Introduction
Lung cancer is the leading cause of cancer-related
mortality worldwide, resulting in ~1.8 million new cases and ~1.6
million deaths annually, with non-small cell lung cancer (NSCLC)
accounting for ~80% of all lung cancers (1). The clinical symptoms and manifestations
of early lung cancer are often hidden and lack specificity;
therefore, most patients are not diagnosed until the disease has
reached an advanced stage (2). The
recommended first-line therapy for advanced NSCLC (without driver
oncogenes targetable by a tyrosine kinase inhibitor and exhibiting
low programmed death ligand-1 expression), platinum-based doublet
chemotherapy, has significantly improved the overall survival and
quality of life for these patients (3,4);
however, after multiple cycles of chemotherapy, tumour cells can
develop resistance to chemotherapeutic drugs, which lowers their
activity against cancer cells (5).
Moreover, the efficacy of chemotherapy is often limited by
multidrug-resistance (MDR)-related proteins which extrude
anticancer drugs (6). Therefore,
investigating the molecular mechanisms underlying cisplatin
resistance in NSCLC has been a focus of recent research (7,8). Data
from many studies show that the mechanism of MDR in tumours is
mainly related to altered cell signaling through key proteins, such
as phosphoinositide 3-kinase-Akt, Janus N-terminal kinase and
Wnt/β-catenin (9–11). Accordingly, disrupting relevant
signaling might be an efficient strategy for reversing acquired
drug resistance. For example, microRNA (miR)-130a upregulation
directly inhibits the expression of the tumour-suppressor gene
runt-related transcription factor-3, leading to activation of
Wnt/β-catenin signaling and increased cisplatin resistance in
hepatocellular carcinoma (12).
Additionally, activation of the Wnt/β-catenin signaling pathway due
to cytoplasmic glycogen synthase kinase-3β (GSK-3β) inhibition
might explain cisplatin resistance observed in
A549/cis-diamminedichloroplatinum (II) (DDP) cells (13). Furthermore, maternally expressed 3
and miR-130b were also shown to enhance cisplatin resistance by
activating the Wnt/β-catenin signaling pathway in lung cancer
(14,15). These findings suggest that
Wnt/β-catenin signaling might be closely related to cisplatin
resistance.
Casein kinase II (CK2) is a highly conserved
serine/threonine kinase commonly found in eukaryotic cells that
most often exists as a tetrameric complex comprising of two
catalytic subunits and two regulatory subunits (16,17). CK2
exerts a variety of biological functions, including enhancing
growth (18), angiogenesis (19), invasion and metastasis (19,20) in
various human tumour xenografts, including lung cancer (19), urothelial cancer (21), mesothelioma (22), hepatocellular cancer (23) and gastric cancer (24), as well as participating in the
regulation of different signaling pathways. Previous findings
suggested CK2 as a positive regulator of Wnt signaling (25,26);
however, to the best of our knowledge, its association with
cisplatin resistance in lung cancer cells remains to be elucidated.
Previous reports have indicated that the inhibition of
dishevelled-2 (DVL-2; the most abundant isoform of the mammalian
DVL family) can re-sensitize cisplatin-resistant lung cancer cells
by downregulating Wnt/β-catenin signaling (27,28).
Additionally, a study has shown that DVL-2 is phosphorylated by CK2
(29). In the present study the
effect of CK2 on MDR in A549 lung cancer cells using A549/DDP cells
as a model system was investigated. Initially, CK2 and DVL-2 levels
in A549 and A549/DDP cells were determined, followed by western
blot analysis to detect levels of MDR-related proteins, including
multidrug-resistance-associated protein 1 (MPR1) and lung
resistance protein (LRP), in A549/DDP cells in the presence or
absence of CX4945 treatment. Thus, the aim of the present study was
to investigate the effects of CX4945 on the cisplatin
(DDP)-resistance of A549/DDP lung cancer cells and determine the
underlying molecular mechanism.
Materials and methods
Cells and cell culture
The human lung adenocarcinoma cell line A549 and its
cisplatin-resistant subline A549/DDP were obtained from the
Institute of Cell Biology. Cells were cultured in a humidified
incubator at 37°C and 5% CO2 in Roswell Park Memorial
Institute (RPMI)-1640 medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% heat-inactivated foetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin (HyClone; GE Healthcare Life Sciences). To maintain
drug resistance, A549/DDP cells were maintained in the presence of
0.5 µg/ml cisplatin (Merck KGaA).
Cell-proliferation analysis
The effect of cisplatin on cell growth was
determined using Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies Inc.) assays. For this, 100 µl A549 and A549/DDP cell
suspensions were distributed in quadruplicate in 96-well plates at
a density of 5×104 cells/well. Culture medium (100 µl)
was added to the wells of 96-well plates as a blank control group,
followed by incubation of both sets of plates at 37°C and 5%
CO2 for 24 h. The cells were then treated with or
without 15 µM CX4945 (Selleck Chemicals Co., Ltd.) for 24 h,
followed by an additional 24 h incubation with 0, 1.25, 2.5, 5, 10,
or 20 µg/ml cisplatin. CCK-8 solution (10 µl) was subsequently
added to each well of the plates and incubated for 1 h, after which
the absorbance at 450 nm was measured. The proliferation of A549
and A549/DDP cells was also determined by the same method after
cells were treated with cisplatin (5 µg/ml) for 0, 12, 24, 36, 48
and 72 h. This procedure was performed three times, and the mean
value was calculated. The concentration at which cisplatin alone or
in combination with CX4945 produced 50% growth inhibition
(IC50) was calculated based on the relative survival
curve.
Flow cytometric analysis of
apoptosis
A total of 5×105 A549/DDP cells/well were
cultured overnight in 6-well plates, followed by the addition of
CX4945 at a concentration of 0, 5, 10, 15, 20, or 25 µM. After 24
h, after cells were digested with trypsin with no EDTA, they were
centrifuged at 350 × g at 4°C for 5 min, washed twice with
phosphate-buffered saline, and resuspended in binding buffer (BD
Biosciences) at a concentration of 2×105 cells/ml.
Annexin V-conjugated fluorescein isothiocyanate and propidium
iodide (BioLegend, Inc.) was added to the binding buffer and
incubated for 15 min at room temperature. After incubation, the
percentage of apoptotic cells was detected using a flow cytometer
(BD FACSCalibur; BD Biosciences) and analysed usig FlowJO version
7.6.1 (Tree Star, Inc.). After dividing the A549/DDP cells into
four groups based on different treatments (RPMI-1640, DDP alone,
CX4949 alone, or combination treatment), apoptosis was detected
using flow cytometry.
Western blot analysis
After the treatment with RPMI-1640 medium, CX4945,
cisplatin or CX4945+cisplatin, and incubation for 24 h, whole-cell
lysates were harvested from A549 and A549/DDP cells using cell
lysis buffer (Beyotime Institute of Biotechnology). The protein
concentration was measured using a bicinchoninic acid assay, and
equal quantities (20 µg) of protein were resolved using SDS-PAGE
with a 10% gel. After resolving, the proteins were transferring to
polyvinylidene difluoride membranes. The membranes were blocked
with 5% bovine serum albumin (Sigma-Aldrich; Merck KGaA) for 1 h at
room temperature, and then incubated with a primary antibody
overnight at 4°C, followed by incubation with either horseradish
peroxidase (HRP) conjugated goat anti-rabbit (cat. no. 111-035-003;
1:10,000) and goat anti-mouse secondary antibodies (cat. no.
115-035-003; 1:10,000) (both from Jackson ImmunoResearch
Laboratories, Inc.) for 1 h at room temperature. Protein bands were
detected using enhanced chemiluminescence reagents (EMD
Millipore).
Antibodies against the following targets were used
for western blot analysis: CK2α (cat. no. sc-12738; 1:500; Santa
Cruz Biotechnology, Inc.), β-catenin (cat. no. sc-7963; 1:500;
Santa Cruz Biotechnology, Inc.), DVL-2 (cat. no. 373413; 1:1,000;
R&D Systems, Inc.), p-DVL-2Ser143 (cat. no.
ab124933; 1:1,000; Abcam), p-DVL-2Thr224 (cat. no.
ab124941; 1:1,000; Abcam), c-Myc (cat. no. 9402; 1:500; Cell
Signaling Technology, Inc.), caspase-3 (cat. no. sc-271759; 1:500;
Santa Cruz Biotechnology, Inc.), cleaved caspase-3 (cat. no. 9661;
1:500; Cell Signaling Technology, Inc.); cyclin D1 (cat. no. 2922;
1:500; Cell Signaling Technology, Inc.), MRP1 (cat. no. sc-365635;
1:1,000; Santa Cruz Biotechnology, Inc.) and LRP (cat. no.
sc-390134; dilution, 1:500; Santa Cruz Biotechnology, Inc.).
Densitometry analysis was performed using ImageJ version 1.52a
(National Institutes of Health). The respective total protein was
used as the loading control for phosphoproteins. β-actin was used
as the loading control for all other proteins.
