Introduction
Prostate cancer is the most frequently diagnosed
type of cancer in men and the second leading cause of
cancer-associated mortality in Western countries (1). In 2013, the incidence rate of prostate
cancer in China was ~9.9/100,000, ranking as the sixth leading
cause of malignancy in men (2,3). Due to
changes in lifestyle and dietary habits, the incidence rate of
prostate cancer has risen significantly in recent years (2,3).
Clinically significant prostate cancer is rarely diagnosed under
the age of 40 years, with a steady rise in incidence and prevalence
after 50 years of age (4,5). Classical treatments for prostate cancer
include surgical resection, radiotherapy, chemotherapy and
endocrine therapy (6). However, the
efficacy of these therapies remains unsatisfactory (6,7).
Immunotherapy may be another potential treatment modality (8), which requires further research. It is
necessary to identify novel prostate cancer regulators for the
diagnosis and treatment of prostate cancer.
To date, the underlying molecular mechanism of
prostate cancer remains to be fully elucidated. Abnormal activation
of androgen receptor is currently considered as the most important
mechanism of prostate cancer (7).
Abnormalities in other signaling molecules, such as overexpression
of transcription factors and oncogenes, and inactivation of tumor
suppressors also participate in different stages of prostate cancer
development (9). Metabolic
reprogramming is one of the important molecular events in the
tumorigenesis and progression of prostate cancer (10). In our previous studies, ankyrin
repeat domain 22 (ANKRD22) was found to be a novel
metabolic-reprogramming molecule, which interacts with the
gatekeeper enzyme of aerobic glycolysis, phosphoinositide-dependent
kinase-1 and multiple subunits of the ATP synthase complex (Pan
et al, unpublished data). ANKRD22 is an ankyrin repeat
protein with four copies of the ankyrin motif. It belongs to the
ankyrin repeat (ANK) family and contains 191 amino acids with a
molecular weight of 21,849 Daltons (11). A number of previous studies have
demonstrated that abnormal ANK family protein function is
associated with disease, including hemolytic anemia, arrhythmia and
autism (12–14). ANK proteins also have important roles
in cancer development and progression (15,16). For
example, ANKRD1 induces renal cancer cell proliferation by
regulation of downstream miRNA (15). By contrast, ANKRD12 may exert
antitumor effects in colorectal cancer by inhibiting liver
metastasis (16). ANKRD22 expression
has been detected in esophageal, gastric and colorectal epithelial
cells, and in ovarian epithelial cells (17). However, the exact function of ANKRD22
in prostate cancer remains unclear.
The present study aimed to analyze the organelle
distribution and expression status of ANKRD22 in prostate cancer
and the function of ANKRD22 in prostate cancer progression.
Materials and methods
Clinical tissue specimens
A total of 11 patients with prostate cancer (age
range, 56–76 years) and 4 male patients with gastric cancer (age
range, 54–68 years) were recruited between August 2017 and March
2018 at The Second Affiliated Hospital of Zhejiang University
(Hangzhou, China). None of patients with prostate cancer had
received hormone therapy, radiotherapy or chemotherapy prior to
radical prostatectomy. Tumor and adjacent non-cancerous tissues
located at least 1 cm from the cancer margin were collected during
the surgery. The tissues were fixed by 10% formaldehyde (Juhua
Group Corporation) at room temperature for 6 h and embedded in
paraffin following dehydration. Subsequently, the paraffin-embedded
tissues were sliced into 5–7 µm sections. The hematoxylin and eosin
(H&E) staining slides of prostate and gastric cancer tissues
were reviewed under light-microscope at ×100 magnification and
diagnoses were verified by an experienced pathologist. The present
study was approved by the Ethics Committee of The Second Affiliated
Hospital of Zhejiang University, and all patients provided written
informed consent.
Bioinformatics analysis
The Cancer Genome Atlas (TCGA) database (https://www.cbioportal.org/study/clinicalData?id=prad_tcga)
and Gene Set Enrichment Analysis (GSEA) of 636 samples was analyzed
by Hope Biotech Company (Hangzhou, China). Human prostate tissue
gene expression profile data sets with corresponding clinical and
follow-up information were downloaded from TCGA database. The
detailed GSEA protocol was obtained from the Broad Institute GSEA
website (www.broad.mit.edu/gsea). The GSEA software v2.0.13
(http://software.broadinstitute.org/gsea/downloads.jsp)
was run using the JAVA 7.0 platform. The dataset and phenotype
label files were created and loaded into the GSEA software. ANKRD22
mRNA level was quantiled into Q0-Q3. Patients
in Q2 and Q3 were considered to exhibit high
ANKRD22 expression, whereas patients in Q0 and
Q1 exhibited low expression of ANKRD22 mRNA.
