Introduction
Osteosarcoma is the most common primary malignant
bone tumor with the majority of cases occurring in the lower long
bones (1). It is predominantly
diagnosed in children and adolescents aged between 10 and 25 years
(2,3). Patients with osteosarcoma are likely to
experience pain and are susceptible to bone fracture due to
weakening of the affected bone. Radiotherapy and chemotherapy alone
may not be sufficient to treat osteosarcoma, and complete cure of
the disease often requires surgical resection. To date, the most
common treatment for osteosarcoma is by limb-salvage, which is the
removal of tumor without amputation. However, due to the high local
recurrence after the initial treatment, it is often combined with
chemotherapy, or further amputation if necessary (4).
Previous studies have revealed that the amount of
non-coding DNA in genomes increased consistently as eukaryotic cell
complexity increased (5,6). Non-coding DNA accounts for the majority
of the human genome, and this is transcribed into non-coding RNA
(ncRNA). Among the ncRNAs, long non-coding RNA (lncRNA), micro RNA
(miRNA/miR) and small interfering RNA (siRNA) serve regulatory
roles in cells. ncRNAs often demonstrate plasticity in expression
(7). Dysregulation of ncRNA
expression and function is associated with a number of different
diseases, including gastric cancer (8), prostate cancer (9), preeclampsia (10), and neurodegenerative diseases
(11). lncRNAs consist of >200
nucleotides and serve important roles in cell development and
differentiation (12–14). Furthermore, alteration of lncRNA
expression is associated with different types of cancer (15–18).
Thus, lncRNAs may serve as potential biomarkers for cancer
(19). The lncRNA LINC00152 has been
implicated in several types of cancer including hepatocellular
carcinoma, gastric cancer and gallbladder cancer, by affecting cell
proliferation, cell cycle arrest and apoptosis (20–23).
LINC00152 interacts with multiple signaling pathways such as the
mTOR signaling pathway by binding onto promoters through
cis-regulation and attenuating the expression of certain proteins,
or by binding directly onto the proteins (20). Furthermore, LINC00152 functions as an
endogenous sponge by binding to a number of miRNAs, including
miR-138, miR-4647, miR-103a-3p, miR-4775 and miR-139-5p, and
inhibiting their functions (24–29).
The aim of the present study was to elucidate the
biological functions of LINC00152 in osteosarcoma, as well as the
underlying molecular mechanisms involved. The expression level of
LINC00152 in osteosarcoma cell lines was analyzed and compared with
that in normal cells. LINC00152 knockdown was used to analyze the
effect of LINC00152 on cell proliferation and to demonstrate
whether LINC00152 exhibits tumorigenic effects similar to those
reported in other studies (30–33). The
current study also aimed to reveal potential miRNA candidates for
LINC00152 binding. Sequence-specific binding between miR-193b-3p
and LINC00152 was validated by a dual-luciferase assay. The overall
effects of the differential expression of miR-193b-3p and LINC00152
was further investigated by performing loss-of-function
experiments. The results obtained in the current study increase the
understanding of the role of LINC00152 in osteosarcoma and may aid
the development of treatment strategies which result in decreased
recurrence and an increased survival rate, as well as decreased
risk and costs associated with surgical resection.
Materials and methods
Cell culture
The human osteosarcoma cell lines U2OS, Saos-2, MG63
and MNNG/HOS, as well as the human osteoblast cell line HFOB 1.19,
were obtained from Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China). The HFOB 1.19 cells were
cultured in Ham's F-12 nutrient mixture containing 10% fetal bovine
serum (FBS); and the MNNG/HOS cell line was cultured in Eagle's
minimum essential medium. All other cell lines were cultured in
RPMI medium 1640 containing 10% FBS. All culture media, as well as
FBS, were purchased from Gibco; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). All cell lines were incubated at 37°C and with
5% CO2 in an incubator.
Cell transfection
Transfections were performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol. The
miR-193b-3p inhibitor and two siRNAs targeting LINC00152 were
purchased from Shanghai GeneChem Co., Ltd. (Shanghai, China). The
following sequences were used: i) miR-193b-3p inhibitor,
5′-AGCGGGACUUUGAGGGCCAGUU-3′; ii) miR-negative control (NC),
5′-CAGUACUUUUGUGUAGUACAA-3′; iii) si-LINC00152-1,
5′-UGAUCGAAUAUGACAGACACCGAAA-3′; iv) si-LINC00152-2,
5′-CAGGGAAUCUUUCAGCUGGAUUCCG-3′; and v) si-NC,
5′-UUCUCCGAACGUGUCACGUdTdT-3′. The concentration of miR-193b-3p
inhibitor and miR-NC was 150 nM. The concentration of
si-LINC00152-1, si-LINC00152-2 and si-NC was 40 nM. Subsequent
experiments were performed 48 h after transfection. The human
osteosarcoma cell lines U2OS, Saos-2, MG63, MNNG/HOS and the human
osteoblast cell line HFOB 1.19 were used for transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from clinical tissues and
cultured cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
First-strand cDNA was synthesized using a Reverse Transcription it
(Takara, Bio, Inc., Otsu, Japan). The reaction was carried out at
42°C for 60 min and was terminated by heating to 70°C for 5 min.