RNA extraction and quantitative
reverse transcription polymerase chain reaction (RT-qPCR)
After the indicated treatments, total RNA was
extracted from cells from the four groups (blank control, cisplatin
alone, CX4945 alone and CX4945+cisplatin) using Trizol®
(Thermo Fisher Scientific, Inc.) according to manufacturer
instructions. Total RNA was reverse transcribed to cDNA at 42°C for
60 min followed by 72°C for 10 min, using a PrimeScript RT reagent
kit (Bio-Rad Laboratories, Inc.). qPCR was conducted using the
UltraSYBR reagent (CWBIO, Technology, Co. Ltd.) on a thermocycler
with the following cycling conditions: Initial denaturation for 3
min at 94°C, followed by 40 cycles of denaturation for 30 sec at
94°C, annealing for 30 sec at 60°C, extension for 30 sec at 72°C,
and a final extension step for 2 min at 72°C. Primer sequences are
listed in Table I. The RNA levels of
target genes were standardized against that of β-actin using the
2−ΔΔCq method (30). All
assays were performed in triplicate to ensure minimum
deviation.
 | Table I.Primer sequences used in the present
study. |
Table I.
Primer sequences used in the present
study.
| Primer name | Sequence |
|---|
| DVL-2 |
|
|
Forward |
5′-GAGGAAGAGACTCCCTACCTG-3′ |
|
Reverse |
5′-CGGGCGTTGTCATCTGAAAT-3′ |
| β-catenin |
|
|
Forward |
5′-CATCTACACAGTTTGATGCTGCT-3′ |
|
Reverse |
5′-GCAGTTTTGTCAGTTCAGGGA-3′ |
| C-myc |
|
|
Forward |
5′-GTCAAGAGGCGAACACACAAC-3′ |
|
Reverse |
5′-TTGGACGGACAGGATGTATGC-3′ |
| Cyclin D1 |
|
|
Forward |
5′-GCTGCGAAGTGGAAACCATC-3′ |
|
Reverse |
5′-CCTCCTTCTGCACACATTTGAA-3′ |
| MRP1 |
|
|
Forward |
5′-AAGGAGGTACTAGGTGGGCTT-3′ |
|
Reverse |
5′-CCAGTAGGACCCTTCGAGC-3′ |
| LRP |
|
|
Forward |
5′-TACATCCGGCAGGACAATGAG-3′ |
|
Reverse |
5′-CTGTGCAGTAGTGACGTGGG-3′ |
| Caspase-3 |
|
|
Forward |
5′-CATGGAAGCGAATCAATGGACT-3′ |
|
Reverse |
5′-CTGTACCAGACCGAGATGTCA-3′ |
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (v5.01; GraphPad Software, inc.). Data are expressed
as the mean ± standard deviation. Independent samples t-tests and
paired t-tests were used to compare data between two groups.
One-way analysis of variance with Bonferroni's correction was used
to compare data among three or more groups. A P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of cisplatin on A549 and
A549/DDP cell growth
A549 and A549/DDP cells were treated with cisplatin
at increasing concentrations (0, 1.25, 2.5, 5, 10 or 20 µg/ml), and
the viability of treated cells was detected using CCK-8 assays. The
viability of the cells was also determined by the same method after
treatment with cisplatin (5 µg/ml) for 0, 12, 24, 36, 48 and 72 h.
Cisplatin significantly inhibited A549 and A549/DDP cell growth in
a dose- and time-dependent manner (Fig.
1A and C), with IC50 values for cisplatin of
3.74±0.22 and 19.28±2.17 µg/ml in A549 and A549/DDP cells,
respectively (P=0.0002; Fig. 1B).
These results indicated that A549/DDP cells were 5.16-fold more
resistant to cisplatin compared with that of A549 cells. Moreover,
A549/DDP cell viability decreased until the concentration increased
to 5 µg/ml; therefore, we chose this as the optimal concentration
for treatment.
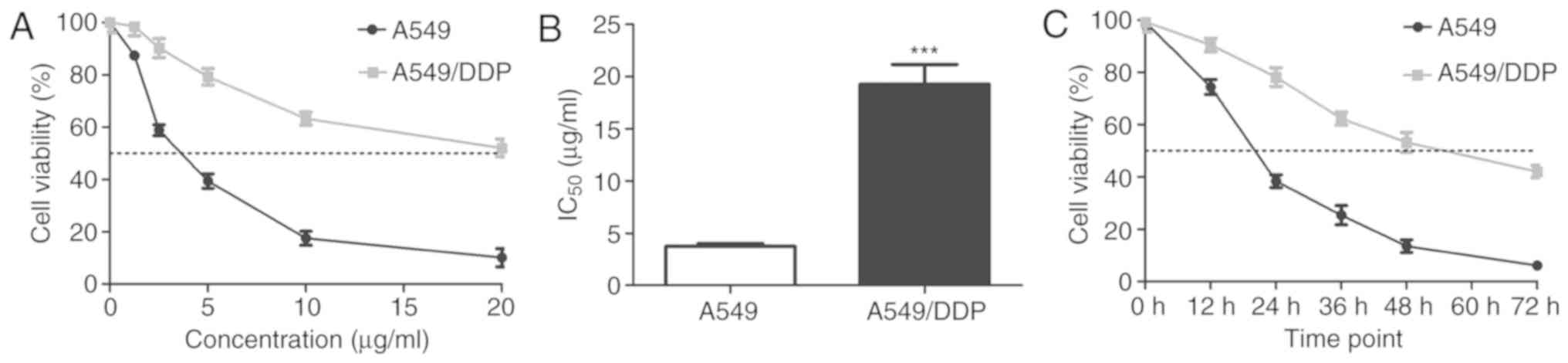 | Figure 1.Effect of cisplatin on A549 and
A549/DDP cell proliferation. (A) A549 and A549/DDP cells were
treated with 0, 1.25, 2.5, 5, 10, or 20 µg/ml cisplatin for 24 h,
and cell viability was detected using performing CCK-8 assays. (B)
IC50 value of cisplatin in A549 and A549/DDP cells (mean
± standard deviation; n=3). ***P<0.001 vs. A549 cells.
Statistical analysis was performed using independent samples
t-tests. (C) A549 and A549/DDP cells were treated with cisplatin (5
µg/ml) for 0, 12, 24, 36, 48 and 72 h and cell viability was
analysed using CCK-8 assay. CCK-8, Cell Counting Kit-8; DDP,
cis-diamminedichloroplatinum (II). |
Levels of
Wnt-signaling-pathway-related proteins and resistance-related
proteins in A549 and A549/DDP cells
Activation of Wnt/β-catenin signaling not only leads
to a variety of human diseases (31)
but it is also associated with cisplatin resistance (13). CK2 and DVL-2 participate in and
activate Wnt/β-catenin signaling (26,32).
Moreover, S143 and T224 of human DVL-2 are phosphorylated by
CK1δ/ε, and the S143A mutation inhibits polo-box domain (PB)-DVL-2
interaction, while the T224A mutation partially inhibits this
interaction (33). The
phosphorylation of DVL-2 and DVL-2-PBD binding are critical for
subsequent interaction with polo-like kinase 1, which is involved
in non-canonical Wnt signaling (33). However, whether these two
phosphorylation sites are associated with the canonical Wnt
signaling pathway remains unclear. Therefore, to determine whether
differences in responses to cisplatin between A549 and A549/DDP
cells were related to disparities in CK2 and DVL-2 levels, western
blot analysis was performed to evaluate CK2, total-DVL-2,
p-DVL-2Ser143 and p-DVL-2Thr224 levels in
both cell lines. As shown in Fig. 2,
CK2 and p-DVL-2Ser143 expression levels were
significantly higher in A549/DDP cells compared with the A549 cells
(P<0.001). Additionally, β-catenin levels were significantly
higher in A549/DDP cells (P<0.001), as were c-Myc and cyclin D1
(P<0.001), which are direct targets of lymphoid-enhancer-binding
factor/T cell factor (LEF/TCF) transcription factors and function
downstream of Wnt signaling (34).
However, the levels of total DVL-2 and p-DVL-2Thr224 in
A549 and A549/DDP cells were not significantly different (Fig. 2A and B). To further investigate
whether the resistance of A549/DDP was associated with
resistance-related proteins, levels of MRP1, LRP, caspase-3 and
cleaved-caspase-3 were analyzed using western blot, revealing
significantly higher MRP1 and LRP levels (P<0.001) and lower
caspase-3 (P<0.001) and cleaved caspase-3 (P<0.001) levels in
A549/DDP cells compared with that in A549 cells (Fig. 2C and D). These results suggested that
differences in cisplatin resistance between A549 cells and A549/DDP
cells might be associated with upregulated levels of MRP1, LRP, CK2
and p-DVL-2Ser143, which are important proteins
associated with the Wnt/β-catenin pathway.