Expression and purification of
recombinant human ANKRD22 protein
The ANKRD22 open reading frame DNA fragment in
ANKRD22/pCMV5-XL5 (OriGene Technologies, Inc.) was amplified by DNA
polymerase (Takara Bio, Inc.). The primers used were as follows:
ANKRD22 forward, 5′-AGTTCTCCATGGAGATGGGAATCCTATACTCTGAGCCCAT-3′ and
reverse, 5′-ACTGAGTGGATCTTACAATGCTTTCCTTAGCATTAATTC-3′. The
following thermocycling conditions were used: Initial denaturation
at 95°C for 30 sec; 35 cycles of 95°C for 10 sec, 55°C for 15 sec
and 72°C for 60 sec. pET-42a vectors (Novagen, Merck KGaA) were
initially digested using NcoI and BamHI
endonucleases, respectively, and mixed at a ratio of 9:1
ANKRD22:pET-42a with T4 ligase at 16°C overnight. The ligation
product was transformed into freshly prepared DH5α E. coli
competent cells (Sangon Biotech Co., Ltd.) and DNA sequencing was
performed by Sanger to determine if transfection had been
successful. Once ANKRD22 was successfully subcloned into pET-42a
(ANKRD22 fused with glutathione S-transferase- and histidine-tags),
ANKRD22/pET-42a plasmids were transformed into Rosetta DE3
competent cells (Novagen; Merck KGaA). The recombinant proteins
were induced using isopropyl-β-D-thiogalactoside (Sangon Biotech
Co., Ltd.) and the inclusion bodies (IB) were collected and
renatured using a Protein Refolding kit (Novagen; Merck KGaA).
Briefly, the cells were resuspended with IB wash buffer (Novagen;
Merck KGaA) and incubated with 100 µg/ml lysozyme (Sangon Biotech
Co., Ltd.) at 30°C for 15 min. Sonication (frequency, 20–25 kHz; 90
W for 10 sec and 10 cycles of 30 sec in an ice bath) was performed
until the solution was not viscous. The lysate was subsequently
centrifuged at 10,000 × g at 4°C for 10 min. The pellet was washed
with 1X IB wash buffer twice and the supernatant was discarded. The
inclusion body pellet was then added to an appropriate amount of
dissolving buffer [50 ml of dissolving buffer consisting of 5 ml
10X IB Solubilization Buffer, 0.5 ml 30% N-lauroylsarcosine, 50 µl
1 M dithiothreitol (DTT) and 44.45 ml deionized water] to make a
final inclusion body concentration of 10–20 mg/ml. The mixture was
fully combined, incubated at room temperature for 15 min and
centrifuged at 10,000 × g at 4°C for 10 min. The supernatant was
collected and dialyzed using 20 mM Tris-HCl buffer (dialysis
buffer; pH 8.5) with 0.01% DTT solution twice (3 h at 4°C each
time), using dialysis buffer twice (3 h at 4°C each time) and using
dialysis buffer with 1 mM reduced glutathione and 0.2 mM oxidized
glutathione overnight at 4°C. The renatured protein was purified
through a Ni2+ column (Bio-Rad Laboratories, Inc.) and
frozen following dialysis with 30% glycerol at −20°C for long-term
storage (1 year).
Establishment of 293T cells
overexpressing ANKRD22
Recombinant ANKRD22-expressing lentivirus particles
(100 µl/ml; multiplicity of infection=100; Cyagen Inc.) were used
to infect 293T cells (Cell Bank of Shanghai Branch of the Chinese
Academy of Sciences) which were maintained in high-glucose DMEM
(Corning, Inc.) supplemented with 10% fetal bovine serum (Corning,
Inc.) and 100 µg/ml gentamicin (Sangon Biotech Co., Ltd.) at 37°C
in a humidified environment with 5% CO2. Following
incubation for 48 h, cells were passaged and transferred to a new
six-well plate and screened with 5 µg/ml puromycin for 7–10 days.
After propagation, 293T cells with high expression levels of
ANKRD22 were digested with 0.25% trypsin solution (Genom
Biotechnology Co. Ltd.) at room temperature for 5 min and collected
by centrifugation at 1,000 × g at room temperature for 5 min and
washed twice with PBS. Following aspiration of the PBS, 500 µl
ice-cold radioimmunoprecipitation assay lysis buffer (pH 7.5; 50 mM
Tris-HCl buffer containing 150 mM NaCl, 1 mM EDTA, 0.25%
deoxycholate acid and 1% NP-40 and 0.25% deoxycholate sodium) was
added, and the solution was mixed by pipetting and gentle
agitation. The solution was lysed on ice for 15 min, centrifuged at
13,000 × g at 4°C for 10 min, and the supernatant was aspirated for
western blot analysis.
Preparation of mouse anti-human
ANKRD22 monoclonal antibodies
A total of 32 female BALB/C mice (age, 6 weeks;
weight, 16–18 g) were purchased from Shanghai Laboratory Animal
Research Center and maintained between February 2017 and July 2017
in the standardized specific pathogen-free-grade experimental
animal center of Zhejiang Chinese Medical University authorized by
the government of Zhejiang province at 22±2°C with 40–70% humidity,
noise <50 dB, light intensity 150–200 Lx, 12:12 h light/dark
cycle, sterile ultrapure water and sterile complete formulated
nutrition granulated feed. The health and behavior of each mouse
was monitored daily. Recombinant ANKRD22 was used as an immunogen
to immunize the mice. The mice were placed in a 10 liter 99%
CO2 chamber (3 l/min) for 5–10 min and were monitored
for signs of life including heartbeat, breath and muscular tension
for 5 min prior to verification of death. All procedures complied
with the Laboratory Animal Guidelines for Ethical Review of Animal
Welfare in China and the animal research protocol was approved by
the Animal Care and Welfare Committee of Zhejiang Chinese Medical
University. Lysate from overexpressed ANKRD22 293T cells was used
for hybridoma screening using western blot analysis. During the
course of antibody production, mice immunization and hybridoma
production was performed by Hangzhou HuaAn Biotechnology Co., Ltd.
