Introduction
Globally, lung cancer is the leading cause of
cancer-associated mortality (1,2).
Non-small cell lung cancer (NSCLC), accounting for ~85% of lung
cancer cases, and is primarily divided into two subtypes: Squamous
cell carcinoma and adenocarcinoma, which are derived from
epithelial cells that line the larger and peripheral small airways,
respectively (3). NSCLC is an
aggressive type of cancer that is accompanied by poor overall
survival (4). Therefore,
investigating the mechanisms of NSCLC progression is of great
importance for the development of targeted treatments for this
disease.
The mitogen-activated protein kinase kinase kinase
kinase (MAP4K) family includes MAP4K1 (5), MAP4K2, MAP4K3 (6), MAP4K4 (7) and MAP4K5, and it is a subtype of the
mammalian sterile 20 (Ste20) family (8). Kinases of the MAP4K family are
important regulators of MAPK, which modulates multiple cellular
functions, including cell proliferation, apoptosis and migration
(9,10). MAP4K3 has been previously reported to
regulate cell death (11). In
hepatocellular carcinoma cell lines, MAP4K3 has also been shown to
induce cell migration and invasion (12). Overexpression of MAP4K3 was
identified in NSCLC tumor tissues (13), and correlated with recurrence in
patients with NSCLC (14).
MicroRNAs (miRs/miRNAs) are a class of small,
non-coding RNAs, with a length of 19–24 nucleotides that can reduce
the translation or induce the degradation of target messenger RNAs
(mRNAs) (15). Specific miRNA
expression changes are associated with prognosis in cancer patients
(16). miRNAs serve important roles
in multiple biological processes, including cell proliferation
(17) and apoptosis (18). For the present study, a list of
potential miRNAs, including miR-98-5p, that may target MAP4K3 were
identified using a bioinformatics approach. A subsequent literature
search revealed that miR-98-5p was a member of the let-7 family
(19), which has previously been
reported to be a regulator of MAP4K3 (13). The current study aimed to explore the
potential of this miRNA to regulate MAP4K3 in NSCLC.
Materials and methods
Cell lines and clinical samples
The 293T cells and human NSCLC cell line A549 were
purchased from the American Type Culture Collection and cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc), 100 U/ml penicillin (Invitrogen; Thermo Fisher
Scientific, Inc.) and 100 µg/ml streptomycin (Invitrogen; Thermo
Fisher Scientific, Inc.) in an incubator at 37°C with 95%
humidified atmosphere and 5% CO2.
A total of 90 NSCLC specimens and matched adjacent
normal tissues were obtained from patients with NSCLC who underwent
surgery at the General Hospital of Xuzhou Mining Group, The Second
Affiliated Hospital of Xuzhou Medical University (Xuzhou, China)
between March 2014 and November 2016. The patients had not received
preoperative radiotherapy or chemotherapy. Written informed consent
was obtained from each patient or the patient's family. The
protocols were approved by the Ethics Committee of the General
Hospital of Xuzhou Mining Group.
Cell transfection
miR-98-5p mimic and a negative control (miR-NC) were
constructed by Shanghai GenePharm Co., Ltd. A total of
1×106 A549 cells/well were seeded in 6-well plates and
incubated at 37°C with 5% CO2 for 24 h to a confluence
of 50–60%. Next, A549 cells were transfected with 50 nM miR-98-5p
mimic (5′-UGAGGUAGUAAGUUGUAUUGUU-3′) or miR-NC
(5′-UCGCUUGGUGCAGGUCGGG-3′) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol, and incubated at 37°C for 48 h before the
following experiments were performed. Cells were randomly divided
into 3 groups: i) Control group, untransfected cells; ii) miR-NC
group, cells transfected with miR-NC; and iii) miR-98-5p group,
cells transfected with miR-98-5p.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from NSCLC tumor tissues,
adjacent normal tissues and NSCLC A549 cells according to the
protocol of the miRNeasy Mini kit (Qiagen, Inc.). For detection of
mature miRNA, poly(A) tails were added by incubation of RNA (100
ng) with poly(A) polymerase (New England BioLabs, Inc.), followed
by RT with oligo-dT adapter primers and Moloney murine leukemia
virus (MMLV) polymerase (Invitrogen; Thermo Fisher Scientific,
Inc.). For quantification of mRNA, complementary DNAs were
synthesized from total RNA using oligo (dT)15 primers and MMLV
polymerase (Invitrogen; Thermo Fisher Scientific, Inc.). mRNA and
miRNA were reverse-transcribed into cDNA under the following
conditions: Incubation at 37°C for 50 min, followed inactivation at
70°C for 15 min. qPCR was performed using SYBR Green PCR Master mix
(Toyobo Life Science) on a Light-Cycler 480 Real-Time PCR System
(Roche Diagnostics), and the thermocycling conditions were as
follows: initial denaturation at 94°C for 2 min, followed by 40
cycles of 94°C for 5 sec and 60°C for 30 sec. The relative levels
of miRNA and mRNA were normalized to U6 and GAPDH, respectively.
The primer sequences were as follows: MAP4K3 forward,
5′-GACTCCCCTGCAAAAAGTCTG-3′ and reverse,
5′-GTCCATAGGTGCCATTTCCAA-3′; GAPDH forward,
5′TTGGTATCGTGGAAGGACTCA-3′ and reverse,
5′-TGTCATCATATTTGGCAGGTT-3′; miR-98-5p forward,
5′-TGAGGTAGTAGTTTGTGCTGTT-3′ and reverse,
5′-GCGAGCACAGAATTAATACGAC-3′; U6 forward,
5′-TGCGGGTGCTCGCTTCGCAGC-3′ and reverse, 5′-CCAGTGCAGGGTCCGAGGT-3′.