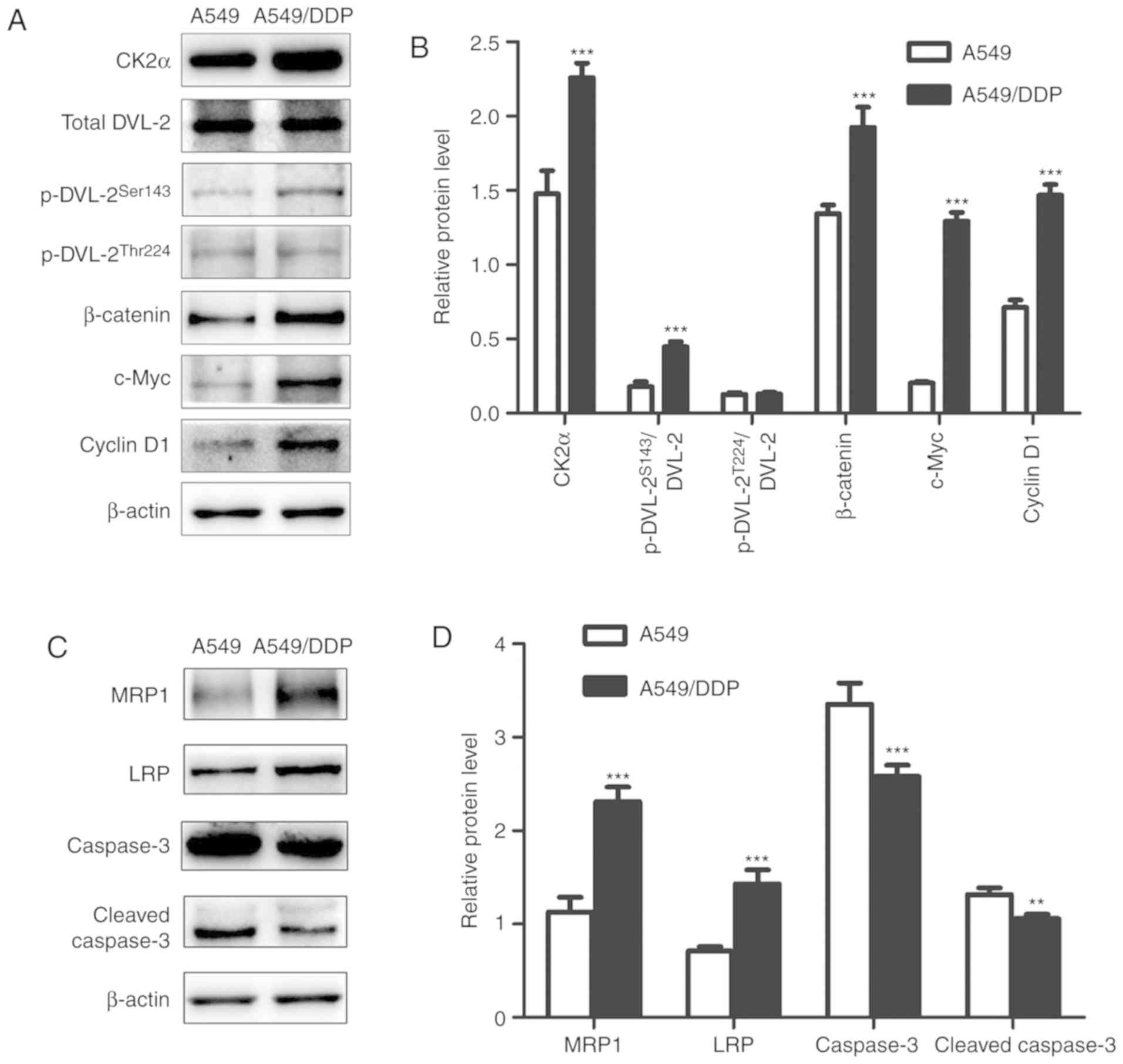 | Figure 2.Expression of proteins associated
with Wnt signaling and drug resistance, respectively, in A549 and
A549/DDP cells. (A and C) Western blot analyses using β-actin as an
internal control. Relative protein levels (mean ± standard
deviation; n=3) associated with the (B) Wnt signaling pathway and
(D) cisplatin resistance, in A549 and A549/DDP cells. **P<0.01,
***P<0.001 vs. A549 cells. Statistical analysis was performed
using independent samples t-tests. DDP,
cis-diamminedichloroplatinum (II); DVL-2, dishevelled-2; p,
phosphorylated; CK2, casein kinase II; MRP1, multidrug
resistance-associated protein 1; LRP, lung resistance protein. |
Effects of cisplatin on
Wnt-signaling-pathway and resistance-related proteins in A549 and
A549/DDP cells
Previous studies have not addressed DVL-mediated
regulation of cancer cell resistance to chemotherapy. To
investigate whether cisplatin resistance in lung adenocarcinoma
cells is associated with DVL-2 activity, total DVL-2,
p-DVL-2Ser143 and p-DVL-2Thr224 levels were
detected after cisplatin treatment of A549 and A549/DDP cells. No
changes in total DVL-2 or p-DVL-2Thr224 levels were
found upon cisplatin treatment in either cell line; however,
cisplatin treatment increased levels of CK2 and
p-DVL-2Ser143 in A549/DDP cells but decreased these
levels in A549 cells. Additionally, β-catenin, c-Myc and cyclin D1
levels were associated with CK2 levels in A549 and A549/DDP cells
(Fig. 3). A previous study showed
that the upregulated expression of β-catenin, cyclin D1 and c-Myc
resulted in the activation of the Wnt/β-catenin pathway (35); therefore, it was hypothesized that
cisplatin activates Wnt/β-catenin signaling in A549/DDP cells,
which might be related to upregulated CK2 and
p-DVL-2Ser143 levels. Moreover, previous studies suggest
that the mechanisms associated with cancer drug resistance are
closely related to resistance proteins and apoptosis (36–38).
Therefore, MRP1 and LRP levels were analyzed after cisplatin
treatment of A549 and A549/DDP cells, which revealed significantly
higher levels of these markers in A549/DDP cells (P<0.001),
whereas they were unchanged in A549 cells (P<0.001).
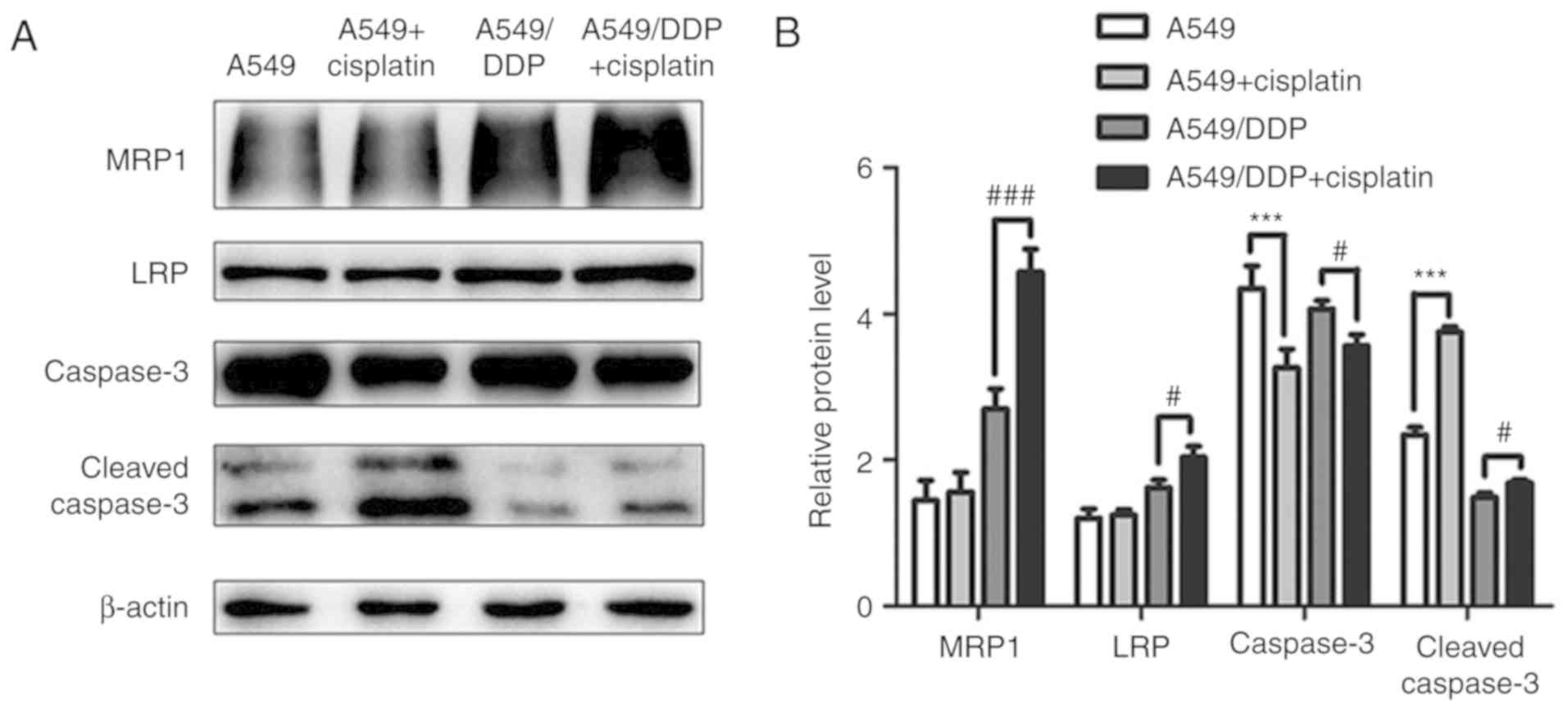
Furthermore, the levels of caspase-3 decreased and the levels of
cleaved caspase-3 increased in both cell lines following cisplatin
treatment compared with no cisplatin treatment (P<0.001), but
this upregulation was more significant in A549 cells compared with
that the A549/DDP cells (Fig.