Following three screening cycles, four hybridoma clones were
obtained: 1A8, 1F3, 2A4 and 2E3.
Western blot analysis
A total of 20 µg protein lysate per lane, as
determined by BCA method (Bio-Rad Laboratories, Inc.), was
separated using 12% SDS-PAGE and loaded onto a nitrocellulose
membrane. The membrane with ANKRD22 protein was cut into strips,
blocked with 5% skimmed milk powder at room temperature for 1 h and
washed with PBS mixed with 0.05% Tween-20 (PBST) five times (3 min
each time). Each membrane strip was then incubated overnight with
the aforementioned mouse anti-ANKRD22 antibodies (1:500) at 4°C.
The strips were then incubated with a horseradish
peroxidase-labeled goat anti-mouse immunoglobulin G (IgG) (H+L)
secondary antibody (1:2,000; cat. no. 115-036-003; Jackson
Immunoresearch Laboratories, Inc.) for 1 h at room temperature. The
protein bands were visualized using an enhanced chemiluminescence
reagent (Pierce; Thermo Fisher Scientific, Inc.) for 1 min, and the
signal was detected using a digital imaging system (LI-COR
Biosciences).
ELISA
For determination of the specificity
of the monoclonal antibodies
A competitive-ELISA was used for validation of the
specificity of the monoclonal antibodies. Briefly, 10 µg/ml
aforementioned recombinant ANKRD22 protein was coated onto
microplate wells and incubated at 4°C overnight. A total of 5 µg/ml
truncated ANKRD22 (2nd-89th) peptide (Pan et al, unpublished
data) was used as a blocking protein and incubated with the
aforementioned four clones of anti-ANKRD22 monoclonal antibody
(1A8, 1F3, 2A4 and 2E3; 1 µg/ml) for 30 min at room temperature
prior to being added to the wells. The ANKRD22 blocking peptide was
replaced with 1% calf bovine serum (Hangzhou Tianhang Biotechnology
Co., Ltd.) as a negative control.
For determination of ANKRD22 protein
in tissue lysates
A total of 10 µg/ml aforementioned recombinant
ANKRD22 protein was coated onto microplate wells and incubated at
4°C overnight. Subsequently, 60 µl lysates from four gastric cancer
tissues or recombinant ANKRD22 protein (10, 50, 200, 500 and 1,000
ng/ml) were mixed with 60 µl anti-ANKRD22 monoclonal antibody
(clone 1A8; 1 µg/ml) for 60 min at room temperature prior to being
added to the wells. The mixed solution was added to the wells and
incubated at 37°C for 60 min.
Subsequently, the microplates were incubated at 37°C
for 2 h. After washing three times with PBST, 100 µl horseradish
peroxidase-labeled goat anti-mouse IgG (1:2,000; cat. no.
115-036-003, Jackson Immunoresearch Laboratories, Inc.) was added
to each well and incubated at 37°C for 1 h. The samples were
subsequently washed three times with PBST, and 100 µl
3,3′5,5′-tetramethylbenzidine (TMB; Sigma-Aldrich; Merck KGaA)
substrate solution was added to each well, incubated at room
temperature for 10 min, and terminated with 1.8 M sulfuric acid (50
µl/well). Absorbance was measured at 450 nm using a microplate
reader.
Reverse transcription-quantitative PCR
(RT-qPCR)
A total of four fresh human gastric cancer tissues
obtained by gastrectomy were used for total RNA extraction using
TRIzol® reagent (Invitrogen, Thermo Fisher Scientific,
Inc.). 1.0 µg extracted RNA was quantified and reverse-transcribed
to cDNA by incubation at 42°C for 60 min using a PrimeScript™ RT
reagent (Takara Bio, Inc.). Subsequently, ANKRD22 was quantified
using qPCR with the Premix Ex Taq™ kit (Takara Bio, Inc.) on a CFX
connect system (Bio-Rad Laboratories, Inc.). The primers and probes
used were as follows: ANKRD22 forward, 5′-CCAGCTTGGACTTCTAGGGA-3′
and reverse, 5′-GGCAGATGGGCTCAGAGTAT-3′; probe,
5′-FAM-TCCCATGCTGGTCCTTCACAGG-TAMRA-3′; GAPDH forward,
5′-ATCATCCCTGCCTCTACTGG-3′ and reverse, 5′-GTCAGGTCCACCACTGACAC-3′;
and probe, 5′-FAM-ACCTTGCCCACAGCCTTGGC-TAMRA-3′ (all from Sangon
Biotech Co., Ltd.). The following thermocycling conditions were
used: Initial denaturation at 95°C for 30; 40 cycles of 95°C for 5
sec and 60°C for 30 sec. GAPDH was used as an internal reference.