qPCR was subsequently performed using the SYBR® Green
PCR Master mix (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol, and an ABI Prism 7500 Sequence Detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
following primer pairs were used for the qPCR: LINC00152, forward
5′-AAAATCACGACTCAGCCCCC-3′ and reverse, 5′-AATGGGAAACCGACCAGACC-3′;
GAPDH forward, 5′-GGGAGCCAAAAGGGTCAT-3′ and reverse,
5′-GAGTCCTTCCACGATACCAA-3′; miR-193b-3p forward,
5′-AACTGGCCCTCAAAGTCCC-3′ and reverse, 5′-ATACCTCGGACCCTGCACTG-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The PCR thermocycling conditions were:
Pre-denaturation at 94°C for 3 min; followed by 35 cycles of 94°C
for 50 sec, 37°C for 1 min, 72°C for 1.5 min; with a final
extension of at 72°C for 7 min. U6 small nuclear RNA (snRNA) was
used as a normalization control for the detection of miR-193b-3p
and GAPDH was used as a normalization control for LINC00152. mRNA
levels were quantified using the 2−ΔΔCq method and
normalized to the controls (U6 snRNA or GAPDH) (34). RT-qPCR was used to determine
expression in U2OS, Saos-2, MG63, MNNG/HOS and HFOB 1.19 cells.
Cell proliferation assay
The U2OS transfected cells were adjusted to a cell
density of 2×103 cells/well, then seeded into 96-well
plates and cultured for 0, 24, 48 and 72 h. A total of 10 µl Cell
Counting Kit-8 reagent (CCK-8; Beyotime Institute of Biotechnology,
Haiman, China) was added to each well. The 96-well plate was
incubated at 37°C with 5% CO2 for 2 h, and absorbance
was measured at a wavelength of 450 nm using a microplate
reader.
Colony formation assay
The U2OS cells transfected with si-LINC00152 or with
si-NC were trypsinized into single-cell suspensions 48 h following
transfection and seeded into six-well plates. The cells were
incubated for ~2 weeks and then fixed with 4% paraformaldehyde for
15 min at room temperature. Colonies were photographed using a
light microscope and counted from three randomly selected fields
(magnification ×4).
Flow cytometry analysis
The apoptosis levels of each the U2OS cells sample
was determined 48 h after transfection. U2OS cells were stained