4).
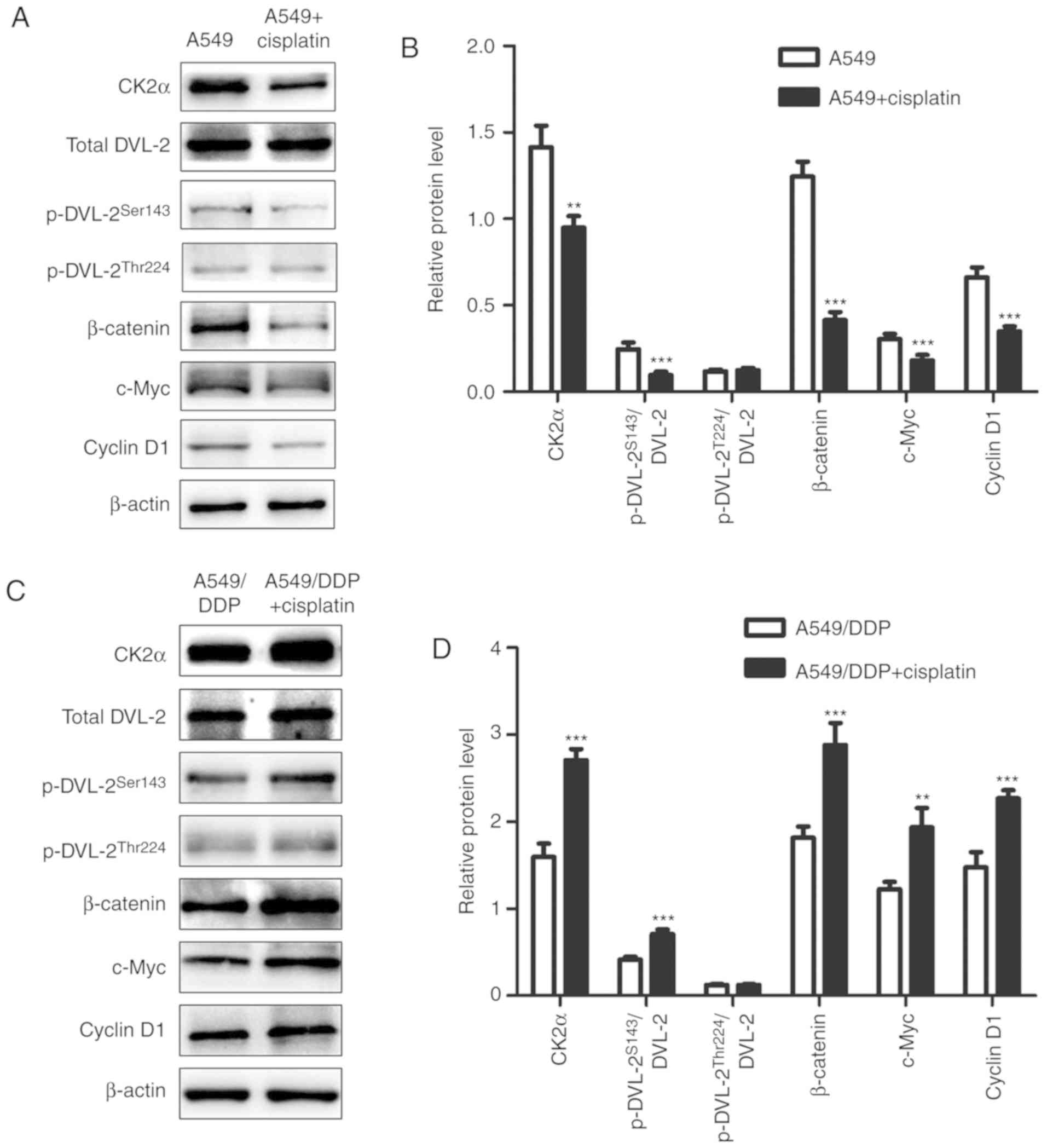 | Figure 3.Expression of CK2, DVL-2, β-catenin,
c-Myc and cyclin D1 in A549 and A549/DDP cells before and after
cisplatin treatment. (A and C) Western blot analyses using β-actin
as an internal control. (B and D) Relative protein levels (mean ±
standard deviation; n=3) of CK2, DVL-2, β-catenin, c-Myc and cyclin
D1 were determined in A549 and A549/DDP cells. **P<0.01,
***P<0.001 vs. A549 cells and A549/DDP cells before cisplatin
treatment. Statistical analysis was performed using paired t-tests.
CK2, casein kinase II; DVL-2, dishevelled-2; DDP,
cis-diamminedichloroplatinum (II). |
Effects of CX4945 on apoptosis and
resistance-related proteins in A549/DDP cells
To assess the effects of CX4945 on apoptosis,
A549/DDP cells were exposed to increasing concentrations of CX4945
(0, 5, 10, 15, 20, or 25 µM) for 24 h, followed by flow cytometric
analysis. Initially, the proportions of apoptotic cells increased
from 4.78 to 5.69%, 6.08 and 6.87% after treatment with 5, 10 and
15 µM CX4945, respectively; however, this percentage increased
markedly to 13.59% at a CX4945 concentration of 20 µM (Fig. 5A). Western blot analysis of similarly
treated A549/DDP cells for proteins associated with drug resistance
revealed decreasing levels of MRP1 and LRP with increasing CX4945
concentrations (Fig. 5B); however,
levels of caspase-3 decreased and cleaved-caspase-3 increased
notably up until a CX4945 concentration of 20 µM, consistent with
the flow cytometry data. Therefore, according to a previous finding
(39) and these results, 15 µM (as
apoptosis only increased at concentrations <20 µM) and 24 h was
chosen as the optimal concentration and time for pretreatment.
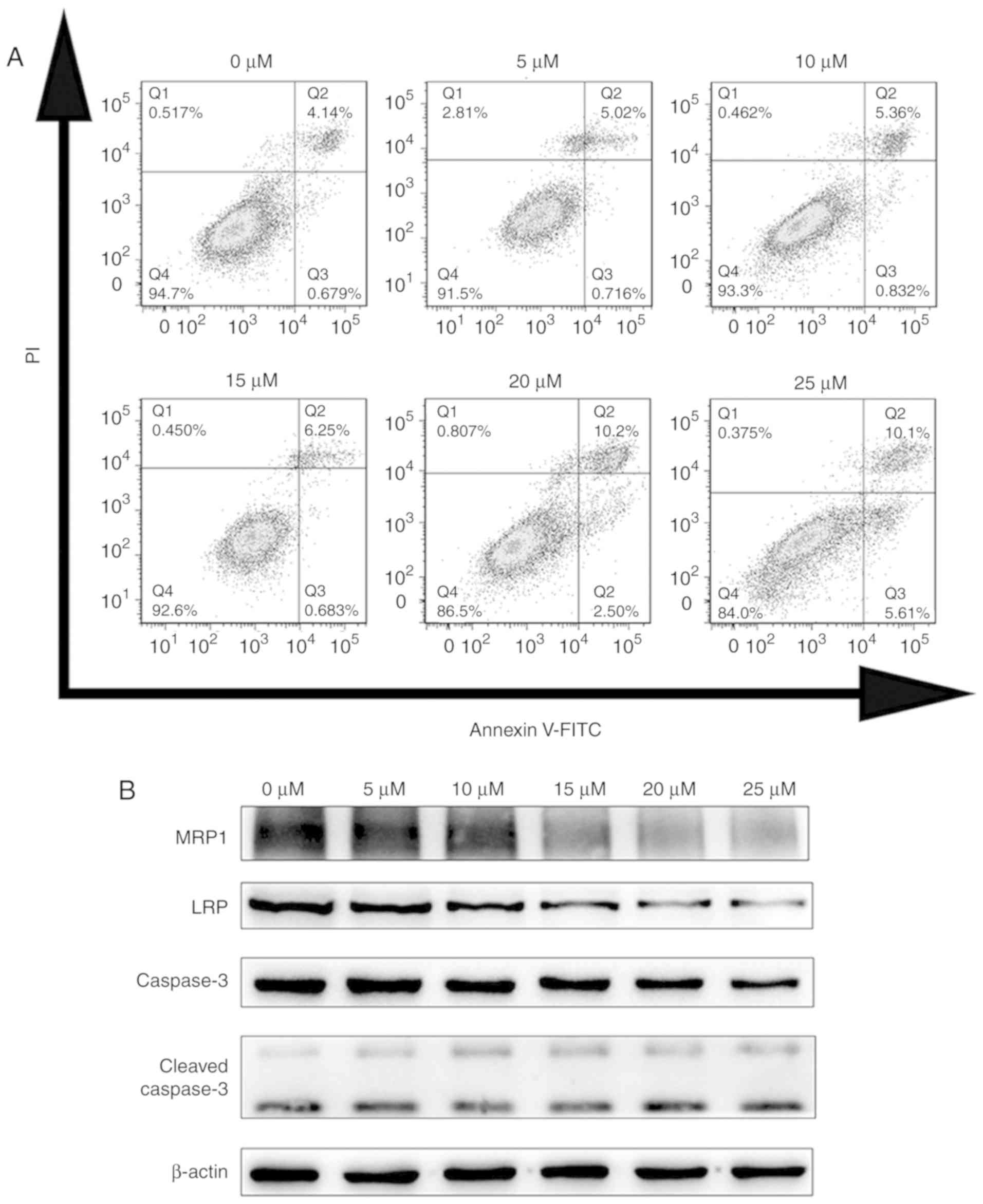 | Figure 5.Effects of CX4945 on apoptosis- and
drug resistance-related proteins in A549/DDP cells. (A) Flow
cytometric analysis of apoptosis in A549/DDP cells treated with 0,
5, 10, 15, 20, or 25 µM CX4945 for 24 h. (B) Protein levels of
MRP1, LRP, caspase-3 and cleaved caspase-3 in response to different
concentrations of CX4945 in A549/DDP cells. MRP1, multidrug
resistance-associated protein 1; LRP, lung resistance protein; DDP,
cis-diamminedichloroplatinum (II); PI, propidium iodide. |
CX4945 increased cisplatin sensitivity
and cisplatin-induced apoptosis in A549/DDP cells
To investigate whether CX4945 affects the
proliferation of A549/DDP cells, they were incubated with different
concentrations of cisplatin (0, 1.25, 2.5, 5, 10, or 20 µg/ml) in
the presence or absence of CX4945 (15 µM) for 24 h, followed by
assessment of cell viability using CCK-8 assay. The IC50
value for cisplatin in CX4945 treated cells was lower compared with
that for parental A549/DDP cells (6.43±0.32 vs. 18.43±1.56 µg/ml,
respectively) (Fig. 6A), indicating
that CX4945 treatment increased the cisplatin sensitivity of