Relative ANKRD mRNA expression was quantified using the
2−ΔΔCq method (18).
Immunohistochemical staining
Immunohistochemical staining was performed using
standard methods. Paraffin-embedded tissues were sliced into 5–7 µm
sections. Following hydration, citric acid buffer was used at 95°C
for 10 min for antigen retrieval. The sections were blocked at room
temperature with 10% normal goat serum (Fuzhou Maxim Biotechnology
Co. Ltd.) and incubated with the mouse anti-ANKRD22 primary
antibody (clone 1A8; 1:200) at room temperature for 1 h.
Subsequently, the sections were washed 5 times in PBST and
incubated for 30 min at room temperature with ready-to-use
streptavidin-peroxidase-labeled goat anti-mouse IgG solution (cat.
no. KIT-9710, Fuzhou Maxim Biotechnology Co. Ltd.). The sections
were developed with 3,3′-diaminobenzidine (cat. no. KIT-9710,
Fuzhou Maxim Biotechnology Co. Ltd.) at room temperature for 15
min, counterstained with hematoxylin for 1 min, and examined by
light-microscopy at ×400 magnification.
Statistical analysis
Data were analyzed using SPSS 22.0 software (IBM
Corp.). The data from TCGA database were imported into an Excel
spreadsheet for normalization, and ANKRD22 mRNA expression was
subsequently retrieved from the dataset. Student's t-test was used
for the comparison of ANKRD22 mRNA expression between prostate
cancer and normal tissues. Kaplan-Meier analysis was used to
investigate the association between ANKRD22 level and survival of
prostate cancer. ANOVA with the Least Significance Difference (LSD)
post hoc test was used to analyze the ANKRD22 expression and
pathological parameters of prostate cancer. P<0.05 was
considered to indicate a statistically significant difference.
Results
Preparation of mouse anti-human
ANKRD22 monoclonal antibodies
To prepare mouse anti-human ANKRD22 monoclonal
antibodies, recombinant human ANKRD22 protein was produced from
E. coli using the pET42a expression system. Human ANKRD22
protein exists in the form of inclusion bodies in E. coli
bacteria. Following renaturing of the inclusion bodies,
hydrosoluble ANKRD22 protein was then purified using
Ni2+ affinity columns (Fig.
1A). BALB/c mice were immunized with the purified recombinant
protein. A total of 93 candidate hybridoma cells were obtained and
were further screened using lysates of 293T cells with
overexpression of ANKRD22 using ELISA and western blot analysis. A
total of four anti-ANKRD22 monoclonal antibodies were identified
(Fig. 1B). Furthermore, all these
antibodies could be almost completely blocked by the 2nd to 89th
ANKRD22 peptide, indicating the high specification of anti-ANKRD22
antibodies (Fig. 1C). Finally, the
association between ANKRD22 protein and ANKRD22 mRNA from gastric
cancer tissues was analyzed. Protein levels determined using ELISA
were in accordance with levels of ANKRD22mRNA detected using
RT-qPCR, thus confirming affinity between these antibodies and
human ANKRD22 protein (Fig. 1D).
 | Figure 1.Preparation and identification of
anti-ANKRD22 monoclonal antibodies. (A) Preparation of recombinant
ANKRD22 protein in prokaryotic cells. M, Marker; lane 1 and 2,
whole lysate of E. coli bacteria transformed with
ANKRD22/pET-42a plasmids induced using isopropyl
β-D-1-thiogalactopyranoside; lane 3, bacterial lysate supernatant;
lane 4, bacterial precipitates; lane 5, partially purified
recombinant ANKRD22 protein by Ni2+ affinity
chromatography following renaturation. The SDS-PAGE gel was stained
using Coomassie brilliant blue dye. (B) ANKRD22 protein was
detected using western blot analysis with different anti-ANKRD22
monoclonal antibodies. Lysates of 293T cells with overexpression of
ANKRD22 were used as test samples. A total of 4 clones of ANKRD22
monoclonal antibodies (clone no. 1A8, 1B1, 2E4 and 1F3) were
successfully verified. (C) Determination of the specificity of the
monoclonal antibodies was validated by competitive-ELISA. The
results showed that activity of each monoclonal antibody could be
blocked by ANKRD22 antigen. Each experiment was repeated 6 times.
