-1573 | Tubular | 2 | ? | ? | ? | ? | ? | ? | Yes | 2.11 |
| B09-20004 | Ductal | 2 | +++ | +++ | − | − | 3 | − | No | 2.16 |
| B09-15644 | Ductal | 2 | +++ | ++ | − | − | 15–20 | +/- | Yes | 2.26 |
| B09-20128 |
Papillary/Ductal | 3 | +++ | +++ | − | − | 10 | − | No | 2.43 |
| B09-20045 | Ductal | 3 | − | − | − | + | 60 | − | No | 3.02 |
| B09-25449 | Ductal | 2 | ++ | +++ | − | − | 25 | + | Yes | 3.70 |
| B09-2726 | Ductal | 3 | − | − | − | + | 50 | + | Yes | 3.98 |
| B09-19151 | Ductal | 3 | +++ | +++ | − | + | 1 | − | No | 6.98 |
| B09-24264 | Ductal | 2 | +++ | +++ | − | − | <5 | − | No | 23.44 |
| B09-17267 | Ductal | 3 | ++ | + | + | + | 15–20 | + | No | 272.58 |
Comparison between healthy and tumour tissue was
performed in cases in which there was enough mRNA from healthy
tissue to perform retrotranscription from mRNA to cDNA.
Protein extraction and
immunoblotting
Total protein extracts were generated using a RIPA
Lysis Buffer System (sc24948; SantaCruz Biotechnology, Dallas, TX,
USA) supplemented with Protease Inhibitor Cocktail set III (539134;
Calbiochem, San Diego, CA, USA) and Phosphatase Inhibitor Cocktail
Set II (524625, Calbiochem). Protein was quantified using a BCA
Protein Assay kit (23225; ThermoFisher Scientific). Protein
extracts (15 µg per sample) were loaded onto SDS-PAGE gels and
electrophoretically transferred to polyvinylidene fluoride (PVDF)
membranes. Membranes were blocked with 5% BSA in TBS-T
(Tris-buffered saline, 0.1% Tween 20). Membranes were incubated
with KLHDC7B antibody (ab126063; Abcam, Cambridge, UK) diluted at a
ratio of 1:500 according to the manufacturer's instructions and
incubated with the membranes overnight at 4°C. Goat horseradish
peroxidase (HRP)-linked secondary antibody was added at a dilution
ratio of (1:5,000) (31460; Pierce ThermoScientific) and incubated
with the membranes at room temperature for 1 h. β-actin (JLA20;
Calbiochem) was used as housekeeping (1:15,000, 1 h at room
temperature, secondary antibody not required). The membranes were
washed 3 times with TBS-T. Immunodetection of proteins was
performed using Amersham ECL Western Blotting Detection Reagent (GE
Healthcare, Chicago, IL, USA) according to the manufacturer's
instructions.
Statistical analysis
Prism 5.0 software (GraphPad Software Inc., La
Jolla, CA, USA) was used for statistics and data representation.
Data are presented as the mean ± standard error of the mean.
Significant differences were determined using ANOVA with Dunnett's
multiple comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Clinicopathological evaluation and
KLHDC7B characterization in breast cancer tumours
We studied the expression of KLHDC7B mRNA in tumours
from different pathological grades (Fig.
1A). Our results showed that KLHDC7B expression tended to
increase as tumour grade increased from grade 1 to 2. Grade 3
tumours showed a significant upregulation of KLHDC7B.