The fold-change in miRNA and mRNA expression was calculated using
the 2−ΔΔCq method (20).
Western blotting
Proteins were isolated from NSCLC tumor tissues,
adjacent normal tissues and the NSCLC cell line A549 using
radioimmunoprecipitation assay buffer (Roche, Diagnostics)
containing a cocktail of complete protease inhibitor (Roche
Diagnostics GmbH). Protein samples (20 µg) were separated by
SDS-PAGE on an 8–10% gel and blotted onto polyvinylidene difluoride
membranes (EMD Millipore). Subsequently, rabbit anti-MAP4K3
antibody (1:1,000 dilution; cat. no. 92427; Cell Signaling
Technology, Inc.), rabbit anti-GAPDH antibody (1:5,000 dilution;
cat. no. 5174; Cell Signaling Technology, Inc.), rabbit
anti-phosphorylated-mammalian target of rapamycin (p-mTOR) antibody
(1:1,000 dilution; cat. no. 5536; Cell Signaling Technology, Inc.),
rabbit anti-mTOR antibody (1:1,000 dilution; cat. no. 2792; Cell
Signaling Technology, Inc.) and rabbit anti-cleaved caspase-3
antibody (1:1,000 dilution; cat. no. 9654; Cell Signaling
Technology, Inc.) were used as primary antibodies, and were
incubated at 4°C overnight. Membranes were incubated with
horseradish peroxidase-conjugated goat anti-rabbit (1:10,000
dilution; cat. no. CW0234S; Beijing ComWin Biotech Co., Ltd.) and
anti-mouse (1: 10,000 dilution; cat. no. CW0221S; Beijing ComWin
Biotech Co., Ltd.) immunoglobulin G secondary antibodies at 37°C
for 1 h. The protein bands were determined by chemiluminescence
(EMD Millipore). Densitometric analysis was performed using ImageJ
version 1.8.0 (National Institutes of Health).
Cell viability
A549 cells were seeded in 96-well plates and
incubated for 48 h following transfection. Subsequently, Cell
Counting kit-8 (CCK-8, Dojindo Molecular Technologies, Inc.) was
used for the examination of cell viability at 0, 12, 24 and 48 h of
incubation. The number of cells was assessed with FLUOstar OPTIMA
(BMG Labtech GmbH) at an absorbance wavelength of 450 nm.
Cell apoptosis
At 72 h after cell transfection, flow cytometry with
Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI)
was used for determination of cell apoptosis in the different
groups. A549 cells were washed, trypsinized and resuspended in the
staining solution provided in the Annexin V-FITC Apoptosis
Detection kit (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. After 1 h of incubation
of the cells with Annexin V-FITC antibody at 37°C, apoptosis was
determined using a flow cytometer (BD Biosciences). The apoptotic
cells presented with a positive Annexin V-FITC signal and a
negative PI signal. The cell number at each phase was determined by
FlowJo software version 7.6.3 (FlowJo LLC).
Dual-luciferase reporter assay
MAP4K3 was predicted to be a gene target for
miR-98-5p by bioinformatics analysis using TargetScan release 7.1
(http://www.targetscan.org/vert_71).
PCR was performed using the following thermocycling conditions:
94°C for 2 min, followed by 35 cycles of 94°C for 2 sec, 60°C for
60 sec and 72°C for 1 min with the following primers: MAP4K3 3′UTR
forward, 5′-GGTACCAAAATAATTTAGTTACT-3′ and reverse,
5′-CTCGAGTGAGGTAGTAACT-3′ and Platinum™ II Green PCR buffer (Thermo
Fisher Scientific, Inc.). The PCR products were amplified using
cDNA from 293T cells and fused to the firefly luciferase gene of
the pGL3-control plasmid (Promega Corporation) with the restriction
enzyme sites of KpnI and XhoI. Two site mutations
were introduced to WT-MAP4K3-3′-UTR to construct the mutant (Mut)
MAP4K3-3′-UTR by a Quick Site-directed mutation kit (Agilent
Technologies, Inc.). The 293T cells were co-transfected with pGL3
constructions including 200 ng pGL3-WT-MAP4K3 and 200 ng
pGL3-Mut-MAP4K3, 10 nM miR-NC or 10 nM miR-98-5p mimic and 26 ng
pRL-TK in 24-well plates using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). At 24 h of
transfection, luciferase activity (firefly and Renilla) was
determined using the dual-luciferase reporter assay system (Promega
Corporation).
Statistical analysis
Statistical analyses were performed using SPSS 13
software (SPSS, Inc.). Differences between two groups and among
multiple groups were analyzed by Student's t-test and one-way
analysis of variance followed by Newman-Keuls test, respectively.
Results were presented as the mean ± standard deviation. P<0.05
was considered to indicate a statistically significant
difference.