*P<0.01. (D) Analysis of the association between ANKRD22 protein
and ANKRD22 mRNA. Comparison of the relative levels of ANKRD22
determined using ELISA and RT-qPCR in four gastric cancer tissue
samples (S1-4). Clone 1A8 was used as the primary antibody in
ELISA. GAPDH was used as the internal inference in RT-qPCR. The
data were normalized to ANKRD22 mRNA of S1. ANKRD22, ankyrin repeat
domain 22; GST, glutathione S-transferase; His, histidine; RT-qPCR,
reverse transcription-quantitative PCR. |
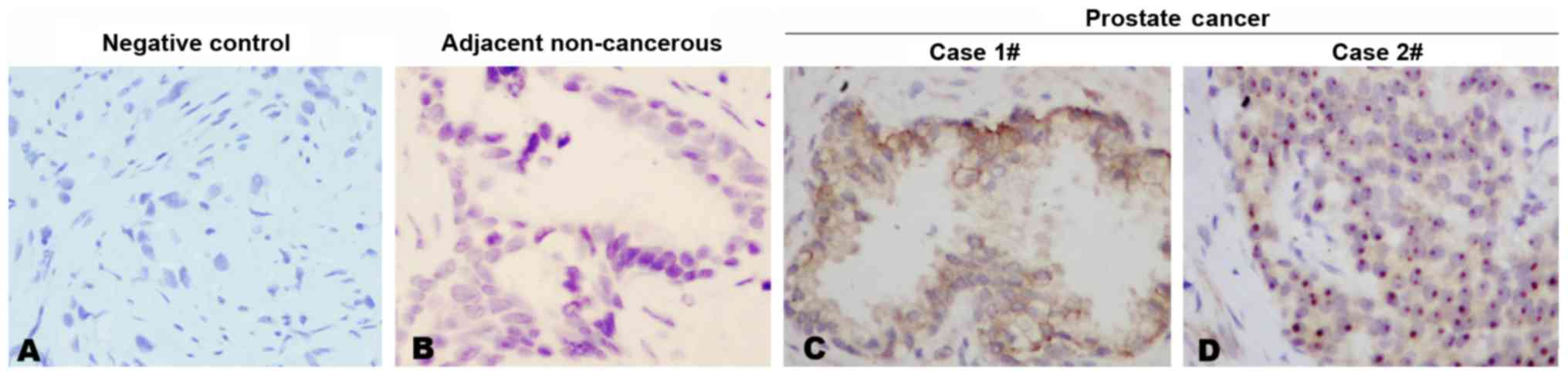
Expression of ANKRD22 in prostate
cancer
Using the generated anti-ANKRD22 monoclonal
antibodies, immunohistochemical staining was performed in prostate
cancer tissue. It was demonstrated that each anti-ANKRD22
monoclonal antibody exhibited a strong reaction with ANKRD22 in
prostate cancer tissues and each prostate cancer tissue sample was
stained positive for ANKRD22 in all 11 clinical samples. Four
clones exhibited similar IHC patterns. Expression of ANKRD22 in
prostate cancer was higher compared with that in adjacent
non-cancerous tissue (Fig. 2A-C).
ANKRD22 was located in the cytoplasm and nucleus of prostate cancer
cells (Fig. 2C and D). These results
indicated that ANKRD22 may be involved in the progression of
prostate cancer.
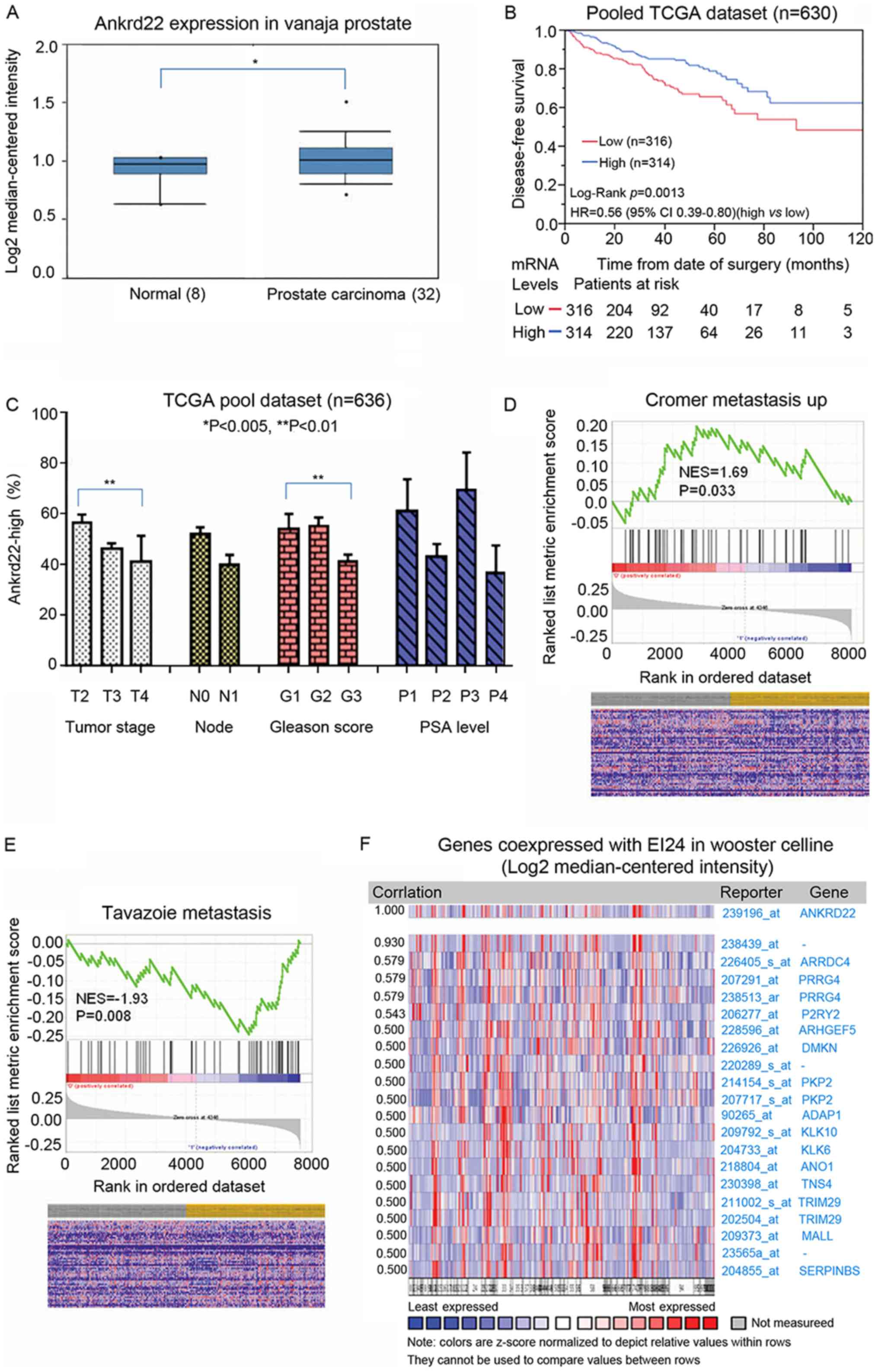
ANKRD22 is associated with prostate
cancer aggressiveness
Our previous study demonstrated that ANKRD22 is a
tumor suppressor in colorectal cancer (Pan et al,
unpublished data). As such, the role of ANKRD22 in the
aggressiveness of prostate cancer and the subsequent patient
outcomes were investigated. In accordance with the
immunohistochemical staining results, TCGA analysis demonstrated
that ANKRD22 was higher in prostate cancer compared with that in
normal prostate (Fig. 3A). Data from
TCGA datasets suggested that patients with high expression of
ANKRD22 exhibited significantly improved prognosis compared with
those with low expression (Fig. 3B),
thus suggesting that ANKRD22 is a good prognostic biomarker for
prostate cancer.
The association between ANKRD22 expression and
prostate cancer staging, lymph node metastasis, Gleason score and
prostate-specific antigen (PSA) level was then explored through
analysis of TCGA datasets. The proportion of patients with high
ANKRD22 expression in stage IV prostate cancer patients was