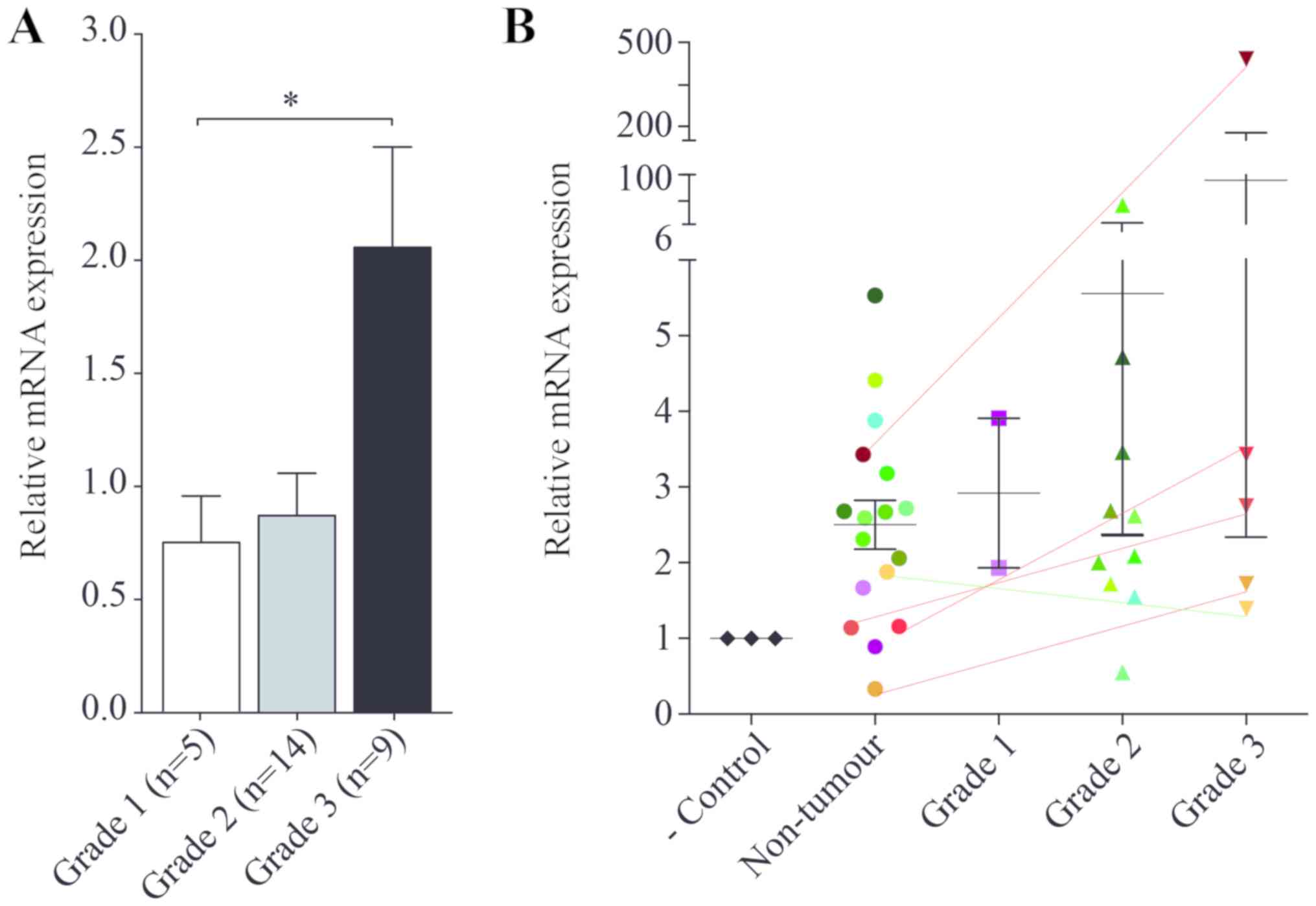 | Figure 1.KLHDC7B mRNA expression in breast
cancer. (A) mRNA expression in grade 1 (n=5), grade 2 (n=14) and
grade 3 (n=9) breast tumours, relative to grade 1 tumours. (B)
Relative mRNA expression in negative control (commercial RNA),
non-tumour surrounding breast tissue, and grade 1, 2 and 3 tumours.
Expression relative to negative control. For each tumour grade,
there is a corresponding mark of the same colour in non-tumour
tissue, allowing a comparison. Lines connect grade 3 tumours with
their respective non-tumour surrounding tissue (red, upregulated
expression; green, downregulated expression). *P<0.05, as
indicated. KLHDC7B, Kelch domain-containing protein 7B. |
Table II shows the
classification of tumours in order of increasing KLHDC7B
expression, with information on tumour type, tumour grade,
hormone-receptor (oestrogen and progesterone), HER and p53 status,
percentage of Ki67 positive cells, metastasis and vascular
invasion. Ki67 cells and tumour type showed a correlation with
KLHDC7B expression. Tumours with more than 10% Ki67 cells had the
highest levels of KLHDC7B expression. For tumour type, lobular
tumours had the lowest expression of KLHDC7B (83.33% of lobular
tumours were classified as having low expression), and ductal
tumours had the highest expression (68% of ductal tumours were
classified as having high expression). Papillary, mucinous,
cribriform and tubular tumours were also analysed, but due to the
low number of each of these tumour types, we cannot draw
conclusions on their relationship to KLHDC7B expression. Besides
tumour type and percentage of positive Ki67 cells, no other
correlations were found between KLHDC7B and the tumour features
described above.