Results
MAP4K3 expression level is increased
in NSCLC tumor tissues
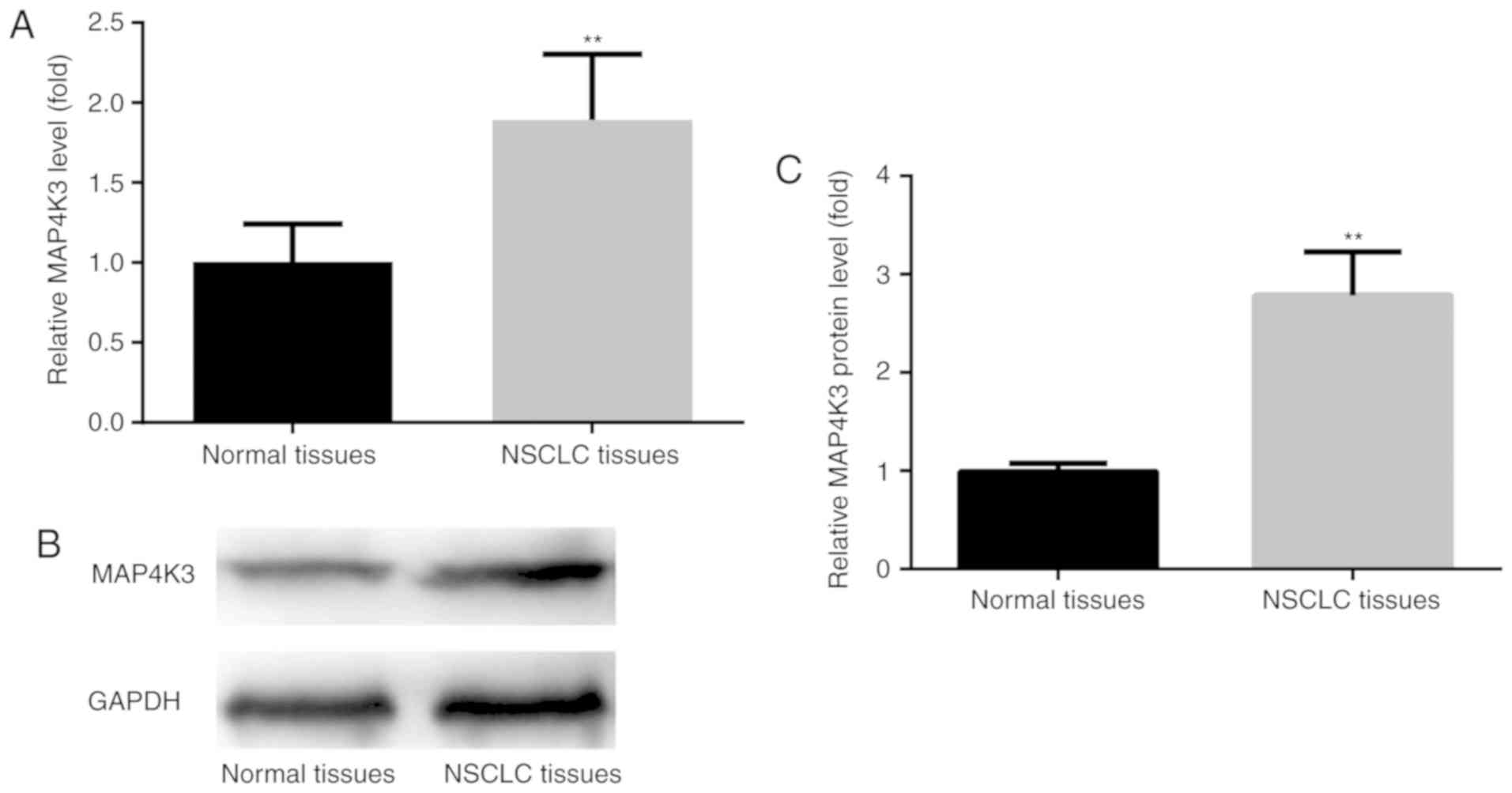
To examine the expression level of MAP4K3, tumor
tissues and their adjacent normal tissues from 90 NSCLC cases were
analyzed using RT-qPCR and western blotting. The results revealed a
significant increase in MAP4K3 mRNA levels in NSCLC tumor tissues
compared with the adjacent normal tissues (Fig. 1A). The western blot results revealed
that, compared with the adjacent normal tissues, MAP4K3 protein
level was increased in NSCLC tumor tissues (Fig. 1B and C). These results were
consistent with a previous report which showed that higher
expression of MAP4K3 was associated with increased recurrence risk
for lung cancer patients (14).
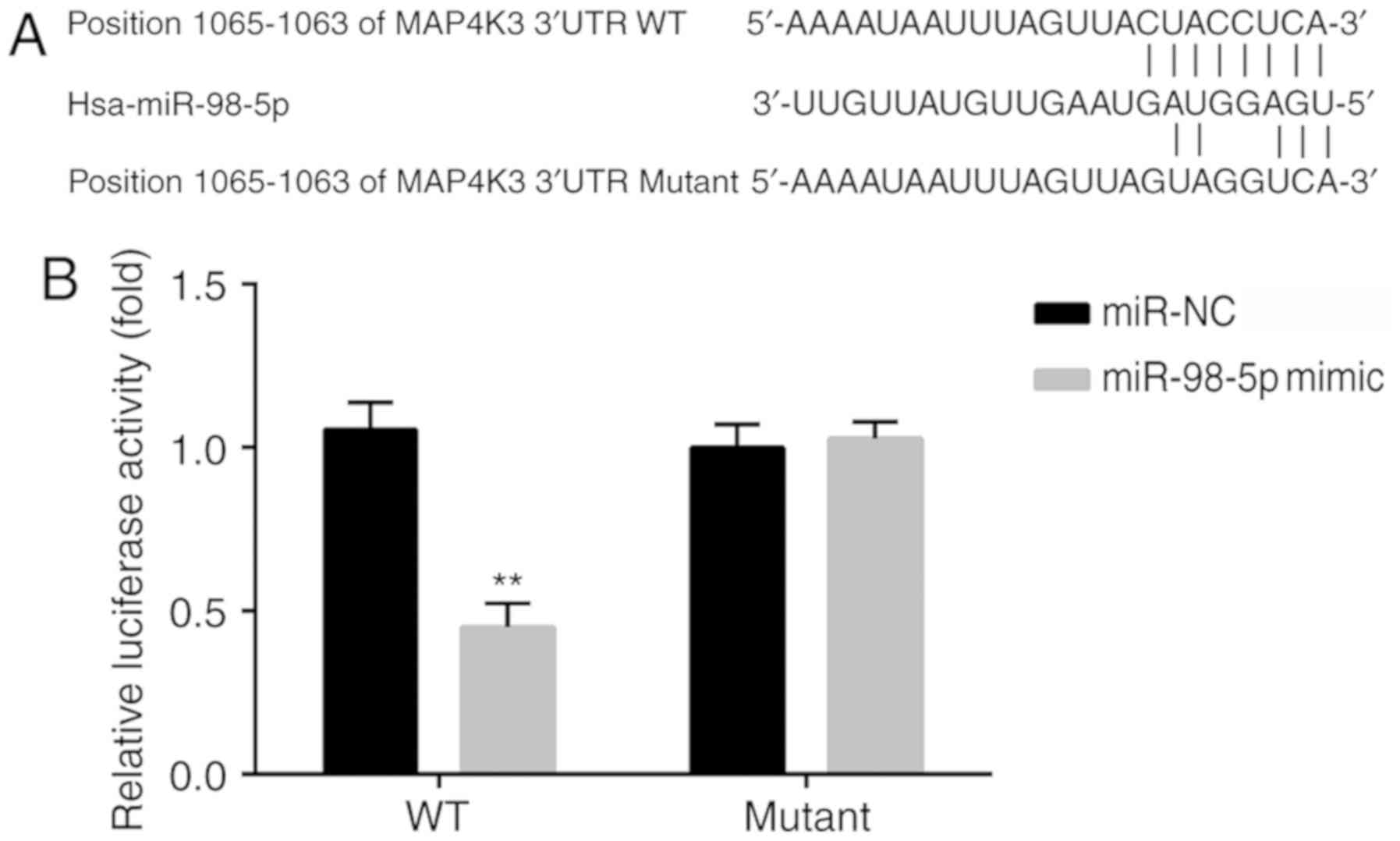
MAP4K3 is a target for miR-98-5p
MAP4K3 was predicted to be a gene target for
miR-98-5p by bioinformatics analysis using TargetScan, and the
binding sequences between positions 1,056-1,063 of the MAP4K3
3′-UTR and miR-98-5p are represented in Fig. 2A. In addition, other miRNAs were
predicted to target MAP4K3 based on the analysis by TargetScan. For