significantly lower compared with that in patients with stage II
prostate cancer, and a low Gleason score was associated with high
ANKRD22 expression (G1 vs. G3; P<0.01). However, no association
was observed between the proportion of ANKRD22-high patients and
lymph node metastasis or PSA level (Fig.
3C). Additionally, in GSEA, high expression of ANKRD22 was
negatively associated with prostate metastasis (metastatic vs.
non-metastatic prostate cancer) (Fig. 3D
and E) and ANKRD22 was significantly co-expressed with tumor
suppressor EI24 (Fig. 3F). These
results indicate that ANKRD22 is a good prognostic target for
prostate cancer and may be a tumor suppressor in prostate
cancer.
Discussion
To the best of our knowledge, the present study is
the first to demonstrate that the level of ANKRD22 has an inverse
association with prostate cancer progression. ANKRD22 may be
expressed in either the cytoplasm or nuclei of prostate cancer
cells, although the underlying molecular mechanism is not clear.
Furthermore, it was discovered that high-grade and high-stage
prostate cancer exhibited a significantly reduced ANKRD22
expression. Patients with higher ANKRD22 mRNA levels exhibited
longer disease-free survival post-prostatectomy.
ANKRD22 is an ankyrin repeat protein with four
copies of the ankyrin motif. Ankyrin motif is one of the most
common protein-protein interaction structural motifs, located in
>400 proteins (19). Each ankyrin
repeat is an L-shaped structure consisting of two inverted α
helices and a β hairpin, with a size of ~33 amino acid residues,
and forms a molecular connection scaffold structure with high
affinity (20). Unlike other
protein-protein binding motifs, ankyrin repeat motifs are primarily
classified based on their specific tertiary structures, which
determine the proteins they interact with and therefore their
specific biological functions. The ankyrin repeats are highly
conserved in evolution, although they interact with a diverse range
of proteins (21). This
protein-protein interaction is involved in the regulation of many
cellular physiological activities, such as transcriptional
regulation, cytoskeleton, ion transport, signal transduction,
inflammation and immunity, and tumorigenesis (22–24).
This suggests, therefore, that ANK family protein members, such as
ANKRD22, are involved in cancer.