In comparison to healthy tissue from the area
surrounding the tumour, with expression normalised to a commercial
RNA sample from a healthy donor (Fig.
1B), grade 3 tumours showed a tendency to KLHDC7B upregulation.
However, the expression of KLHDC7B in grade 3 tumours was not
always higher than non-tumour samples from other patients (from
grade 1- and 2-matched tissue).
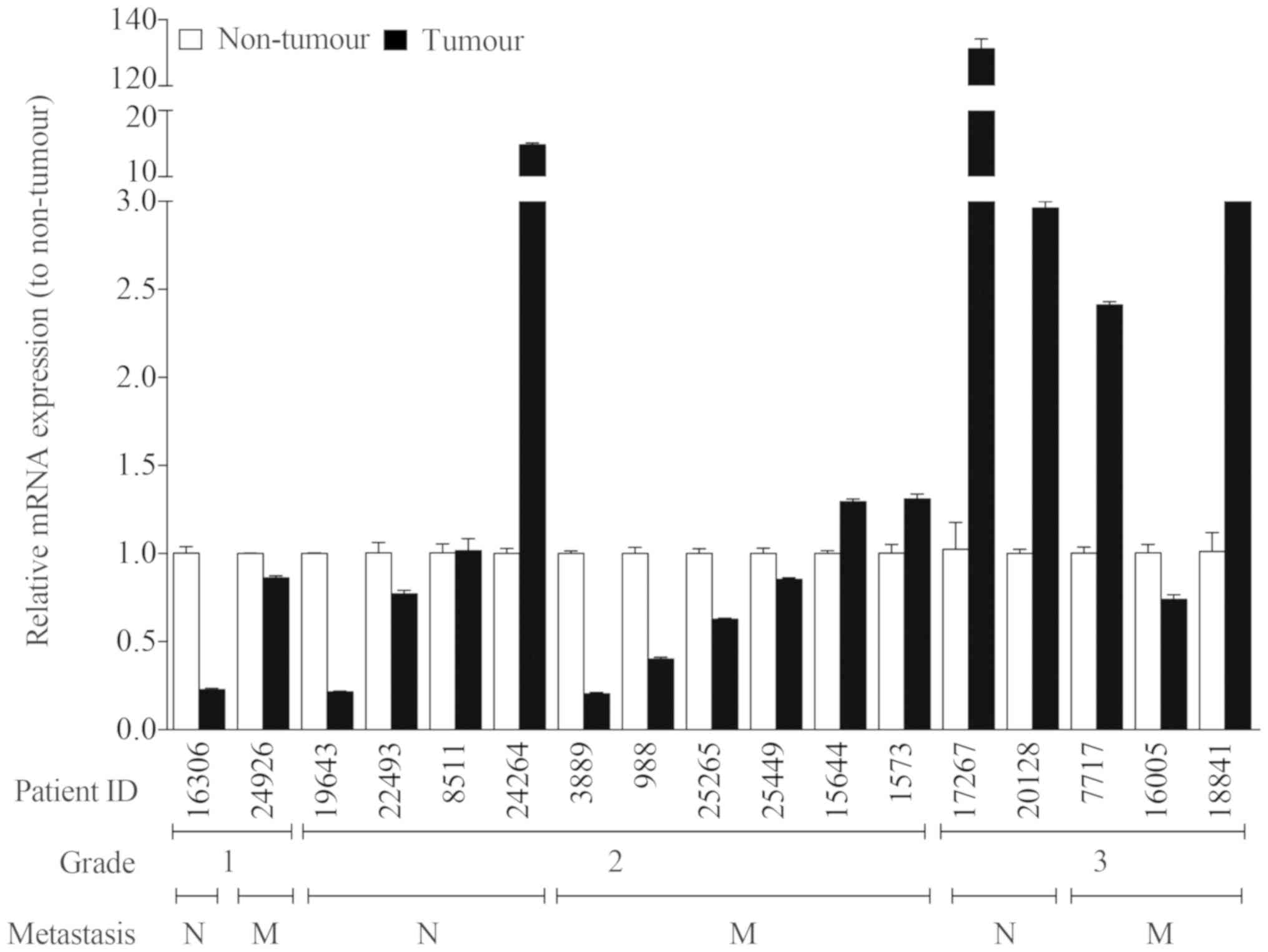
To avoid the influence of tumour heterogeneity among
patients, we compared the KLHDC7B expression in breast tumours and
in the surrounding healthy tissue in the same patient (Fig. 2). Seven out of 12 (58%) grade 1 and 2
tumours showed a significantly lower KLHDC7B expression compared to
non-tumour surrounding tissue. Only 3 out of 10 (30%) grade 2
tumours showed a significantly higher KLHDC7B expression in tumour
tissue. Four out of 5 (80%) grade 3 tumours showed a significantly
higher KLHDC7B expression than healthy tissue.