with annexin V-fluorescein isothiocyanate (FITC) and propidium
iodide (PI) using an annexin V-FITC/PI apoptosis detection kit
(Becton, Dickinson and Company, Franklin Lakes, NJ, USA). For cell
cycle analysis, cells were analyzed using a Cycletest Plus DNA
Reagent kit (Becton, Dickinson and Company). The kits were used
according to the manufacturer's protocol. Following 15 min of
incubation with Cycletest Plus DNA Reagent kit, cells were examined
using FACSCalibur flow cytometry system (FACScan; BD Biosciences,
Franklin Lakes, NJ, USA).
Luciferase reporter assay
U2OS cells were seeded into a 96-well plate for 24 h
and subsequently co-transfected with: pmirGLO-LINC00152-wild-type
(WT) and miR-NC; pmirGLO-LINC00152-WT and miR-193b-3p;
pmirGLO-LINC00152-mutant (MT) and miR-NC; or pmirGLO-LINC00152-MT
and miR-193b-3p (Promega Corporation, Madison, WI, USA), using
Lipofectamine® 2000, respectively. Following 48 h at
37°C, Firefly and Renilla luciferase activities were
measured using the Dual-Luciferase Reporter assay system (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
protocol. Firefly luciferase activity was normalized to
Renilla luciferase activity.
Statistical analysis
SPSS software (version 17; SPSS, Inc., Chicago, IL,
USA) was used for statistical analysis throughout the study.
StarBase software (version 2.0; http://starbase.sysu.edu.cn/) was used for
bioinformatics analyses in the study. All quantitative results are
presented as the mean ± standard deviation. The independent-samples
t-test was used for comparisons between two groups. Comparisons
among multiple groups were performed by one-way analysis of
variance followed by Bonferroni's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Knockdown of LINC00152 inhibits cell
proliferation, and induces G0/G1 cell cycle arrest and apoptosis in
osteosarcoma cells
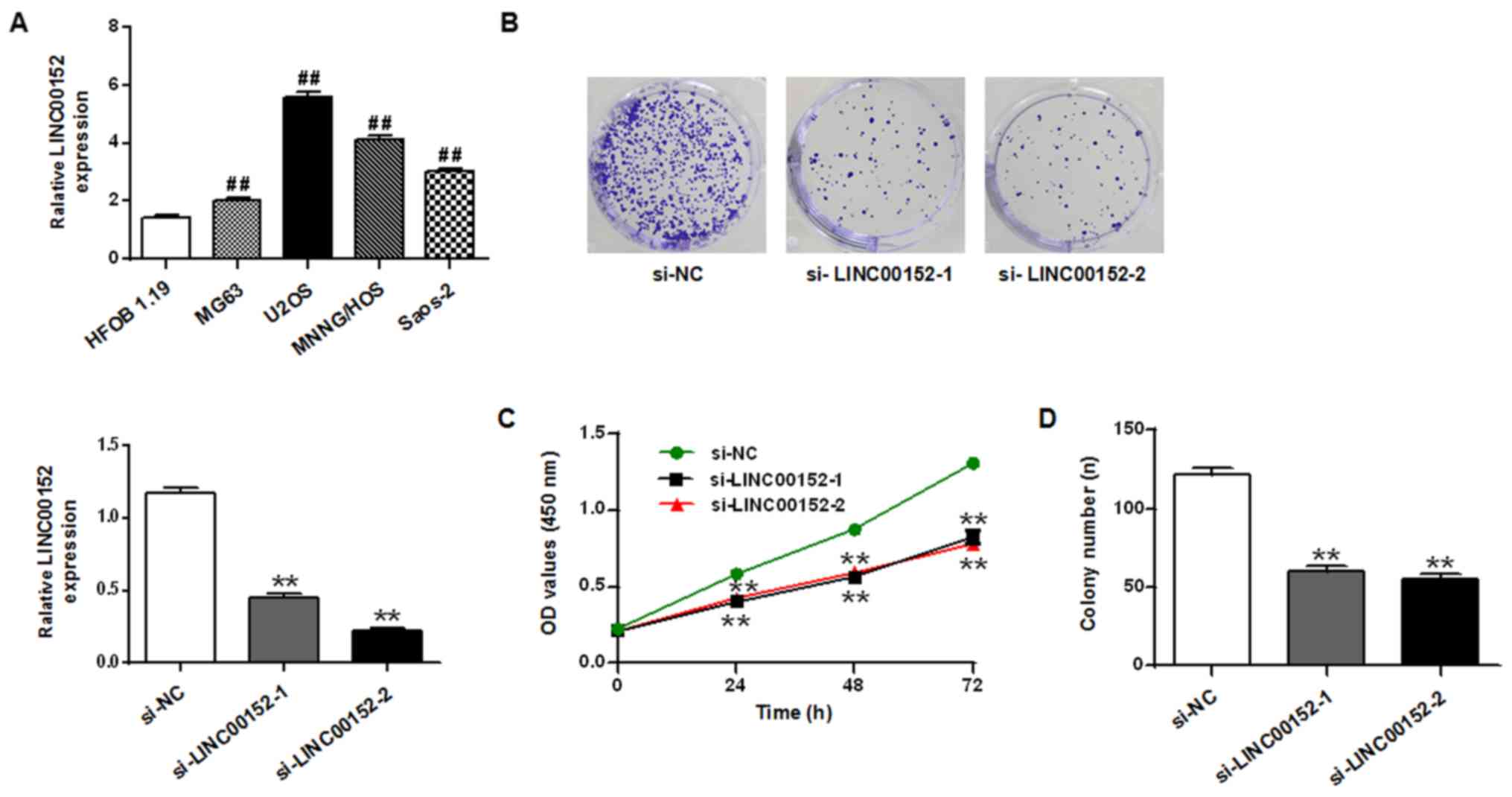
The expression level of LINC00152 in the
osteosarcoma cell lines U2OS, Saos-2, MG63 and MNNG/HOS was
quantified and compared with the expression level in the osteoblast
cell line HFOB 1.19 (P<0.01; Fig.
1). The expression levels of LINC00152 were significantly