ANKRD22 is expressed in normal digestive tract
epithelial and tumor cells. Yin et al (11) observed high expression of ANKRD22 in
non-small cell lung cancer cells, which is induced by upregulation
of transcription factor E2F1. Venner et al (25) observed an increase of ANKRD22
expression in macrophages in acute renal allograft rejection,
indicating that ANKRD22 is a negative regulator of the interferon
pathway similar to interferon-inducible protein AIM2 (26). Both oncogenic and tumor suppressor
functions have previously been reported in ANK proteins. For
example, the Notch pathway receptors (Notch 1–4) are ANK family
members that contain six ankyrin repeats (27). Activation of the Notch pathway
increases stemness and inhibits differentiation (28). Compared with non-stem cells, ANKRD28
is upregulated in acute myeloid leukemia stem cells (29). Apoptosis-stimulation of p53 protein
2, which contains four ankyrin repeats, was previously reported to
induce stemness and chemoresistance in hepatoma cells (30). By contrast, ankyrin repeat and SOCS
box protein 4 is highly expressed in the early stage of placental
development and mediates differentiation of embryonic stem cells
into the vascular system (31).
ANKRD3 is reported to be associated with differentiation of neural
stem cells (32). ANKRD6 recruits
casein kinase I isoform-ϵ to the β-catenin degradation complex and
inhibits β-catenin/transcription factor signaling (33). In the present study, the potential
role of ANKRD22 in prostate cancer as a tumor suppressor was
uncovered, similar to the study by Schoenborn et al
(34) where the copy number of
ANKRD22 was decreased in prostate cancer. By contrast, a recent
study demonstrated that ANKRD22 promoted progression of non-small
cell lung cancer (11). This
suggests that ANKRD22 may exhibit either pro- or antitumor function
depending on the origin of the tumor.
Taken together, these results indicate that ANKRD22
may be involved in the progression of prostate cancer. The
mechanisms of how ANKRD22 affects tumorigenesis of prostate cancer
remains to be defined. Identifying the targets of ANKRD22 and
biological functions of their interaction in prostate cancer
progression remains to be further discussed.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Natural Science of Foundation of Zhejiang Province (grant. no.
Y17H050034).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YQ and HW contributed to the conception and design
of the present study. HW also provided administrative support. YQ,
SY and TP performed experiments. YQ, LY and JL were responsible for
the collection and assembly of data. YQ, LY and YZ performed data
analysis and interpretation. All authors contributed to the writing
of the manuscript and its final approval.
Ethics approval and consent to
participate
The research protocol was approved by the Second
Affiliated Hospital of Zhejiang University Ethics Committee, and
all patients provided written informed consent. The animal research
protocol was approved by the Animal Care and Welfare Committee of
Zhejiang Chinese Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Zhang S, Zeng H, Xia C,
Zuo T, Yang Z, Zou X and He J: Cancer incidence and mortality in
China 2013. Cancer Lett. 401:63–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scher HI, Solo K, Valant J, Todd MB and
Mehra M: Prevalence of prostate cancer clinical states and
mortality in the United States: Estimates using a dynamic
progression model. PLoS One. 10:e01394402015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye D and Zhu Y: Epidemiology of prostate
cancer in China: An overview and clinical implication. Zhonghua Wai
Ke Za Zhi. 53:249–252. 2015.(In Chinese). PubMed/NCBI
|
|
6
|
Attard G, Parker C, Eeles RA, Schröder F,
Tomlins SA, Tannock I, Drake CG and de Bono JS: Prostate cancer.
Lancet. 387:70–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Watson PA, Arora VK and Sawyers CL:
Emerging mechanisms of resistance to androgen receptor inhibitors
in prostate cancer. Nat Rev Cancer. 15:701–711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schepisi G, Farolfi A, Conteduca V,
Martignano F, De Lisi D, Ravaglia G, Rossi L, Menna C, Bellia SR,
Barone D, et al: Immunotherapy for prostate cancer: Where we are
headed. Int J Mol Sci. 18:E26272017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Taylor BS, Schultz N, Hieronymus H,
Gopalan A, Xiao Y, Carver BS, Arora VK, Kaushik P, Cerami E, Reva
B, et al: Integrative genomic profiling of human prostate cancer.
Cancer Cell. 18:11–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schrecengost R and Knudsen KE: Molecular
pathogenesis and progression of prostate cancer. Semin Oncol.
40:244–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yin J, Fu W, Dai L, Jiang Z, Liao H, Chen
W, Pan L and Zhao J: ANKRD22 promotes progression of non-small cell
lung cancer through transcriptional up-regulation of E2F1. Sci Rep.
7:44302017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hennink I, van Leeuwen MW, Penning LC and
Piek CJ: Increased number of tissue factor protein expressing
thrombocytes in canine idiopathic immune mediated hemolytic anemia.