A549/DDP cells. To confirm that CX4945 enhances cisplatin-mediated
apoptosis, flow cytometry was performed to detect apoptosis in
A549/DDP cells treated with RPMI-1640 (as a blank control group),
CX4945, cisplatin, or CX4945+cisplatin. The results revealed that
total apoptotic rates in groups treated with CX4945, cisplatin, or
CX4945+cisplatin were 8.62±0.43% (compared with the control group;
P<0.05), 9.64±0.38% (compared with the control group; P<0.05)
or 18.54±0.74% (compared with the control group; P<0.05), which
were higher compared with that of the control group (8.05±0.36%).
Furthermore, the apoptotic rate in the combination treatment group
(18.54±0.74%) was significantly higher compared with that in the
groups treated with CX4945 alone (8.65±0.43%) compared with
CX4945+cisplatin group (P<0.001), or cisplatin alone
(9.64±0.38%) compared with CX4945+cisplatin group (P<0.001;
Fig. 6B). These data indicated that
inhibition of CK2 activity by CX4945 enhanced cisplatin-induced
apoptosis in A549/DDP cells.
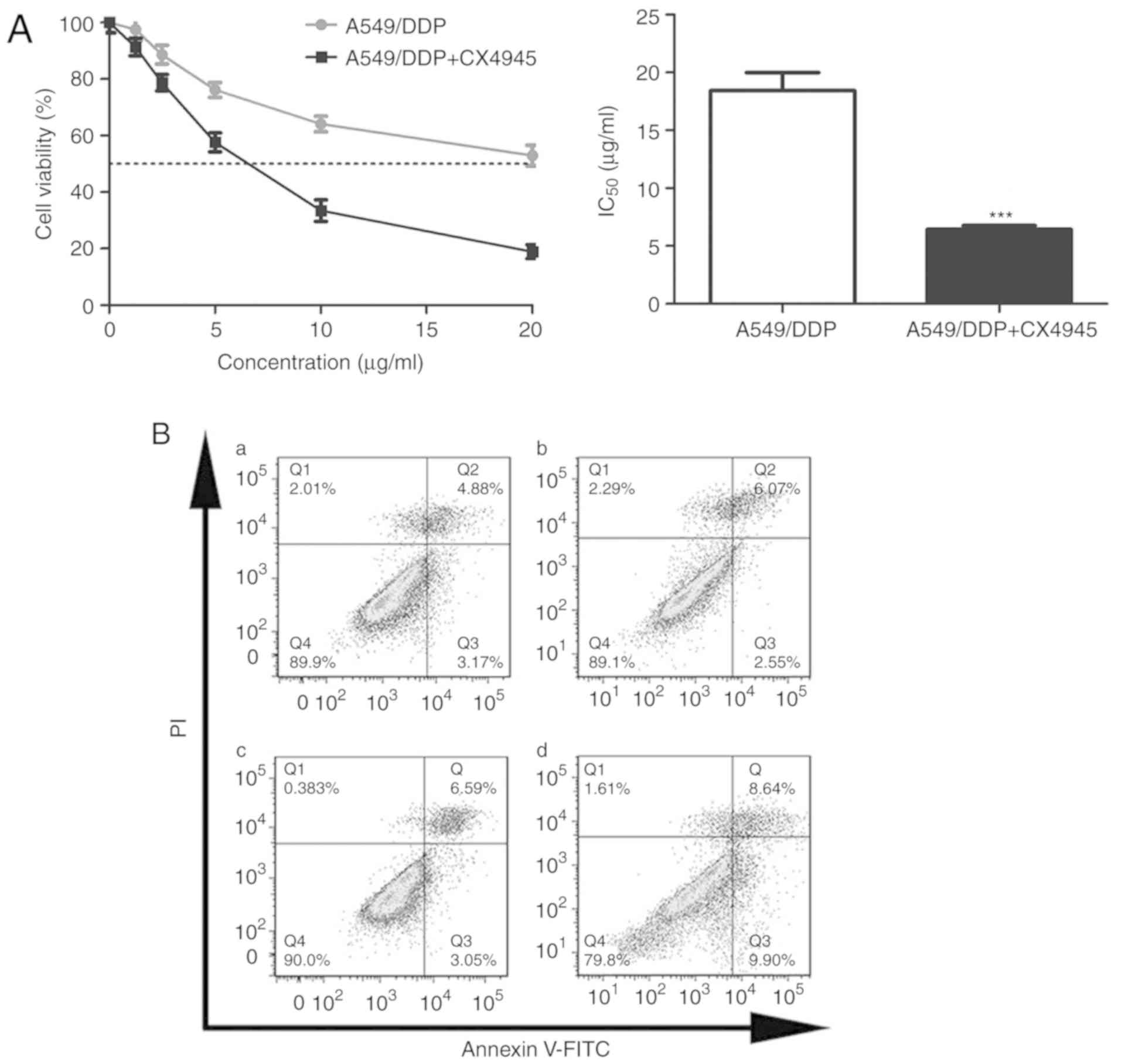 | Figure 6.Effect of CX4945 on the proliferation
and apoptosis of A549/DDP cells. (A) After pre-treatment with
CX4945 for 24 h, A549/DDP cells were treated with 0, 1.25, 2.5, 5,
10, or 20 µg/ml cisplatin for 24 h, and the IC50 values
of cisplatin in A549/DDP cells were determined (mean ± standard
deviation; n=3). ***P<0.001 vs. A549/DDP cells without CX4945
pre-treatment. Statistical analysis was performed using independent
samples t-tests. (B) A549/DDP cells were treated with (Ba)
RPMI-1640 medium (blank control), (Bb) CX4945, (Bc) cisplatin, or
(Bd) CX4945+cisplatin and incubated for 24 h. Apoptotic cells were
analysed by flow cytometry. DDP, cis-diamminedichloroplatinum (II);
PI, propidium iodide; IC50, 50% growth inhibition. |
CX4945 reversed cisplatin resistance
by blocking Wnt/β-catenin signaling in A549/DDP cells
The present study revealed that cisplatin can lead
to the activation of Wnt/β-catenin signaling, which might be
associated with CK2 and p-DVL-2Ser143 upregulation. To
supporting this finding, A549/DDP cells were treated with
RPMI-1640, CX4945, cisplatin, or CX4945+cisplatin, followed by
detection of mRNA and protein levels of Wnt signaling-related
molecules using RT-qPCR and western blot analysis, respectively.
p-DVL-2Ser143, β-catenin, c-Myc and cyclin D1 mRNA and
protein levels decreased significantly in the combined treatment
group compared with cisplatin treatment alone, whereas total DVL-2
and p-DVL-2Thr224 levels did not differ significantly
among the groups (P>0.05). Similar results were obtained upon
comparison of CX4945 only group with the control group (Fig. 7A-D). These results revealed that
inhibition of CK2 activity decreased p-DVL-2Ser143
levels and implied that DVL-2 phosphorylation at Ser143 by CK2
promotes activation of Wnt/β-catenin signaling.