instance, through literature review, let-7c was identified to be a
regulator of MAP4K3 (19). As
miR-98-5p is a member of the let-7 family, this miRNA was selected
for investigation in the current study (13).
The 3′-UTR of MAP4K3 mRNA was verified to be a
target for miR-98-5p by dual-luciferase reporter analysis. The
relative luciferase activity in the cells transfected with wild
type (WT) MAP4K3 3′-UTR and miR-98-5p mimic was significantly
decreased compared with that observed in cells transfected with WT
MAP4K3 3′-UTR and miR-NC. There was no significant difference in
relative luciferase activity between cells transfected with mutant
MAP4K3 3′-UTR and miR-98-5p mimic and cells transfected with mutant
MAP4K3 3′-UTR and miR-NC (Fig.
2B).
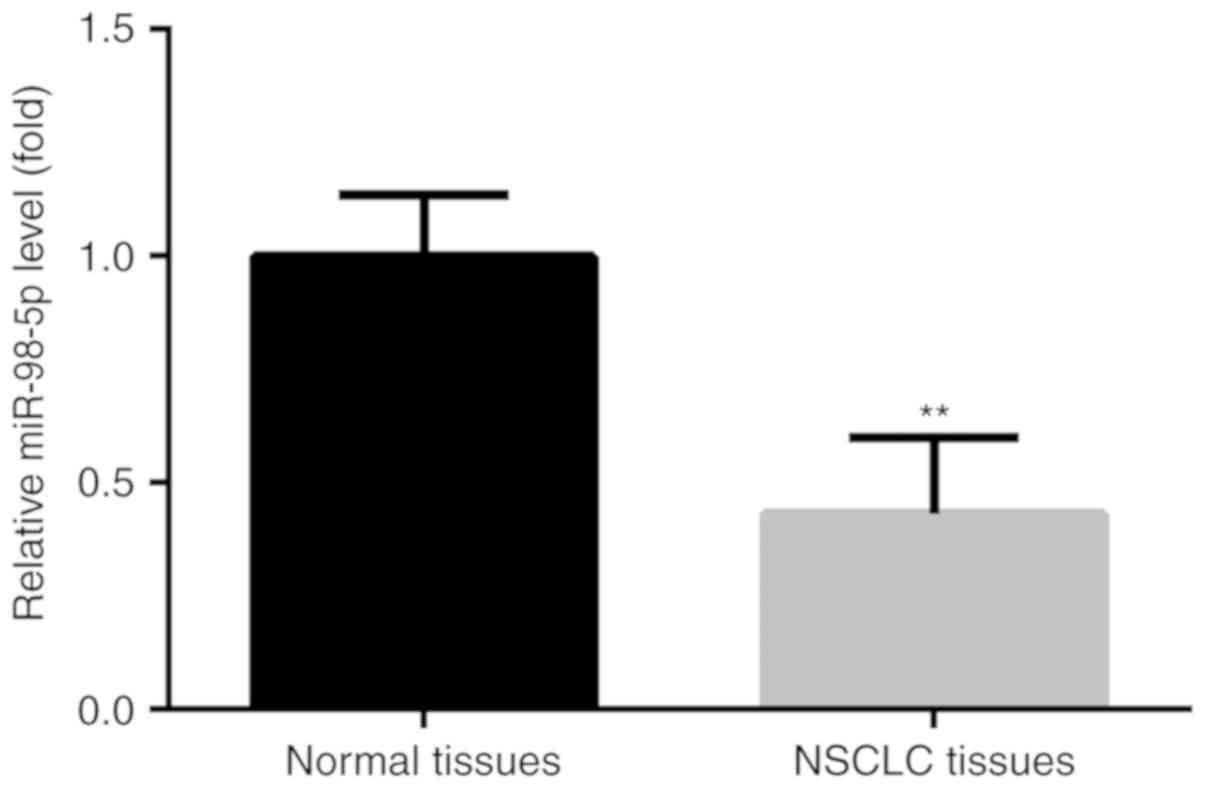
miR-98-5p expression is decreased in
NSCLC tumor tissues
The expression levels of miR-98-5p was determined in
tumor tissues and adjacent normal tissues from 90 patients with
NSCLC. The results revealed a significant decrease in miR-98-5p in
NSCLC tumor tissues compared with the adjacent normal tissues
(Fig. 3).