upregulated in the four osteosarcoma cell lines compared with HFOB
1.19 cells, particularly in the U2OS cells. This was statistically
significant compared to the other osteosarcoma cell line.
The biological function of LINC00152 in osteosarcoma
cells was subsequently investigated. Two siRNAs targeting LINC00152
were used in the knockdown experiments in U2OS cells. Transfecting
with either one of the siRNAs suppressed the expression of
LINC00152 compared with the si-NC. Compared to si-NC group, the
relative LINC00152 expression of si-LINC00152-1 and si-LINC00152-2
groups were decreased (P<0.01; Fig.
1A). Results from the CCK-8 assay revealed that si-LINC00152-1
and si-LINC00152-2 significantly suppressed cell proliferation in
U2OS cells compared with the si-NC group (P<0.01; Fig. 1C). Similarly, the colony formation
assay revealed that U2OS cells transfected with si-LINC00152-1 and
si-LINC00152-2 exhibited significantly fewer colonies compared with
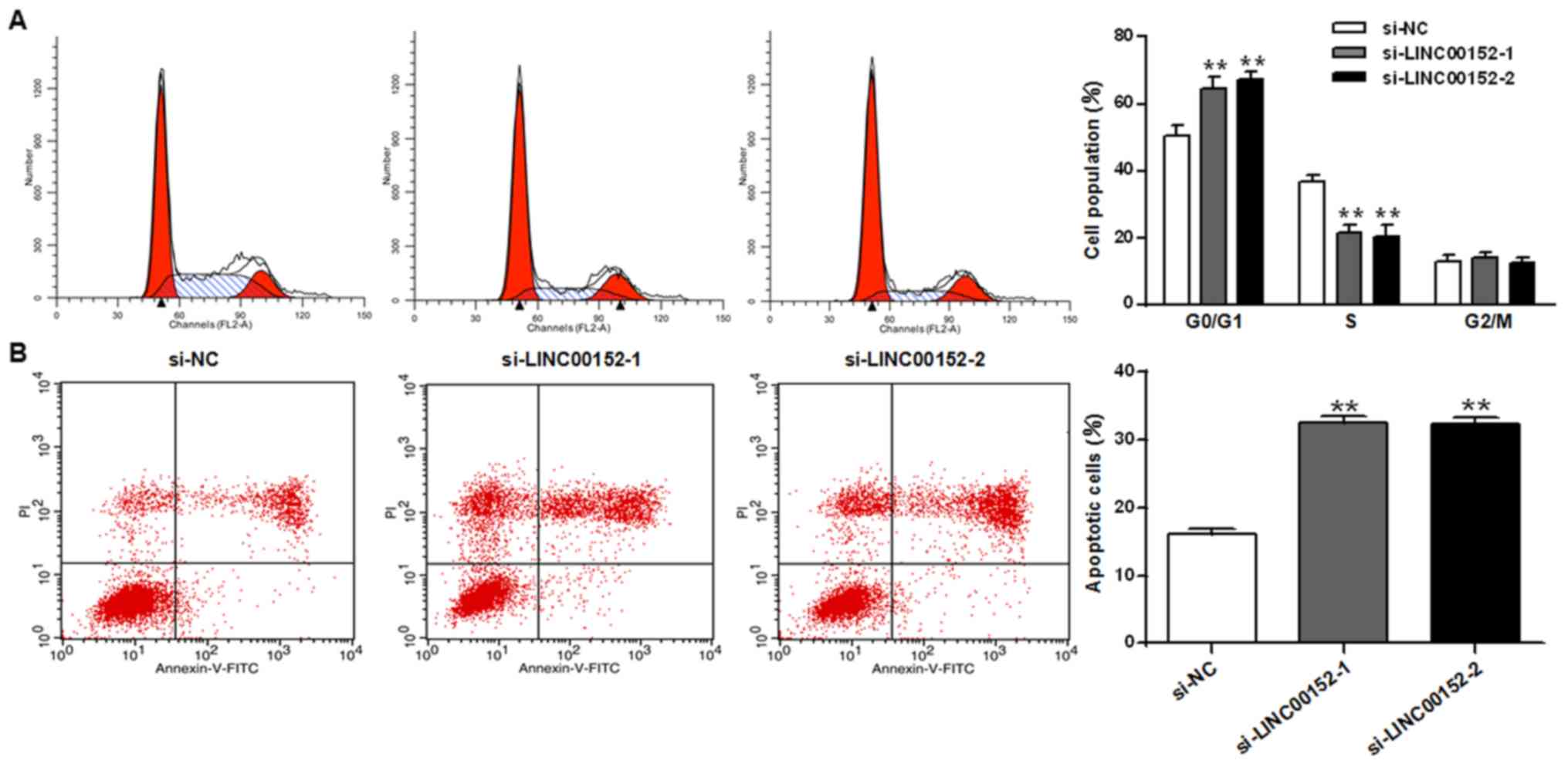
those transfected with si-NC (P<0.01; Fig. 1D). Flow cytometry analysis was
performed to determine whether LINC00152 impacted the growth of
osteosarcoma cells by affecting the cell cycle and apoptosis. The
results revealed that the LINC00152 knockdown significantly induced
G0/G1 cell cycle arrest (P<0.01; Fig.
2A) and increased apoptosis (P<0.01; Fig. 2B) in U2OS cells compared with cells
transfected with si-NC.
LINC00152 acts as a direct target of
miR-193b-3p
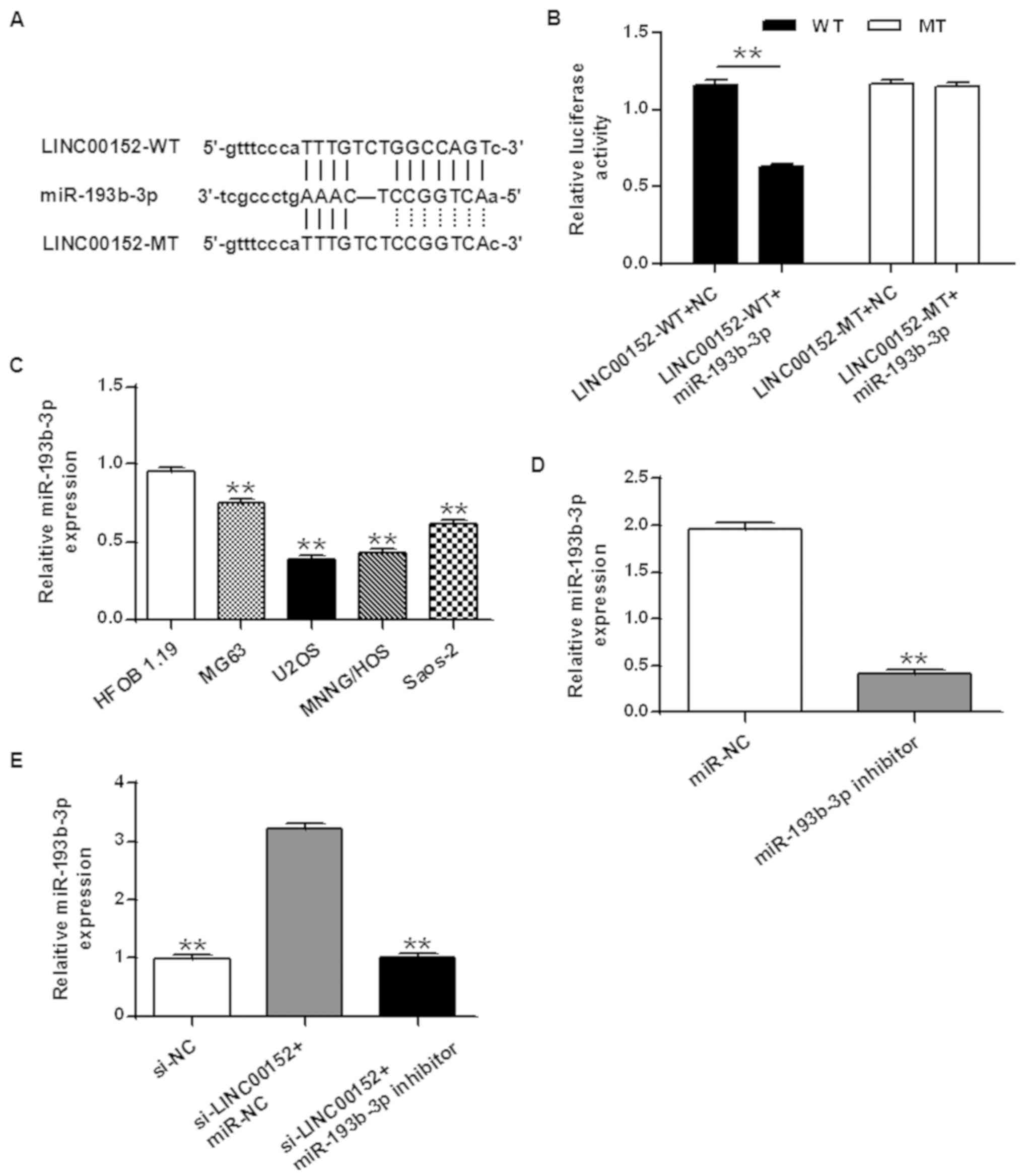
lncRNAs act as endogenous sponges for miRNAs and
affect their expression (35). In
order to elucidate the mechanisms underlying the effects of
LINC00152 on osteosarcoma cells, bioinformatics analyses (Fig. 3) were performed to identify miRNAs
which may bind to LINC00152. Bioinformatics analyses revealed that
LINC00152 contains a binding site for miR-193b-3p (Fig. 3A). A dual-luciferase assay was
performed to confirm the binding of miR-193b-3p to LINC00152. The
results obtained revealed that reduced luciferase activity was
observed only when both the LINC00152-WT reporter vector and
miR-193b-3p were co-transfected (P<0.01 vs. LINC00152-WT