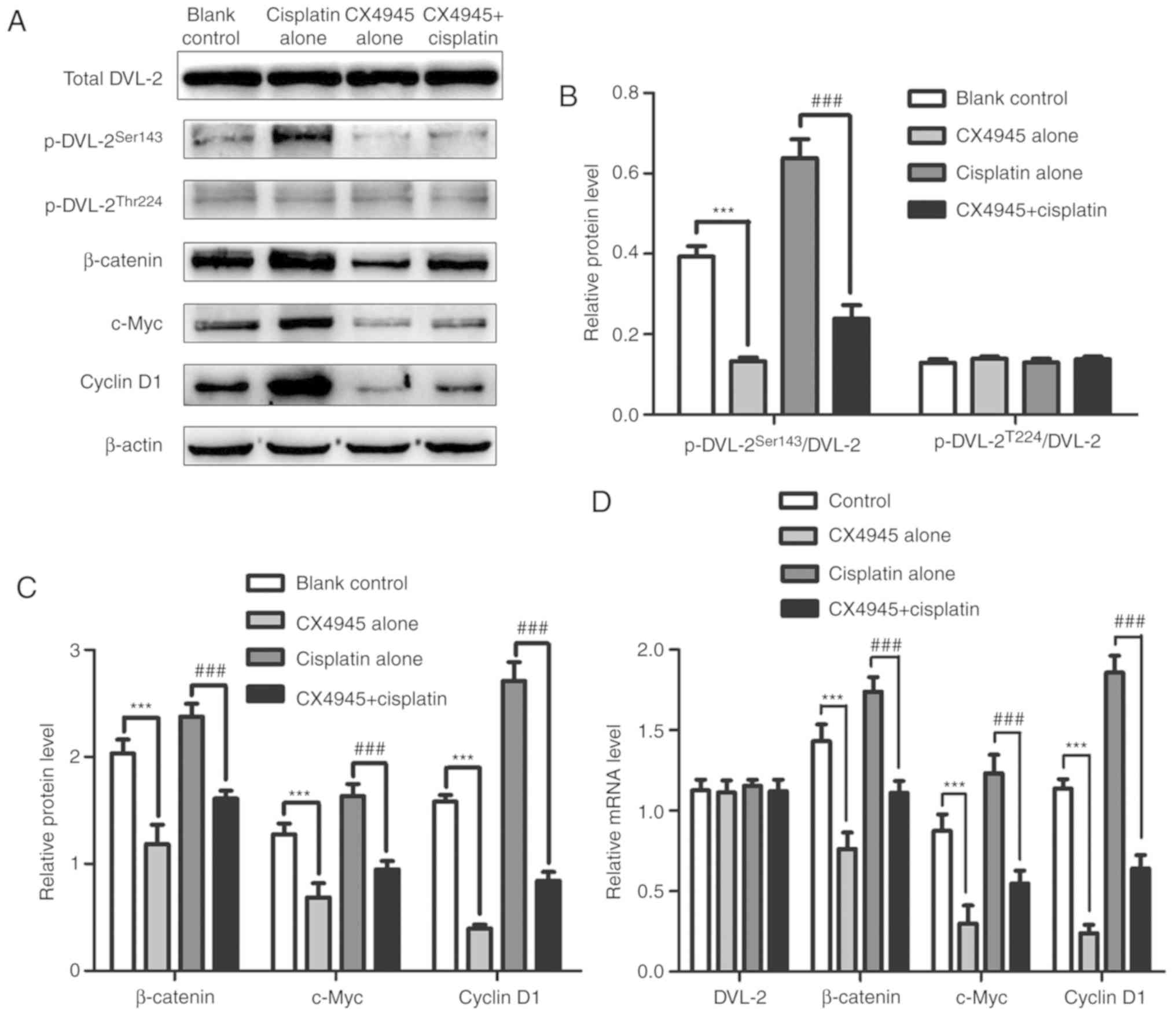 | Figure 7.Protein and mRNA levels of DVL-2,
β-catenin, c-Myc and cyclin D1 in A549/DDP cells treated with
RPMI-1640, CX4945, cisplatin, or CX4945+cisplatin. RPMI-1640 served
as the blank control. (A) Western blot analyses using β-actin as an
internal control. Relative protein expression levels of (B) DVL-2,
(C) β-catenin, c-Myc and cyclin D1 based on densitometry analysis
of (A). (D) mRNA expression of DVL2, β-catenin, c-Myc and cyclin D1
determined using reverse transcription-quantitative PCR. Data
represent the mean ± standard deviation of three independent
experiments and one-way analysis of variance. ***P<0.001 vs.
blank control group; ###P<0.001 vs. the cisplatin
only group. DDP, cis-diamminedichloroplatinum (II); DVL-2,
dishevelled-2. |
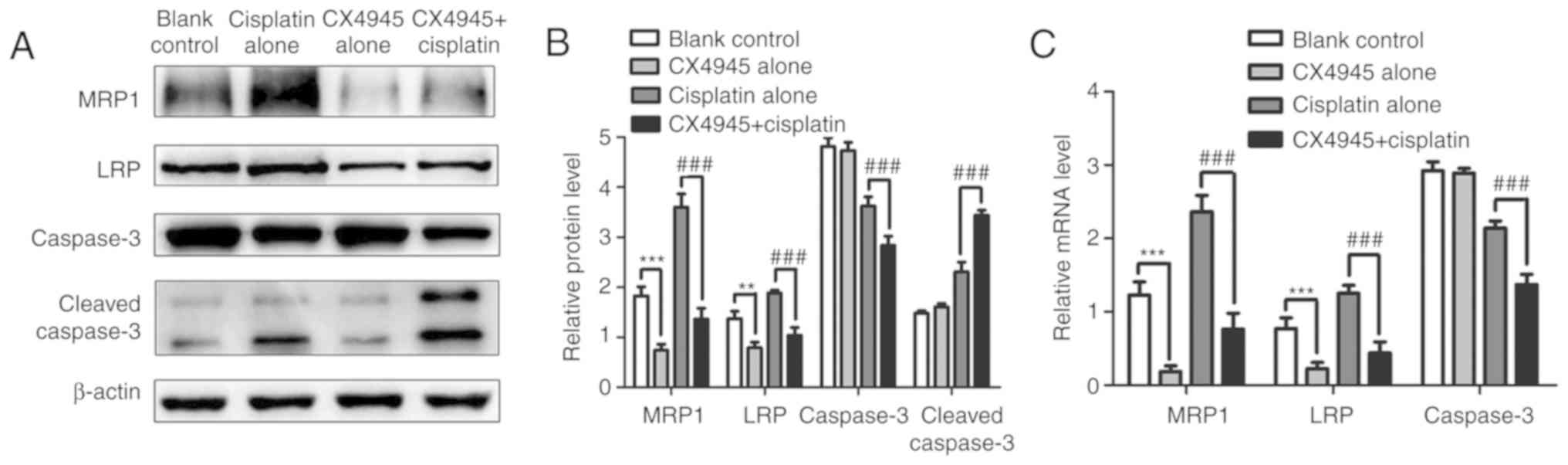
Additionally, MRP1, LRP, caspase-3 and
cleaved-caspase-3 protein and mRNA levels were analyzed, and
revealed that MRP1, LRP and caspase-3 levels in the
combined-treatment group were significantly lower compared with
that in the cisplatin-only group (P<0.01), whereas
cleaved-caspase-3 levels were significantly upregulated in the
combined-treatment group (P<0.001). Moreover, compared with that
in the control group, MRP1 and LRP mRNA and protein levels
decreased in the CX4945-only group, whereas caspase-3 mRNA and
protein levels and cleaved-caspase-3 protein levels remained
virtually unchanged between both groups. Levels of MRP1, LRP and
caspase-3 mRNA are consistent with their protein levels (Fig. 8A-C). These results indicated that
CX4945 reversed cisplatin resistance by blocking Wnt/β-catenin
signaling, downregulating levels of resistance-associated proteins,
and inducing apoptosis.
Discussion
Platinum drugs alone or in combination with other
drugs are considered the standard first-line chemotherapy for
patients with advanced lung cancer (3). However, intrinsic and acquired
resistance to cisplatin is a major obstacle for the clinical
treatment of NSCLC. The mechanism of drug resistance in lung cancer
is very complex, and many factors are involved. First, studies
identified mechanisms associated with membrane-transport
protein-mediated drug efflux pumps and associated with upregulation
of membrane proteins with efflux functions, such as P-glycoprotein,
MRP and LRP (38,40). Second, abnormal activation of
intracellular enzyme systems, such as topoisomerase (Topo) and
glutamyl transpeptidase, can combine with various chemotherapeutic
drug, such as platinum compounds, melphalan, cyclophosphamide,
chlorambucil, doxorubicin and nitrogen mustards to reduce their
activity (41). For example,
decreased DNA Topo II expression can cause tumour cells to rapidly
repair DNA and evade DNA-targeted chemotherapeutics, thereby
mediating drug resistance (42).