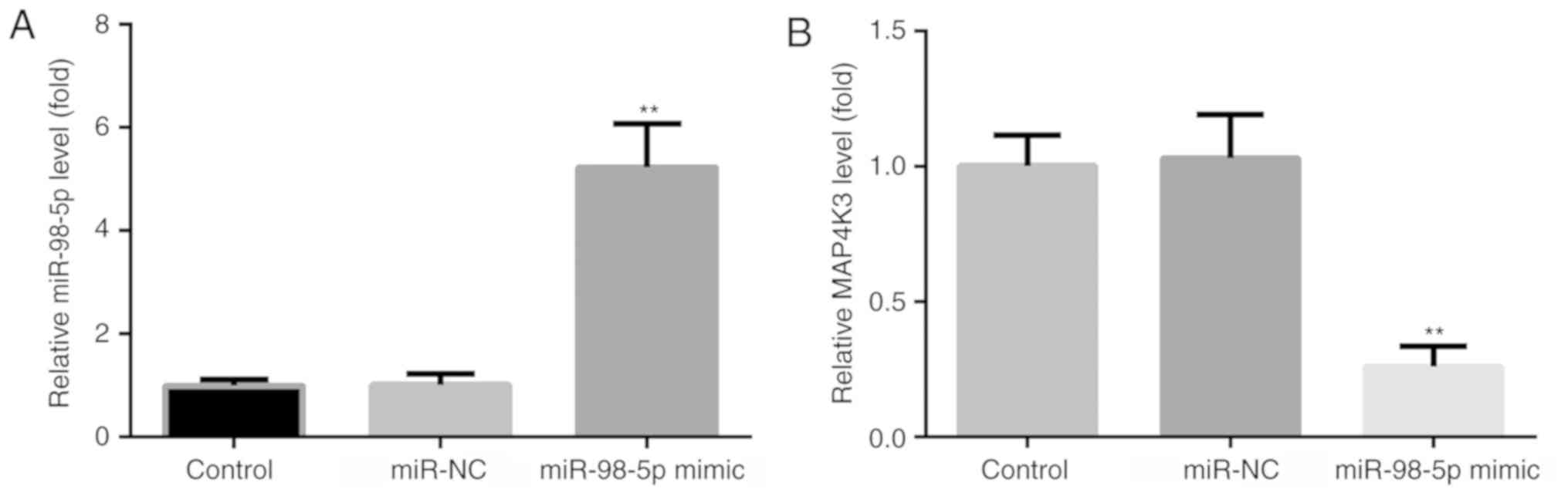
miR-98-5p overexpression increases
miR-98-5p and decreases MAP4K3 levels
To examine the effects of miR-98-5p mimic on the
expression changes of miR-98-5p and MAP4K3 in A549 cells, RT-qPCR
was used. Compared with the control and miR-NC groups, an increase
in miR-98-5p (Fig. 4A) and a
decrease in MAP4K3 (Fig. 4B) levels
were observed in the miR-98-5p mimic group.
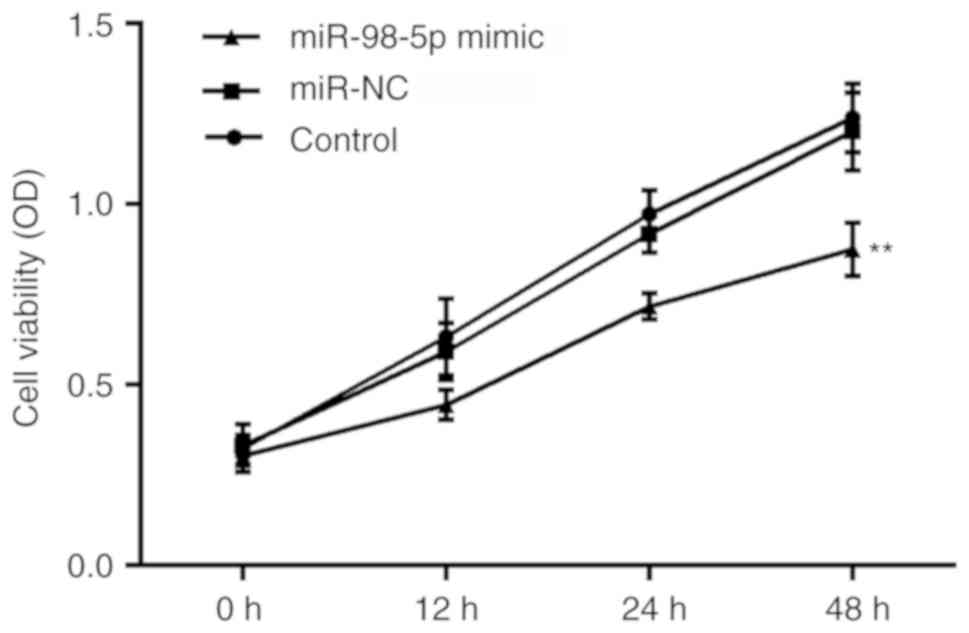
miR-98-5p overexpression reduces cell
viability
The effects of miR-98-5p on cell viability in NSCLC
A549 cells were determined by CCK-8 assay. The growth curves
demonstrated the effects of miR-98-5p on NSCLC cell viability. At
48 h of incubation following cell transfection, a significant
decrease in cell viability was observed in the miR-98-5p mimic
group compared with the control and miR-NC groups, whereas no
significant differences were present at 0, 12 or 24 h of incubation
(Fig. 5).