When we correlated KLHDC7B expression with
metastatic capacity (Fig. 2), 4 out
of 10 (40%) of the tumours that produced metastases showed
upregulation of this gene, but 5 out of 10 (50%) had
downregulation.
Characterization of KLHDC7B mRNA and
protein expression in cell lines
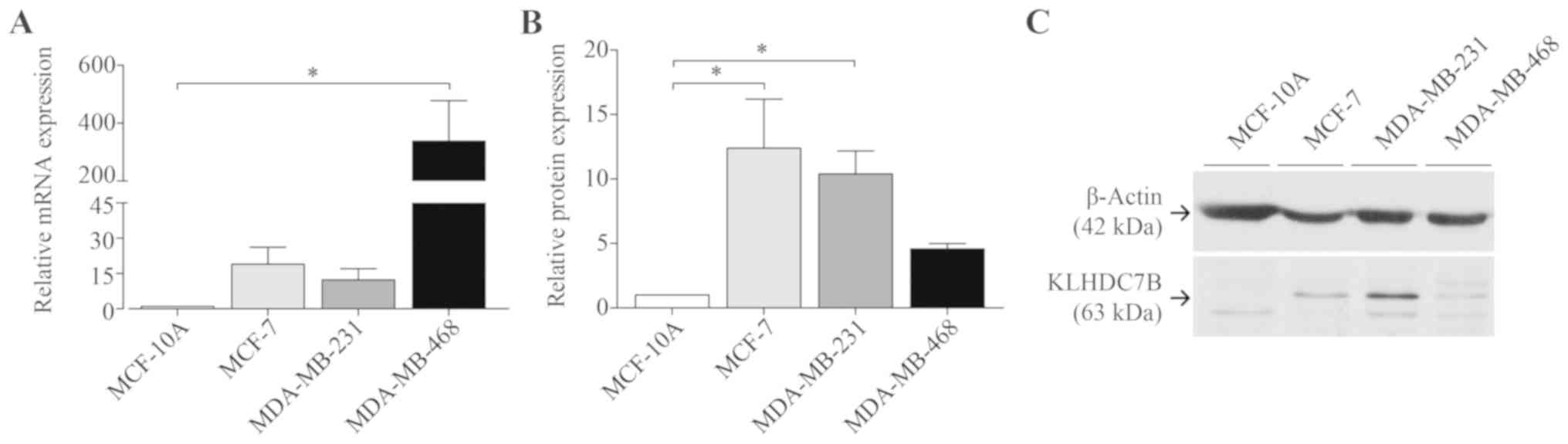
KLHDC7B mRNA expression was evaluated in different
cell lines, using MCF-10A as the non-tumour cell lines and MCF-7,
MDA-MB-231 and MDA-MB-468 as the tumour cell lines. As expected,
KLHDC7B was overexpressed in tumour cell lines (Fig. 3A).
Protein expression was determined by western-blot
(Fig. 3B and C). All four tumour
cell lines showed higher expression than MCF-10A (no expression).
Surprisingly, MDA-MB-468 showed lower expression than the other
tumour cell lines, although the mRNA expression was higher.
Discussion
KLHDC7B has been found in different tumours
including breast, ovary and cervical cancer (13,17–18,20),
revealing its possible role in tumour biology. The KLHDC7B gene has
been found to be hypermethylated and upregulated in breast cancer
and consequently has been postulated as an epigenetic marker of
breast cancer (17). However, its
role in the development and progression of these cancers is largely
unknown.
To understand the role of KLHDC7B in tumour
progression we studied the expression of KLHDC7B mRNA in tumours of
different pathological grades. Grade 3 tumours, tumours with more
than 10% Ki67 positive cells and ductal tumours had the highest
expression of KLHDC7B (Table II).