Third, upregulation of antiapoptotic markers, such as B-cell
lymphoma-2 and c-Myc, can lead to a decrease in cancer cell
apoptosis (43).
Cisplatin is the most commonly used chemotherapeutic
drug and causes tumour cell death by disrupting DNA structure and
function (44). Therefore, we
hypothesized that cisplatin resistance might be related to the
aberrant expression of signaling molecules induced by DNA
destruction, resulting in altered activation of associated
pathways. Activation of the Wnt/β-catenin pathway leads to
cisplatin resistance in lung cancer (13–15).
Moreover, CK2 is a positive regulator of Wnt signaling (25,26).
Therefore, we hypothesized that Wnt signaling might be associated
with cisplatin resistance in lung cancer cells; however, the role
of CK2 in cisplatin resistance in lung cancer cells, as well as the
potential underlying mechanism, remained unknown. To the best of
our knowledge, this is the first study investigating an association
between CK2 and cisplatin resistance. The results from the present
study showed that levels of CK2, p-DVL-2Ser143,
β-catenin, cyclin D1 and c-Myc were significantly higher in
A549/DDP cells compared with that in A549 cells, whereas no
differences were observed in total DVL-2 and
p-DVL-2Thr224 levels. Furthermore, significantly higher
levels of MRP1 and LRP were found in A549/DDP cells compared with
that in A549 cells, although the level of cleaved caspase-3 was
significantly lower in A549/DDP cells. As well-established
components of Wnt/β-catenin signaling, upregulated β-catenin,
cyclin D1 and c-Myc represent an activated state (or ‘on-state’) of
this pathway (35). Therefore, the
differences between A549 cells and A549/DDP cells in terms of
cisplatin resistance might be connected to CK2 and
p-DVL-2Ser143 upregulation and activation of Wnt
signaling.
Previous studies show that the co-receptor LRP5/6
forms a complex with Wnt-bound Frizzled in the presence of a Wnt
ligand (on-state), leading to the activation of DVL proteins via
sequential phosphorylation, which displaces GSK-3β from the
adenomatous polyposis coli/axin/GSK-3β complex (35,45,46).
Destruction of this complex stabilizes β-catenin, which accumulates
in the cytoplasm and results in its subsequent translocation to the
nucleus, where it binds LEF/TCF transcription factors to activate
the expression of c-Myc and cyclin D1 (45). DVL-2 participates in and activates
Wnt/β-catenin signaling (32);
however, the mechanism associated with DVL phosphorylation during
Wnt/β-catenin signaling remains unclear. To investigate whether CK2
and p-DVL-2Ser143 are associated with cisplatin
resistance, similar analyses was performed in the presence of this
drug, finding that following cisplatin treatment, CK2 and
p-DVL-2Ser143 levels in A549/DDP cells significantly
increased while they were significantly reduced in A549 cells.
However, no significant changes were observed in total DVL-2 and
p-DVL-2Thr224 levels upon cisplatin treatment of either
A549/DDP or A549 cells. Additionally, cisplatin treatment
significantly increased β-catenin, cyclin D1 and c-Myc levels in
A549/DDP cells but significantly downregulated these markers in
A549 cells. These data revealed a positive association between
activation of Wnt/β-catenin signaling and CK2 upregulation in
A549/DDP cells. Furthermore, MRP1 and LRP protein levels were
significantly upregulated after cisplatin treatment in A549/DDP
cells; however, these levels remained unchanged in A549 cells.
Furthermore, cleaved-caspase-3 levels increased significantly in
both cell lines after cisplatin treatment, although their
upregulation in A549/DDP cells was not as significant relative to
that in A549 cells. Based on these results, we hypothesized that
cisplatin activates Wnt/β-catenin signaling by upregulating CK2 in
A549/DDP cells but inactivates this pathway in A549 cells.
Moreover, DVL-2 phosphorylation at Ser143 by CK2 might account for
the activation of Wnt/β-catenin signaling, which was also
accompanied by MRP1 and LRP upregulation.
CX4945 is a potent and selective ATP-competitive
inhibitor of CK2 (47) and
reportedly inhibits not only the migration and invasion of A549
cells (39,48) but also suppresses cell growth through
induction of cell cycle arrest at the G2/M phase and
apoptosis (49). To address our
hypothesis regarding cisplatin resistance, A549/DDP cells were
divided into four groups, and similar analyses were performed,
revealing significantly decreased p-DVL-2Ser143 levels
in the combined treatment group compared with that in the cisplatin
only group and accompanied by the inactivation of Wnt/β-catenin
signaling and MRP1 and LRP downregulation. Additionally, inhibition
of CK2 activity decreased p-DVL-2Ser143 levels, implying
DVL-2 phosphorylation at Ser143 by CK2 promoted activation of
Wnt/β-catenin signaling. Analysis of cleaved-caspase-3 levels in
the four groups revealed that CX4945 treatment increased
cisplatin-induced apoptosis in A549/DDP cells by downregulating
MRP1 and LRP, with this finding confirmed by flow cytometric
analysis. Furthermore, the IC50 value of CX4945
pre-treated cells was lower compared with that of parental A549/DDP
cells. These results indicated that CX4945 reversed cisplatin
resistance by blocking Wnt/β-catenin signaling and downregulating
MRP1 and LRP levels.
In conclusion, the results from the present study
suggest that Wnt/β-catenin signaling can be clinically targeted
using CK2 inhibitors to induce sensitivity to platinum-based
chemotherapeutics used for treating lung cancer. These results
support the combined application of cisplatin with CX4945 for the
treatment of NSCLC. Further studies are required to confirm that
CX4945 will also function in vivo in order to predict the
clinical benefit for patients. Additionally, the use of only one
DDP resistant cell line may be a potential limitation of the
present study. Future studies will address this and also
investigate the involvement of associated signaling pathways,
besides the Wnt signaling pathway.
Acknowledgements
Not applicable.
Funding
This study was supported by the Scientific Research
Foundation Projects of the Affiliated Hospital of Jiangsu
University (grant no. 2015-013) and funds from the Zhenjiang
Science and Technology Development Project (grant no.
SH2013033).
Availability of data and materials
All data generated or analysed during the present
study are included in this published article.
Authors' contributions
CJ and PS conceived the study and designed the
experiments. CJ performed the experiments. CJ and JP collected and
analysed the experimental results. CJ drafted and revised the
article.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CCK-8
|
Cell Counting Kit-8
|
|
CK2
|
casein kinase II
|
|
DDP
|
cis-diamminedichloroplatinum (II)
|
|
DVL-2
|
dishevelled-2
|
|
GSK-3β
|
glycogen synthase kinase-3β
|
|
IC50
|
50% growth inhibition
|
|
LRP
|
lung resistance protein
|
|
MRP1
|
multidrug resistance-associated
protein 1
|
|
NSCLC
|
non-small cell lung cancer
|
|
RPMI
|
Roswell Park Memorial Institute
|
|
Topo
|
topoisomerase
|
References
|
1
|
Torre LA, Siegel RL and Jemal A: Lung
cancer statistics. Adv Exp Med Biol. 893:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maconachie R, Mercer T, Navani N and
McVeigh G; Guideline Committee, : Lung cancer: Diagnosis and
management: Summary of updated NICE guidance. BMJ. 364:l10492019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hellmann MD, Li BT, Chaft JE and Kris MG:
Chemotherapy remains an essential element of personalized care for
persons with lung cancers. Ann Oncol. 27:1829–1835. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lunacsek OE, Ravelo A, Coutinho AD, Hazard
SJ, Green MR, Willey J, Eaddy M and Goertz HP: First-line treatment
with bevacizumab and platinum doublet combination in Non-squamous
non-small cell lung cancer: A retrospective cohort study in US
oncology community practices. Drugs Real World Outcomes. 3:333–343.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Willers H, Azzoli CG, Santivasi WL and Xia
F: Basic mechanisms of therapeutic resistance to radiation and
chemotherapy in lung cancer. Cancer J. 19:200–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mirzaei SA, Reiisi S, Ghiasi TP, Shekari
A, Aliakbari F, Azadfallah E and Elahian F: Broad blocking of MDR
efflux pumps by acetylshikonin and acetoxyisovalerylshikonin to
generate hypersensitive phenotype of malignant carcinoma cells. Sci
Rep. 8:34462018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Zhang L, Yang J, Li B and Wang J:
CDH13 promoter methylation regulates cisplatin resistance of
non-small cell lung cancer cells. Oncol Lett. 16:5715–5722.
2018.PubMed/NCBI
|
|
8
|
Rose MC, Kostyanovskaya E and Huang RS:
Pharmacogenomics of cisplatin sensitivity in non-small cell lung
cancer. Genomics Proteomics Bioinformatics. 12:198–209. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou BG, Wei CS, Zhang S, Zhang Z and Gao
HM: Matrine reversed multidrug resistance of breast cancer
MCF-7/ADR cells through PI3K/AKT signaling pathway. J Cell Biochem.