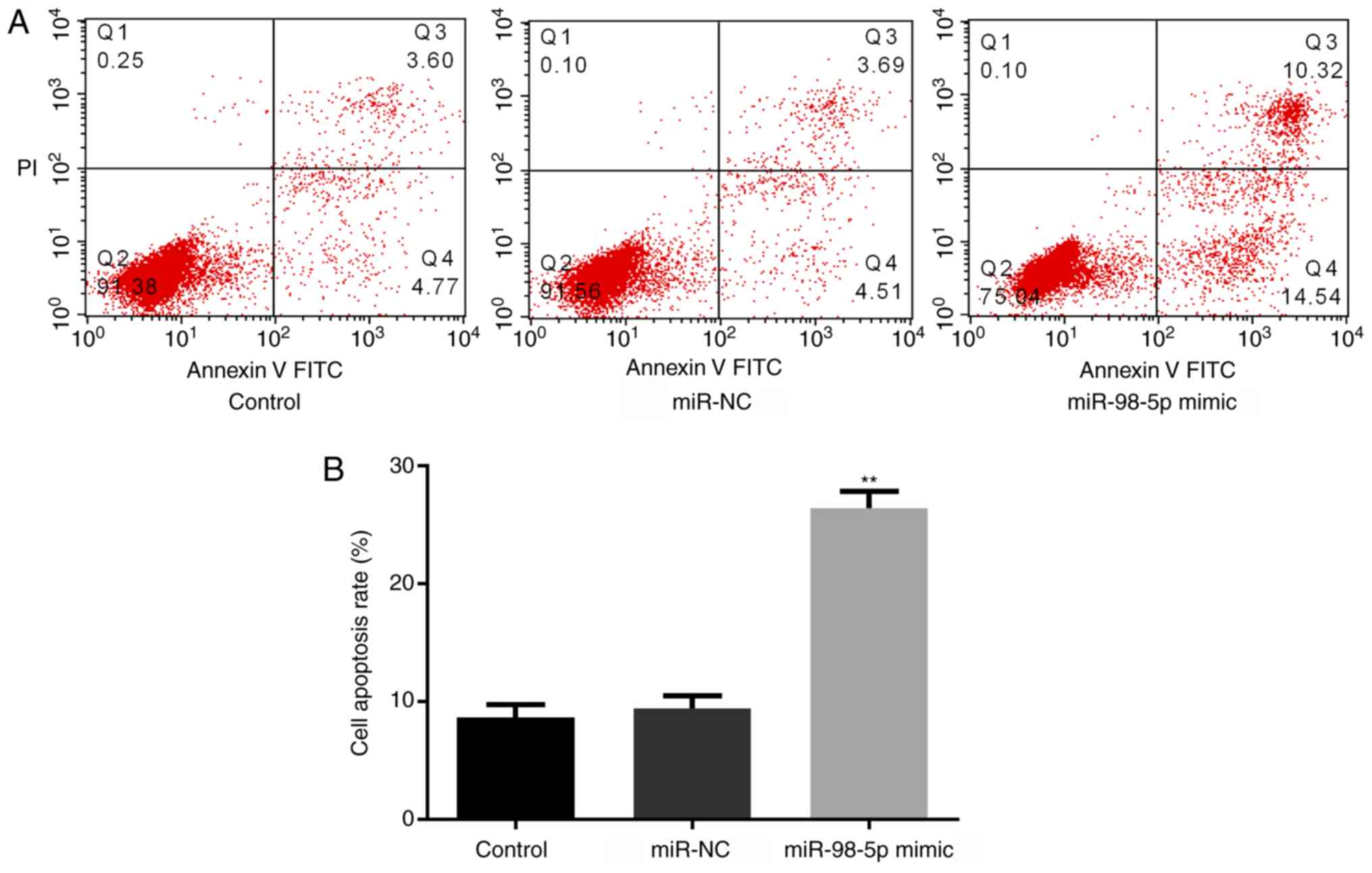
miR-98-5p overexpression induces cell
apoptosis
The effects of miR-98-5p on cell apoptosis in the
NSCLC cell line A549 were determined by flow cytometry. The results
demonstrated that, compared with the control and miR-NC groups,
there was a significant increase in cell apoptosis in the miR-98-5p
mimic group (Fig. 6A and B).