Vet Immunol Immunopathol. 196:22–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang J, Chen Y, Xiong X, Zhou X, Han L, Ni
L, Wang W, Wang X, Zhao L, Shao D and Huang C: Peptidome analysis
reveals novel serum biomarkers for children with autism spectrum
disorder in China. Proteomics Clin Appl. 12:e17001642018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roell W, Klein AM, Breitbach M, Becker TS,
Parikh A, Lee J, Zimmermann K, Reining S, Gabris B, Ottersbach A,
et al: Overexpression of Cx43 in cells of the myocardial scar:
Correction of post-infarct arrhythmias through heterotypic
cell-cell coupling. Sci Rep. 8:71452018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fragiadaki M and Zeidler MP: Ankyrin
repeat and single KH domain 1 (ANKHD1) drives renal cancer cell
proliferation via binding to and altering a subset of miRNAs. J
Biol Chem. 293:9570–9579. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bai R, Li D, Shi Z, Fang X, Ge W and Zheng
S: Clinical significance of Ankyrin repeat domain 12 expression in
colorectal cancer. J Exp Clin Cancer Res. 32:352013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wissing ML, Kristensen SG, Andersen CY,
Mikkelsen AL, Høst T, Borup R and Grøndahl ML: Identification of
new ovulation-related genes in humans by comparing the
transcriptome of granulosa cells before and after ovulation
triggering in the same controlled ovarian stimulation cycle. Hum
Reprod. 29:997–1010. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chakrabarty B and Parekh N: Identifying
tandem Ankyrin repeats in protein structures. BMC Bioinformatics.
15:65992014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miller MK, Bang ML, Witt CC, Labeit D,
Trombitas C, Watanabe K, Granzier H, McElhinny AS, Gregorio CC and
Labeit S: The muscle ankyrin repeat proteins: CARP, ankrd2/Arpp and
DARP as a family of titin filament-based stress response molecules.
J Mol Biol. 333:951–964. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Parra RG, Espada R, Verstraete N and
Ferreiro DU: Structural and energetic characterization of the
ankyrinrepeat protein family. PLoS Comput Biol. 11:e10046592015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tanno H, Yamaguchi T, Goto E, Ishido S and
Komada M: The Ankrd 13 family of UIM-bearing proteins regulates EGF
receptor endocytosis from the plasma membrane. Mol Biol Cell.
23:1343–1353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wette SG, Smith HK, Lamb GD and Murphy RM:
Characterization of muscle ankyrin repeat proteins in human
skeletal muscle. Am J Physiol Cell Physiol. 313:C327–C339. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pan W, Sun K, Tang K, Xiao Q, Ma C, Yu C
and Wei Z: Structural insights into ankyrin repeat-mediated
recognition of the kinesin motor protein KIF21A by KANK1, a
scaffold protein in focal adhesion. J Biol Chem. 293:1944–1956.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Venner JM, Famulski KS, Badr D, Hidalgo
LG, Chang J and Halloran PF: Molecular landscape of T cell-mediated
rejection in human kidney transplants: Prominence of CTLA4 and PD
ligands. Am J Transplant. 14:2565–2576. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
VanDussen KL, Carulli AJ, Keeley TM, Patel
SR, Puthoff BJ, Magness ST, Tran IT, Maillard I, Siebel C, Kolterud
Å, et al: Notch signaling modulates proliferation and
differentiation of intestinal crypt base columnar stem cells.
Development. 139:488–497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lubman OY, Korolev SV and Kopan R:
Anchoringnotchgenetics and biochemistry; structural analysis of the
ankyrin domain sheds light on existing data. Mol Cell. 13:619–626.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huan YW, Bengtsson RJ, MacIntyre N,
Guthrie J, Finlayson H, Smith SH, Archibald AL and Ait-Ali T:
Lawsonia intracellularis exploits β-catenin/Wnt and Notch
signalling pathways during infection of intestinal crypt to alter
cell homeostasis and promote cell proliferation. PLoS One.
12:e01737822017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
de Jonge HJ, Woolthuis CM, Vos AZ, Mulder
A, van den Berg E, Kluin PM, van der Weide K, de Bont ES, Huls G,
Vellenga E and Schuringa JJ: Gene expression profiling in the
leukemic stem cell-enriched CD34+ fraction identifies
target genes that predict prognosis in normal karyotype AML.
Leukemia. 25:1825–1833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu L, Tong X, Zhang S, Yin F, Li X, Wei H,
Li C, Guo Y and Zhao J: ASPP2 suppresses stem cell-like
characteristics and chemoresistance by inhibiting the Src/FAK/Snail
axis in hepatocellular carcinoma. Tumour Biol. 37:13669–13677.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lahiani A, Zahavi E, Netzer N, Ofir R,
Pinzur L, Raveh S, Arien-Zakay H, Yavin E and Lazarovici P: Human
placental eXpanded (PLX) mesenchymal-like adherent stromal cells
confer neuroprotection to nerve growth factor (NGF)-differentiated
PC12 cells exposed to ischemia by secretion of IL-6 and VEGF.
Biochim Biophys Acta. 1853:422–430. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang L, Ju X, Cheng Y, Guo X and Wen T:
Identifying Tmem59 related gene regulatory network of mouse neural
stem cell from a compendium of expression profiles. BMC Syst Biol.
5:1522011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hino S, Michiue T, Asashima M and Kikuchi
A: Casein kinase I epsilon enhances the binding of Dvl-1 to Frat-1
and is essential for Wnt-3a-induced accumulation of beta-catenin. J
Biol Chem. 278:14066–14073. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schoenborn JR, Nelson P and Fang M:
Genomic profiling defines subtypes of prostate cancer with the
potential for therapeutic stratification. Clin Cancer Res.
19:4058–4066. 2013. View Article : Google Scholar : PubMed/NCBI
|