reporter vector + NC; Fig. 3B).
Co-transfecting the MT LINC00152 and miR-193-3p revealed no changes
in luciferase activity compared with the LINC00152-MT + NC. The
results obtained suggested that miR-193b-3p may exhibit
sequence-specific binding to LINC00152.
The expression levels of miR-193b-3p in the
osteosarcoma cell lines MG63, U2OS, MNNG/HOS and Saos-2, as well as
in the osteoblast cell line HFOB 1.19 were measured. The expression
levels of miR-193b-3p in all four osteosarcoma cell lines were
significantly lower compared with that in HFOB 1.19 (P<0.01;
Fig. 3C).
Loss-of-function experiments were performed and the
expression level of miR-193b-3p was monitored. The transfection
efficiency of the miR-193b-3p inhibitor was demonstrated by RT-qPCR
(P<0.01 vs. miR-NC; Fig. 3D).
Results revealed that knockdown of LINC00152 by transfecting with
si-LINC00152 increased the expression of miR-193b-3p (P<0.01 vs.
si-NC group); whereas co-transfecting si-LINC00152 and the
miR-193b-3p inhibitor decreased the expression of miR-193b-3p
compared with the si-LINC00152 + miR-NC group (Fig. 3E; P<0.01). The results obtained
suggested that LINC00152 decreased the expression of miR0193b-3p in
osteosarcoma cells, by acting as a sponge for sequence-specific
binding.
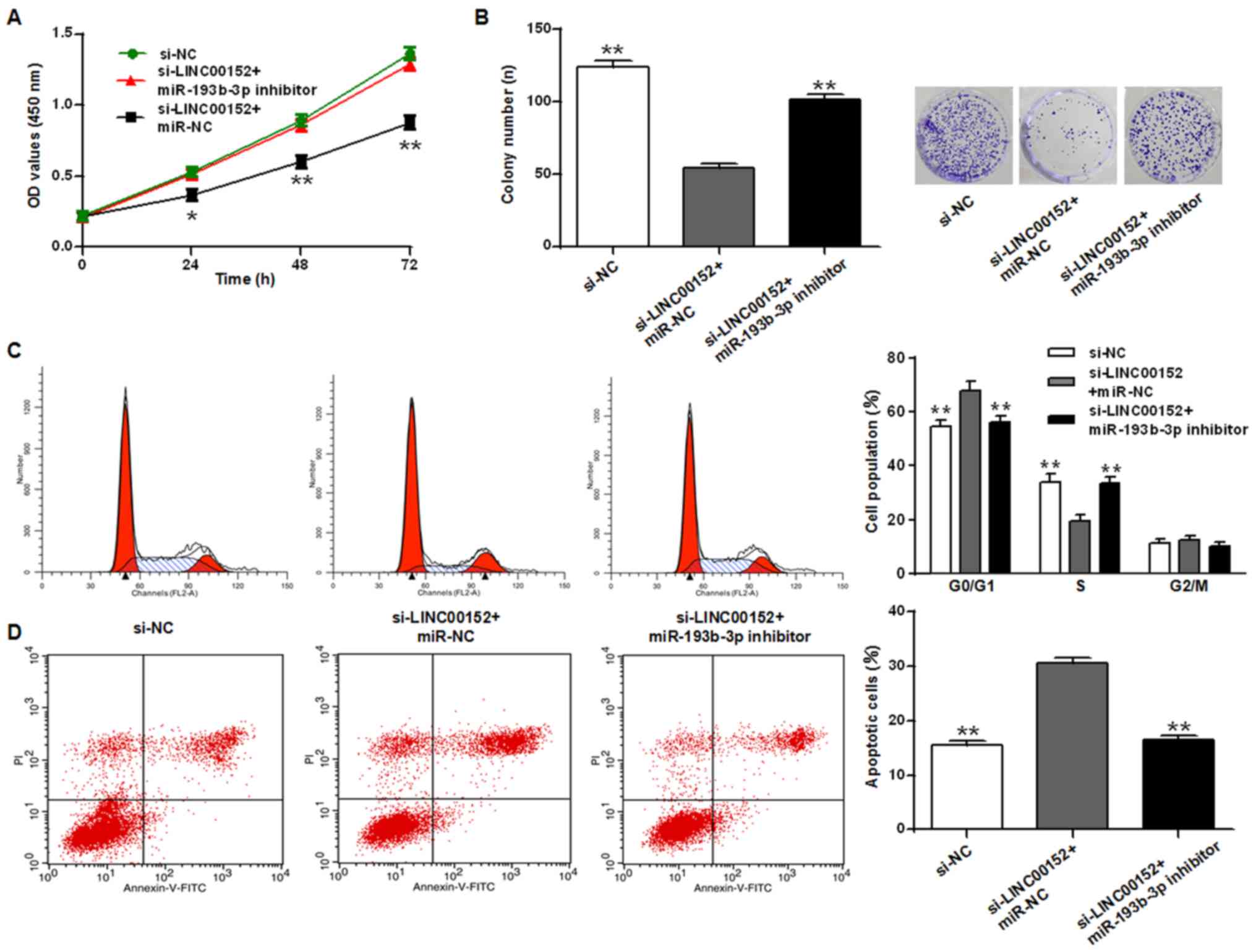
miR-193b-3p inhibitor rescues the
growth inhibition induced by LINC0012 knockdown in U2OS cells
The CCK-8 assay (Fig.
4A) presented different significance levels at 24, 48 and 72 h,
and colony formation assay (Fig. 4B)
revealed that the miR-193b-3p inhibitor reversed the growth
inhibition resulting from LINC00152 knockdown in osteosarcoma cells
(P<0.05 and P<0.01 vs. si-NC group, respectively). Similarly,
cell cycle analysis revealed that LINC00152 knockdown induced cycle
arrest at the G0/G1 phase and promoted apoptosis, while the
miR-193b-3p inhibitor reduced these effects (Fig. 4C and D; P<0.01). The miR-193b-3p
inhibitor may reverse the growth inhibition effect of LINC00152
knockdown in osteosarcoma cell lines, indicating the antagonistic
effects of LINC00152 and miR-193b-3p.
Discussion