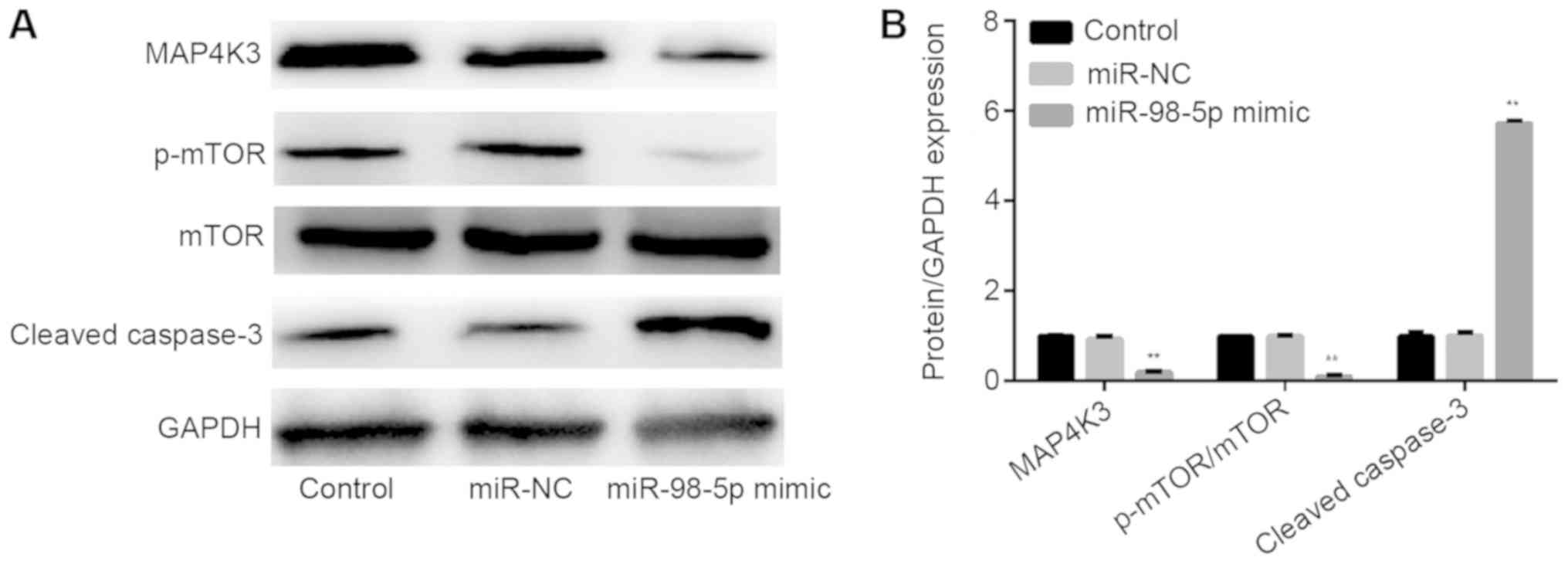
miR-98-5p regulates MAP4K3 expression
and the mTOR signaling pathway
To further examine the association between miR-98-5p
and MAP4K3, as well as the potential regulatory mechanisms of
miR-98-5p in the mTOR signaling pathway, the protein levels of
associated molecules were determined by western blotting. As
presented in Fig. 7A and B, the
expression levels of MAP4K3 and p-mTOR were decreased, cleaved
caspase-3 expression was increased, and the expression of mTOR was
not significantly changed upon transfection with miR-98-5p mimic
compared to control and miR-NC groups.
Discussion
Patients with NSCLC exhibit a median 5-year survival
rate ranging from 26% (stage IIIA) to 51% (stage IA), which is
lower than that of other types of cancer, including breast and
prostate cancer (21). Despite the
uncertain efficacy of chemotherapy, the most frequently used
therapeutic method for the treatment of early-stage NSCLC is
complete surgical resection alongside cisplatin-based chemotherapy
(22). The results of the present
study indicate that miR-98-5p, which was significantly decreased in
NSCLC tumor tissues, could target MAP4K3 and inhibit the activation
of the mTOR signaling pathway. These findings support an apical
role for miR-98-5p/MAP4K3 in intracellular signal transduction
cascades in NSCLC.
MAP4K3 is a protein kinase of the Ste20 family that
is activated by ultraviolet radiation and the proinflammatory
cytokine tumor necrosis factor-α (6). The present study demonstrated that
MAP4K3, a target gene of miR-98-5p, was overexpressed in NSCLC
tissues, while miR-98-5p was found to be downregulate. These
findings indicate that miR-98-5p and MAP4K3 may serve roles in the
progression of NSCLC.
The ability of tumor cells to increase cell number
is determined by the cell proliferation rate and the extent of cell
removal through apoptosis (23).
Acquired resistance to apoptosis may be a hallmark of all cancer
types (23). The present study
investigated the effects of miR-98-5p on proliferation and
apoptosis in A549 cells. It was observed that overexpression of
miR-98-5p significantly inhibited cell proliferation and promoted
apoptosis. These findings suggest a potential function of miR-98-5p
as a therapeutic target for NSCLC. Previous studies have revealed
that miRNA levels in tumors are variable, even among commercial
cell lines, suggesting that further research is required to
determine the potential association between miRNA changes and
clinical diagnostics or therapeutic treatments (24,25).
mTOR signaling is deregulated in multiple diseases
including cancer (26). The present
study revealed that miR-98-5p could significantly inhibit the
activation of mTOR. This phenomenon indicated that miR-98-5p was
able to regulate MAP4K3 expression and affect NSCLC cell activities
via the mTOR signaling pathway.
However, the present study had limitations, as it
was not directly demonstrated that altered MAP4K3 expression
impacted proliferation and apoptosis of lung cancer cells. Future
studies will investigate MAP4K3 overexpression in NSCLC cells and
the interaction of MAP4K3 and miR-98-5p.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZW, ZH, LZ, SZ performed the experiments and
analyzed the data. BW conceived the study, analyzed the data and
prepared the manuscript. All the authors read and approved the
final version of manuscript for publication.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of General Hospital of Xuzhou Mining Group, The Second
Affiliated Hospital of Xuzhou Medical University (Xuzhou, China).