119:3885–3891. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pang L, Lu J, Huang J, Xu C, Li H, Yuan G,
Cheng X and Chen J: Upregulation of miR-146a increases cisplatin
sensitivity of the non-small cell lung cancer A549 cell line by
targeting JNK-2. Oncol Lett. 14:7745–7752. 2017.PubMed/NCBI
|
|
11
|
Cui J, Jiang W, Wang S, Wang L and Xie K:
Role of Wnt/β-catenin signaling in drug resistance of pancreatic
cancer. Curr Pharm Des. 18:2464–2471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu N, Shen C, Luo Y, Xia L, Xue F, Xia Q
and Zhang J: Upregulated miR-130a increases drug resistance by
regulating RUNX3 and Wnt signaling in cisplatin-treated HCC cell.
Biochem Biophys Res Commun. 425:468–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao Y, Liu Z, Zhang X, He J, Pan Y, Hao F,
Xie L, Li Q, Qiu X and Wang E: Inhibition of cytoplasmic GSK-3b
increases cisplatin resistance through activation of Wnt/β-catenin
signaling in A549/DDP cells. Cancer Lett. 336:231–239. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xia Y, He Z, Liu B, Wang P and Chen Y:
Downregulation of Meg3 enhances cisplatin resistance of lung cancer
cells through activation of the WNT/β-catenin signaling pathway.
Mol Med Rep. 12:4530–4537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Q, Zhang B, Sun L, Yan Q, Zhang Y,
Zhang Z, Su Y and Wang C: MicroRNA-130b targets PTEN to induce
resistance to cisplatin in lung cancer cells by activating
Wnt/β-catenin pathway. Cell Biochem Funct. 36:194–202. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hanif IM, Hanif IM, Shazib MA, Ahmad KA
and Pervaiz S: Casein Kinase II: An attractive target for
anti-cancer drug design. Int J Biochem Cell Biol. 42:1602–1605.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Litchfield DW: Protein kinase CK2:
Structure, regulation and role in cellular decisions of life and
death. Biochem J. 369:1–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zou J, Luo H, Zeng Q, Dong Z, Wu D and Liu
L: Protein kinase CK2α is overexpressed in colorectal cancer and
modulates cell proliferation and invasion via regulating
EMT-related genes. J Transl Med. 9:972011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Benavent AF, Capobianco CS, Garona J,
Cirigliano SM, Perera Y, Urtreger AJ, Perea SE, Alonso DF and
Farina HG: CIGB-300, an anti-CK2 peptide, inhibits angiogenesis,
tumor cell invasion and metastasis in lung cancer models. Lung
Cancer. 107:14–21. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Amin EB, Mayo MW, Chudgar NP,
Bucciarelli PR, Kadota K, Adusumilli PS and Jones DR: CK2α′ drives
lung cancer metastasis by targeting BRMS1 nuclear export and
degradation. Cancer Res. 76:2675–2686. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shimada K, Anai S, Marco DA, Fujimoto K
and Konishi N: Cyclooxygenase 2-dependent and independent
activation of Akt through casein kinase 2α contributes to human
bladder cancer cell survival. BMC Urol. 11:82011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang S, Yang YL, Wang Y, You B, Dai Y,
Chan G, Hsieh D, Kim IJ, Fang LT, Au A, et al: CK2α, over-expressed
in human malignant pleural mesothelioma, regulates the Hedgehog
signaling pathway in mesothelioma cells. J Exp Clin Cancer Res.
33:932014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang HX, Jiang SS, Zhang XF, Zhou ZQ, Pan
QZ, Chen CL, Zhao JJ, Tang Y, Xia JC and Weng DS: Protein kinase
CK2α catalytic subunit is overexpressed and serves as an
unfavorable prognostic marker in primary hepatocellular carcinoma.
Oncotarget. 6:34800–34817. 2015.PubMed/NCBI
|
|
24
|
Bae JS, Park SH, Kim KM, Kwon KS, Kim CY,
Lee HK, Park BH, Park HS, Lee H, Moon WS, et al: CK2α
phosphorylates DBC1 and is involved in the progression of gastric
carcinoma and predicts poor survival of gastric carcinoma patients.
Int J Cancer. 136:797–809. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dowling JE, Alimzhanov M, Bao L, Chuaqui
C, Denz CR, Jenkins E, Larsen NA, Lyne PD, Pontz T, Ye Q, et al:
Potent and selective CK2 kinase inhibitors with effects on Wnt
pathway signaling in vivo. ACS Med Chem Lett. 7:300–305. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seldin DC, Landesman-Bollag E, Farago M,
Currier N, Lou D and Dominguez I: CK2 as a positive regulator of
Wnt signalling and tumourigenesis. Mol Cell Biochem. 274:63–67.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee YN, Gao Y and Wang HY: Differential
mediation of the Wnt canonical pathway by mammalian Dishevelleds-1,
−2 and −3. Cell Signal. 20:443–452. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo K, Gu X, Liu J, Zeng G, Peng L, Huang
H, Jiang M, Yang P, Li M, Yang Y, et al: Inhibition of disheveled-2
resensitizes cisplatin-resistant lung cancer cells through
down-regulating Wnt/β-catenin signaling. Exp Cell Res. 347:105–113.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Song DH, Sussman DJ and Seldin DC:
Endogenous protein kinase CK2 participates in Wnt signaling in
mammary epithelial cells. J Biol Chem. 275:23790–23797. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smalley MJ, Signoret N, Robertson D,
Tilley A, Hann A, Ewan K, Ding Y, Paterson H and Dale TC:
Dishevelled (Dvl-2) activates canonical Wnt signalling in the
absence of cytoplasmic puncta. J Cell Sci. 118:5279–5289. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee KH, Johmura Y, Yu LR, Park JE, Gao Y,
Bang JK, Zhou M, Veenstra TD, Yeon Kim B and Lee KS: Identification
of a novel Wnt5a-CK1ε-Dvl2-Plk1-mediated primary cilia disassembly
pathway. EMBO J. 31:3104–3117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cadigan KM and Waterman ML: TCF/LEFs and
Wnt signaling in the nucleus. Cold Spring Harb Perspect Biol.
4(pii): a0079062012.PubMed/NCBI
|
|
35
|
Angers S and Moon RT: Proximal events in
Wnt signal transduction. Nat Rev Mol Cell Biol. 10:468–477. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu C, Gong Q, Chen T, Lv J, Feng Z, Liu P
and Deng Z: Treatment with 20(S)-ginsenoside Rg3 reverses multidrug
resistance in A549/DDP xenograft tumors. Oncol Lett. 15:4376–4382.
2018.PubMed/NCBI
|
|
37
|
Wu Q, Yang Z, Nie Y, Shi Y and Fan D:
Multi-drug resistance in cancer chemotherapeutics: Mechanisms and
lab approaches. Cancer Lett. 347:159–166. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ku MJ, Park JW, Ryu BJ, Son YJ, Kim SH and
Lee SY: CK2 inhibitor CX4945 induces sequential inactivation of
proteins in the signaling pathways related with cell migration and
suppresses metastasis of A549 human lung cancer cells. Bioorg Med
Chem Lett. 23:5609–5613. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei H, Lu W, Li M, Zhang Q and Lu S:
Concomitance of P-gp/LRP expression with EGFR mutations in exons 19
and 21 in non-small cell lung cancers. Yonsei Med J. 57:50–57.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Backos DS, Franklin CC and Reigan P: The
role of glutathione in brain tumor drug resistance. Biochem
Pharmacol. 83:1005–1012. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang H, Liu J, Meng Q and Niu G:
Multidrug resistance protein and topoisomerase 2 alpha expression
in non-small cell lung cancer are related with brain metastasis
postoperatively. Int J Clin Exp Pathol. 8:11537–11542.
2015.PubMed/NCBI
|
|
43
|
Javid J, Mir R, Mirza M, Imtiyaz A,
Prasant Y, Mariyam Z, Julka PK, Mohan A, Lone M, Ray PC and Saxena
A: CC genotype of anti-apoptotic gene BCL-2 (−938 C/A) is an
independent prognostic marker of unfavorable clinical outcome in
patients with non-small-cell lung cancer. Clin Transl Oncol.
17:289–295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kelland L: The resurgence of
platinum-based cancer chemotherapy. Nat Rev Cancer. 7:573–584.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Macdonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim J and Kim SH: Druggability of the CK2
inhibitor CX-4945 as an anticancer drug and beyond. Arch Pharm Res.
35:1293–1296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim J and Hwan KS: CK2 inhibitor CX-4945
blocks TGF-β1-induced epithelial-to-mesenchymal transition in A549
human lung adenocarcinoma cells. PLoS One. 8:e743422013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
So KS, Rho JK, Choi YJ, Kim SY, Choi CM,
Chun YJ and Lee JC: AKT/mTOR down-regulation by CX-4945, a CK2
inhibitor, promotes apoptosis in chemorefractory non-small cell
lung cancer cells. Anticancer Res. 35:1537–1542. 2015.PubMed/NCBI
|