The expression level of LINC00152 in osteosarcoma
cell lines was investigated, and significantly higher expression
levels of LINC00152 were observed in the osteosarcoma cell lines
compared with an osteoblast cell line. Two siRNAs targeting
LINC00152 were demonstrated to inhibit the expression of LINC00152
in U2OS cells. Knockdown of LINC00152 resulted in growth
inhibition, demonstrated by a reduction in cell proliferation and
colony formation. Knockdown of LINC00152 in U2OS cells
significantly induced G0/G1 cell cycle arrest and apoptosis. These
results suggested a positive association between LINC00152 and
osteosarcoma. LINC00152 was previously associated with colon
carcinomas, prostate cancer, leukemias, bladder and kidney cancer
(15–18).
The current study aimed to explore the mechanism
underlying the tumorigenic effect of LINC00152. LINC00152 acts as
an endogenous sponge by binding several miRNAs and inhibiting their
activity (35). The bioinformatics
analyses performed in the current study revealed that LINC00152
contains a putative binding site for miR-193b-3p. Previous studies
revealed that the miR-193b cluster demonstrated tumor-suppressive
effects by reducing cell proliferation, as well as by reducing the
total number of cells in the S-phase (36–38).
Furthermore, miR-193b acts as a tumor suppressor and is
epigenetically silenced in prostate cancer cells (37), and may serve a similar role in
osteosarcoma cells. miR-193b-3p was therefore selected as a
potential target of LINC00152 in the current study.
Sequence-specific binding of miR-193b-3p to LINC00152 was validated
by the dual-luciferase assay. Furthermore, the expression of
miR-193b-3p was significantly lower in the four osteosarcoma cell
lines, particularly in U2OS, compared with the osteoblast cell
line. LINC00152 may downregulate miR-193b-3p in osteosarcoma cells.
The expression of miR-193b-3p in osteosarcoma cells with LINC00152
knockdown was investigated. The results obtained indicated a
significantly increased miR-193b-3p expression resulting from
LINC00152 knockdown compared with si-NC, while co-transfecting with
a miR-193b-3p inhibitor reversed this effect.