All patients or their families provided written informed consent
prior to the start of the research.
Patient consent for publication
Not applicable.
Competing of interests
The authors declare that they have no competing
interests.
References
|
1
|
Dent AG, Sutedja TG and Zimmerman PV:
Exhaled breath analysis for lung cancer. J Thorac Dis. 5:S540–S550.
2013.PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
World Health Organization Classification
of Tumors, . Pathology and Genetics of Tumours of the Lung, Pleura,
Thymus and HeartWilliam D, Travis WD, Elisabeth Brambilla E, Konrad
Müller-Hermelink H and Harris CC: IARC Press; Lyon: 2004
|
|
4
|
Wang T, Nelson RA, Bogardus A and Grannis
FW Jr: Five-year lung cancer survival: Which advanced stage
nonsmall cell lung cancer patients attain long-term survival?
Cancer. 116:1518–1525. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu MC, Qiu WR, Wang X, Meyer CF and Tan
TH: Human HPK1, a novel human hematopoietic progenitor kinase that
activates the JNK/SAPK kinase cascade. Genes Dev. 10:2251–2264.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diener K, Wang XS, Chen C, Meyer CF,
Keesler G, Zukowski M, Tan TH and Yao Z: Activation of the c-Jun
N-terminal kinase pathway by a novel protein kinase related to
human germinal center kinase. Proc Natl Acad Sci USA. 94:9687–9692.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chuang HC, Sheu WH, Lin YT, Tsai CY, Yang
CY, Cheng YJ, Huang PY, Li JP, Chiu LL, Wang X, et al: HGK/MAP4K4
deficiency induces TRAF2 stabilization and Th17 differentiation
leading to insulin resistance. Nat Commun. 5:46022014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen YR and Tan TH: Mammalian c-Jun
N-terminal kinase pathway and STE20-related kinases. Gene Ther Mol
Biol. 4:83–98. 1999.
|
|
9
|
Chen YR and Tan TH: The c-Jun N-terminal
kinase pathway and apoptotic signaling (review). Int J Oncol.
16:651–662. 2000.PubMed/NCBI
|
|
10
|
MacCorkle RA and Tan TH: Mitogen-activated
protein kinases in cell-cycle control. Cell Biochem Biophys.
43:451–461. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lam D, Dickens D, Reid EB, Loh SH, Moisoi
N and Martins LM: MAP4K3 modulates cell death via the
post-transcriptional regulation of BH3-only proteins. Proc Natl
Acad Sci USA. 106:11978–11983. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu L, Lu L, Zheng A, Xie J, Xue Q, Wang
F, Wang X, Zhou H, Tong X, Li Y, et al: miR-199a-5p and let-7c
cooperatively inhibit migration and invasion by targeting MAP4K3 in
hepatocellular carcinoma. Oncotarget. 8:13666–13677.
2017.PubMed/NCBI
|
|
13
|
Zhao B, Han H, Chen J, Zhang Z, Li S, Fang
F, Zheng Q, Ma Y, Zhang J, Wu N and Yang Y: MicroRNA let-7c
inhibits migration and invasion of human non-small cell lung cancer
by targeting ITGB3 and MAP4K3. Cancer Lett. 342:43–51. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsu CP, Chuang HC, Lee MC, Tsou HH, Lee
LW, Li JP and Tan TH: GLK/MAP4K3 overexpression associates with
recurrence risk for non-small cell lung cancer. Oncotarget.
7:41748–41757. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Markou A, Tsaroucha EG, Kaklamanis L,
Fotinou M, Georgoulias V and Lianidou ES: Prognostic value of
mature microRNA-21 and microRNA-205 overexpression in non-small
cell lung cancer by quantitative real-time RT-PCR. Clin Chem.
54:1696–1704. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bueno MJ, Pérez de Castro I and Malumbres
M: Control of cell proliferation pathways by microRNAs. Cell Cycle.
7:3143–3148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jovanovic M and Hengartner MO: miRNAs and
apoptosis: RNAs to die for. Oncogene. 25:6176–6187. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Bao W, Liu Y, Wang S, Xu S, Li X,
Li Y and Wu S: miR-98-5p contributes to cisplatin resistance in
epithelial ovarian cancer by suppressing miR-152 biogenesis via
targeting Dicer1. Cell Death Dis. 9:4472018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using realtime quantitative PCR and
the 2(Delta Delta C(T)) method. Methods. 25:4024082001. View Article : Google Scholar
|
|
21
|
Chuang JC, Neal JW, Niu XM and Wakelee HA:
Adjuvant therapy for EGFR mutant and ALK positive NSCLC: Current
data and future prospects. Lung Cancer. 90:172015. View Article : Google Scholar
|
|
22
|
NSCLC Metaanalyses Collaborative Group, ;
Arriagada R, Auperin A, Burdett S, Higgins JP, Johnson DH, Le
Chevalier T, Le Pechoux C, Parmar MK, Pignon JP, et al: Adjuvant
chemotherapy, with or without postoperative radiotherapy, in
operable nonsmallcell lung cancer: Two metaanalyses of individual
patient data. Lancet. 375:126712772010.
|
|
23
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dumache R, Rogobete AF, Andreescu N and
Puiu M: Genetic and epigenetic biomarkers of molecular alterations
in oral carcinogenesis. Clin Lab. 61:1373–1381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zahran F, Ghalwash D, Shaker O, Al-Johani
K and Scully C: Salivary microRNAs in oral cancer. Oral Dis.
21:739–747. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Soulard A and Hall MN: SnapShot: mTOR
signaling. Cell. 129:4342007. View Article : Google Scholar : PubMed/NCBI
|