Ki67 expression is a well-known marker of active proliferation and
the association between high proliferation and poor prognosis is
well stablished (42–44). Lobular, mucinous, tubular, and
papillary carcinomas have been associated with lower risk of
mortality than ductal carcinomas (45). Together, these data indicate that
KLHDC7B is associated with more aggressive tumours and worse
prognosis.
The upregulation of KLHDC7B in advanced tumours
could suggest a positive association with metastatic capacity,
although we did not find such a difference in our analysis.
When we compared tumour tissue with healthy breast
tissue, KLHDC7B expression in tumour tissue was not always higher
than in non-tumour samples from other patients with grade 1 and 2
tumours. These data reveal a huge variability among individuals,
demonstrating one of the most relevant issues in
oncology-intertumour heterogeneity. To improve understanding of the
role of KLHDC7B in breast cancer, we compared KLHDC7B expression in
breast tumours and surrounding healthy tissue from the same
patient. This new approach confirmed previously published data on
KLHDC7B upregulation in breast tumours (17), but our results also revealed that the
expression of this gene is grade-dependent and only significantly
upregulated in grade 3 tumours. Additionally, we found interesting
results in grade 1 and 2 tumours, that KLHDC7B was downregulated in
well-differentiated and moderately-differentiated tumours. These
new data would suggest a dual role of KLHDC7B during tumour
progression, which we will analyse in future studies.
We can conclude that when using KLHDC7B expression
as a marker of breast cancer, it should be correlated against
healthy tissue from the same patient, rather than the general
population, as comparisons with the general population are likely
to lead to false results (a consequence of intertumour
heterogeneity). Additionally, use of KLHDC7B as a marker without
considering tumour grade could lead to inaccurate diagnoses.
The results of this study could increase
understanding of the involvement of KLHDC7B in breast cancer,
although the sample size poses a potential limitation, and future
studies should use a larger sample.
These data indicate that KLHDC7B is associated with
more aggressive tumours and worse prognosis, however they do not
explain the functional role of KLHDC7B in breast tumours. KLHDC7B
protein could have an anti- or pro-tumour role or even a dual role
that could explain the differences according to tumour grade. To
unravel the functional role of KLHDC7B, future experiments should
be performed including up- and down-regulation of KLHDC7B
expression to establish its role in the progression of breast
tumours. These studies should preferably be performed in breast
cell lines.
We studied the expression of KLHDC7B in tumour cell
lines and healthy cell lines. MCF-10A, MDA-MB-231 and MDA-MB-468
are hormone-receptor and HER negative cell lines; MCF-10A does not
express KLHDC7B, while the two MDA-MB cell lines do. In contrast,
MCF-7 is a hormone-receptor positive cell line and expresses
KLHDC7B (Table I). Our results
therefore suggest that hormone-receptor and HER status are not
related to KLHDC7B expression.
We have confirmed previously published results on
mRNA expression and added protein expression studies, which have
not previously been described. Our data show that mRNA expression
does not correlate exactly with protein expression. A previous
study by Gry et al (46)
showed that RNA does not always correlate with protein expression
and, more importantly, the correlation of the same protein can vary
depending on the cell line. This weak correlation is due to several
factors, including various post-transcriptional processes: For
example, some mRNAs are strongly retained in the nucleus, which may
lead to overestimation of RNA levels relative to protein levels.
The lack of correlation between RNA and protein levels in
MDA-MB-468 cells could be the result of complex regulatory
mechanisms. For future in vitro studies, protein expression
should be taken into consideration when selecting the appropriate
cell line model, and we recommend that MDA-MB-468 not be used as a
model to study the role of KLHDC7B protein in breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by Fondo de
Investigaciones Sanitarias (grant nos. PI17/02247 and PI14/01320),
Centro de Investigación Biomédica en Red de Cáncer (grant no.
CB16/12/00363), Generalitat de Catalunya (grant nos. 2017 SGR 1799
and 2014 SGR 1131), and the FP7 Marie Sklodowska-Curie COFUND
program (grant no. 267128; INCOMED program).
Availability of data and materials
All data generated or analysed during the present
study are included in this published article.