A recent study performed a comprehensive deep
sequencing analysis on osteosarcoma cell samples to identify
differentially expressed RNAs. The authors revealed that 65 miRNAs,
233 lncRNAs and 1,405 mRNAs were differentially expressed in
osteosarcoma cells compared with normal cells, suggesting that
dysregulation of RNA signaling may be involved in the progression
of osteosarcoma (39). The present
study revealed a novel miRNA-lncRNA target pair that is
dysregulated in osteosarcoma cell lines. The overexpression of
LINC00152 downregulated miR-193b-3p in osteosarcoma cells,
potentially by acting as a competing endogenous RNA that binds to
miR-193b-3p. Cell cycle analysis revealed that this dysregulation
reduced G0/G1 arrest and promoted cell cycle progression, which
resulted in increased cell proliferation and decreased apoptosis. A
previous study demonstrated that LINC00152 may be potentially
downregulated by miR-376c-3p (40),
suggesting that ncRNAs form a complex signaling network that is
highly associated with a number of different diseases, such as
gastric and hepatocellular carcinoma.
The current study revealed that the dysregulation of
LINC00152 and miR-193b-3p may contribute to osteosarcoma; however,
further experiments using clinical samples are required to verify
these results. This may improve the understanding of epigenetic
regulation via LINC00152 and miR-193b-3p in osteosarcoma and may
provide insights for potential therapeutic treatment
strategies.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of Liaoning Provincial Department of Science and
Technology (grant no. 20170540383) and the Youth Foundation of
Liaoning Provincial Department of Education (grant no.
JYTQN201726).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PL, WH, YL and YW performed the experiments. PL
analyzed the data and was a major contributor in writing the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meyers PA and Gorlick R: Osteosarcoma.
Pediatr Clin North Am. 44:973–989. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Picci P: Osteosarcoma (osteogenic
sarcoma). Orphanet J Rare Dis. 2:62007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harrison DJ, Geller DS, Gill JD, Lewis VO
and Gorlick R: Current and future therapeutic approaches for
osteosarcoma. Expert Rev Anti Infect Ther. 18:39–50. 2017.
View Article : Google Scholar
|
|
5
|
Taft RJ, Pheasant M and Mattick JS: The
relationship between non-protein-coding DNA and eukaryotic
complexity. Bioessays. 29:288–299. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mattick JS: The central role of RNA in
human development and cognition. FEBS Lett. 585:1600–1616. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang JX, Rastetter RH and Wilhelm D:
Non-coding RNAs: An introduction. Adv Exp Med Biol. 886:13–32.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang Q, Zhang RW, Sui PC, He HT and Ding
L: Dysregulation of non-coding RNAs in gastric cancer. World
Gastroenterol J. 21:10956–10981. 2015. View Article : Google Scholar
|
|
9
|
Massillo C, Dalton GN, Paula L Farré PL,
De Luca P and De Siervi A: Implications of microRNA dysregulation
in the development of prostate cancer. Reproduction. 154:R81–R97.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song X, Luo X, Gao Q, Wang Y, Gao Q and
Long W: Dysregulation of lncRNAs in; lacenta and pathogenesis of
preeclampsia. Curr Drug Targets. 18:1165–1170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tan L, Yu JT and Tan L: Causes and
consequences of microRNA dysregulation in neurodegenerative
diseases. Mol Neurobiol. 51:1249–1262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blackshaw S, Harpavat S, Trimarchi J, Cai
L, Huang H, Kuo WP, Weber G, Lee K, Fraioli RE, Cho SH, et al:
Genomic analysis of mouse retinal development. PLoS Biol.
2:E2472004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dinger ME, Amaral PP, Mercer TR, Pang KC,
Bruce SJ, Gardiner BB, Askarian-Amiri ME, Ru K, Soldà G, Simons C,
et al: Long noncoding RNAs in mouse embryonic stem cell
pluripotency and differentiation. Genome Res. 18:1433–1445. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E and
Chang H: Functional demarcation of active and silent chromatin
domains in human HOX loci by noncoding RNAs. Cell. 129:1311–1323.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pibouin L, Villaudy J, Ferbus D, Muleris
M, Prospéri MT, Remvikos Y and Goubin G: Cloning of the mRNA of
overexpression in colon carcinoma-1: A sequence overexpressed in a
subset of colon carcinomas. Cancer Genet Cytogenet. 133:55–60.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu X, Ravindranath L, Tran N, Petrovics G
and Srivastava S: Regulation of apoptosis by a prostate-specific
and prostate cancer-associated noncoding gene, PCGEM1. DNA Cell
Biol. 25:135–141. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calin GA, Liu CG, Ferracin M, Hyslop T,
Spizzo R, Sevignani C, Fabbri M, Cimmino A, Lee EJ, Wojcik SE, et
al: Ultraconserved regions encoding ncRNAs are altered in human
leukemias and carcinomas. Cancer Cell. 12:215–229. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martens-Uzunova ES, Böttcher R, Croce CM,
Jenster G, Visakorpi T and Calin GA: Long noncoding RNA in
prostate, bladder, and kidney cancer. Eur Urol. 65:1140–1151. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luka B, Metka RG and Damjan G: Long
noncoding RNAs as biomarkers in cancer. Dis Markers. 2017:1–14.
2017.
|
|
20
|
Ji J, Tang J, Deng L, Xie Y, Jiang R, Li G
and Sun B: LINC00152 promotes proliferation in hepatocellular
carcinoma by targeting EpCAM via the mTOR signaling pathway.