Authors' contributions
AMP designed and carried out the experiments,
interpreted the results, and wrote the manuscript. SRYC obtained
the human samples and patient data, and contributed to project
funding and manuscript revision. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The project was approved by the Clinical Research
Ethics Committee at Vall d'Hebron Hospital (PR(AG)309/2016).
Patients provided written informed consent for sample
collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Curado MP, Edwards B, Shin HR, Storm H,
Ferlay J, Heanue M and Boyle P: Cancer incidence in five
continents. 9. IARC Press; Lyon, France: 2007
|
|
3
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe:
Estimates for 40 countries in 2012. Eur J Cancer. 49:1374–1403.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo P, You JO, Yang J, Jia D, Moses MA and
Auguste DT: Inhibiting metastatic breast cancer cell migration via
the synergy of targeted, pH-triggered siRNA delivery and chemokine
axis blockade. Mol Pharm. 11:755–765. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elston CW and Ellis IO: Assessment of
histological grade. The breast. 13. Churchill Livingstone;
Edinburgh-New York: pp. 356–384. 1998
|
|
7
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leal JF, Fominaya J, Cascón A, Guijarro
MV, Blanco-Aparicio C, Lleonart M, Castro ME, Ramon Y Cajal S,
Robledo M, et al: Cellular senescence bypass screen identifies new
putative tumor suppressor genes. Oncogene. 27:1961–1970. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
López-Vicente L, Armengol G, Pons B, Coch
L, Argelaguet E, Lleonart M, Hernández-Losa J, de Torres I, Ramon Y
and Cajal S: Regulation of replicative and stress-induced
senescence by RSK4, which is down-regulated in human tumors. Clin
Cancer Res. 15:4546–4553. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
López-Vicente L, Pons B, Coch L, Teixidó
C, Hernández-Losa J, Armengol G, Ramon Y and Cajal S: RSK4
inhibition results in bypass of stress-induced and oncogene-induced
senescence. Carcinogenesis. 32:470–476. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rowinsky EK: Signal events: Cell signal
transduction and its inhibition in cancer. Oncologist (3 Suppl 8).
S5–S17. 2003. View Article : Google Scholar
|
|
12
|
Wade PA: Transcriptional control at
regulatory checkpoints by histone deacetylases: Molecular
connections between cancer and chromatin. Hum Mol Genet.
10:693–698. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Wu LY, Zhang XS and Zhang S:
Discovery of co-occurring driver pathways in cancer. BMC
Bioinformatics. 15:2712014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schones DE and Zhao K: Genome-wide
approaches to studying chromatin modifications. Nat Rev Genet.
9:179–191. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jones PA, Issa JP and Baylin S: Targeting
the cancer epigenome for therapy. Nat Rev Genet. 17:630–641. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hutchinson L: Breast cancer: Challenges,
controversies, breakthroughs. Nat Rev Clin Oncol. 7:669–670. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim TW, Kim YJ, Lee HJ, Min SY, Kang HS
and Kim SJ: Hs.137007 is a novel epigenetic marker hypermethylated
and up-regulated in breast cancer. Int J Oncol. 36:1105–1111. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Khirade MF, Lal G and Bapat SA: Derivation
of a fifteen gene prognostic panel for six cancers. Sci Rep.
5:132482015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jeong G, Bae H, Jeong D, Ham J, Park S,
Kim HW, Kang HS and Kim SJ: A Kelch domain-containing KLHDC7B and a
long non-coding RNA ST8SIA6-AS1 act oppositely on breast cancer
cell proliferation via the interferon signaling pathway. Sci Rep.
8:129222018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo P, Wang D, Wu J, Yang J, Ren T, Zhu B
and Xiang Y: The landscape of alternative splicing in cervical
squamous cell carcinoma. Onco Targets Ther. 8:73–79. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
NCBI Unigene: Hs.137007-Kelch domain
containing 7B (KLHDC7B). http://www.ncbi.nlm.nih.gov/UniGene/clust.cgi?UGID=150709&TAXID=9606&SEARCH=Hs.137007December.
2018
|
|
22
|
Hung-Chun Y: Comparative and functional
genomics study of the telomeric 7.73-Mb region from marker D22S418
through D22S1726 on human chromosome 22. UMI Dissertations
Publishing. 2008.ISBN 9780549576495 (pag 74, 75).
|
|
23
|
NCBI Reference Sequence: Kelch
domain-containing protein 7B [Homo sapiens]. https://www.ncbi.nlm.nih.gov/protein/NP_612442December.
2018
|
|
24
|
Laird PW: Cancer epigenetics. Hum Mol
Genet. 1:R65–R76. 2005. View Article : Google Scholar
|
|
25
|
Nabilsi NH, Broaddus RR and Loose DH: DNA
methylation inhibits p53-mediated survivin repression. Oncogene.
28:2046–2050. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cox GS, Gutkin DW, Haas MJ and Cosgrove
DE: Isolation of an Alu repetitive DNA binding protein and effect
of CpG methylation on binding to its recognition sequence. Biochim
Biophys Acta. 1396:67–87. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dobbelstein M and Moll U: Targeting
tumour-supportive cellular machineries in anticancer drug
development. Nat Rev Drug Discov. 13:179–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Issa JP: DNA methylation as a therapeutic
target in cancer. Clin Cancer Res. 13:1634–1637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
UniProtKB: Q96G42 (KLD7B_HUMAN), .
http://www.uniprot.org/uniprot/Q96G42December.
2018
|
|
30
|
Hara T, Ishida H, Raziuddin R, Dorkhom S,
Kamijo K and Miki T: Novel kelch-like protein, KLEIP, is involved
in actin assembly at cell-cell contact sites of Madin-Darby canine
kidney cells. Mol Biol Cell. 15:1172–1184. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stogios PJ and Privé GG: The BACK domain
in BTB-kelch proteins. Trends Biochem Sci. 29:634–637. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mantzouranis L, Bagattini R and Souza GM:
KeaA, a Dictyostelium Kelch-domain protein that regulates the
response to stress and development. BMC Dev Biol. 10:792010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Taguchi K, Motohashi H and Yamamoto M:
Molecular mechanisms of the Keap1-Nrf2 pathway in stress response
and cancer evolution. Genes Cells. 16:123–140. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gupta VA and Beggs AH: Kelch proteins:
Emerging roles in skeletal muscle development and diseases. Skelet
Muscle. 4:112014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sesé M, Fuentes P, Esteve-Codina A, Béjar
E, McGrail K, Thomas G, Aasen T, Ramón Y and Cajal S:
Hypoxia-mediated translational activation of ITGB3 in breast cancer
cells enhances TGF-β signaling and malignant features in vitro and
in vivo. Oncotarget. 8:114856–114876. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hevir N, Trošt N, Debeljak N and Rižner
TL: Expression of estrogen and progesterone receptors and estrogen
metabolizing enzymes in different breast cancer cell lines. Chem
Biol Interact. 191:206–216. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Woods Ignatoski KM, Dziubinski ML,
Ammerman C and Ethier SP: Cooperative interactions of HER-2 and
HPV-16 oncoproteins in the malignant transformation of human
mammary epithelial cells. Neoplasia. 7:788–798. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lim LY, Vidnovic N, Ellisen LW and Leong
CO: Mutant p53 mediates survival of breast cancer cells. Br J
Cancer. 101:1606–1612. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dai X, Cheng H, Bai Z and Li J: Breast
cancer cell line classification and its relevance with breast tumor
subtyping. J Cancer. 8:3131–3141. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jiang G, Zhang S, Yazdanparast A, Li M,
Pawar AV, Liu Y, Inavolu SM and Cheng L: Comprehensive comparison
of molecular portraits between cell lines and tumors in breast
cancer. BMC Genomics. 7 (Suppl 17):S5252016. View Article : Google Scholar
|
|
41
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Veronese SM, Gambacorta M, Gottardi O,
Scanzi F, Ferrari M and Lampertico P: Proliferation index as a
prognostic marker in breast cancer. Cancer. 71:3926–3931. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
van Diest PJ, van der Wall E and Baak JP:
Prognostic value of proliferation in invasive breast cancer: A
review. J Clin Pathol. 57:675–681. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Patani N, Martin LA and Dowsett M:
Biomarkers for the clinical management of breast cancer:
International perspective. Int J Cancer. 133:1–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li CI, Uribe DJ and Daling JR: Clinical
characteristics of different histologic types of breast cancer. Br
J Cancer. 93:1046–1052. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gry M, Rimini R, Strömberg S, Asplund A,
Pontén F, Uhlén M and Nilsson P: Correlations between RNA and
protein expression profiles in 23 human cell lines. BMC Genomics.
10:3652009. View Article : Google Scholar : PubMed/NCBI
|