Oncotarget. 6:42813–42824. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Wang X, Tang J, Jiang R, Zhang W, Ji
J and Sun B: HULC and Linc00152 act as novel biomarkers in
predicting diagnosis of hepatocellular carcinoma. Cell Physiol
Biochem. 37:687–696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou J, Zhi X, Wang L, Wang W, Li Z, Tang
J, Wang J, Zhang Q and Xu Z: Linc00152 promotes proliferation in
gastric cancer through the EGFR-dependent pathway. J Exp Clin
Cancer Res. 34:1352015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Liu J, Bai H, Dang Y, Lv P and Wu
S: Long intergenic non-coding RNA 00152 promotes renal cell
carcinoma progression by epigenetically suppressing P16 and
negatively regulates miR-205. Am J Cancer Res. 7:312–322.
2017.PubMed/NCBI
|
|
24
|
Cai Q, Wang ZQ, Wang SH, Li C, Zhu ZG,
Quan ZW and Zhang WJ: Upregulation of long non-coding RNA LINC00152
by SP1 contributes to gallbladder cancer cell growth and tumor
metastasis via PI3K/AKT pathway. Am J Transl Res. 8:4068–4081.
2016.PubMed/NCBI
|
|
25
|
Teng W, Qiu C, He Z, Wang G, Xue Y and Hui
X: Linc00152 suppresses apoptosis and promotes migration by
sponging miR-4767 in vascular endothelial cells. Oncotarget.
8:85014–85023. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu M, Xue Y, Zheng J, Liu X, Yu H, Liu L,
Li Z and Liu Y: Linc00152 promotes malignant progression of glioma
stem cells by regulating miR-103a-3p/FEZF1/CDC25A pathway. Mol
Cancer. 16:1102017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu Z, Dai J, Liao Y, Ma J and Zhou W:
Knockdown of long noncoding RNA LINC0000125 suppresses cellular
proliferation and invasion in glioma cells by regulating MiR-4775.
Oncol Res. 5:857–867. 2017.
|
|
28
|
Bian Z, Zhang J, Li M, Feng Y, Yao S, Song
M, Qi X, Fei B, Yin Y, Hua D and Huang Z: Long non-coding RNA
LINC00152 promotes cell proliferation, metastasis, and confers 5-FU
resistance in colorectal cancer by inhibiting miR-139-5p.
Oncogenesis. 6:3952017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cai Q, Wang Z, Wang S, Weng M, Zhou D, Li
C, Wang J, Chen E and Quan Z: Long non-coding RNA LINC00152
promotes gallbladder cancer metastasis and epithelial-mesenchymal
transition by regulating HIF-1α via miR-138. Open Biol.
7:1602472017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu J, Shuang Z, Zhao J, Tang H, Liu P,
Zhang L, Xie X and Xiao X: Linc00152 promotes tumorigenesis by
regulating DNMTs in triple-negative breast cancer. Biomed
Pharmacother. 97:1275–1281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yuan W, Sun Y, Liu L, Zhou B, Wang S and
Gu D: Circulating lncRNAs serve as diagnostic markers for
hepatocellular carcinoma. Cell Physiol Biochem. 44:125–132. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun K, Hu P and Xu F: LINC00152/miR-139-5p
regulates gastric cancer cell aerobic glycolysis by targeting
PRKAA1. Biomed Pharmacother. 97:1296–1302. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma P, Wang H, Sun J, Liu H, Zheng C, Zhou
X and Lu Z: LINC00152 promotes cell cycle progression in
hepatocellular carcinoma via miR-193a/b-3p/CCND1 axis. Cell Cycle.
17:974–984. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Militello G, Weirick T, John D, Döring C,
Dimmeler S and Uchida S: Screening and validation of lncRNAs and
circRNAs as miRNA sponges. Brief Bioinform. 1:780–788. 2016.
|
|
36
|
Mets E, Van der Meulen J, Van Peer G,
Boice M, Mestdagh P, Van de Walle I, Lammens T, Goossens S, De
Moerloose B, Benoit Y, et al: MicroRNA-193b-3p acts as a tumor
suppressor by targeting the MYB oncogene in T-cell acute
lymphoblastic leukemia. Leukemia. 29:798–806. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rauhala HE, Jalava SE, Isotalo J, Bracken
H, Lehmusvaara S, Tammela TL, Oja H and Visakorpi T: miR-193b is an
epigenetically regulated putative tumor suppressor in prostate
cancer. Int J Cancer. 127:1363–1372. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gastaldi C, Bertero T, Xu N,
Bourget-Ponzio I, Lebrigand K, Fourre S, Popa A, Cardot-Leccia N,
Meneguzzi G, Sonkoly E, et al: MiR-193b/365a cluster controls
progression of epidermal squamous cell carcinoma. Carcinogenesis.
35:1110–1120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xie L, Yao Z, Zhang Y, Li D, Hu F, Liao Y,
Zhou L, Zhou Y, Huang Z and He Z: Deep RNA sequencing reveals the
dynamic regulation of miRNA, lncRNAs, and mRNAs in osteosarcoma
tumorigenesis and pulmonary metastasis. Cell Death Dis. 9:7722018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang YH, Fu J, Zhang ZJ, Ge CC and Yi Y:
LncRNA-LINC00152 down-regulated by miR-376c-3p restricts viability
and promotes apoptosis of colorectal cancer cells. Am J Transl Res.
8:5286–5297. 2016.PubMed/NCBI
|